
Abstract
Aim: SNPs in the gene for TPMT exemplify one of the most successful translations of pharmacogenomics into clinical practice. This study explains the correlation between common SNPs and variable number of tandem repeats (VNTR) in promoter of the gene. Materials & methods: We determined VNTR polymorphisms, as well as TPMT*2 and TPMT*3 SNPs and TPMT activity in Slovenian and Italian individuals and lymphoblastoid cell lines. Results: We observed a previously unreported VNTR allele, AB7C, in a TPMT*3A heterozygous individual. VNTRs with two (AB2C) and three or more (ABnC, n ≥ 3) B motifs were statistically significant in complete linkage disequilibrium (D′ = 1, r2 = 1, p < 0.0001) with the TPMT*3C and TPMT*3A alleles, respectively. Conclusion: The study provides insights into the stepwise evolution of TPMT*3 alleles from *3C to *3A, with increasing number of B motifs in the VNTR region.
Graphical abstract
Due to high correlation of its activity with common genetic polymorphisms, TPMT exemplifies one of the most successful translations of pharmacogenomics into clinical practice [Citation1]. TPMT plays an important part in the deactivation pathway of thiopurine drugs by directing conversion of thiopurines, 6-mercaptopurine (6MP) and 6-thioguanine, to the methylated entities, 6-methyl-MP (6MMP) or 6-methylthioguanine, respectively [Citation2]. This S-adenosylmethionine-dependent methylation decreases levels of 6-thioguanine nucleotides, the active metabolites of thiopurine drugs, which are used in patients with acute lymphoblastic leukemia (ALL), transplantation procedures and autoimmune disorders [Citation3–5]. The population distribution of TPMT activity is trimodal [Citation2]. The majority of individuals (89%) have normal TPMT activity, around 11% of population have decreased activity and around 0.3% bears completely deficient enzyme [Citation6]. Often, the decrease in TPMT activity is due to SNPs in the TPMT gene, among which TPMT*3A, TPMT*3C and TPMT*2 are the most common [Citation7]. Individual genetic variability of TPMT is mirrored in levels of 6-thioguanine nucleotides. When patients with lower enzyme activity are treated with standard thiopurine doses, they exhibit a lower relapse rate, but are more prone to severe hematologic side effects [Citation3,Citation8].
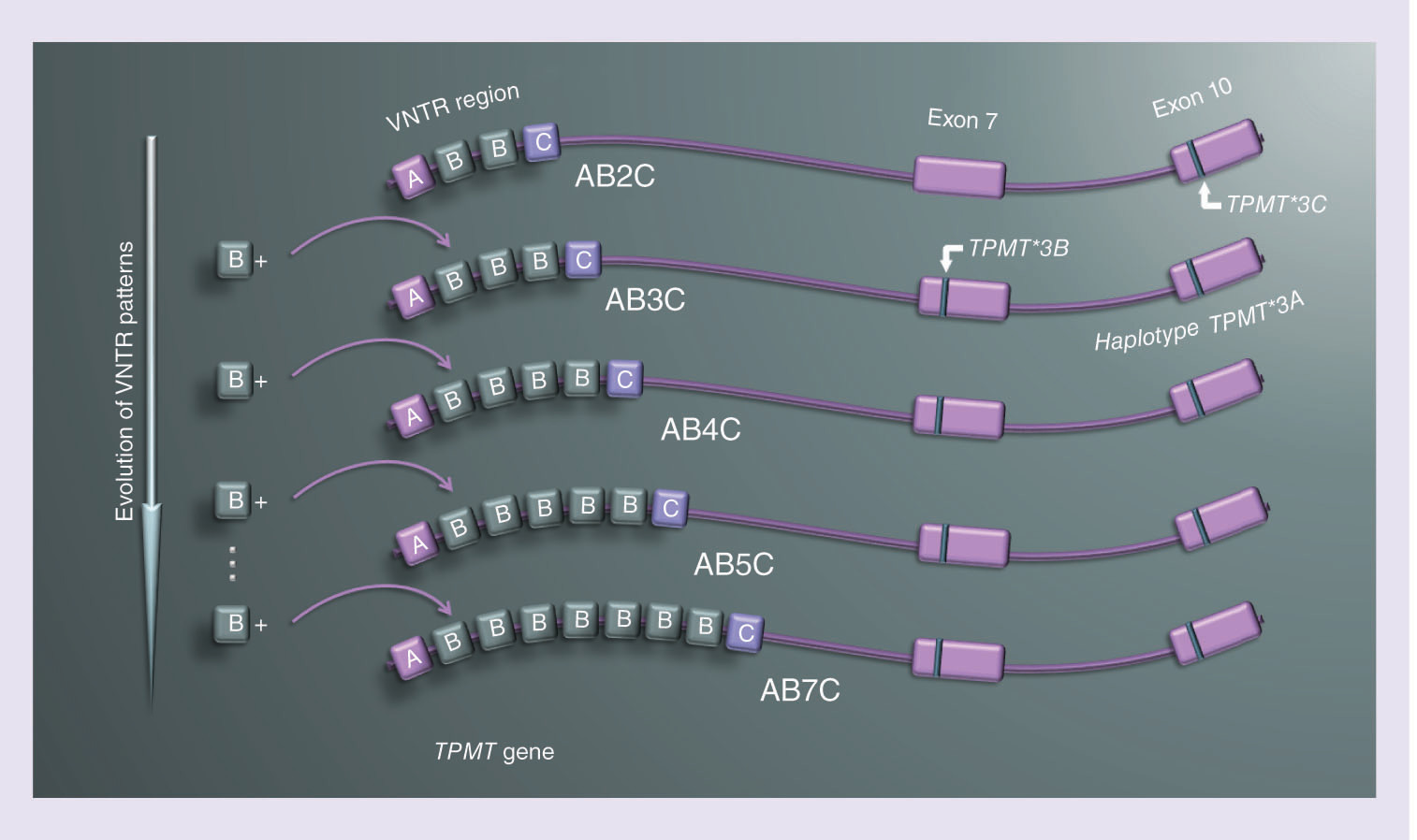
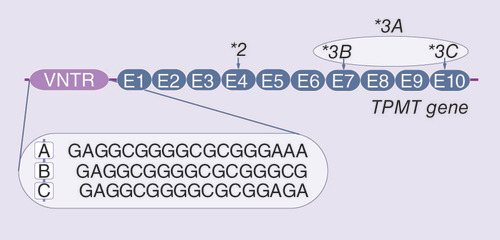
In addition to the variability caused by SNPs in the coding region of TPMT, its G/C rich promoter is variable as well, containing DNA motifs in the form of a variable number of tandem repeats (VNTR) [Citation9]. This repetitive DNA region consists of three motifs in its internal structure, named A, B and C () [Citation10]. Reported human VNTR alleles are made up of 1–7 repeats of motif A, 1–6 repeats of motif B and a single motif C. The motifs always appear in the same order: AmBnC [Citation10]. The overall length of repeats ranges from three to nine motifs, which classifies VNTR alleles in the TPMT promoter as minisatellites [Citation11]. The evolutionary mechanism, that best describes the formation of novel VNTR alleles in the TPMT promoter, is the stepwise mutation model (SMM) [Citation12,Citation13]. The model assumes gradual gaining (or losing) of one single motif at the time, explaining the phylogenetic relationships between VNTR alleles [Citation13].
The sequence of A, B and C VNTR motifs and location of *3 polymorphisms in the coding region of TPMT are shown.
