Abstract
Objectives
The release of nitric oxide is controlled by cholinergic and adrenergic receptors. Recent observations suggest that activation of β-adrenoceptors can inhibit the release of nitric oxide. The aim of the present study was to examine the effect of α1- and α2-adrenoceptor activation on nitric oxide-mediated vasodilation.
Methodology
In a first set of experiments, the endothelium-dependent vasodilators acetylcholine (ACh), 5-hydroxytryptamine (5HT), and bradykinin (BK), and the nitric oxide donor sodium nitroprusside (SNP) were administered in a random order in the brachial artery together with saline, or the nonselective α-adrenoceptor agonists norepinephrine or clonidine, or the nitric oxide synthase inhibitor NG-monomethyl-L-arginine (L-NMMA). The infusions of saline, norepinephrine, clonidine, and L-NMMA started 10 minutes before the infusions of ACh, BK, 5HT, and SNP. In a second set of experiments, cumulative doses of ACh, BK, and 5HT were infused, in a random order, intra-arterially together with saline or the selective α1-adrenoceptor agonist methoxamine. The infusions of saline and methoxamine started 5 minutes before the infusions of ACh, BK, and 5HT. Forearm blood flow was measured using computerized venous occlusion plethysmography.
Results
ACh, 5HT, BK, and SNP induced a significant increase in forearm blood flow (p < 0.05 for all). These vasodilator responses were significantly attenuated by norepinephrine, clonidine, and L-NMMA (p < 0.05 for all), except for SNP. In the second set of experiments, all three endothelium-dependent vasodilators induced a dose-dependent vasodilation, which was significantly inhibited by methoxamine (p < 0.05).
Conclusion
These results show that endothelium-dependent nitric oxide-mediated vasodilation is inhibited by activation of peripheral α1-adrenoceptors.
Introduction
There is evidence to suggest that the release of vasodilator or vasoconstrictor mediators from endothelial cells is, in part, influenced by the autonomic nervous system (Sartori and Scherrer 1999; CitationZanzinger 1999). The classic example is acetylcholine (ACh), the parasympathetic neurotransmitter, eliciting a nitric oxide-mediated vasodilation via activation of muscarinic type-3 (M3) receptors located on the endothelial cells (CitationFurchgott and Zawadzki 1980; CitationIgnarro 1989; CitationAngus and Lew 1992; CitationBruning et al 1995). Besides this cholinergic receptor, it has been shown that activation of adrenergic receptors can modulate the release of nitric oxide (CitationLiao and Homey 1993). Vasodilatation induced by β-adrenoceptors and α2-adrenoceptors was partially inhibited following inhibition of nitric oxide synthase (CitationDawes et al 1997; CitationLembo et al 1997; CitationMajmudar et al 1999). Recently, CitationHijmering et al (2002) have demonstrated that activation of the sympathetic nervous system inhibits flow-mediated vasodilation, suggesting inhibition of nitric oxide by an α-adrenergic mechanism.
It was the aim of the present study to investigate the effect of α1- and α2-adrenoceptor activation on endothelium-dependent nitric oxide-mediated vasodilation in humans.
Materials and methods
Subjects
Twenty-three healthy non-smoking male volunteers, mean age range 26.0 ± 2.2 years, participated in this study. All subjects gave informed consent. The results of their medical history, physical examination, screening laboratory tests, and ECG were within normal ranges. All volunteers were free from all risk factors that could be associated with impaired endothelium function. The Leiden University Medical Center ethics review committee approved the protocol of the study, and the investigations complied with the principles outlined in the Declaration of Helsinki.
Experimental procedures
The experiments were performed in a quiet room with a constant temperature of 21–23°C. For the duration of the experiment, subjects were in a supine position with both arms stabilized slightly above the level of the heart. On the day of the study and 12 hours before, all subjects refrained from alcohol and caffeine-containing beverages. After local anesthesia of the skin, a 20-gauge polyethylene catheter (Ohmeda, Swindon, UK) was inserted into the brachial artery of the non-dominant arm for measuring intra-arterial blood pressure and infusion of the study drugs using Harvard volumetric precision pumps (Harvard '22, Harvard Apparatus Ltd, Edenbridge, Kent, UK).
