Abstract
In the past few years the classical concept of the renin–angiotensin system (RAS) has experienced substantial conceptual changes. The identification of the renin/prorenin receptor, the angiotensin converting enzyme homologue ACE2 as an angiotensin peptide processing enzyme, Mas as a receptor for Ang-(1–7) and the possibility of signaling through ACE, have contributed to switch our understanding of the RAS from the classical limited proteolysis linear cascade to a cascade with multiple mediators, multiple receptors, and multi functional enzymes. In this review we will focus on the recent findings related to RAS and, in particular, on its role in diabetes by discussing possible interactions between RAS mediators, endothelium function, and insulin signaling transduction pathways as well as the putative role of ACE2-Ang-(1–7)-Mas axis in disease pathogenesis.
Introduction
In a classical view, the renin–angiotensin system (RAS) is considered an endocrine system whose active metabolite, the angiotensin II (Ang II), is produced by enzymatic sequential cleavage from the angiotensinogen substratum of hepatic origin (CitationWeber et al 2001; CitationZaman et al 2002). In the bloodstream, the renin, a highly specific protease, converts the circulating angiotensinogen into the decapeptide Ang I, which, in turn is converted to Ang II by the converting enzyme (ACE) action (CitationWeber et al 2001; CitationZaman et al 2002). This enzyme also inactivates bradykinin (BK) and is highly found in the endothelial cells membranes of the pulmonary circulation (CitationWeber et al 2001; CitationZaman et al 2002; CitationCarey and Siragy 2003a).
In the past few years the classical concept of the RAS has experienced substantial conceptual changes. Although this system is known for decades, recent discoveries in cellular and molecular biology, as well as cardiovascular and renal physiology, have introduced larger understanding about its function in physiological situations and in several diseases (CitationCarey and Siragy 2003a; CitationChappell et al 2004; CitationSimões e Silva et al 2006a; CitationSantos and Ferreira 2007; CitationChappell 2007).
One of the most relevant conceptual changes in our understanding of the RAS was the discovery of local or tissue RAS (CitationMiyazaki and Takai 2006; CitationPaul et al 2006). A local system is characterized by the presence of RAS components, such as angioten sinogen, processing enzymes, angiotensins, and specific receptors at tissue level. In the 70’s, CitationGanten and colleagues (1971) demonstrated, for the first time, that all RAS’s components could be produced locally in several organs and tissues. The local RAS has been found in the heart, blood vessels, kidney, adrenal gland, pancreas, central nervous system, reproductive system, lymphatic and adipose tissue (CitationNielsen et al 2000; CitationSernia 2001; CitationLavoie and Sigmund 2003; CitationSpät and Hunyady 2004). The local systems appear to be regulated independently of the circulatory RAS but can also interact with the latter. In this way, local RAS’s effects could occur in the cell that produces the peptides (intracrine and autocrine functions), in adjacent cells (paracrine function) or through the bloodstream to a specific organ or tissue (endocrine function) (CitationMiyazaki and Takai 2006; CitationPaul et al 2006). The main importance of tissue RAS is linked to local regulatory mechanisms that contribute to a great number of homeostatic pathways, including cellular growth, extracellular matrix formation, vascular proliferation, endothelium function and apoptosis (CitationMiyazaki and Takai 2006; CitationPaul et al 2006).
Other important landmarks for the new concept of RAS were the characterization of angiotensin-(1–7) [Ang-(1–7)] as a biological active metabolite of RAS (CitationSimões e Silva et al 2006a; CitationSantos and Ferreira 2007), the Ang IV receptor as an insulin regulated aminopeptidase (IRAP) (CitationAlbiston et al 2001), the renin/prorenin receptor (CitationNguyen et al 2002), the ACE homologue ACE2 as an angiotensin peptide processing enzyme (CitationDonoghue et al 2000; CitationTipnis et al 2000), Mas as a receptor for Ang-(1–7) (CitationSantos et al 2003), and the possibility of signaling through ACE (CitationKohlstedt et al 2004). These discoveries have contributed to switch our understanding of the RAS from the classical limited-proteolysis linear cascade to a cascade with multiple mediators, multiple receptors, and multi functional enzymes (CitationSimões e Silva et al 2006a; CitationSantos and Ferreira 2007; CitationChappell 2007) ().
Figure 1 Novel components of the renin–angiotensin system and its interactions.
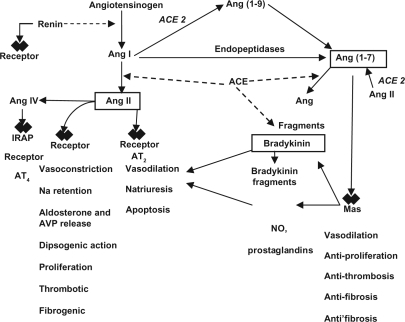
In regard to Ang-(1–7), the identification of ACE2 and Mas as a receptor implicated in its actions contributed to decisively establish this heptapeptide as a biologically active member of the RAS cascade. Most evidence supports a counter-regulatory role for Ang-(1–7) by opposing many Ang II-AT1 receptor mediated actions, especially vasoconstriction and proliferation (CitationSimões e Silva et al 2006a; CitationSantos and Ferreira 2007; CitationChappell 2007). Furthermore, Ang-(1–7) has been associated to the physiopathology of several diseases such as hypertension (CitationLuque et al 1996; CitationFerrario et al 1998; CitationSimões e Silva et al 2004, Citation2006b, Citation2006c), pre-eclampsia (CitationMerril et al 2002; CitationBrosnihan et al 2004; CitationShah 2005), hypertrophic myocardial disease and congestive heart failure, myocardial infarct (CitationZisman et al 2003; CitationSantos and Ferreira 2005), chronic renal diseases (CitationSimões e Silva et al 2006b), hepatic cirrhosis (CitationPaizis et al 2005; CitationPereira et al 2007; CitationHerath et al 2007; CitationWarner et al 2007), diabetic nephropathy (CitationCarey and Siragy 2003b; CitationTikelis et al 2003; CitationKalantrina and Okusa 2006), and gestational diabetes (CitationNogueira et al 2007).
This review will summarize the current aspects of RAS, its role in diabetes, and its well known and putative interactions with endothelium function and insulin signaling. Moreover, the importance of the RAS to diabetes prevention and its involvement on cardiovascular and renal events in diabetic patients will be discussed.
New aspects of the RAS components
Angiotensinogen
Angiotensinogen (AGT) is a glycoprotein of 452 aminoacids produced in the liver as well as in other tissues (including the heart, vessels, kidneys, and adipose tissue), which circulates as an inactive biological peptide. Through renin action, it is converted into Ang I, the precursor peptide in classical RAS cascade (CitationWeber et al 2001).
The M235T polymorphism of the AGT gene is known to be associated with circulating and tissue AGT (CitationBloem et al 1995; CitationWinkelmann et al 1999; CitationGumprecht et al 2000). However, the finding of genetic linkage between the AGT gene and hypertension in many studies and the presence of increased levels of plasma AGT in subjects with higher parental and personal blood pressure only support a role for increased synthesis of angiotensinogen in hypertension but do not explain how this translates into elevated blood pressure (CitationNaber and Siffert 2004). One hypothesis is that the genetic increase in RAS activity due to the T235 allele of AGT may lead to greater formation of Ang II in cardiac, vascular, and renal tissues, predisposing to cardiovascular and renal damage (CitationWinkelmann et al 1999; CitationGumprecht et al 2000).
Renin
Renin is an aspartyl protease enzyme produced by the juxtaglomerular apparatus of the kidney. This enzyme acts on AGT and forms the decapeptide Ang I. Renin is considered a key enzyme of the RAS due to the rate-limiting nature of its hydrolytic activity on the precursor AGT (CitationWeber et al 2001; CitationZaman et al 2002).
In recent years, this view has acquired new and important features after the discovery of the renin/prorenin receptor by CitationNguyen and colleagues (2002). Thus, according to our new understanding of the role of renin within the RAS, it serves as an Ang II-generating enzyme through the limited-proteolysis process started by its action on AGT and it can act as an agonist of the RAS inducing signaling through its receptor. In addition, binding of renin to its receptor appears to increase its catalytic efficiency on its substrate. We are still searching for the biological and physiopathological implications of these actions, but there are suggestions that it may be implicated in target-organ lesion, especially in the kidney (CitationStaessen et al 2006).
Angiotensin-converting enzyme
The key role of angiotensin-converting enzyme (ACE) within the RAS is well established since the pioneering work of CitationSkeggs and colleagues (1956), which showed that ACE is the main Ang II-generating enzyme. About 40 years later, the action of ACE in the catabolism of Ang-(1–7) was clearly established (CitationDeddish et al 1998). As shown in , by forming the potent vasoconstrictor Ang II and inactivating the vasodilator Ang-(1–7), ACE plays a central role as a pressor enzyme. Another action outside the RAS, the inactivation of the vasodilator BK, further strengthens the role of ACE as a pro-hypertensive enzyme.
However, the role of ACE is not restricted to its hydrolytic activity. Fleming and colleagues (CitationKohlstedt et al 2004) have identified a new action of ACE: the out in signaling. According to this novel view, upon binding by bradykinin (BK) and ACE inhibitors (ACEi), ACE is phosphorylated at the amino acid Ser1270 and triggers a signal cascade, which was able to increase ACE and COX-2 synthesis. In addition, the signaling through ACE was described in adipocytes and pre-adipocytes (CitationKohlstedt et al 2004).