Early studies attempted to determine the influence of VNTRs in the TPMT promoter on TPMT enzyme activity. Lower activity of the enzyme was associated with VNTR genotypes containing more than five repeats in total [Citation9,Citation14,Citation15]. However, several of these studies did not take into account the structure of the VNTR region (i.e., the exact number of A and B motifs), or did not label the alleles consistently, preventing exact conclusions. Recently, the number of A motifs in VNTR has been negatively correlated with the transcription rate of TPMT [Citation16,Citation17]. The results have been replicated in a clinical trial on patients treated with thiopurines [Citation18], suggesting the importance of precise characterization of VNTR patterns in terms of the number of A and B motifs for clinical applications.
The function and the mechanism of action of the VNTR region remain unknown. In this study, we investigated the association of VNTR architecture with TPMT activity and its connection to common clinically relevant genetic polymorphisms in the TPMT gene. We sequenced the TPMT promoter, genotyped TPMT for TPMT*2 and TPMT*3 alleles and measured TPMT activity in healthy Slovenian volunteers. Results were validated in a cohort enriched for variant TPMT*3 alleles, comprising nonrandomly selected Italian patients with ALL or inflammatory bowel disease (IBD) and lymphoblastoid cell lines (LCL) from healthy Israeli and Estonian donors.
Materials & methods
Blood samples
Venous blood (3–5 ml) from 132 unrelated healthy Caucasian volunteers from Slovenia [Citation6] was used for TPMT genotyping. TPMT activity was measured in the red blood cells (RBC) of 29, randomly chosen, Slovene individuals [Citation6]. Since the frequency of TPMT*3 alleles in the Slovenian cohort was too low to properly perform the linkage disequilibrium (LD) analysis, randomly selected Slovenian cohort was complemented by the samples of individuals previously determined to be carriers of TPMT*3 alleles. For VNTR analysis 24 such samples were nonrandomly selected from Italian ALL or IBD patients [Citation19,Citation20].
Lymphoblastoid cell lines
In order to further enrich the study cohort with TPMT*3 alleles, we conducted experiments on 41 LCLs of consenting healthy adults, 30 of which were obtained from the National Laboratory for the Genetics of Israeli populations, Tel-Aviv University, Israel [Citation21]. LCLs from the National Laboratory for the Genetics of Israeli populations were generated from the lymphocytes of 30 healthy donors – 27 Ashkenazi individuals, two Palestinian individuals and one Ethiopian individual as previously described [Citation21]. The other 11 LCLs were prepared from the lymphocytes of selected Caucasian subjects from the Estonian Biobank, Estonian Genome Center, University of Tartu [Citation22]. The lymphocytes of healthy adult donors were immortalized in vitro by Epstein–Barr virus. Cells were cultured in RPMI medium (Merck, Sigma-Aldrich, Darmstadt, Germany), supplemented with 10% fetal bovine serum (Thermo Fisher Scientific, Gibco, MA, USA), 4mM l-glutamine and antibiotics (100 U/ml penicillin; 100 μg/ml streptomycin; 250 ng/ml amphotericin B; all from Merck, Sigma-Aldrich) and kept under humidified conditions at 37°C and 5% CO2 [Citation21].
Extraction of DNA and genotyping
For blood samples, the extraction of DNA and determination of TPMT*2 and TPMT*3 alleles has been described previously [Citation6,Citation19,Citation20].
For LCLs, DNA was extracted from 2.5 × 106 cells according to the manufacturer's instructions using the QIAamp DNA Blood Mini Kit (Qiagene, Venlo, The Netherlands). TPMT*2 and TPMT*3 alleles were determined in samples with 10 ng of DNA using TaqMan SNP Genotyping Assay (Thermo Fisher Scientific; Applied Biosystems, CA, USA) on a Roche LightCycler 480 system, following the manufacturers’ instructions. SNPs and part numbers of all TaqMan SNP Genotyping Assays are listed in .