The subjects rested for at least 45 minutes after the insertion of the intra-arterial catheter to achieve a stable baseline. Forearm blood flow was measured bilaterally, using computerized R-wave–triggered venous occlusion plethysmography, as described previously (CitationChang et al 1988). During the intra-arterial infusions, the hand was excluded from the circulation using a small wrist cuff inflated to 40 mmHg above the systolic blood pressure. Between the various experiments, the wrist cuffs were deflated and sufficient time (30–45 min) was allotted to let forearm blood flow return to baseline levels.
Blood flow in the control arm was measured to exclude systemic vascular effects of the drugs used. The forearm blood flow results were not corrected by changes in forearm blood flow in the control arm by calculating the forearm ratio, for this would reduce the accuracy of the measurements with the computerized methods used (CitationKamper et al 2001). Heart rate was recorded from a triple-lead electrocardiogram.
Drugs and solutions
The following drugs were used for intra-arterial infusions: acetylcholine•HCl (ACh) (Bournonville Pharma BV, Den Haag, The Netherlands), bradykinin (BK) (Clinalpha, Darmstadt, Germany), 5-Hydroxytryptamine•HCl (5HT) (prepared by the Clinical Pharmacy of the Leiden University Medical Center), sodium-nitroprusside (SNP) (prepared by the Clinical Pharmacy of the Leiden University Medical Center), clonidine (Boehringer Ingelheim BV, Alkmaar, The Netherlands), methoxamine (Glaxo Smith Kline BV, Zeist, The Netherlands), norepinephrine (prepared by the Clinical Pharmacy of the Leiden University Medical Center), and the competitive nitric oxide synthase inhibitor NG-monomethyl-L-arginine (L-NMMA) (Clinalpha, Darmstadt, Germany). All drugs were dissolved in 0.9% NaCl except for SNP, which was dissolved in 5% glucose. All solutions were prepared from sterile stock solutions and ampoules on the day of the experiments.
ACh, BK, and 5HT were used as endothelium-dependent nitric oxide-mediated vasodilators, and SNP as an exogenous nitric oxide donor to induce vasodilatation independent of the endogenous nitric oxide pathway. Both norepinephrine and clonidine were used as non-selective α1- and α2-receptor agonists, and methoxamine as a selective α1-receptor agonist (CitationBradshaw et al 1981; CitationJie et al 1984). L-NMMA was used as an inhibitor of endothelium-dependent nitric oxide-mediated vasodilatation (CitationAngus and Lew 1992; CitationBruning et al 1993; CitationCockcroft et al 1994).
Study protocol
To limit the length of the study day for the participant and to preclude carry-over effects of the various study drugs used, the study was done in two sets of experiments.
Study protocol 1
Seven subjects participated in study protocol 1. First, intra-arterial infusions of ACh (100 ng/kg/min), BK (0.1 ng/kg/min), 5HT (1.0 ng/kg/min), and SNP (100 ng/kg/min) were administered in a random order together with a continuous infusion of saline (0.4 mL/min). Then, intra-arterial infusions of ACh, BK, 5HT, and SNP were repeated under continuous intra-arterial infusion of norepinephrine (1.6 ng/kg/min) and clonidine (60 ng/kg/min) in a random order. Finally, the intra-arterial infusions of ACh, BK, 5HT, and SNP were administered during simultaneous infusion of L-NMMA (30 μg/kg/min).
Each dose of the ACh, BK, and SNP infusions was administered for five minutes. The dose of 5HT was infused for seven minutes. The infusions of saline, norepinephrine, clonidine, and L-NMMA started 10 minutes before the infusions of ACh, BK, 5HT, and SNP. Forearm blood flow, intra-arterial blood pressure and heart rate were measured during the two minutes immediately prior to the start of each infusion and during the last two minutes of each infusion.
