
The potential of bendamustine, a purine-like alkylating agent, for treating blood cancers associated with the Epstein-Barr virus (EBV) led us to examine interactions between drug and virus. Bendamustine and other alkylating agents such as chlorambucil and melphalan disrupt latency. Bendamustine and rituximab, an anti-CD20 monoclonal antibody, can combine to increase lytic replication. We discuss how this ability to reactivate latent virus informs clinical strategies.
EBV infection is associated with human hematologic malignancies. This herpesvirus evades immune surveillance over the host’s lifetime by entering a quiescent phase in B-cells. Compared to lytic replication, during which ∼100 gene products amplify virus, latency restricts transcription to ∼1–10 genes [Citation1]. EBV adopts this expression program to immortalize B-cells in vitro and inhabit tumors of B-cell malignancies like a proportion of Burkitt, Hodgkin, post-transplantation, diffuse large B-cell, primary central nervous system, primary effusion, and plasmablastic lymphoma in vivo [Citation1,Citation2].
Bendamustine is a chemotherapy agent increasingly used to treat blood cancers. Recently rediscovered, this small molecule contains a mechlorethamine nitrogen mustard that generates DNA damage distinct from other alkylating drugs. Bendamustine combined with other drugs [Citation3,Citation4] effectively treats indolent non-Hodgkin lymphoma. Preliminary trials also show success against relapsed or refractory diffuse large B-cell lymphoma [Citation5,Citation6]. Similarly aggressive cancers are associated with EBV infection [Citation1,Citation2], so the potential of bendamustine treatment for EBV-associated malignancies led us to examine interactions between drug and virus.
Bendamustine and other alkylating chemotherapy drugs induce EBV reactivation in a cell culture model of non-Hodgkin lymphoma. MutuI cells [Citation7] were grown at 37 °C with 5% CO2 in RPMI-1640 containing 2 mM HEPES and 10% fetal bovine serum. We treated cells at a density of 0.5 × 106/mL with bendamustine (Sigma-Aldrich, St. Louis, MO or EMD Millipore, Billerica, MA). Reactivation was measured by staining for the immediate-early lytic transactivator BZLF1 using the paraformaldehyde-methanol method [Citation8] with a BZ1 antibody (Santa Cruz Biotechnology, Dallas, TX) and goat anti-mouse IgG-FITC (Santa Cruz Biotechnology, Dallas, TX). Flow cytometry was performed on a FACSCaliburDxP8 (BD Biosciences, San Jose, CA). Bendamustine evokes both a dose- and time-dependent response in that 2- or 3-day treatment with 10–100 μM bendamustine induces ∼10–30% reactivation []. Lytic induction also modestly occurs in EBV-infected Akata and Daudi cells, but not detectably in KemI, KemIII, and GM12878 cells (data not shown). Deep sequencing of the EBV transcriptome (Phan & Miranda, manuscript in preparation) reveals that bendamustine increases RNA levels genome-wide by ∼100-fold []. BHLF1 shares a promoter with the lytic origin of replication and its expression correlates with reactivation [Citation9]; BcLF1, a true late gene, requires viral DNA replication to encode a capsid protein [Citation10]. Bendamustine increases BHLF1 and BcLF1 expression as well as the entire lytic program. Curious if other alkylating agents have similar effects, we treated cells with either 10 μM chlorambucil (Sigma-Aldrich, St. Louis, MO) or 10 μM melphalan (Sigma-Aldrich, St. Louis, MO) for 3 days. Both drugs also reactivate EBV []. We can therefore add bendamustine and other alkylators to the list of DNA-damaging agents, such as cisplatin, 5-fluorouracil, gemcitabine, and doxorubicin [Citation11,Citation12], capable of disrupting EBV latency.
Figure 1. EBV reactivation induced by bendamustine and other alkylating agents. (A) Time- and dose-dependent response of EBV reactivation upon bendamustine treatment. BZLF1 expression in MutuI cells was measured by flow cytometry. Error bars represent the standard deviation of n = 4 replicates. (B) Deep sequencing of the EBV transcriptome in response to bendamustine treatment. The X axis denotes nucleotide position and the Y axis denotes the number of counts per million mapped reads. An inset of the top panel depicts low levels of transcription not visible on the scale used to compare conditions. Specific lytic genes are annotated. One representative experiment from two independent replicates is shown. (C) EBV reactivation upon chlorambucil and melphalan treatment. BZLF1 expression in MutuI cells was measured by flow cytometry. Error bars represent the standard deviation of n = 4 replicates. (D) Increased reactivation induced by bendamustine plus rituximab. BZLF1 expression in MutuI cells was measured by flow cytometry. Error bars represent the standard deviation of n = 4 replicates.
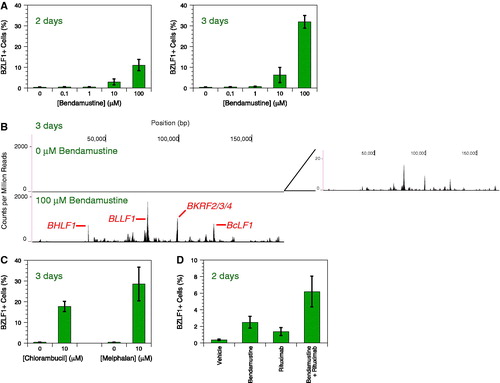
Because bendamustine plus rituximab, an anti-CD20 monoclonal antibody, effectively treats indolent non-Hodgkin lymphoma [Citation3,Citation4], we investigated this combination’s interaction with latent EBV Rituximab (Genentech, South San Francisco, CA) at 100 μg/mL rituximab and bendamustine at 10 μM independently induce EBV reactivation after 2 days, but combination generates even more []. Rituximab similarly enhances reactivation induced by dexamethasone [Citation13]. CD20 engagement combined with other latency disruptors may be a general means of enhancing induction.