Table 1. The list of analyzed SNPs in the TPMT coding region, their rs numbers, nucleotide changes and TaqMan Assay IDs.
The promoter region of TPMT was genotyped for VNTR in the DNA isolated from both, blood samples and LCLs. The region of interest was amplified using primers F (5′-GTCACCCGAAATCCGCCAC-3′) and R (5′-CGTTCCCTTTCTCACCCGC-3′) with the PCR reaction mix containing 80–160 ng of DNA per sample. PCR reagent from FIREPol DNA Polymerase, PCR kit, with 30% Solution S for GC rich regions (i.e., G/C rich enhancer; all from Solis BioDyne, Tartu, Estonia) was used according to the manufacturer's instructions. The following thermal cycling conditions were applied: 15 min 95°C, 30× (1 min 95°C, 30 s 58°C, 1 min 72°C), 8 min 72°C and cooling. The presence of PCR fragments containing 622–730 base pairs was confirmed by gel electrophoresis composed of 2% agarose with 1× SYBR safe DNA gel stain (Thermo Fisher Scientific, Invitrogen). The separation of fragments was performed at 100 V and 400 A and lasted 20 min. PCR products were purified by QIAquick PCR purification Kit (Qiagene) according to the manufacturer's instructions. Samples were sequenced by McLab (CA, USA).
TPMT activity assay
TPMT activity in blood samples of 29, randomly chosen, healthy individuals was measured as previously described [Citation6].
Activity of the enzyme in LCLs was measured by the reversed-phase HPLC method [Citation6,Citation23]. Briefly, 10 × 106 cells were resuspended in 150 mM potassium-phosphate buffer, pH 7.4, and lysed by sonication. An aliquot of all samples was taken from the lysates for determination of proteins. The amount of produced 6MMP was measured in the rest of the cell lysate after 2-h incubation with 50 μM S-adenosylmethionine and 2.25 mM 6MP on 37°C. Total protein concentration was determined using the BioRad Protein Assay Kit (Bio-Rad, CA, USA), in accordance with the manufacturer's instructions. The activity was calculated as the amount of 6MMP produced in an hour per mg of protein. The mean TPMT activity for each individual was obtained from at least three independent experiments.
Statistical methods
Sequences were determined using Finch TV software (Geospiza, Inc., WA, USA; http://www.geospiza.com).
LD: since SNPs and VNTRs were determined separately, we first performed an analysis to estimate the haplotypes using PHASE platform for Windows 2.1.1 [Citation24,Citation25], which allowed for preservation of VNTR architecture in its original form. Motifs A and B were assigned as two separate alleles in the immediate vicinity of one another (1 bp). Standardized LD (D′) was then calculated from the estimated haplotype frequencies individually for each VNTR allele in combination with each SNP (*1, *3C, *3A [*3B]) (). The target VNTR allele was compared against a pool of other VNTR alleles. VNTR minisatellites have a significantly higher recombination rate than SNPs. Thus, SNPs can persist in all VNTR alleles formed in subsequent evolutionary steps. When addressing LD between VNTR patterns and TPMT*3A allele, we therefore merged haplotype information of samples containing VNTR alleles with ABnC, n >2 and compared them against a pool of other VNTR alleles. The significance of calculated LD was then determined by χ2 test.
Table 2. Frequency of TPMT*3 genotypes in a cohort of randomly selected healthy Slovenian individuals and in the cohort enriched for TPMT*3 alleles.
Table 3. Allele frequencies of variable number of tandem repeats in the present and previous studies.
Table 4. Linkage disequilibrium for all combinations of variable number of tandem repeats and SNPs in TPMT, obtained from estimated haplotype combinations.
It is noteworthy that the analysis of the VNTR region and SNP genotype analyses were performed separately by two researchers in a double-blind manner, to avoid bias. Neither of the researchers knew results on the genotype of the regions prior to the conclusion of the data analysis.
Results
Frequency of TPMT*3 & VNTR alleles
To evaluate the genetic background of TPMT in each individual, we first genotyped 132 randomly selected, healthy Slovenian blood donors for polymorphisms in coding regions and promoter of TPMT. The allele frequencies of TPMT*3A and TPMT*3C resembled those in other Caucasian populations. However, no TPMT*3A and TPMT*3C homozygous individuals were detected (). Furthermore, no TPMT*2 alleles were found in the Slovenian cohort. We determined 14 different VNTR alleles generating 24 different VNTR genotypes in this cohort. In accordance with previous studies, the most frequent allele was A2BC (43.1%), followed by A2B2C (37.9%) and A2B3C (8.5%) ( & ). All other alleles were represented at frequencies of less than 2% in the population. The most common genotypes were, as expected, A2BC/A2B2C (37.3%), A2BC/A2BC (17.9%) and A2B2C/A2B2C (14.2%). Interestingly, the VNTR allele AB2C was present only in the individual with allele TPMT*3C. Moreover, all TPMT*3A individuals carried one of the following VNTR alleles: AB3C, AB4C, AB5C and AB7C. None of these VNTR alleles (ABnC, n ≥2) were present in wild-type individuals. To our knowledge, the VNTR allele AB7C was observed for the first time.
†Nonrandomly selected samples: lymphoblastoid cell lines from Estonian and Israeli donors (n = 41) and blood samples from Italian inflammatory bowel disease or acute lymphoblastic leukemia patients (n = 24).