Study protocol 2
Sixteen subjects participated in the study for protocol 2. Six subjects received intra-arterial cumulative dose infusions of ACh (100 and 1000 ng/kg/min); another six subjects received intra-arterial infusions of BK (0.01 and 0.1 ng/kg/min); and four subjects received intra-arterial infusion of 5HT (0.3 and 1.0 ng/kg/min). All these infusions were administered under continuous infusion of saline (of 0.4 mL/min). Subsequently, all intra-arterial infusions of ACh, BK, and 5HT were repeated during a continuous intra-arterial infusion of methoxamine (1 μg/kg/min).
Each dose of the ACh and BK infusions was administered for 5 minutes. The doses of 5HT were infused for 7 minutes. The infusions of saline, and methoxamine started 5 minutes before the infusions of ACh, BK, and 5HT. Forearm blood flow, intra-arterial blood pressure, and heart rate were measured during the two minutes immediately prior to the start of each infusion and during the last two minutes of each dose infusion.
Analysis
Forearm blood flow is expressed as mL/min per 100 mL forearm tissue and as percent changes from baseline. Changes in forearm blood flow were analyzed by two-way analysis of variance. The other results were evaluated by Student's t-test for paired and unpaired observations. All p-values less than 0.05 were considered statistically significant. All results are given as means ± standard error.
Results
During the experiments, blood flow in the control arm, heart rate, and intra-arterial blood pressure did not show any relevant changes (data not shown). Therefore, changes in forearm blood flow induced by the study drugs can be interpreted as local vascular effects of these drugs in the forearm (CitationChang et al 1988; CitationBruning el al 1993).
Study protocol 1
The intra-arterial infusion of norepinephrine alone reduced forearm blood flow significantly from 3.71 ± 0.28 to 2.31 ± 0.12 mL/min per 100 mL forearm tissue (p < 0.05). Clonidine reduced the baseline forearm blood flow from 2.51 ± 0.19 to 2.03 ± 0.14 mL/min per 100 mL forearm tissue (p < 0.05). The infusion of L-NMMA alone reduced forearm blood flows from 2.73 ± 0.27 to 2.35 ± 0.27 mL/min per 100 mL forearm tissue (p < 0.05).
The effects of the intra-arterial infusions of the endothelium-dependent vasodilators ACh, BK, 5HT and the nitric oxide donor SNP on forearm blood flow are shown in and . ACh 100 ng/kg/min induced a significant increase in forearm blood flow from 3.59 ± 0.47 to 14.56 ± 4.03 mL/100 mL/min (p < 0.05). BK 0.1 ng/kg/min and 5HT 1.0 ng/kg/min increased forearm blood flow from 3.40 ± 0.55 to 5.67 ± 0.64 mL/100 mL/min and from 3.60 ± 0.72 to 7.02 ± 1.11 mL/100mL/min, respectively (p < 0.05 for both). The infusion of SNP 100 ng/kg/min elevated forearm blood from 4.11 ± 0.68 to 18.89 ± 2.71 mL/100 mL/min (p < 0.05).
Figure 1 Forearm blood flows (mL/min per 100 mL forearm tissue) before and during intra-arterial infusions of acetylcholine (100 ng/kg/min), bradykinin (0.1 ng/kg/min), 5-hydroxytryptamine (1.0 ng/kg/min), and sodium-nitroprusside (100 ng/kg/min). *p < 0.05 compared with saline.
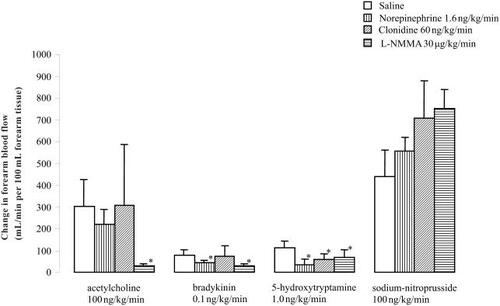
Table 1 Forearm blood flows (mL/min per 100 mL forearm tissue) before and during intra-arterial infusions of acetylcholine (100 ng/kg/min), bradykinin (0.1 ng/kg/min), 5-hydroxytryptamine (1.0 ng/kg/min), and sodium-nitroprusside (100 ng/kg/min)
Concomitant infusion of norepinephrine significantly inhibited the vasodilator responses induced by ACh with 37 ± 12%, BK with 39 ± 12%, and 5HT with 44 ± 10% when compared with the responses under concomitant infusion of saline (p < 0.05 for all) (see ). For concomitant infusion of clonidine, the inhibition of Ach-, BK-, and 5HT-mediated vasodilatation was 48 ± 18%, 45 ± 12%, and 40 ± 12%, respectively (p < 0.05 for all compared with saline) (see ), whereas it was 65 ± 12%, 34 ± 10% and 44 ± 8% for concomitant infusion with L-NMMA (p < 0.05 for all compared with saline) (see ).