Angiotensin-converting enzyme 2
In 2000, two independent groups (CitationDonoghue et al 2000; CitationTipnis et al 2000) identified a new enzyme homologue of ACE, which they called angiotensin-converting enzyme 2 (ACE2) (CitationDonoghue et al 2000) and captopril insensitive carboxypeptidase (CitationTipnis et al 2000). This enzyme shows a homology of about 42% with ACE, but different biochemical activities, being highly specific for angiotensin cleavage. For example, in contrast to classical ACE, ACE2 does not metabolize BK.
In the original work of CitationDonoghue and colleagues (2000), ACE2 was described as having as a primary activity the hydrolysis of Ang I, generating Ang-(1–9), serving as an indirect pathway to generate Ang II. However, this interpretation was challenged by the demonstration that Ang II was the preferable substrate for ACE2 (CitationTipnis et al 2000; CitationVickers et al 2002). Indeed, the catalytic efficiency of ACE2 against Ang II is 400-fold higher than for Ang I and leads to Ang-(1–7) formation (CitationRice et al 2004). It is also functionally remarkable that ACE2 is the main enzyme for Ang-(1–7) generation in many tissues (CitationRice et al 2004; CitationFerrario et al 2005).
Taking into account the enzymatic properties of the two ACEs and of the two main RAS mediators, Ang II and Ang-(1–7), we can make out a new vision of the RAS. In this novel concept, the RAS can be seen as a dual function system in which the vasoconstrictor/proliferative or vasodilator/antiproliferative actions are primarily driven by the ACE/ACE2 balance (CitationSimões e Silva et al 2006a; CitationSantos and Ferreira 2007; CitationChappell 2007; CitationWarner et al 2007). According to this concept, an elevated ACE activity concomitant with a reduced ACE2 activity will lead to increased Ang II generation and increased catabolism of Ang-(1–7)-favoring vasoconstriction, while the opposite profile (elevated ACE2 activity with reduced ACE activity) will decrease Ang II levels by converting it into Ang-(1–7), which, in turn, promotes vasodilation (CitationSimões e Silva et al 2006a; CitationSantos and Ferreira 2007; CitationChappell 2007; CitationWarner et al 2007).
Angiotensin II
Ang II was isolated in 1940 by CitationBraun Menendez and colleagues (1940) and Page and Helmer (1940) and was firstly characterized as a potent vasoconstrictor that increases peripheral vascular resistance and consequently elevates arterial pressure. In situations of extracellular fluid volume contraction, Ang II reduces renal sodium excretion via alterations in renal hemodynamics, direct enhancement of proximal tubule sodium bicarbonate reabsorption, and aldoster one mediated increases in late distal tubule and cortical collecting duct reabsorption (CitationHall 1991; CitationIchikawa and Harris 1991; CitationHall et al 1999; CitationBrewster and Perazella 2004). Ang II also increases thirst, salt appetite, and intestinal sodium absorption, all of which increase the extracellular fluid volume. In this context, the RAS was initially defined as an endocrine system in which circulating Ang II regulates blood pressure and electrolyte balance via its actions on vascular tone, aldosterone secretion, renal sodium handling, thirst, water intake, sympathetic activity, and vasopressin release.
Additional experimental studies as well as clinical trials using ACE inhibitors and Ang II type 1 (AT1) receptor blockers have shown that the actions of this system extend far beyond blood pressure control and electrolyte balance (CitationFerrario et al 2004). It has become clear that the deleterious actions of RAS on cardiovascular remodeling and renal function account for the beneficial effects of the ACE inhibitors and AT1receptor blockers in patients with hypertension, left ventricular hypertrophy, heart failure, and diabetic nephropathy (CitationFerrario et al 2004). However, besides circulating RAS, Ang II as well as other RAS mediators can also be locally produced and exert paracrine, intracrine, and autocrine effects (CitationMyiazaki and Takai 2006). One of the first evidence for the tissue formation of Ang II came from studies on angiotensin metabolism in sheep (CitationFei et al 1980). The octapeptide was subsequently found to be produced in numerous tissues, including the adrenal, brain, heart, kidney, vasculature, adipose tissue, gonads, pancreas, prostate, eye, and placenta (CitationNielsen et al 2000; CitationSernia 2001; CitationLavoie and Sigmund 2003; CitationSpät and Hunyady 2004; CitationNavar and Nishiyama 2004; CitationBrewster and Perazella 2004). Further investigations proposed that in the circulating RAS, renin originating from the kidney is a rate-limiting factor for Ang II generation in plasma, but in the vascular tissue, ACE and other peptidases such as chymase regulate Ang II generation (CitationMyiazaki and Takai 2006). Although not all components of the classical RAS are synthesized locally in some tissues, alternative enzymatic pathways or the presence of the renin receptor, which binds to and activates circulating renin and prorenin (CitationNguyen et al 2002), may also permit local Ang II formation. In addition, the RAS is also able to exert intracrine effects due to the intracellular formation of Ang II (CitationRe 2003; CitationBaker et al 2004). However, kinetic studies suggest that intracellular Ang II is largely derived from receptor mediated uptake of the extracellular peptide (CitationDanser 2003).
The cellular Ang II receptor was identified in 1974 as a high-affinity plasma membrane-binding site that is sensitive to guanyl nucleotides (CitationGlossmann et al 1974). Later, distinct AT1 and AT2receptor subtypes were identified by selective ligands (CitationDe Gasparo et al 2000) and were subsequently characterized as seven transmembrane receptors by molecular cloning (CitationMurphy et al 1991; CitationSasaki et al 1991; CitationKambayashi et al 1993; CitationMukoyama et al 1993). Thus, Ang II mediates effects via complex intracellular signaling pathways that are stimulated following binding of the peptide to its cell-surface receptors, AT1 and AT2(CitationMurphy et al 1991; CitationTouyz and Schiffrin 2000). In humans, AT1 receptor is widely expressed in blood vessels, heart, kidney, adrenal glands, and liver, whereas AT2 receptor is present mainly in fetal tissue, decreasing rapidly after birth, with relatively low amounts normally expressed in adult tissue. The G protein-coupled AT1 receptor mediates the known physiological and pathological actions of Ang II and undergoes rapid desensitization and internalization after agonist stimulation. In contrast, the AT2 receptor does not exhibit the latter features and acts mainly through Gi and tyrosine phosphatases to exert pre-dominantly inhibitory actions on cellular responses mediated by the AT1 receptor and growth factor receptors (CitationNouet and Nahmias 2000; CitationHunyady and Catt 2006). For example, both receptors play a role in regulating vascular smooth muscle cell (VSMC) function, although they differ in their actions. While the AT1 receptor is associated with growth, inflammation and vasoconstriction, the AT2receptor is generally associated with opposite actions stimulating apoptosis and vasodilatation (CitationTouyz and Schiffrin 2000).
Animal models and clinical data have also helped to establish that inhibition of Ang II action in nonclassical target sites, such as immune cells, explains some of the unanticipated therapeutic effects of ACE inhibitors and AT1 receptor blockers (CitationFerrario et al 2004). Parallel studies on the molecular mechanism of action of Ang II have revealed that its main target, the AT1 receptor, is one of the most versatile members of the G protein-coupled receptor (GPCR) family (CitationHunyady and Catt 2006). Ang II exerts several cytokine-like actions via the AT1receptor and can stimulate multiple signaling pathways, activate several growth factor receptors, and promote the formation of reactive oxygen species (ROS) and other proinflammatory responses (CitationHunyady and Catt 2006).
Angiotensin-(1–7)
Ang-(1–7) was regarded as an inactive component of the RAS for many years. However after the demonstration that Ang-(1–7) was the main product formed from Ang I through an ACE-independent pathway, in dog brain micro-punches homogenates (CitationSantos et al 1988) and the demonstration that this heptapeptide was equipotent to Ang II to elicit vasopressin release from neurohyphophyseal explants (CitationSchiavone et al 1988) and to decrease blood pressure upon microinjection into the nTS (CitationCampagnole-Santos et al 1989), the importance of Ang-(1–7) within the RAS became increasingly evident.
Ang-(1–7) is formed from Ang II by prolylendopeptidase, prolylcarboxipeptidase or ACE2 or directly from Ang I through hydrolysis by prolylendopeptidase and endopeptidase 24.11 and it is metabolized by ACE to Ang-(1–5) (Donoughue et al 2000; CitationTipnis et al 2000; CitationChappell et al 2004; CitationRice et al 2004). ACE inhibitors elevate plasma Ang-(1–7) concentrations by both increasing Ang I, the substrate for Ang-(1–7), as well as by preventing Ang-(1–7) degradation (see ). Ang-(1–7) is present in the circulation and in many tissues including heart, blood vessels, kidney and liver (CitationSimões e Silva et al 2006a; CitationSantos and Ferreira 2007; CitationChappell 2007).
The recent identification of ACE2 which forms Ang-(1–7) from Ang II (Donoughue et al 2000; CitationTipnis et al 2000), and the G-protein coupled receptor Mas as an Ang-(1–7) receptor (CitationSantos et al 2003) have provided biochemical and molecular evidence for the biological significance of the Ang-(1–7). In addition, the inhibitory effects of Ang-(1–7) on Ang II-induced vasoconstriction, and its growth inhibitory, antiarrhythmogenic and antithrombogenic effects imply a counter regulatory role for this angiotensin within the RAS and suggest that this may be a potential target for development of new drugs (CitationSimões e Silva et al 2006a; CitationSantos and Ferreira 2007; CitationChappell 2007).