‡Variable number of tandem repeats (VNTR) alleles with frequency lower than 2%: AB4C (1.89%), AB5C and A5BC (1.52%), A4BC and A6BC (0.76%), ABC, AB2C, AB3C, AB7C, A7BC and A3B2C (0.38%).
§VNTR alleles with frequency lower than 2%: A6BC and ABC (1.54%), A4BC and A2B5C (0.77%).
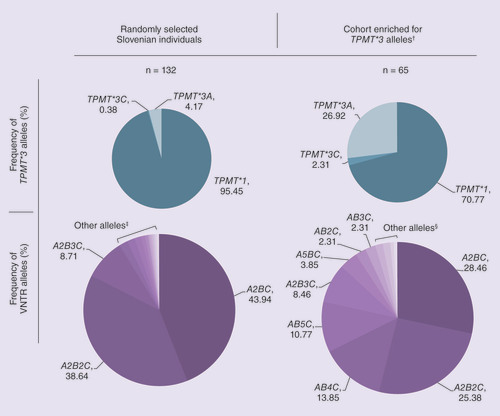
As the frequency of TPMT*3 alleles in the random population is too low to convincingly confirm the association between TPMT*3 and VNTR alleles, we next selected additional samples from individuals previously determined to carry TPMT*3 allele(s). Additionally, a few samples of wild-type TPMT individuals were added. These samples originated from IBD or ALL patients (n = 24) and from LCLs obtained from Estonian and Israeli donors (n = 41). By combining samples from LCLs, IBD and ALL patients, we intentionally created a cohort with elevated frequency of TPMT*3 alleles ( & ). DNA was extracted from 2 TPMT*1/*3C heterozygous, 32 TPMT*1/*3A heterozygous, one homozygous and one mixed heterozygous (*3A/*3A and *3A/*3C), as well as 29 wild-type individuals, and TPMT*3 genotype was reconfirmed (). The sequencing of the TPMT promoter region determined 12 structurally different VNTR alleles in this TPMT*3 enriched cohort ().
In all three cohorts combined (healthy Slovenian, Italian ALL and IBD patients and LCLs), AB3C, AB4C, AB5C and AB7C patterns were detected only in individuals with at least one TPMT*3A allele. Furthermore, the individual with genotype *3A/*3A was also AB4C/AB4C homozygous at the VNTR locus. These results suggest that TPMT*3A allele is in linkage with ABnC (n ≥3) VNTR patterns and corroborate results from Slovenian population study. In all three cohorts, AB2C was detected in individuals with *1/*3C and *3A/*3C genotypes. The latter had AB4C/AB2C genotype at the VNTR locus. Since AB4C associates with the TPMT*3A allele, the data further suggest that the TPMT*3C allele corresponds to AB2C VNTR motif. The high percentage of ABnC alleles in the cohort enriched for TPMT*3 variants () confirmed the association between VNTR motifs AB2C and ABnC (n ≥3) and TPMT*3C and TPMT*3A alleles, respectively.
Evolutionary linkage of VNTR & TPMT*3 polymorphisms
To statistically evaluate the correlation between TPMT*3 and VNTR alleles, we performed an analysis of LD, which further confirmed our hypothesis. Genotype data of merged samples from the Slovenian cohort, LCLs, ALL and IBD patients were used to estimate the haplotypes, and calculate the LD and its level of significance (). VNTR polymorphism AB2C was in statistically significant complete LD with TPMT*3C polymorphism (D′ = 1, r2 = 1; p < 0.0001), and pooled polymorphisms AB3C, AB4C, AB5C and AB7C were in complete LD with TPMT*3A polymorphism (D′ = 1, r2 = 1; p < 0.0001). Other VNTR patterns were in statistically significant LD with wild-type (TPMT*1) allele ().
All TPMT*3 alleles occur with VNTR pattern ABnC, where n is at least two. Therefore, we investigated the evolutionary basis for such an association. TPMT*3A allele is a haplotype that contains TPMT*3C and TPMT*3B SNPs in a coding region of TPMT () [Citation27]. TPMT*3C allele is quite common, especially in populations of African origin [Citation28,Citation29]. Meanwhile, TPMT*3B allele almost never occurs separately from TPMT*3C [Citation30,Citation31]. In all analyzed samples, TPMT*3C allele was present together with VNTR polymorphisms ABnC with two B motifs (n = 2) and TPMT*3A allele co-existed with the same VNTR patterns that contained three or more B motifs (n >2). Assuming a stepwise mutation model as an evolutionary description of VNTR formation and their phylogenetic evaluation [Citation12], we hypothesize that TPMT*3C variation occurred concurrently with the AB2C polymorphism (). In further evolutionary genetic changes, an additional B motif in the VNTR region was gained. At that time, the TPMT*3B mutation appeared, creating the TPMT*3A haplotype, suggesting that the TPMT*3B variation occurred sequentially after TPMT*3C and not in parallel or separately from it. Since the mutation rate of VNTR polymorphisms is significantly higher compared with the mutation rate of variations resulting in SNPs, TPMT*3A was retained, while the VNTR region gained additional B motifs, generating several haplotypes with TPMT*3A and VNTR ABnC polymorphisms ().
□ signifies other possible variable number of tandem repeats alleles, found in lymphoblastoid cell lines or previously reported by other studies [Citation12,Citation16].