The ability of bendamustine to reactivate latent EBV informs future clinical strategies. Taken pessimistically, our work warns that drug treatment may exacerbate risk of adverse effects caused by viremia in an already weakened immune system. Impairment of lymphocyte recovery by bendamustine [Citation14] particularly warrants heightened precautions. Personalized medicine that tailors chemotherapy regimens to EBV status of the tumor may suggest the preferred treatment. Taken optimistically, this study contends that bendamustine may be useful in lytic induction therapy, an alternative to standard cytotoxic treatment that attempts to leverage the presence of virus to target cancer cells. Latent EBV coerced into reactivation can specifically mark tumor cells for killing by cytopathic effects, epitope recognition by cytotoxic T lymphocytes, or small molecule drugs that target lytic proteins [Citation15]. Our transcriptome profiling reveals that bendamustine induces the lytic EBV expression program, including true late genes, which demonstrates that prerequisite proteins must have been translated. These early proteins comprise the targets for multiple cure strategies. Drugs like bendamustine and other alkylating agents, which are already efficacious antitumor agents, could form the foundation of effective lytic induction therapy combinations. In whatever way this knowledge is applied, future clinical work should recognize that bendamustine reactivates latent EBV.
GLAL-2015-0568-File003.pdf
Download PDF (1.2 MB)GLAL-2015-0568-File002.pdf
Download PDF (1.2 MB)Acknowledgements
We thank An T. Phan for optimizing the BZLF1 expression assay, Shannon C. Kenney (University of Wisconsin, Madison) for providing the MutuI cell line, and Marielle Cavrois and Marianne L. Gesner at the Gladstone Institutes Flow Cytometry Core for assisting with experiments. This publication was supported by the National Center for Advancing Translational Sciences, National Institutes of Health, through UCSF-CTSI Grant Number UL1 TR000004. This publication was made possible with help from the University of California San Francisco-Gladstone Institute of Virology & Immunology Center for AIDS Research (CFAR), an NIH-funded program (P30 AI027763).
Potential conflict of interest
Disclosure forms provided by the authors are available with the full text of this article at www.informahealthcare.com/lal.
References
- Young LS, Rickinson AB. Epstein-Barr virus: 40 years on. Nature Rev Cancer 2004; 4: 757–768.
- Carbone A, Cesarman E, Spina M, et al. HIV-associated lymphomas and gamma-herpesviruses. Blood 2009; 113: 1213–1224.
- Rummel MJ, Niederle N, Maschmeyer G, et al. Bendamustine plus rituximab versus CHOP plus rituximab as first-line treatment for patients with indolent and mantle-cell lymphomas: an open-label, multicentre, randomised, phase 3 non-inferiority trial. Lancet 2013; 381:1203–1210.
- Flinn IW, van der Jagt R, Kahl BS, et al. Randomized trial of bendamustine-rituximab or R-CHOP/R-CVP in first-line treatment of indolent NHL or MCL: the BRIGHT study. Blood 2014; 123: 2944–2952.
- Ohmachi K, Niitsu N, Uchida T, et al. Multicenter phase II study of bendamustine plus rituximab in patients with relapsed or refractory diffuse large B-cell lymphoma. J Clin Oncol 2013; 31: 2103–2109.
- Vacirca JL, Acs PI, Tabbara IA, et al. Bendamustine combined with rituximab for patients with relapsed or refractory diffuse large B cell lymphoma. Ann Hematol 2014; 93: 403–409.
- Gregory CD, Rowe M, Rickinson AB. Different Epstein-Barr virus-B cell interactions in phenotypically distinct clones of a Burkitt’s lymphoma cell line. J Gen Virol 1990; 71(Pt 7): 1481–1495.
- Imbert-Marcille BM, Coste-Burel M, Robillard N, et al. Sequential use of paraformaldehyde and methanol as optimal conditions for the direct quantification of ZEBRA and rta antigens by flow cytometry. Clin Diagnostic Lab Immunol 2000; 7: 206–211.
- Arvey A, Tempera I, Tsai K, et al. An atlas of the Epstein-Barr virus transcriptome and epigenome reveals host-virus regulatory interactions. Cell Host Microbe 2012; 12: 233–245.
- Chang PJ, Chang YS, Liu ST. Characterization of the BcLF1 promoter in Epstein-Barr virus. J Gen Virol 1998; 79(Pt 8):2003–2006.
- Feng WH, Israel B, Raab-Traub N, et al. Chemotherapy induces lytic EBV replication and confers ganciclovir susceptibility to EBV-positive epithelial cell tumors. Cancer Res 2002; 62: 1920–1926.
- Feng WH, Hong G, Delecluse HJ, et al. Lytic induction therapy for Epstein-Barr virus-positive B-cell lymphomas. J Virol 2004; 78: 1893–1902.
- Daibata M, Bandobashi K, Kuroda M, et al. Induction of lytic Epstein-Barr virus (EBV) infection by synergistic action of rituximab and dexamethasone renders EBV-positive lymphoma cells more susceptible to ganciclovir cytotoxicity in vitro and in vivo. J Virol 2005; 79: 5875–5879.
- Garcia Munoz R, Izquierdo-Gil A, Munoz A, et al. Lymphocyte recovery is impaired in patients with chronic lymphocytic leukemia and indolent non-Hodgkin lymphomas treated with bendamustine plus rituximab. Ann Hematol 2014; 93: 1879–1887.
- Israel BF, Kenney SC. Virally targeted therapies for EBV-associated malignancies. Oncogene 2003; 22: 5122–5130.