In this regard, recent studies suggest that, at least in part, the beneficial effects of ACE inhibitors in heart and kidney diseases may be attributed to Ang-(1–7) (CitationSimões e Silva et al 2006a; CitationSantos and Ferreira 2007). These findings are in agreement with the hypothesis that the RAS is capable to self regulate its activity through the formation of Ang-(1–7). In addition, CitationKostenis and colleagues (2005) recently demonstrated that the Ang-(1–7) G-protein coupled receptor Mas can hetero-oligomerize with AT1 receptor and by so inhibit the actions of Ang II. Indeed, Mas receptor acts in vivo as an antagonist of the AT1 receptor (CitationKostenis et al 2005). We currently believe that RAS can act through two opposite arms: the major one responsible for the main actions of this system is constituted by the ACE-Ang II-AT1 receptor axis and the other, a counterregulatory arm, is formed by the ACE2-Ang-(1–7)-Mas axis (CitationSimões e Silva et al 2006a; CitationSantos and Ferreira 2007).
Angiotensin III
Ang III is generated from the metabolism of Ang II by ami nopeptidase A, which cleaves the Asp1-Arg2 bound. Ang III is a biologically active peptide of the RAS whose effects are essentially similar to those observed for Ang II though less potent. Ang III enhances blood pressure, vasopressin release, and thirst when it is centrally administrated (CitationCesari et al 2002). Ang III infusion increases blood pressure in healthy volunteers and hypertensive patients (CitationSuzuki et al 1984) as well as augments aldosterone release (CitationPlovsing et al 2003). Ang III does not affect renal function in humans (CitationPlovsing et al 2003), but it induces natriuresis probably through AT2 receptors stimulation in AT1receptor blocked rats (CitationPadia et al 2006). In addition, in cultured renal cells this peptide stimulates the expression of many growth factors, proinflammatory mediators, and extracellular matrix proteins (CitationRuiz-Ortega et al 2000). Ang III normally binds to AT1and with more affinity to AT2 receptors (CitationPadia et al 2007). However, it is still not established whether these effects of Ang III are due to direct action of this peptide at angiotensins` receptors or they are mediated by smaller peptides formed as a consequence of its degradation.
Angiotensin IV
Ang IV was generated by the enzyme aminopeptidase N, which removes the amino acid arginine from the N-terminus of Ang III sequence. Alternatively, this angiotensin fragment can be also formed directly from Ang II by the enzyme D-aminopeptidase. Central administration of Ang IV markedly enhances learning and memory in normal rodents and reverses memory deficits observed in animal models of amnesia (CitationChay et al 2004; CitationBraszko et al 2006). In the kidney, Ang IV increases renal cortical blood flow without altering systemic blood pressure and decreases Na+ transport in isolated renal proximal tubules (CitationChay et al 2004). On the other hand, other studies have shown that this peptide can decrease total and regional renal blood flow in rats (CitationLi et al 2006). It has been reported that Ang IV induces vasodilation in preconstricted endothelium-intact but not endothelium-denuded pulmonary artery through a specific binding site that is blocked by divalinal (CitationChen et al 2000). In addition, this peptide increases endothelial NO synthase activity and cellular cGMP content in porcine pulmonary arterial endothelial cells (CitationPatel et al 1998). In the heart, Ang IV reduced left ventricular pressure development and ejection capabilities but increased the sensitivity of pressure development during the systole and speeded relaxation (CitationSlinker et al 1999).
Most Ang IV actions are mediated by a specific binding site called AT4 receptor. This receptor is found in many organs such as brain, adrenal gland, kidney, lung, and heart and it mediates the majority of Ang IV actions (CitationChay et al 2004; CitationAlbiston et al 2007). However, in some circumstances, Ang IV can induce its effects by interacting with AT1 receptor (CitationLi et al 2006). It is important to mention that Ang IV is not the only ligand for AT4 receptor, since other peptides such as LVV-hemorphin 7 can bind to this receptor (CitationAlbsiton et al 2004). The AT4 receptor has been also identified as the transmembrane enzyme, insulin-regulated membrane aminopeptidase (IRAP) (CitationAlbiston et al 2001). IRAP is a type II integral membrane spanning protein belonging to the M1 family of aminopeptidases and is predominantly found in GLUT4 vesicles in insulin-responsive cells. Three hypotheses for the memory-potentiating effects of the AT4 receptor/IRAP ligands, Ang IV and LVV-hemorphin 7, are proposed: acting as potent inhibitors of IRAP, they may prolong the action of endogenous promnestic peptides; they may modulate glucose uptake by modulating trafficking of GLUT4; IRAP may act as a receptor, transducing the signal initiated by ligand binding to its C-terminal domain to the intracellular domain that interacts with several cytoplasmic proteins (CitationChay et al 2004; CitationAlbiston et al 2007). However, the precise meaning of this finding remains to be elucidated.
Other angiotensin peptides
The physiological relevance of other angiotensin peptides is still unclear. Indeed, many studies have been showing the presence of these angiotensins in diverse clinical conditions. It is possible that the formation of other angiotensins could be an alternative mechanism to override different levels of RAS blockade, thus interfering with the action of ACE inhibitors or AT1receptor blockers.
Recent reports have attributed some biological actions to the smaller fragment of the RAS as the Ang-(3–7). A result of the degradation of Ang-(1–7), Ang II, and Ang IV by amino-peptidase or carboxypeptidase, it is believed that this angiotensin plays important role in the brain (CitationKarwowska-Poleska et al 1997) and kidney (CitationHanda 1999). Indeed, Ang-(3–7) has an affinity for AT4 receptors that did differ from Ang IV.
The issue whether Ang-(1–9) itself has direct biological effects or whether its supposed actions are elicited by generation of different metabolites, especially Ang-(1–7), remains unsolved. In some pathological conditions, its concentration changes significantly, indicating that it could be involved in those diseases (CitationOcaranza et al 2006). Ang-(1–9) can be formed direct from Ang I through catalytic action of ACE2 and it is metabolized by ACE and NEP to generate Ang-(1–7) (CitationRice et al 2004). It has been reported that this peptide enhances BK actions, increases NO and arachidonic acid release (CitationJackman et al 2002), and is involved in the regulation of platelet function (CitationMogielnicki et al 2003).
Ang A, also a novel angiotensin peptide, was firstly detected in plasma from healthy humans and in end-stage renal disease patients (CitationJankowski et al 2007). Ang A concentrations were less than 20% of the Ang II concentrations in healthy subjects, but the ratio of Ang A/Ang II has been shown as higher in end-stage renal disease patients. In a different way from other angiotensin fragments, Des[Asp1]-[Ala1]-Ang II is generated by decarboxylation reaction instead of cleavage process. Thus, the first amino acid of the Ang II sequence (aspartic acid) is converted into alanine. Conventional enzyme immunoassays do not distinguish between Ang II and Ang A. Otherwise, these assays quantify the sum of Ang II and Ang A. Ang A has similar affinity to the AT1 receptor as Ang II, but a higher affinity to the AT2 receptor. However, its activity in in vitro preparations is lower when compared with Ang II. Indeed, Ang A is a less potent and only partial agonist at the AT1receptor. On the other hand, due to its high affinity to the AT2 receptor, it could be an endogenous Ang II regulatory peptide (CitationJankowski et al 2007).
The demonstration of endogenous Ang-(1–12) in the rat may foretell renin independent pathways that lead to the formation of biologically active peptides (CitationNagata et al 2006). Indeed, the peptide bond Tyr12-Tyr13 hydrolyzed to produce Ang-(1–12) from rat angiotensinogen is structurally distinct from the Leu10-Leu11 bond recognized by rat renin to form Ang I. Moreover, these bonds are also distinct for human angiotensinogen (CitationNagata et al 2006). Despite the absence of a functional effect attributed to Ang-(1–12), the processing to this intermediate peptide may comprise another level of RAS regulation.
and summarize the results obtained by recent basic and clinical research on RAS.
Table 1 Summary of recent basic research on renin–angiotensin system
Table 2 Summary of recent clinical research on renin–angiotensin system
RAS and diabetes
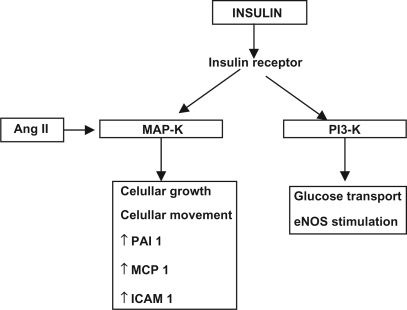
The association of the RAS with the endocrine system is particularly illustrated by the prominent role of Ang II in diabetes and metabolic syndrome. The frequent association of diabetes mellitus (DM) with hypertension, retinopathy, nephropathy, and cardiovascular disease has implicated the RAS in the initiation and progression of these disorders. This has been demonstrated by clinical trials in which RAS inhibitors significantly reduced the incidence of vascular complications in DM patients (UKPDS 1998; CitationHOPE 2000; CitationBrenner et al 2001; CitationLewis et al 2001; CitationParving et al 2001; CitationJandeleit-Dahm et al 2005). These improvements appear to result from protective actions upon skeletal muscle (CitationMuller et al 1997; CitationFrossard et al 2000) and pancreatic islets (CitationLupi et al 2006), and also from enhanced insulin sensitivity associated with decreased adipocyte size (CitationFuruhashi et al 2004), as well as increased transcapillary glucose transport (CitationMuller et al 1997; CitationFrossard et al 2000). Ang II can also cause insulin resistance by interfering with the insulin-stimulated increase in insulin receptor substrate 1-associated PI3K activity (CitationFolli et al 1999) ().
Figure 2 Interactions between angiotensin II and insulin receptor in vascular smooth muscle cells.