![Figure 3. Phylogenetic associations between different alleles of variable number of tandem repeats in TPMT and their frequency distribution in the present set of Slovenian individuals, designated by the diameter of circles.□ signifies other possible variable number of tandem repeats alleles, found in lymphoblastoid cell lines or previously reported by other studies [Citation12,Citation16].](/cms/asset/9e1a42da-6436-4d15-9fd9-d8cb98f3c93c/ipgs_a_12348625_f0003.jpg)
Influence of TPMT*3 & VNTR alleles on the TPMT activity
To evaluate whether VNTR polymorphisms affect TPMT activity, we measured the basal activity of the enzyme in 29 of 132 healthy Slovenian volunteers and in 38 of 41 LCLs. Median TPMT activity of Slovenian donors with wild-type TPMT genotype (*1/*1) was 16.6 pmol 6MMP/h/107 RBC, whereas the median activity of heterozygous (*1/*3) individuals was 10.0 pmol 6MMP/h/107 RBC (A). Due to the low frequency of TPMT*3 alleles in the randomly selected population, the size of the Slovenian cohort was too small to allow the statistical analysis of TPMT activity distribution (A). However, the activity of two heterozygous individuals was in the range of previously reported intermediate TPMT activities in the Slovenian and other populations [Citation6,Citation32,Citation33]. The broad distribution of activities in individuals with wild-type TPMT is also in accordance with several previously published studies [Citation1,Citation34]. TPMT activity, measured in LCL lysates, significantly correlated with the number of TPMT*3 alleles (p < 0.0001; B). LCLs with wild-type TPMT (*1/*1) had the highest activity (median 137.4 pmol 6MMP/h/mg prot.), heterozygous LCLs (*1/*3) an intermediate level of activity (median 56.2 pmol 6MMP/h/mg prot.) and variant homozygotes (*3/*3) had the lowest TPMT activity (5.73 pmol 6MMP/h/mg prot.).
(A) Distribution of TPMT activity according to TPMT*3 genotype in Slovenian individuals. (B) Classification of TPMT activity based on TPMT genotype in lymphoblastoid cell lines (LCLs). Groups of LCLs with different TPMT*3 genotype significantly differ in TPMT activity (p = 0.0001). (C) and (D) TPMT activity of healthy Slovenian individuals and LCLs, respectively, in relation to the number of repeats in the VNTR pattern. TPMT activity trend with the total number of repeats is represented by ○, solid line, while the trend of only wild-type (*1/*1) individuals and LCLs is shown by □, dashed line. (E) Distribution of TPMT activity of LCLs across the groups with different VNTR patterns. LCLs carrying at least one TPMT*3 allele are shown in darker purple. Pooled subgroups of variable number of tandem repeats motifs (i.e., AmBC, ABnC and A2BnC) exhibited statistically significant differences in TPMT activity (p = 0.0001). Results are presented as median ± interquartile difference.
*p = 0.0001.
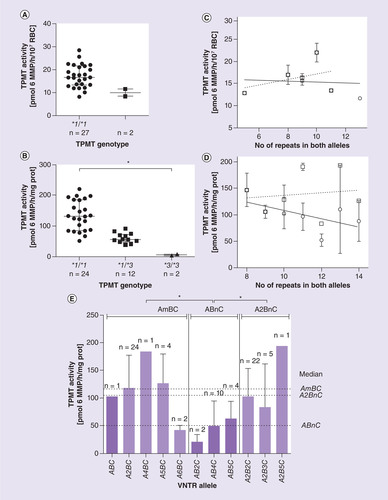
In accordance with previous studies [Citation9,Citation14], we observed a decrease in median TPMT activity with an increase in the number of repeats in both alleles, irrespective of the number of A and B motifs (C & D, solid line). The same trend of decreasing TPMT activity with the number of VNTR motifs was absent in only wild-type (*1/*1) individuals and LCLs (C & D, dashed line). In addition, we noticed a trend of fewer LCLs with *1/*1 genotype within the groups with more than ten tandem repeats, suggesting that individuals with variant TPMT*3 allele also have longer VNTR region. Not only the total number of tandem repeats, but also their VNTR patterns (i.e., number of A and B motifs) were associated with TPMT activity (E). ABnC patterns had significantly lower enzyme activity compared with A2BnC (p < 0.0001) and AmBC (p < 0.0001) patterns (E).
Discussion
The present study finds that there is a LD between VNTR patterns with ABnC structure (n ≥2) and TPMT*3 alleles. Specifically, motif AB2C is associated with TPMT*3C and ABnC (n ≥3) motifs with TPMT*3A allele. Furthermore, we identified a previously unreported VNTR pattern AB7C. The individual with this pattern is a carrier of TPMT*3A allele. Further functional studies for this individual (e.g., the analysis of TPMT activity) could not have been performed, as we have not been able to obtain the red blood cell sample from this individual.
Our data are strongly supported by other studies. The most demonstrable linkage disequilibria observed in previous studies were between TPMT*3A and AB4C (*6b), and between TPMT*3C and AB2C (*4b) [Citation26]. In that study all individual subjects (who originated from Pakistan and India) had only the AB2C VNTR pattern: no other ABnC polymorphisms were detected [Citation26]. This is in accordance with the distribution of TPMT*3 alleles in the ethnic groups of Asia [Citation35]. In another study, which assessed a Mozambican population, VNTR motifs AB4C (*V6b) and AB5C (*V7b) were absent, while AB2C motif was present; in this population TPMT*3C is the prevalent allele and TPMT*3A is rare [Citation12]. This is in line with our results and in accordance with the distribution of these variants in populations of African and Asian origin [Citation12]. Furthermore, VNTR*6, which could represent either AB4C, A4BC or A2B3C motifs, has been linked to the TPMT*3A polymorphism in early studies of VNTR alleles in TPMT [Citation9,Citation12]. Unfortunately, the information on qualitative structure of VNTR polymorphisms was lacking at that time, making it impossible to determine which of the three VNTR patterns was in linkage with TPMT*3A allele. Based on these findings, we can predict that African population would have extremely low frequencies of ABnC, n ≥3, alleles but higher frequency of AB2C allele, compared with the Caucasian population. Populations of Asian origin have a low frequency of TPMT*3C and an even lower frequency of TPMT*3A [Citation35,Citation36]. Therefore, AB2C with similar frequency to the Caucasian population and low frequency of ABnC, n ≥3, would be expected in these populations.