Study protocol 2
The intra-arterial infusion with methoxamine alone reduced forearm blood significantly from 3.51 ± 0.29 to 2.03 ± 0.23 mL/min per 100 mL forearm tissue (n = 16; p < 0.05). shows the effects of intra-arterially infused cumulative dose infusions of ACh, BK, and 5HT on forearm blood flow. During the infusion of saline, all three endothelium-dependent vasodilators induced a significant dose-dependent increase in forearm blood flow (p < 0.05 for all). The effects of ACh, BK, and 5HT were significantly attenuated by the concomitant infusion of the selective α1-receptor agonist methoxamine compared with saline (p < 0.05), except for the highest dose of ACh used.
Table 2 Forearm blood flows (mL/min per 100 mL forearm tissue) before and during intra-arterial infusions of acetylcholine (100 and 1000 ng/kg/min), bradykinin (0.01 and 0.1 ng/kg/min), and 5-hydroxytryptamine (0.3 and 1.0 ng/kg/min)
Discussion
The main finding of this study is that activation of α1-adrenoceptors inhibits endothelium-dependent nitric oxide-mediated vasodilation. That the α-adrenoceptor activation did not influence the vasodilator response induced by the nitric oxide donor SNP indicates that α-adrenoceptor activation attenuates the release of endogenous nitric oxide and not the vascular response to nitric oxide per se.
It is well established that ACh, BK, and 5HT are endothelium-dependent nitric oxide-mediated vasodilators (CitationFurchgott and Zawadzki 1980; CitationIgnarro 1989; CitationAngus and Lew 1992; CitationBruning et al 1993; CitationCockcroft et al 1994). This was confirmed in the present study by showing that the vasodilator responses evoked by these vasoactive substances were significantly attenuated by L-NMMA. The fact that the vasodilation induced by the exogenous nitric oxide donor SNP was not influenced by L-NMMA confirms that L-NMMA attenuates specifically the production of endogenous nitric oxide and not the vascular response to nitric oxide (CitationJie et al 1984; CitationAngus and Lew 1992; CitationBruning et al 1993).
In the first set of experiments, we showed that the vasodilator responses induced by ACh, BK, and 5HT were significantly attenuated by a simultaneous infusion of the non-selective α1- and α2-agonists norepinephrine and clonidine, indicating α1- and/or α2-receptor activation inhibits endothelium-dependent vasodilation. The finding in the second set of experiments that the selective α1-receptor agonist methoxamine also reduced the vasodilation induced by the three different endothelium-dependent vasodilators used provides evidence that in any case α1-adrenoceptor activation inhibits endothelium-dependent nitric oxide-mediated vasodilation. Based on the present findings, the role of the α2-receptor remains unclear because we had no selective α2-receptor agonist available for human use except clonidine. Although clonidine has preferential affinity for the α2-receptor, it has also affinity for the α1-adrenoceptor (CitationBradshaw et al 1981).
That norepinephrine and clonidine did not influence the vasodilator response elicited by SNP shows that reduction of basal forearm blood flow per se (ie, vasoconstriction) does not explain the attenuated responses of the endothelium-dependent vasodilators. Although it can be argued that the vasodilator response induced by SNP was much greater than those induced by BK and 5HT, it was of the same magnitude as the response induced by ACh. Furthermore, since SNP is an exogenous donor of nitric oxide, it provides evidence that α-adrenoceptor activation does not inhibit the vasodilator response induced by nitric oxide.