In addition, the renal RAS is clearly activated in DM, with increased tissue Ang II (CitationGiacchetti et al 2005) that leads to the development of diabetic nephropathy, a major cause of end-stage renal disease. Blockade of the RAS could thus reduce tissue Ang II levels, with beneficial effects on cardiovascular and renal function.
The so-called metabolic syndrome, now regarded as a global epidemic, includes obesity, insulin resistance, dyslipidemia, and hypertension, often with hypertriglyceridemia and reduced high-density lipoprotein, as well as hyperuricemia and increased C-reactive protein (CitationMeigs 2003). These features are often associated with endothelial cell dysfunction with impaired control of vascular tone, increased adhesiveness of leukocytes, and increased formation of growth factors and cytokines. These processes have been implicated in the development of diabetes, hypertension, heart failure, atherosclerosis, and renal failure. In this context, RAS components, mostly Ang II, have a potential role in endothelial cell dysfunction, insulin resistance, inflammation, and proliferative effects (CitationWatanabe et al 2005).
Interactions between RAS, endothelium, and insulin signaling pathways
The endothelium is a dynamic autocrine and paracrine organ that regulates vascular tonus and the interaction of the vascular wall with circulating substances and plasma cells (CitationCardillo and Panza 1998). It produces vasodilator and vasoconstrictor substances that are in balance under normal conditions. The health of the vascular net depends though on the normal operation of the endothelium. The endothelial dysfunction has been considered an imbalance in which the vasoconstrictor effects overcome the vasodilator ones in the vascular tonus (CitationJansson 2007; CitationPechánová et al 2007). This imbalance leads to a decrease of NO activity, which implies in a damage of the vascular protection (CitationPechánová et al 2007). The endothelium generally acts as a preventing barrier to the exocytose of macrophages and of low-density lipoproteins (LDL) to the subendothelium, and, thus protects against the atherosclerosis process (CitationRomeo et al 2007).
The RAS can influence the vascular function by several ways, and studies have shown AT1 and AT2 receptors in the endothelium cells (CitationJacques et al 2003), besides evidence of Mas receptors, the Ang-(1–7) receptor (CitationSampaio et al 2007a, Citation2007b). Ang II generally exhibits a vasoconstrictor effect on the endothelium while Ang-(1–7) is preferentially a vasodilator peptide (CitationSampaio et al 2007b). Thus, the balance of these angiotensins’ effects is probably involved in the maintenance of endothelium integrity. It is well established that Ang II produces endothelial dysfunction through different pathways, such as increasing the oxidative stress (CitationAgarwal 2003) and exerting proliferative (CitationClark et al 2008) and prothrombotic activities (CitationSpillert et al 1994; CitationWatanabe et al 2005). Ang II also stimulates the production of superoxide radicals, TGF-β, endothelin, and plasminogen activator inhibitor (PAI-1), which ultimately interferes in NO action (CitationRodrigues et al 2007). Thus, there is enough evidence that Ang II accumulation damages the endothelium and leads to atherosclerosis (CitationWatanabe et al 2005; CitationCippolone et al 2006). However, other mediators are also responsible for endothelial function regulation (CitationSavoia and Schiffrin 2007). In this regard, the vasoconstriction actions produced by Ang II and endothelin are probably opposed by the release of NO, prostaglandins, bradykinin as well as by Ang-(1–7). Furthermore, Ang-(1–7) promotes the release of NO and prostaglandins (CitationBrosnihan et al 1996; CitationPörsti et al 1994) and potentiates bradykinin effects in different experimental models (CitationPaula et al 1995; CitationBrosnihan et al 1996; CitationMaia et al 2004). In addition, Ang (1–7) also inhibits the growth of VSMC (CitationFreeman et al 1996), platelet aggregation and thrombosis (CitationKucharewicz et al 2002; CitationFraga-Silva et al 2008), inflammation, fibrosis (CitationPereira et al 2007), and oxidative stress (CitationBenter et al 2008), which in turn might lead to endothelial function restoration.
Indeed, Ang-(1–7) can antagonize Ang II effects not only by the stimulation of other vasodilators but also through AT1 receptor inhibition. In this regard, CitationKostenis and colleagues (2005) reported that Mas receptor seems to act as a physiological antagonist of AT1 receptor and by so counteracts many Ang II actions ().
Alterations in the synthesis and degradation of endothelial factors have recently been described as a contributor to the beginning or maintenance of a series of complications, including changes in vascular reactivity, blood flow, and tissue perfusion, which are all present in diabetes, renal diseases, heart failure, and hypertension (CitationKang et al 2002; CitationSimões e Silva et al 2006c; CitationNakagawa et al 2007). Recently, studies have shown the presence of reduced levels of Ang-(1–7) in pregnancy complicated by pre-eclampsia (CitationMerril et al 2002) and gestational diabetes (CitationNogueira et al 2007), corroborating to the hypothesis that changes in circulating levels of Ang-(1–7) can be a marker of a new onset endothelial dysfunction.
Clinical and experimental evidence show that ACE inhibitors and AT1 receptor blockers clearly improve endothelial dysfunction. Many mechanisms might take part in this effect and one of them is the increase of bradykinin levels with ACE inhibitors (CitationWeir 2007). Considering that the treatment with RAS blockers (ACE inhibitors and AT1 receptor blockers) also increases Ang-(1–7) levels (CitationCampbell 2003; CitationSimões e Silva et al 2006b), our group and other investigators have speculated that the beneficial action of RAS blockade could be, at least in part, due to Ang-(1–7) effects (CitationSimões e Silva et al 2006a; CitationSantos and Ferreira 2007; CitationChappell 2007).
In regard to diabetes, ACE inhibitors have also been associated to a decrease in the incidence of type 2 diabetes in hypertensive patients (CitationYusuf et al 2001; CitationVijayaraghavan and Deedwania 2005; CitationSchmieder et al 2007), besides their wide spread utilization to prevent renal and cardiovascular diabetes complications (CitationWeir 2007). Recent studies have shown an interaction between RAS and the insulin signaling pathways through AT1 receptors (CitationVelloso et al 2006), and the utilization of AT1 receptor blockers have shown similar protective effects on renal and cardiovascular diabetes complications (CitationBrenner et al 2001; CitationLindholm et al 2002). While Ang II may contribute to endothelial dysfunction and chronic diabetes complications, especially in the presence of insulin resistance (CitationHsueh and Quiñones 2003), treatment with ACE inhibitors or AT1 receptor blockers has shown to be protective in diabetes (CitationBrenner et al 2001; CitationLindholm et al 2002). However, these studies have not addressed the possible role of Ang-(1–7) in this unclear mechanism by which ACE inhibitors or AT1 receptor blockers prevent diabetes complications and novel diabetes onset.
Interactions between endothelial factors and insulin signaling
Several studies in humans and animals have shown that insulin stimulated NO from the vascular endothelium represents an important physiologic role in the increase of the capillary recruitment and of the total blood flow to the skeletal muscle (CitationVelloso et al 2006). The blunted endothelium NO-release has also been implicated in the reduction of capillary recruitment and of total blood flow, leading to a state of low glucose availability that characterizes insulin resistance (CitationHsueh and Quiñones 2003; CitationVincent et al 2003).
Besides its NO-dependent effects in the vascular endothelium, insulin also interferes with hemodynamic homeostasis in insulin resistant states (CitationYki-Jarvinen 2003). In these conditions, insulin can act through MAP-K pathway, promoting cellular growth and other pro-inflammatory and pro-thrombotic actions (CitationVelloso et al 2006). In obese individuals, it has been speculated a possible shift from PI3-K pathway, the molecular via for insulin signal transduction into action, to the MAP-K pathway, offering a molecular explanation by which hyperinsulinemia of these individuals might play a pro-atherogenic role (CitationHsueh and Quiñones 2003). There is also a possible interaction between insulin and local RAS at this level, since Ang II through AT1 receptors may stimulate the MAP-K pathway and, by doing so, promotes cellular proliferation, inflammation and thrombosis (CitationVelloso et al 2006), while the activation of AT2 receptors could inhibit this same via, meanwhile activating PI3-K (CitationBedecs et al 1997) (). Although a possible antiproliferative action of Ang-(1–7) in this scenario is only speculative, experimental evidence with cardiomyocytes (CitationTallant et al 2005), pulmonary tumor cells (CitationGallagher and Tallant 2004), and liver tissue (CitationPereira et al 2007; CitationHerath et al 2007; CitationWarner et al 2007) corroborates this possibility.
Figure 3 Interactions between renin–angiotensin system, kinin–kallikrein system, and insulin signaling transduction pathways.
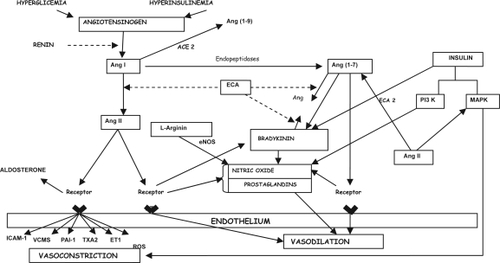
In addition, many studies have reported a connection between insulin resistance and RAS (CitationMatayoshi et al 2007). It was shown that Ang II infusion induced insulin resistance (CitationRichey et al 1999) and the blockade of the RAS improved insulin sensibility (CitationFuruhashi et al 2004). In this regard, CitationSantos and colleagues (2008) recently reported that, despite normal body weight, Mas-knockout mice in FVB/N background presented dyslipidemia, increased levels of insulin and leptin, and an approximately 50% increase in abdominal fat mass. Mas deletion also led to glucose intolerance and reduced insulin sensitivity as well as a decrease in insulin-stimulated glucose uptake by adipocytes and decreased GLUT4 in adipose tissue. These results indicated that Mas deficiency in FVB/N mice induces a metabolic syndrome-like state. Further studies are obviously necessary to address the pathophysiological role of ACE2-Ang-(1–7)-Mas axis in human diabetes.