To compare our results with those in 1000 Genomes Project, we endeavored to analyze the data from the database [Citation37], using NCBI Genome Browser for GRCh37.p13 [Citation38]. The data confirmed population genetics and frequency distribution of TPMT*3C and TPMT*3A alleles, and complete LD between TPMT*3C and TPMT*3B alleles. Individuals with TPMT*3 alleles were further analyzed for the VNTR pattern. The coverage of this region, surrounding nucleotide 18,155,419 (Chr 6), was very low or nonexistent, which in the majority of cases made it impossible to identify these long and complex VNTR polymorphisms lying within a G/C-rich region in the TPMT promoter. Nevertheless, in approximately 20% of TPMT*3 individuals the VNTR pattern in the TPMT promoter could be partially predicted with the sequence pointing toward AB- and not AA- pattern, which is in line with our study, where patterns with single A and several B VNTR motifs occur in individuals carrying TPMT*3 allele. Despite the great advances of the next-generation sequencing, it has been previously reported, that genome assembly for highly G/C-rich regions is challenging and results in reduced or no coverage [Citation39]. Alternative approaches in sequencing complete VNTR assemblies such as single molecule real time sequencing have the potential to facilitate future high throughput studies of such polymorphisms [Citation40].
Conclusion
We report a novel VNTR pattern AB7C, which was observed in a TPMT*1/*3A individual. Furthermore, our findings provide novel insights into the concurrent development of VNTR and TPMT*3 alleles. We confirmed that TPMT*3 alleles correlate with TPMT activity. However, we could not conclude what influence (if any) VNTR polymorphisms have on the activity of the enzyme, since patterns with ABnC, n ≥2, are in significant LD with TPMT*3 alleles. Our study suggests that VNTR pattern AB2C is associated with TPMT*3C, while patterns ABnC, n ≥3, are associated with TPMT*3A allele, suggesting these VNTR sequences as indirect pharmacogenomic markers.
Future perspective
Based on the present results, it is possible that the VNTR region has no influence on the TPMT activity and that the previously observed association is the sole consequence of the linkage between ABnC, n ≥2, VNTR motifs and TPMT*3 alleles. Conversely, the same results could indicate the joint influence of the VNTR region and TPMT*3 alleles on the enzyme activity. Interesting insights in function of TPMT promoter region have been previously reported in terms of TPMT expression and binding of transcription factors [Citation10,Citation16,Citation41]; however, the role of VNTR in the promoter region of TPMT remains unresolved. Further functional in vitro studies are necessary to decipher the possible influence of the VNTR region on TPMT activity.
Based on its sequence, the B motif is strongly indicated as predicted binding site for transcription factor Sp1, which is a recognized regulator of TPMT transcription [Citation16,Citation41]. At the same time, sequence of motif B carries determinants that enable formation of G-quadruplexes [Citation42]. Even though their formation in promoter of TPMT has not been investigated, G-quadruplexes can be envisaged as regulators of VNTR stability or an element that directs transcription of the gene. In light of our findings, future studies of such secondary structures would be extremely important, since multiple B motifs are present in the promoter regions of TPMT*3 alleles. They might be even assumed as regulatory domains counteracting TPMT-activity-lowering SNPs in coding region of the TPMT.
We report a novel pattern of variable number of tandem repeats (VNTR) in the TPMT promoter: AB7C.
Only TPMT*3A individuals carried one of the following VNTR alleles: AB3C, AB4C, AB5C and AB7C.
Only TPMT*3C individuals carried AB2C allele.
VNTR patterns with single A and multiple B motifs are associated with TPMT*3 alleles. VNTR patterns with three or more B motifs developed concurrently with the TPMT*3B SNP from pattern AB2C, which coexisted with the TPMT*3C allele.
Author's contributions
D Urbančič, I Mlinarič-Raščan and N Karas Kuželički designed research. D Urbančič and A Šmid performed research. D Urbančič analyzed data. I Mlinarič-Raščan, G Stocco and G Decorti contributed reagents/clinical samples/analysis tools. D Urbančič, A Šmid and N Karas Kuželički wrote the paper.
Ethical disclosure
The authors state that, ‘the study has been approved by Local and National Medical Ethics Committees and has followed the principles outlined in the Declaration of Helsinki for all human experimental investigations. Written informed consent has been obtained from all participating subjects. The experiments comply with the current laws of the Republic of Slovenia and the Italian Republic’.
Acknowledgements
The authors wish to thank all participants who entered the study cohorts and clinical coordinators, M Rabusin and M Bramuzzo from the Institute for Maternal and Child Health IRCCS and BG Trieste for overseeing the recruitment of ALL and IBD patients. We would like to thank D Gurwitz from the Department of Human Molecular Genetics and Biochemistry, Sackler School of Medicine, Tel Aviv and A Metspalu from The Estonian Genome Center, University of Tartu for providing in vitro LCL model. We thank R Franca from the Institute for Maternal and Child Health IRCCS Burlo Garofolo Trieste for technical assistance and coordinating ALL repository, and M Lucafò from the Department of Medical, Surgical and Health Sciences, University of Trieste for coordinating IBD repository. The authors thank P Ferkov from the Faculty of Pharmacy, University of Ljubljana, D Selvestrel and O Montecchini from the Developmental and Reproductive Sciences, University of Trieste for technical assistance, and M Milek for providing useful information on TPMT in Slovene population. We would like to thank TJ Bevec for the proof reading of this manuscript.