These findings corroborate the results of the recent experiments of CitationHijmering et al (2002) showing that sympathetic activation inhibits flow-mediated vasodilation in the human forearm. Flow-mediated vasodilation is mediated via shear stress, inducing the release of nitric oxide (CitationKoller et al 1994; CitationDavies et al 1997). It is often argued that shear stress-induced release of nitric oxide is under the control of different mechanisms than agonist- or microvascular-mediated nitric oxide release. This is based on the observations that adrenoceptor activation does not influence ACh- and metacholine-induced vasodilation (CitationRector et al 1993; CitationEngelke et al 1997). However, in these experiments, relatively high doses of ACh and metacholine were used, and it has been shown that ACh and its synthetic equivalent, metacholine, induce a nitric oxide-independent vasodilation in the higher dose range (CitationBruning et al 1996; CitationKamper et al 2002). This is in accordance with the present finding that, in contrast with the lower dose of ACh used, the vasodilator response induced by the highest dose of ACh was not influenced by the selective α1-receptor agonist, methoxamine. However, when CitationEngelke et al (1997) used a lower dose of ACh comparable to our dose of 100 ng/kg/min, the responses to lower body suction and phentolamine were significantly attenuated, corroborating our results.
From the present findings and the literature, evidence emerges that the endocrine function of the endothelial cells is controlled, in part, by the autonomic nervous system (CitationToda and Okamura 2003). Stimulation of the cholinergic M3-receptors and β2-adrenoceptors induce the release of nitric oxide (CitationBruning et al 1995; CitationDawes et al 1997; CitationMajmudar et al 1999); whereas, as shown in the present study, activation of α1-adrenoceptors attenuates the release of nitric oxide. The α2-receptors seem to have a dual effect on nitric oxide release, whereby the inhibitory effects seem to be most prominent in the human forearm vascular bed (CitationJie et al 1984). These findings might be an explanation for the better survival of heart failure patients treated with the non-selective β-blocker carvedilol, which has α1-receptor blocking properties, compared with patients treated with the selective β1-blocker metoprolol (CitationPoole-Wilson et al 2003). Heart failure is associated with sympathetic activation and endothelial dysfunction (CitationCohn et al 1984; CitationFerguson et al 1990). It could be hypothesized that the additional α1-receptor-blocking properties of carvedilol might lead to improved endothelial mediated, nitric oxide-induced vasodilation in these patients; thus further resulting in a reduction in peripheral vascular resistance and reducing impedance to flow, and hence lowering the work-load placed on the myocardium, which is always beneficial in a patient with a failing heart. This, however, needs to be further investigated.
Limitations of the present study
The conclusion of the present study – that in humans, endothelium-dependent nitric oxide-mediated vasodilation is attenuated by activation of peripheral α1-adrenoceptors – is based on the lack of change to SNP-mediated blood flow. There exists an indirect measurement of nitric oxide by changes in nitrite/nitrate levels (CitationKawano et al 1999). This measurement was not performed in this study. Another explanation of a reduced nitric oxide-mediated vasodilation is that the bioavailability of the endothelium-generated nitric oxide is reduced due to impaired NADPH-oxidase activity (CitationDe Vriese et al 2000).
From the present findings, we conclude that in humans, endothelium-dependent nitric oxide-mediated vasodilation is attenuated by activation of peripheral α1-adrenoceptors.