RAS, diabetes, and the endothelium
The cardiovascular morbidity and mortality related to diabetes, especially type 2 diabetes, are very high. Recently, cardiovascular risk has been reaching enormous proportions due to the obesity epidemic, metabolic syndrome, and type 2 diabetes. Large clinical trials have been conducted to address the increase in renal and cardiovascular complications in the diabetic patient. The results of United Kingdom Prospective Diabetes Study (UKPDS), Reduction of Endpoints in Non Insulin Dependent Diabetes Mellitus with the Angiotensin II Antagonist Losartan (RENAAL), Irbesartan Diabetic Nephropathy Trial (IDNT) Patients with Type 2 Diabetes and Microalbuminuria Study (IRMA2), and Heart Outcomes Prevention Evaluation (HOPE and MICRO-HOPE) have shown that a well defined treatment of diabetic patients translated into renal and cardiovascular protection. Through these studies, it is now well established that prevention or reduction in proteinuria, blood pressure control, glycemic control, and particularly, the blockade of RAS are essential to prevent or delay the vascular diabetes complications. As already mentioned, RAS blockade has been shown not only a protective effect upon renal and cardiovascular complications of type 2 diabetes (CitationWeir 2007), but also reduce novel type 2 diabetes onset in hypertensive patients as shown by HOPE (CitationLindholm et al 2002) and LIFE studies (CitationDahlof et al 2002).
Studies of RAS in diabetes have shown conflictive results regarding the activation of this system (CitationPrice et al 1999; CitationBojestig et al 2000). For example, in diabetes nephropathy, studies have shown a suppression of plasma renin activity (CitationBjork 1990), while tissue renin are generally elevated (CitationAnderson et al 1993). Therefore, although the evaluation of the circulating RAS components may be of importance, it may not give full information of intra renal RAS components.
Adipose tissue secretes a variety of hormones (known collectively as adipokines) that can modulate endothelial function. For example, TNF-α is a proinflammatory cytokine secreted by adipose cells that may cause insulin resistance and endothelial dysfunction (CitationRydén and Arner 2007). In addition, components of RAS, including AGT, renin, ACE, Ang II, and the AT1 and AT2 receptors are present in adipose tissue (CitationSarzani et al 2008). Ang II, through AT1 receptor, is able to modulate insulin actions, and this may be due to inhibition of cross talk between Ang II signaling and insulin signaling in metabolic and vascular tissues (CitationCaglayan et al 2005). In this regard, the addition of Ang II to endothelial cells activates MAP-K pathways, leading to increased serine phosphorylation of IRS-1, impaired PI3-K activity, and endothelial dysfunction, which is abolished by AT1 receptor blockers (CitationNakashima et al 2006). In addition to effects on insulin signaling, the activation of AT1 receptors stimulates the production of ROS via NADPH oxidase, increases expression of ICAM-1, and ET-1 release from the endothelium (CitationCaglayan et al 2005). Therefore, there are multiple direct and indirect mechanisms by which RAS contribute to insulin resistance and endothelial dysfunction through the modulation of insulin signaling and other metabolic and vascular pathways (CitationCaglayan et al 2005), supporting the therapeutic institution of RAS inhibition.
RAS blockade may be achieved through the inhibition of Ang II formation (with ACE or renin inhibitors), and through direct inhibition of AT1 receptors. In clinical scenario, ACE inhibitors and AT1 receptor blockers have been considered the most important ways of RAS blockade. The mechanism of action of ACE inhibitors results in accumulation of BK, whereas it does not occur with the AT1 receptor blockers. Other differences include the blockade of Ang II formation through ACE-independent pathways by AT1 receptor blockers (CitationLi et al 2004), and a more prominent reduction of plasma blood flow by AT1 receptor blockers and renin inhibitors (CitationLi et al 2004). However, these agents have exhibited similar beneficial results in renal and cardiovascular diabetes complications (CitationYusuf et al 2001; CitationLindholm et al 2002).
In the HOPE study, diabetes incidence was lower in the group treated with ramipril when compared with placebo (CitationYusuf et al 2001). In the LIFE (Losartan Intervention For Endpoint Reduction in Hypertension) study, the use of losartan was associated with a reduction in the occurrence of new diabetes onset as compared with atenolol (CitationLindholm et al 2002). Although the precise mechanisms involved on the reduction of new type 2 diabetes diagnosis by RAS inhibition are not well established, the increase in insulin mediated glucose uptake (CitationJamerson et al 1996), the improvement of endothelial function (CitationMukai et al 2002), a decrease in inflammatory response (CitationMervaala et al 1999), an increase in BK and Ang-(1–7) levels (CitationMaia et al 2004), and an effect in local system in the pancreatic islets (CitationLau et al 2004) have been suggested as some of the putative factors (CitationCooper et al 2006). To date, many studies have placed RAS inhibition as of great importance to prevent cardiovascular diabetes complications (HOPE, LIFE, and RENAAL Studies).
Concerning its preventive actions in type 2 diabetes, RAS inhibition has also been shown to improve insulin sensitivity, allowing a better insulin action or a reduction in insulin dosages in some patients (CitationDe Mattia et al 1996). Several studies, utilizing clamps for insulin sensibility evaluation, have confirmed these data in patients receiving ACE inhibitors or AT1 blockers (Yki-Jarvinen 1995).
RAS inhibition is of importance in the prognosis of renal diabetes complications of both type 1 and type 2 diabetes. Renal Ang II increases the capillary glomerular pressure, which ultimately translates into hyperfiltration and protein uria (CitationNavar et al 2000; CitationBrewster and Perazella 2004). Deleterious Ang II effects have been demonstrated in experimental studies in which its infusion resulted in increase of intraglomerular pressure and protein excretion (CitationDoria et al 1997). The resulting proteinuria associates to arterial hypertension and to the elevated risk of end stage renal disease, besides predicting cardiovascular complications of type 2 diabetes (CitationMogensen 1984; CitationDeckert 1994). The beneficial effects of ACE inhibitors or AT1 receptor blockers upon diabetes nephropathy have been understood, at least in part, as a result of the attenuation of Ang II effects on blood pressure and glomerular hemodynamics (CitationCarey and Siragy 2003b; CitationKalantrina and Okusa 2006). Besides these hemodynamic actions, Ang II also stimulates TGFβ 1 and protein kinase C through AT1 receptors and thus increases extracellular matrix and vascular permeability (CitationKagami et al 1994; CitationWolf and Ziyadeh 1997; CitationCarey and Siragy 2003b). These effects are quite similar to the results obtained with chronic hyperglycemia on the kidney, suggesting that the harmful renal hyperglycemic effects may be partially mediated by Ang II (CitationSingh et al 1999). Of interest, hyperglycemia, the insulin resistance and the elevated free fatty acids may also stimulate renal Ang II generation, thus contributing to the pathogenesis of diabetes nephropathy (CitationCarey and Siragy 2003b; CitationKalantrina and Okusa 2006). Furthermore, in the diabetic kidney, there was reduced ACE2 protein expression that was prevented by ACE inhibitor therapy (CitationTikellis et al 2003).
Many studies have also proven the role of ACE2 as a protective enzyme for cardiovascular diseases (see ). As the ACE2 gene is on the X chromosome, the studies correlating the ACE2 gene polymorphisms with cardiovascular and metabolic diseases have to take the gender into consideration meanwhile performing data analyses (CitationLieb et al 2006; CitationZhong et al 2006; CitationYang et al 2007). Although there is a lack of studies in this field, particularly in diabetes, a Chinese study has shown that patients with the metabolic syndrome exhibit an association of ACE2 gene A/G polymorphism and elevated blood pressure (CitationZhong et al 2006). Further studies should investigate the importance of these polymorphisms in diabetes, and also verify if the known polymorphisms in the RAS genes that have been studied in restenosis after percutaneous coronary intervention (CitationGross et al 2007; CitationWijpkema et al 2006) would also play a prominent role in diabetes and coronary disease.
Therefore, the identification of ACE2 in the kidney and cardiovascular system, its modulation in diabetes, and the recent description that this enzyme plays a biological role in the generation and degradation of various angiotensin peptides provide a rationale to further explore the role of ACE2-Ang-(1–7)-Mas axis in diabetic complications.
Concluding remarks
In conclusion, this review shows the physiology of the RAS, including the importance of novel mediators, receptors, and enzymatic pathways. It also shows how the current view of this system might impact diabetes and its complications, highlighting the importance of RAS inhibition in this disease treatment. Furthermore, we also provide recent evidence for putative role of ACE2-Ang-(1–7)-Mas axis in diabetes pathogenesis.
Disclosure
The authors report no conflict of interest.