Financial & competing interests disclosure
This work was supported by the Slovenian Research Agency (J3-6792, P1-0208, MR34512). The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed.
No writing assistance was utilized in the production of this manuscript.
Additional information
Funding
References
- Lennard L , CartwrightCS , WadeR , VoraA . Thiopurine methyltransferase and treatment outcome in the UK acute lymphoblastic leukaemia trial ALL2003 . Br. J. Haematol.170 ( 4 ), 550 – 558 ( 2015 ).
- Weinshilboum RM , SladekSL . Mercaptopurine pharmacogenetics: monogenic inheritance of erythrocyte thiopurine methyltransferase activity . Am. J. Hum. Genet.32 ( 5 ), 651 – 662 ( 1980 ).
- Lennard L , CartwrightCS , WadeR , VoraA . Thiopurine dose intensity and treatment outcome in childhood lymphoblastic leukaemia: the influence of thiopurine methyltransferase pharmacogenetics . Br. J. Haematol.169 ( 2 ), 228 – 240 ( 2015 ).
- Lee M-N , KangB , ChoiSYet al. Relationship between azathioprine dosage, 6-thioguanine nucleotide levels, and therapeutic response in pediatric patients with IBD treated with azathioprine . Inflamm. Bowel Dis.21 ( 5 ), 1054 – 1062 ( 2015 ).
- Liang JJ , GeskeJR , BoilsonBAet al. TPMT genetic variants are associated with increased rejection with azathioprine use in heart transplantation . Pharmacogenet. Genomics23 ( 12 ), 658 – 665 ( 2013 ).
- Milek M , MurnJ , JaksicZ , Lukac BajaloJ , JazbecJ , Mlinaric RascanI . Thiopurine S-methyltransferase pharmacogenetics: genotype to phenotype correlation in the Slovenian population . Pharmacology77 ( 3 ), 105 – 114 ( 2006 ).
- Tamm R , MägiR , TremmelRet al. Polymorphic variation in TPMT is the principal determinant of TPMT phenotype: a meta-analysis of three genome-wide association studies . Clin. Pharmacol. Ther.101 ( 5 ), 684 – 695 ( 2017 ).
- Lee M-N , KangB , ChoiSYet al. Impact of genetic polymorphisms on 6-thioguanine nucleotide levels and toxicity in pediatric patients with IBD treated with azathioprine . Inflamm. Bowel Dis.21 ( 12 ), 2897 – 2908 ( 2015 ).
- Spire-Vayron de la Moureyre C , DebuysereH , MastainBet al. Genotypic and phenotypic analysis of the polymorphic thiopurine S-methyltransferase gene (TPMT) in a European population . Br. J. Pharmacol.125 ( 4 ), 879 – 887 ( 1998 ).
- Spire-Vayron de la Moureyre C , DebuysèreH , FazioFet al. Characterization of a variable number tandem repeat region in the thiopurine S-methyltransferase gene promoter . Pharmacogenetics9 ( 2 ), 189 – 198 ( 1999 ).
- Vergnaud G , DenoeudF . Minisatellites: mutability and genome architecture . Genome Res.10 ( 7 ), 899 – 907 ( 2000 ).
- Alves S , AmorimA , PrataMJ . Evolution of a VNTR located within the promoter region of the thiopurine methyltransferase gene: inferences from population and sequence data . Hum. Genet.111 ( 2 ), 172 – 178 ( 2002 ).
- Kimura M , OhtaT . Stepwise mutation model and distribution of allelic frequencies in a finite population . Proc. Natl Acad. Sci. USA75 ( 6 ), 2868 – 2872 ( 1978 ).
- Yan L , ZhangS , EiffBet al. Thiopurine methyltransferase polymorphic tandem repeat: genotype–phenotype correlation analysis . Clin. Pharmacol. Ther.68 ( 2 ), 210 – 219 ( 2000 ).
- Alves S , AmorimA , FerreiraF , PrataMJ . Influence of the variable number of tandem repeats located in the promoter region of the thiopurine methyltransferase gene on enzymatic activity . Clin. Pharmacol. Ther.70 ( 2 ), 165 – 174 ( 2001 ).
- Zukic B , RadmilovicM , StojiljkovicMet al. Functional analysis of the role of the TPMT gene promoter VNTR polymorphism in TPMT gene transcription . Pharmacogenomics11 ( 4 ), 547 – 557 ( 2010 ).
- Kotur N , StankovicB , KasselaKet al. 6-mercaptopurine influences TPMT gene transcription in a TPMT gene promoter variable number of tandem repeats-dependent manner . Pharmacogenomics13 ( 3 ), 283 – 295 ( 2012 ).
- Kotur N , DokmanovicL , JanicDet al. TPMT gene expression is increased during maintenance therapy in childhood acute lymphoblastic leukemia patients in a TPMT gene promoter variable number of tandem repeat-dependent manner . Pharmacogenomics16 ( 15 ), 1701 – 1712 ( 2015 ).
- Stocco G , MartelossiS , ArrigoSet al. Multicentric case–control study on azathioprine dose and pharmacokinetics in early-onset pediatric inflammatory bowel disease . Inflamm. Bowel Dis.23 ( 4 ), 628 – 634 ( 2017 ).