References
- AngusJALewMJInterpretation of the acetylcholine test of endothelial cell dysfunction in hypertensionJ Hypertens199210S17986
- BradshawCMStokerMJSzabadiEThe effects of clonidine on cortical neurones: evidence for its action as a partial agonist at α1-adrenoceptorsBrit J Pharmacol1981747578
- BruningTAChangPCBlauwGJSerotonin-induced vasodilatation in the human forearm is mediated by the “nitric oxide-pathway”: no evidence for involvement of the 5-HT3-receptorJ Cardiovasc Pharm1993224451
- BruningTAChangPCHendriksMGIn vivo characterization of muscarinic receptor subtypes that mediate vasodilatation in patients with essential hypertensionHypertension1995267077607735
- BruningTAChangPCKemmeMJComparison of cholinergic vasodilator responses to acetylcholine and methacholine in the human forearmBlood Press19965333418973750
- ChangPCVerlindeRBruningTA microcomputer-based, R-wave triggered system for hemodynamic measurements in the forearmComput Biol Med198818157633396336
- CockcroftJRChowienczykPJBrettSEEffect of NG-monomethyl-L-arginine on kinin-induced vasodilation in the human forearmBr J Clin Pharmacol199438307107530473
- CohnJNLevineTBOlivariMTPlasma norepinephrine as a guide to prognosis in patients with chronic congestive heart failureN Engl J Med1984311819236382011
- DaviesPFVolinMVJosephLBornCVRSchwartzCJEndothelial responses to hemodynamic shear stress: spatial and temporal considerationsVascular endothelium1997StuttgartSchattauer16776
- DawesMChowienczykPJRitterJMEffects of inhibition of the L-arginine/nitric oxide pathway on vasodilation caused by beta-adrenergic agonists in human forearmCirculation199795229379142007
- de VrieseASVerbeurenTJVan de VoordeJEndothelial dysfunction in diabetesBr J Pharmacol20001309637410882379
- EngelkeKAWilliamsMMDietzNMDoes sympathetic activation blunt nitric oxide-mediated hyperemia in the human forearm?Clin Auton Res1997785919174656
- FergusonDWBergWJSandersJSClinical and hemodynamic correlates of sympathetic nerve activity in normal humans and patients with heart failure: evidence from direct microneurographic recordingsJ Am Coll Cardiol1990161125342229759
- FurchgottRFZawadzkiJVThe obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholineNature198028837366253831
- HijmeringMLStroesESOlijhoekJSympathetic activation markedly reduces endothelium-dependent, flow-mediated vasodilationJ Am Coll Cardiol200239683811849869
- IgnarroLJEndothelium-derived nitric oxide relaxes nonvascular smooth muscleFASEB J198933162642868
- JieKVan BrummelenPVermeyPIdentification of vascular postsynaptic alpha 1- and alpha 2-adrenoceptors in manCirc Res198454447526325037
- KamperAMde CraenAJMBlauwGForearm blood flow measurements using computerized strain-gauge venous occlusion plethysmography: unilateral versus bilateral measurementsClin Physiol20015524711576152
- KamperAMPaulLCBlauwGJProstaglandins are involved in acetylcholine- and 5-hydroxytryptamine-induced, nitric oxide-mediated vasodilatation in human forearmJ Cardiovasc Pharmacol200240922912451326
- KawanoHMotoyamaTHirashimaOHyperglycemia rapidly suppresses flow-mediated endothelium-dependent vasodilation of brachial arteryJ Am Coll Cardiol1999341465410400004
- KollerASunDHuangACorelease of nitric oxide and prostaglandins mediates flow-dependent dilation of rat gracilis muscle arteriolesAm J Physiol1994267H326328048598
- LemboGIaccarinoGVecchioneCInsulin modulation of an endothelial nitric oxide component present in the alpha2- and beta-adrenergic responses in human forearmJ Clin Inves1997100200714
- LiaoJKHomeyCJThe release of endothelium-derived relaxing factor via alpha 2-adrenergic receptor activation is specifically mediated by Gi alpha 2J Biol Chem199326819528338103519
- MajmudarNGAnumbaDRobsonSCContribution of nitric oxide to beta2-adrenoceptor mediated vasodilatation in human forearm arterial vasculatureBr J Clin Pharmacol199947173710190652
- Poole-WilsonPASwedbergKClelandJGComparison of carvedilol and metoprolol on clinical outcomes in patients with chronic heart failure in the Carvedilol Or Metoprolol European Trial (COMET): randomised controlled trialLancet200336271312853193
- RectorTSBankAJde BruynVHEffects of norepinephrine on endothelium-dependent vasodilation of forearm resistance vesselsClin Pharmacol Ther19935337498453857
- SartoriCScherrerUInsulin, nitric oxide and the sympathetic nervous system: at the crossroads of metabolic and cardiovascular regulationJ Hypertens19911715172510608463
- TodaNOkamuraTThe pharmacology of nitric oxide in the peripheral nervous system of blood vesselsPharmacol Rev20035527132412773630
- ZanzingerJRole of nitric oxide in the neural control of cardiovascular functionCardiovasc Res1999436394910690335