References
- AbdallaSLotherHQuittererU2000AT1-receptor heterodimers show enhanced G-protein activation and altered receptor sequestrationNature40794810993080
- AgarwalR2003Proinflammatory effects of oxidative stress in chronic kidney disease: role of additional angiotensin II blockadeAm J Physiol Renal Physiol284F863912505865
- AlbistonALMcDowallSGMatsacosD2001Evidence that the angiotensin IV (AT(4)) receptor is the enzyme insulin-regulated ami nopeptidaseJ Biol Chem27648623611707427
- AlbistonALPeckGRYeatmanHR2007Therapeutic targeting of insulin-regulated aminopeptidase: heads and tails?Pharmacol Ther1164172717900701
- AlbistonALPedersonESBurnsP2004Attenuation of scopolamine-induced learning deficits by LVV-hemorphin-7 in rats in the passive avoidance and water maze paradigmsBehav Brain Res1542394315302130
- AndersonSJungFFIngelfingerJR1993Renal renin-angiotensin system in diabetes: functional immunohistochemical and molecular biological correlationsAm J Physiol265F4774868238377
- BakerKMCherninMISchreiberT2004Evidence of a novel intracrine mechanism in angiotensin II-induced cardiac hypertrophyRegul Pept12051315177915
- BedecsKElbazNSutremM1997Angiotensin II type 2 receptors mediate inhibition of mitogen activated protein kinase cascade and functional activation of SHP-1 tyrosine phosphataseBiochem J325449549230127
- BenterIFYousifMHDhaunsiGS2008Angiotensin (1-7) prevents activation of NADPH oxidase and renal vascular dysfunction in diabetic hypertensive ratsAm J Nephrol28253317890855
- BjorkS1990The renin-angiotensin system in diabetes mellitus: a physiological and therapeutic studyScand J Urol Nephrol126150
- BloemLJManatungaAKTewksburyDA1995The serum angio-tensinogen concentration and variants of the angiotensinogen gene in white and black childrenJ Clin Invest95948537883995
- BojestigMNystromFHArnqvistHJ2000The renin-angiotensin-aldosterone system is suppressed in adults with Type 1 diabetesJ Renin Angiotensin Aldosterone Syst1353611967822
- BraszkoJJWalesiukAWielgatP2006Cognitive effects attributed to angiotensin II may result from its conversion to angiotensin IVJ Renin Angiotensin Aldosterone Syst71687417094054
- Braun-MenendezEFascioloJCLeloirLF1940The substance causing renal hypertensionJ Physiol (Lond)982839816995204
- BrennerBMCooperMEde ZeeuwD2001Effects of losartan on renal and cardiovascular outcomes in patients with type 2 diabetes and nephropathyN Eng J Med3458619
- BrewsterUCPerazellaMA2004The renin-angiotensin-aldosterone system and the kidney: effects on kidney diseaseAm J Med1162637214969655
- BrosnihanKBLiPFerrarioCM1996Angiotensin-(1–7) dilates canine coronary arteries through kinins and nitric oxideHypertension2752388613197
- BrosnihanKBNevesLAAntonL2004Enhanced expression of Ang (1–7) during pregnancyBraz J Med Biol Res3712556215273828
- CaglayanEBlaschkeFTakataY2005Metabolic syndrome interdependence of the cardiovascular and metabolic pathwaysCurr Opin Pharmacol51354215780821
- Campagnole-SantosMJDizDISantosRAS1989Cardiovascular effects of angiotensin-(1–7) injected into the dorsal medulla of ratsAm J Physiol257H32492750946
- CampbellDJ2003The renin angiotensin and kallikrein kinin systemsInt J Biochem Cell Biol357849112676165
- CardilloCPanzaJA1998Impaired endotelial regulation of vascular tone in patients with systemic arterial hypertensionVasc Med3138449796077
- CareyRMSiragyHM2003aNewly recognized components of the renin-angiotensin system: potential roles in cardiovascular and renal regulationEndocr Rev2432617112788798
- CareyRMSiragyHM2003bThe intrarenal renin-angiotensin system and diabetic nephropathyTrends Endocrinol Metab1462748112890592
- CesariMRossiGPPessinaAC2002Biological properties of the angiotensin peptides other than angiotensin II: implications for hypertension and cardiovascular diseasesJ Hypertens20793912011628
- ChappellMCModrallJGDizDI2004Novel aspects of the renal Renin-Angiotensin System: Angiotensin-(1–7), ACE2 and blood pressure regulationContrib Nephrol143778915248357
- ChappellMC2007Emerging evidence for a functional angiotensin-converting enzyme 2-angiotensin-(1–7)-Mas receptor axis: More than regulation of blood pressure?Hypertension50596917785634
- ChaySRFernandoRPeckG2004The angiotensin IV/AT4 receptorCell Mol Life Sci6127283715549174
- ChenSPatelJMBlockER2000Angiotensin IV-mediated pulmonary artery vasorelaxation is due to endothelial intracellular calcium releaseAm J Physiol279L84956
- CipolloneFFaziaMLMezzetiA2006Role of angiotensin II receptor blockers in atherosclerotic plaque stabilityExpert Opin Pharmacother72778516448322
- ClarkMAGuillaumeGPierre-LouisHC2008Angiotensin II induces proliferation of cultured rat astrocytes through c-Jun N-terminal kinaseBrain Res Bull75101618158102
- CooperMETikellisCThomasMC2006Preventing diabetes in patients with hypertension: one more reason to block the renin-angiotensin systemJ Hypertens Suppl24S576316601575
- DahlofBDevereuxRBKjeldsenSE2002Cardiovascular morbidity and mortality in the Losartan Intervention for Endpoint Reduction in Hypertension Study (LIFE): a randomized trial against atenololLancet359995100311937178
- DanserAH2003Local renin-angiotensin systems: the unanswered questionsInt J Biochem Cell Biol357596812676161
- De GasparoMCattKJInagamiT2000International union of pharmacology. XXIII. The angiotensin II receptorsPharmacol Rev524157210977869
- De MattiaGFerriCLaurentiO1996Circulating cathecolamines and metabolic effects of captopril in NIDDM patientsDiabetes Care19226308742566
- DeckertT1994Nephropathy and coronary death: the fatal twins in diabetes mellitusNephrol Dial Transplant91069717800202
- DeddishPAMarcicBJackmanHL1998N-domain-specific substrate and C-domain inhibitors of angiotensin-converting enzyme: angiotensin-(1–7) and keto-ACEHypertension31912179535414
- DonoghueMHsiehFBaronasE2000A novel angiotensin-converting enzyme-related carboxypeptidase (ACE2) converts angiotensin I to angiotensin 1–9Circ Res87E1910969042
- DoriaAOnumaTWarramJH1997Synergistic effect of angiotensin II type 1 receptor genotype and poor glycaemic control on risk of nephropathy in IDDMDiabetologia401293999389421
- FeiDTCoghlanJPFernleyRT1980Peripheral production of angiotensin II and III in sheepCirc Res46I13577379257
- FerrarioCAbdelhamedAIMooreM2004Angiotensin II antagonists in hypertension, heart failure, and diabetic nephropathy: focus on losartanCurr Med Res Opin202799315025837
- FerrarioCMJessupJGallagherPE2005Effects of renin-angiotensin system blockade on renal angiotensin-(1–7) forming enzymes and receptorsKidney Int6821899616221218
- FerrarioCMMartellNYunisC1998Characterization of angiotensin-(1–7) in the urine of normal and essential hypertensive subjectsAm J Hypertens11137469524041
- FolliFSaadMJVellosoL1999Crosstalk between insulin and angiotensin II signalling systemsExp Clin Endocrinol Diabetes107133910320054
- Fraga-SilvaRAPinheiroSVGonçalvesAC2008The Antithrombotic Effect of Angiotensin-(1–7) Involves Mas-Mediated NO Release from PlateletsMol Med14283518026570
- FreemanEJChisolmGMFerrarioCM1996Angiotensin (1–7) inhibits vascular smooth muscle cell growthHypertension2810488675248
- FrossardMJoukhadarCSteffenG2000Paracrine effects of angiotensin-converting-enzyme-and angiotensin-II-receptor-inhibition on transcapillary glucose transport in humansLife Sci6614754
- FuruhashiMUraNTakizawaH2004Blockade of the renin-angiotensin system decreases adipocyte size with improvement in insulin sensitivityJ Hypertens2219778215361770
- GallagherPETallantEA2004Inhibition of human lung cancer cell growth by angiotensin-(1–7)Carcinogenesis2520455215284177
- GantenDMarquez-JulioAGrangerP1971Renin in dog brainAm J Physiol221173374330904
- GiacchettiGSechiLARilliS2005The renin-angiotensin-aldosterone system, glucose metabolism and diabetesTrends Endocrinol Metab16120615808810
- GlossmannHBaukalACattKJ1974Angiotensin II receptors in bovine adrenal cortex. Modification of angiotensin II binding by guanyl nucleotidesJ Biol Chem24966464358562
- GrossCMPerrotAGeierC2007Recurrent in-stent restenosis is not associated with the angiotensin converting enzyme D/I, angiotensinogen Thr174Met and Met235Thr, and the angiotensin-II receptor 1 A1166C polymorphismJ Invasive Cardiol19261417541127
- GumprechtJZychmaMJGrzeszczakW2000End-stage renal disease study group. Angiotensin I-converting enzyme gene insertion/deletion and angiotensinogen M235T polymorphisms: risk of chronic renal failureKidney Int585131910916074
- HallJEBrandsMWHenegarJR1999Angiotensin II and long term arterial pressure regulation: the overriding dominance of the kidneyJ Am Soc Nephrol10S2586510201880
- HallJE1991The renin-angiotensin system: renal actions and blood pressure regulationCompr Ther178171879129
- HandaRK1999Angiotensin-(1–7) can interact with the rat proximal tubule AT(4) receptor systemAm J Physiol277F75F8310409300