- Stocco G , MartelossiS , BarabinoAet al. Glutathione-S-transferase genotypes and the adverse effects of azathioprine in young patients with inflammatory bowel disease . Inflamm. Bowel Dis.13 ( 1 ), 57 – 64 ( 2007 ).
- Morag A , KirchheinerJ , RehaviM , GurwitzD . Human lymphoblastoid cell line panels: novel tools for assessing shared drug pathways . Pharmacogenomics11 ( 3 ), 327 – 340 ( 2010 ).
- Leitsalu L , HallerT , EskoTet al. Cohort profile: Estonian Biobank of the Estonian Genome Center, University of Tartu . Int. J. Epidemiol.44 ( 4 ), 1137 – 1147 ( 2015 ).
- Keizer-Garritsen JJ , BrouwerC , LambooyLHJet al. Measurement of thiopurine S-methyltransferase activity in human blood samples based on high-performance liquid chromatography: reference values in erythrocytes from children . Ann. Clin. Biochem.40 ( Pt 1 ), 86 – 93 ( 2003 ).
- Stephens M , SmithNJ , DonnellyP . A new statistical method for haplotype reconstruction from population data . Am. J. Hum. Genet.68 ( 4 ), 978 – 989 ( 2001 ).
- Stephens M , ScheetP . Accounting for decay of linkage disequilibrium in haplotype inference and missing-data imputation . Am. J. Hum. Genet.76 ( 3 ), 449 – 462 ( 2005 ).
- Marinaki AM , ArenasM , KhanZHet al. Genetic determinants of the thiopurine methyltransferase intermediate activity phenotype in British–Asians and Caucasians . Pharmacogenetics13 ( 2 ), 97 – 105 ( 2003 ).
- Szumlanski C , OtternessD , HerCet al. Thiopurine methyltransferase pharmacogenetics: human gene cloning and characterization of a common polymorphism . DNA Cell Biol.15 ( 1 ), 17 – 30 ( 1996 ).
- Hon YY , FessingMY , PuiCH , RellingMV , KrynetskiEY , EvansWE . Polymorphism of the thiopurine S-methyltransferase gene in African–Americans . Hum. Mol. Genet.8 ( 2 ), 371 – 376 ( 1999 ).
- Hiratsuka M , InoueT , OmoriF , AgatsumaY , MizugakiM . Genetic analysis of thiopurine methyltransferase polymorphism in a Japanese population . Mutat. Res.448 ( 1 ), 91 – 95 ( 2000 ).
- Rossino R , VincisC , AlvesSet al. Frequency of the thiopurine S-methyltransferase alleles in the ancient genetic population isolate of Sardinia . J. Clin. Pharm. Ther.31 ( 3 ), 283 – 287 ( 2006 ).
- Bahari A , HashemiM , BariZ , Moazeni-RoodiA , KaykhaeiM-A , NarouieB . Frequency of thiopurine S-methyltransferase (TPMT) alleles in southeast Iranian population . Nucleosides Nucleotides Nucleic Acids29 ( 3 ), 237 – 244 ( 2010 ).
- Karas-Kuželički N , ŠmidA , TammR , MetspaluA , Mlinarič-RaščanI . From pharmacogenetics to pharmacometabolomics: SAM modulates TPMT activity . Pharmacogenomics15 ( 11 ), 1437 – 1449 ( 2014 ).
- Chouchana L , NarjozC , RocheDet al. Interindividual variability in TPMT enzyme activity: 10 years of experience with thiopurine pharmacogenetics and therapeutic drug monitoring . Pharmacogenomics15 ( 6 ), 745 – 757 ( 2014 ).
- Hindorf U , AppellML . Genotyping should be considered the primary choice for pre-treatment evaluation of thiopurine methyltransferase function . J. Crohns Colitis6 ( 6 ), 655 – 659 ( 2012 ).
- Kubota T , ChibaK . Frequencies of thiopurine S-methyltransferase mutant alleles (TPMT*2, *3A, *3B and *3C) in 151 healthy Japanese subjects and the inheritance of TPMT*3C in the family of a propositus . Br. J. Clin. Pharmacol.51 ( 5 ), 475 – 477 ( 2001 ).
- Lu Y , KhamSK-Y , FooT-Cet al. Genotyping of eight polymorphic genes encoding drug-metabolizing enzymes and transporters using a customized oligonucleotide array . Anal. Biochem.360 ( 1 ), 105 – 113 ( 2007 ).
- 1000 Genomes Project Consortium ; AutonA , BrooksLDet al.A global reference for human genetic variation . Nature526 ( 7571 ), 68 – 74 ( 2015 ).
- Geer LY , Marchler-BauerA , GeerRCet al. The NCBI BioSystems database . Nucleic Acids Res.38 ( Database issue ), D492 – D496 ( 2010 ).
- Chen Y-C , LiuT , YuC-H , ChiangT-Y , HwangC-C . Effects of GC bias in next-generation-sequencing data on de novo genome assembly . PLoS ONE8 ( 4 ), e62856 ( 2013 ).
- Wenzel A , AltmuellerJ , EkiciABet al. Single molecule real time sequencing in ADTKD- MUC1 allows complete assembly of the VNTR and exact positioning of causative mutations . Sci. Rep.8 ( 1 ), 4170 ( 2018 ).
- Fessing MY , KrynetskiEY , ZambettiGP , EvansWE . Functional characterization of the human thiopurine S-methyltransferase (TPMT) gene promoter . Eur. J. Biochem.256 ( 3 ), 510 – 517 ( 1998 ).
- Todd AK , JohnstonM , NeidleS . Highly prevalent putative quadruplex sequence motifs in human DNA . Nucleic Acids Res.33 ( 9 ), 2901 – 2907 ( 2005 ).