- [HOPE] Heart Outcomes Prevention Evaluation Study Investigators2000Effects of ramipril on cardiovascular and microvascular outcomes in people with diabetes mellitus: results of the HOPE study and MICRO-HOPE substudyLancet355253910675071
- HerathCBWarnerFJLubelJS2007Upregulation of hepatic angiotensin-converting enzyme 2 (ACE2) and Angiotensin-(1–7) levels in experimental billiary fibrosisJ Hepatol473879517532087
- HsuehWAQuiñonesMJ2003Role of endothelial dysfunction in insulin resistanceAm J Card9210J17J
- HunyadyLCattKJ2006Pleiotropic AT1 receptor signaling pathways mediating physiological and pathogenic actions of angiotensin IIMol Endocrinol209537016141358
- IchikawaIHarrisRC1991Angiotensin actions in the kidney. Renewed insight into the old hormoneKidney Int40583961745006
- JackmanHLMassadMGSekosanM2002Angiotensin 1–9 and 1–7 release in human heart: role of cathepsin AHypertension399768112019279
- JacquesDAbdel MalakNASaderS2003Angiotensin II and its receptors in human endocardial endothelial cells : role in modulating intracellular calciumCan J Physiol Pharmacol812596612733824
- JamersonKANesbittSDAmerenaJV1996Angiotensin mediates forearm glucose uptake by hemodynamic rather than direct effectsHypertension2785488613260
- Jandeleit-DahmKATikellisCReidCM2005Why blockade of the renin-angiotensin system reduces the incidence of new-onset diabetesJ Hypertens234637315716683
- JankowskiVVanholderRvan der GietM2007Mass-spectrometric identification of a novel angiotensin peptide in human plasmaArterioscler Thromb Vasc Biol2729730217138938
- JanssonPA2007Endothelial dysfunction in insulin resistance and type 2 diabetesJ Intern Med2621738317645585
- KagamiSBorderWAMillerDE1994Angiotensin II stimulates extracellular matrix protein synthesis through induction of transforming growth factor-beta expression in the rat glomerular mesangial cellsJ Clin Invest93243178200978
- KalantariniaKOkusaMD2006The renin-angiotensin system and its blockade in diabetic renal and cardiovascular diseaseCurr Diab Rep681616522275
- KambayashiYBardhanSTakahashiK1993Molecular cloning of a novel angiotensin II receptor isoform involved in phosphotyrosine phosphatase inhibitionJ Biol Chem2682454368227011
- KangD-HKanellisJHugoC2002Role of the microvascular endothelium in progressive renal diseaseJ Am Soc Nephrol138061611856789
- Karwowska-PoleckaWKulakowskaAWisniewskiK1997Losartan influences behavioral effects of angiotensin II (3–7) in ratsPharmacol Res36275839425616
- KohlstedtKBrandesRPMuller-EsterlW2004Angiotensin-converting enzyme is involved in outsidein signaling in endothelial cellsCirc Res9460714615289
- KostenisEMilliganGChristopoulosA2005G-protein-coupled receptor Mas is a physiological antagonist of the angiotensin II type 1 receptorCirculation11118061315809376
- KucharewiczIPawlakRMatysT2002Antithrombotic effect of captopril and losartan is mediated by angiotensin-(1–7)Hypertension40774912411476
- LauTCarlssonPOLeungPS2004Evidence for a local angiotensin-generating system and dose dependent inhibition of glucose stimulated insulin release by angiotensin II in isolated pancreatic isletsDiabetologia47240814722647
- LavoieJLSigmundCD2003Minireview: overview of the renin-angiotensin system-an endocrine and paracrine systemEndocrinology14421798312746271
- LewisEJHunsickerLGClarkWRfor The Collaborative Study Group2001Renoprotective effect of the angiotensin receptor antagonist irbersartan in patients with nephropathy due to type 2 diabetesN Engl J Med3458516011565517
- LiMLiuKMichalicekJ2004Involvement of chymase-mediated angiotensin II generation in blood pressure regulationJ Clin Invest1141122015232618
- LiXCCampbellDJOhishiM2006AT1 receptor-activated signaling mediates angiotensin IV-induced renal cortical vasoconstriction in ratsAm J Physiol290F102433
- LiebWGrafJGotzA2006Association of angiotensin-converting enzyme 2 (ACE2) gene polymorphisms with parameters of left ventricular hypertrophy in men. Results of the MONICA Augsburg echocardiographic substudyJ Mol Med84889616283142
- LindholmLHIbsenHDahlofB2002Cardiovascular morbidity and mortality in patients with diabetes in the Losartan Intervention For Endpoint reduction in hypertension study (LIFE). A randomized trial against atenololLancet35910041011937179
- LupiRDel GuerraSBuglianiM2006The direct effects of the angiotensin-converting enzyme inhibitors, zofenoprilat and enalaprilat, on isolated human pancreatic isletsEur J Endocrinol1543556116452552
- LuqueMMartinPMartellN1996Effects of captopril related to increased levels of prostacyclin and angiotensin-(1–7) in essential hypertensionJ Hypertens147998058793704
- MaiaLGRamosMCFernandesL2004Angiotensin – (1–7) antagonist A-779 attenuates the potentiation of bradykinin by captopril in ratsJ Cardiovasc Pharmacol436859115071356
- MatayoshiTKamideKTakiuchiS2007Relationship between insulin resistance and the renin-angiotensin system: analysis for patients with essential and renovascular hypertensionClin Exp Hypertens294798718041161
- MeigsJB2003The metabolic syndromeBr Med J32761212855493
- MerrilDCKarolyMChenK2002Angiotensin–(1–7) in normal and pre-elamptic pregnancyEndocrine182394512450315
- MervaalaEMMullerDNParkJK1999Monocyte infiltration and adhesion molecules in a rat model of high human renin hypertensionHypertension33389959931135
- MiyazakiMTakaiS2006Tissue angiotensin II generating system by angiotensin-converting enzyme and chymaseJ Pharmacol Sci100391716799256
- MogensenCE1984Microalbuminuria predicts clinical proteinuria and early mortality in maturity onset diabetesN Eng J Med31035660
- MogielnickiAKramkowskiKChabielskaE2003Angiotensin 1–9 influences hemodynamics and hemostatics parameters in ratsPol J Pharmacol555034
- MukaiYShimokawaHHigashiM2002Inhibition of renin-angiotensin system ameliorates endothelial dysfunction associated with aging ratsArterioscler Thromb Vasc Biol2214455012231564
- MukoyamaMNakajimaMHoriuchiM1993Expression cloning of type 2 angiotensin II receptor reveals a unique class of seven-transmembrane receptorsJ Biol Chem26824539428227010
- MullerMFaschingPSchmidR1997Inhibition of paracrine angiotensin-converting enzyme in vivo: effects on interstitial glucose and lactate concentrations in human skeletal muscleEur J Clin Invest27825309373760
- MurphyTJAlexanderRWGriendlingKK1991Isolation of a cDNA encoding the vascular type-1 angiotensin II receptorNature35123362041570
- NaberCKSiffertW2004Genetics of human arterial hypertensionMinerva Med953475615467511
- NagataSKatoJSasakiK2006Isolation and identification of pro-angiotensin-12, a possible component of the renin-angiotensin systemBiochem Biophys Res Comm35010263117045572
- NakagawaTSegalMCrokerB2007A breakthrough in diabetic nephropathy: the role of endothelial dysfunctionNephrol Dial Transplant222775717595179
- NakashimaHSuzukiHOhtsuH2006Angiotensin II regulates vascular and endothelial dysfunction: recent topics of Angiotensin II type-1 receptor signaling in the vasculatureCurr Vasc Pharmacol4677816472178
- NavarLGHarrisonBLImigJD2000Renal responses to AT(1) receptor blockadeAm J Hypertens13S4554
- NavarLGNishiyamaA2004Why are angiotensin concentrations so high in the kidney?Curr Opin Nephrol Hypertens131071515090867
- NguyenGDelarueFBurckleC2002Pivotal role of the renin/pro-renin receptor in angiotensin II production and cellular responses to reninJ Clin Invest10914172712045255
- NielsenAHSchauserKHPoulsenK2000Current topic: the uteropla cental renin-angiotensin systemPlacenta214687710940196
- NogueiraAISantosRASSimões e SilvaAC2007The pregnancy-induced increase of plasma angiotensin-(1–7) is blunted in gestational diabetesRegul Pep1415560
- NouetSNahmiasC2000Signal transduction from the Angiotensin II AT2 receptorTrends Endocrinol Metab111610652498
- OrcaranzaMPGodoyIJalilJE2006Enalapril attenuates downregulation of Angiotensin-converting enzyme 2 in the late phase of ventricular dysfunction in myocardial infarcted ratHypertension48572816908757
- PadiaSHHowellNLSiragyHM2006Renal angiotensin type 2 receptors mediate natriuresis via angiotensin III in the angiotensin II type 1 receptor-blocked ratHypertension475374416380540
- PadiaSHKempBAHowellNL2007Intrarenal aminopeptidase N inhibition augments natriuretic responses to angiotensin III in angiotensin type 1 receptor-blocked ratsHypertension496253017190872
- PaizisGTikellisCCooperME2005Chronic liver injury in rat and man upregulates the novel enzyme angiotensin converting enzyme IIGut541790616166274
- ParvingHHLehnertHBrochner-MortensenJfor the Irbesartan in Patients with Type 2 Diabetes and Microalbuminuria Study Group2001The effect of irbesartan on the development of diabetic nephropathy in patients with type 2 diabetesN Engl J Med345870811565519
- PatelJMMartensJRLiYD1998Angiotensin IV receptor-mediated activation of lung endothelial NOS is associated with vasorelaxationAm J Physiol275L106189843842
- PaulMMehrAPKreutzR2006Physiology of local renin-angiotensin systemsPhysiol Rev8674780316816138
- PaulaRDLimaCVKhoslaMC1995Angiotensin -(1–7) potentiates the hypotensive effect of bradykinin in conscious ratsHypertension26115497498987
- PechánováOSimkoF2007The role of nitric oxide in the maintenance of vasoactive balancePhysiol Res56Suppl 2S71617824812
- PereiraRMSantosRASTeixeiraMM2007The renin-angiotensin system in a rat model of hepatic fibrosis: Evidence for a protective role of angiotensin-(1–7)J Hepatol466748117188388
- PlovsingRRWambergCSandgaardNC2003Effects of truncated angiotensins in humans after double blockade of the renin systemAm J Physiol285R98191
- PorstiIBaraATBusseR1994Release of nitric oxide by angiotensin-(1–7) from porcine coronary endothelium: implications for a novel angiotensin receptorBr J Pharmacol11165248019744
- PriceDADe’OliveiraJMFisherND1999The state and responsiveness of the renin-angiotensin system in patients with type II diabetes mellitusAm J Hypertens123485510232494
- ReRN2003Intracellular renin and the nature of intracrine enzymesHypertension421172212860832
- RiceGIThomasDAGrantPJ2004Evaluation of angiotensin-converting enzyme (ACE), its homologue ACE2 and neprilysin in angiotensin peptide metabolismBiochem J383455115283675
- RicheyJMAderMMooreD1999Angiotensin II induces insulin resistance independent of changes in interstitial insulinAm J Physiol2779206
- RodriguezAFortuñoAGómez-AmbrosiJ2007The inhibitory effect of leptin on angiotensin II-induced vasoconstriction in vascular smoth muscle cells is mediated via a nitric oxide-dependent mechanismEndocrinology1483243117038553
- RomeoGRMoultonKSKazlauskasA2007Attenuated expression of profilin-1 confers protection from atherosclerosis in the LDL receptor null mouseCirc Res1013283017702977
- Ruiz-OrtegaMLorenzoOEgidoJ2000Angiotensin III increases MCP-1 and activates NF-kappaB and AP-1 in cultured mesangial and mononuclear cellsKidney Int5722859810844599
- RydénMArnerP2007Tumour necrosis factor-alpha in human adipose tissue – from signaling mechanisms to clinical implicationsJ Intern Med262431817875179
- SampaioWOHenrique de CastroCSantosRA2007aAngiotensin-(1–7) counterregulates angiotensin II signaling in human endothelial cellsHypertension501093817984366
- SampaioWOSouza dos SantosRAFaria-SilvaR2007bAngiotensin-(1–7) through receptor Mas mediates endothelial nitric oxide synthese activation via Akt-dependent pathwaysHypertension491859217116756
- SantosRASBrosnihanKBChappellMC1988Converting enzyme activity and angiotensin metabolism in the dog brainstemHypertension1115373277912
- SantosRASFerreiraAJ2005Cardiovascular actions of Angiotensin-(1–7)Braz J Med Biol Res3849950715962175
- SantosRASFerreiraAJ2007Angiotensin-(1–7) and the reninangiotensin systemCurr Opin Nephrol Hypertens16122817293687
- SantosRASSimões e SilvaACMaricC2003Angiotensin-(1–7) is an endogenous ligand for the G protein-coupled receptor MasProc Natl Acad Sci USA10082586312829792
- SantosSHFernandesLRMarioEG2008Mas deficiency in FVB/N mice produces marked changes in lipid and glycemic metabolismDiabetes57340718025412
- SarzaniRMarcucciPSalviP2008Angiotensin II stimulates and atrial natriuretic peptide inhibits human visceral adipocyte growthInt J Obes3225967
- SasakiKYamanoYBardhanS1991Cloning and expression of a complementary DNA encoding a bovine adrenal angiotensin II type-1 receptorNature35123032041569
- SavoiaCSchiffrinEL2007Vascular inflammation in hypertension and diabetes: molecular mechanisms and therapeutic interventionsClin Sci1123758417324119
- SchiavoneMTSantosRASBrosnihanKB1988Release of vasopressin from the rat hypothalamo-neurohypophysial system by angiotensin-(1–7) heptapeptideProc Natl Acad Sci USA85409583375255
- SchmiederREHilgersKFSchlaichPM2007Renin-angiotensin system and cardiovascular riskLancet36912081917416265
- SerniaC2001A critical appraisal of the intrinsic pancreatic angiotensin-generating systemJOP250511862023
- ShahDM2005Role of the renin-angiotensin system in the pathogenesis of preeclampsiaAm J Physiol Renal Physiol288F6142515753325
- Simões e SilvaACDinizJSPereiraRM2006bCirculating renin angiotensin system in childhood chronic renal failure: Marked increase of angiotensin-(1–7) in end-stage renal diseasePediatr Res607349
- Simões e SilvaACDinizJSRegueira-FilhoA2004The renin angiotensin system in childhood hypertension: Selective increase of angiotensin-(1–7) in essential hypertensionJ Pediatr14593815238914
- Simões e SilvaACPinheiroSVBPereiraRM2006aThe therapeutic potential of Angiotensin-(1–7) as a novel Renin Angiotensin System mediatorMini Rev Med Chem66039
- Simões e SilvaAC2006cPathophysiology of arterial hypertension: Insights from pediatric studiesCurr Pediatr Rev220923
- SinghRAlaviNSinghAK1999Role of angiotensin II in glucose-induced inhibition of mesangial matrix degradationDiabetes4820667310512375
- SkeggsLTJrKahnJRShumwayNP1956The preparation and function of the hypertensin-converting enzymeJ Exp Med103295913295487
- SlinkerBKWuYBrennanAJ1999Angiotensin IV has mixed effects on left ventricle systolic function and speeds relaxationCardiovasc Res42660910533606
- SpätAHunyadyL2004Control of aldosterone secretion: a model for convergence in cellular signaling pathwaysPhysiol Rev8448953915044681
- SpillertCRSunSMillerMA1994Hypertension-related coronary thrombosis: prothrombic role of angiotensin IIJ Natl Med Assoc8668687966432
- StaessenJALiYRichartT2006Oral renin inhibitorsLancet36814495617055947
- SuzukiSDoiYAoiW1984Effect of angiotensin III on blood pressure, renin-angiotensin-aldosterone system in normal and hypertensive subjectsJpn Heart J2575856376863
- TallantEAFerrarioCMGallagherPE2005Angiotensin-(1–7) inhibits growth of cardiac myocytes through activation of the mas receptorAm J Physiol289H15606
- TikellisCJohnstonCIForbesJM2003Characterization of renal angiotensin-converting enzyme 2 in diabetic nephropathyHypertension41392712623933
- TipnisSRHooperNMHydeR2000A human homolog of angiotensin-converting enzyme. Cloning and functional expression as a captopril-insensitive carboxypeptidaseJ Biol Chem275332384310924499
- TouyzRMSchiffrinEL2000Signal transduction mechanisms mediating the physiological and pathophysiological actions of angiotensin II in vascular smooth muscle cellsPharmacol Rev526397211121512
- VellosoLAFolliFPeregoL2006The multi-faceted crosstalk between the insulin and angiotensin II signaling systemsDiabetes Metab Res Rev229810716389635
- VickersCHalesPKaushikV2002Hydrolysis of biological peptides by human angiotensin-converting enzyme-related carboxypeptidaseJ Biol Chem277148384311815627
- VijayaraghavanKDeedwaniaPC2005The renin angiotensin system as a therapeutic target to prevent diabetes and its complicationsCardiol Clin231658315694745
- VincentMAMontagnaniMQuonMJ2003Inhibiting NOS blocks microvascular recruitment and blunts muscle glucose uptake in response to insulinAm J Physiol285E12330
- WarnerFJLubelJSMcCaughanGW2007Liver fibrosis: a balance of ACEs?Clin Sci (Lond)11310818
- WatanabeTBarkerTABerkBC2005Angiotensin II and the endothelium: Diverse signals and effectsHypertension45163915630047
- WeberKT2001Aldosterone in congestive heart failureN Engl J Med34516899711759649
- WeirMR2007Effects of renin-angiotensin system inhibition end-organ protection: can we do better?Clin Ther2918032418035185
- WijpkemaJSvan HaelstPLMonraatsPS2006Restenosis after percutaneous coronary intervention is associated with the angiotensin-II type-1 receptor 1166A/C polymorphism but not with polymorphisms of angiotensin-converting enzyme, angiotensin-II receptor, angiotensinogen or heme oxygenase-1Pharmacogenet Genomics16331716609364
- WinkelmannBRRussAPNauckM1999Angiotensinogen M235T polymorphism is associated with plasma angiotensinogen and cardiovascular diseaseAm Heart J13769870510097233
- WolfGZiyadehFN1997The role of angiotensin II in diabetic nephropathy: enphasis on non-hemodynamic mechanismsAm J Kidney Dis29153669002545
- YangWHuangWSuS2007Association study of angiotensin I converting enzyme 2 gene polymorphisms with coronary heart disease and myocardial infarction in Chinese Han populationClin Sci (Lond)1113334016822235
- Yki-JarvinenH2003Non-glycemic effects of insulinClin Cornerstone, Suppl4S612
- YusufSGersteinHHoogwerfB2001Ramipril and the development of diabetesJAMA28618828511597291
- ZamanMAOparilSCalhoumDA2002Drugs targeting the renin-angiotensin aldosterone systemNature162136
- ZhongJYanZLiuD2006Association of angiotensin converting enzyme 2 gene A/G polymorphism and elevated blood pressure in Chi nese patients with metabolic syndromeJ Lab Clin Med14791516459167
- ZismanLSKellerRSWeaverB2003Increased angiotensin-(1–7)-forming activity in failing human heart ventricles: evidence for up regulation of the angiotensin-converting enzyme homologue ACE2Circulation10817071214504186