
Abstract
The present study was performed to determine the effects of different antioxidants on testicular histopathology and oxidative damage induced by cadmium (Cd) in rat testis and prostate. Twenty five rats were equally divided into five groups (n = 5/group). The control group was injected subcutaneously with saline while the Cd alone treated group received a subcutaneous injection of 0.2 mg/kg CdCl2. Other groups were treated with sulphoraphane (25 µg/rat), vitamin E (75 mg/kg), and Ficus Religiosa plant extract (100 mg/kg) orally along with subcutaneous injections of 0.2 mg/kg CdCl2 for fifteen days. Oxidative damage in the testicular and prostate tissues were assessed by the estimation of catalase (CAT), peroxidase (POD), superoxide dismutase (SOD), and glutathione reductase (GSR) activity. Lipid peroxidation (TBARS), protein estimation, and histomorphology were also assessed. Cadmium exposure caused a significant decrease in antioxidant enzymes like CAT, POD, SOD, GSR, protein concentrations, and a marked increase in TBARS activity in rat testis and prostate. Histological examination of adult male rat testes showed a disruption in the arrangement of seminiferous tubules along with a reduction in the number of germ cells, Leydig cells, tunica albuginea thickness, diameter of seminiferous tubules, and height of germinal epithelium. Co-treatment with vitamin E, sulphoraphane, and Ficus religiosa were found to be effective in reversing Cd induced toxicity, representing potential therapeutic options to protect the reproductive tissues from the detrimental effects of Cd toxicity.
Introduction
Humans are generally exposed to Cd toxicity in drinking water, smoke, and food. Unlike organic compounds, Cd is not biodegradable and has a long biological half-life. Persistent Cd poisoning can lead to osteoporosis, nephrotoxicity, and necrosis of the prostate and testes. It also causes cardiovascular disease, neurodegenerative diseases, renal failure, growth retardation, infertility, and embryotoxic effects [El-Shahat et al. 2009; Yu et al. Citation2007].
Cadmium exposure results in tissue toxicity, mainly by causing oxidative damage to the tissue. Oxidative stress is known to be an important element in almost half of the men with infertility and is seen as a key factor, causing oxidative damage by producing reactive oxygen species (ROS) [Tremellen Citation2008]. Cadmium is not a redox metal and adopts a single oxidized state generating superoxide anion radicals, H2O2, NO, and OH-radicals, which alter the system of antioxidant enzymes. It causes a disturbance of the defense system and induces high levels of lipid peroxidation (LPO) [Martin et al. Citation2001].
Testicular tissue is highly susceptible to Cd toxicity, as this may lead to alterations in steroid biosynthesis in the Leydig cells [Nandi et al. Citation2001]. Cadmium exposure leads to oxidative damage, which in turn causes cell death and necrosis within the tissue [Bonda et al. Citation2004; El-Demerdash et al. 2004; Sen-Gupta et al. Citation2004]. Similarly chronic toxic exposure to Cd produces significant histological changes in the ventral prostate [Martin et al. Citation2001]. It is assumed that Cd at a dose as low as 1–2 mg/kg body weight can cause histological changes in the testes of adult male rats without causing pathological changes in other organs [Prozialeck et al. Citation2007].
This damage to the tissue is prevented by enzymatic and non-enzymatic products, produced within the cells. Vitamin E is a well-known antioxidant which is lipid-soluble and may stop the generation of ROS in the oxidation of fats. It is the principal continuous inductor component of the enzyme complex protection system that can protect cells against oxidative stress induced damage [Azzi et al. Citation1993]. Sulforaphane is a phytochemical natural substance and belongs to the family of isothiocyanates, that has been shown to reduce diazinon and cisplatin induced oxidative damage in liver, kidney, and lungs of rats [Guerrero-Beltran et al. Citation2010; Harun et al. 2011]. Ficus religiosa is an antioxidant that reduces stress in the body [Arora et al. Citation1999] that is also used as a treatment for disorders of the endocrine system and the reproductive system [Panda Citation2000]. Vitamin E is a well known antioxidant that provides protection against Cd induced testicular damage in rats [Kini et al. Citation2009], however, little is known about the protective effects of the other antioxidants used in this study against metal intoxication in the reproductive tissues. With reference to the previous study of Jahan et al. [Citation2014], in which it was found that these antioxidants increase the concentration of Zn and daily sperm production, and reduces Cd deposition in testicular tissues, we extended our observations by determining the effect of these antioxidants on oxidative stress and histological changes induced by Cd within the testicular and prostate tissues of male rats.
Results
No significant difference was observed in left, right, and total testicular weight, length, and width of all treated groups as compared to the control group. Cadmium treatment caused a significant reduction in protein content (5.92 ± 0.17) in testicular tissues as compared to the control group (10.53 ± 0.39). Co-treatment with the antioxidants prevented this reduction in tissue protein content significantly (10.34 ± 0.53, 11.09 ± 0.60, 9.61 ± 0.20 in sulforaphane, vitamin E, and plant extract (PE) co-treated groups, respectively).
The concentration of the antioxidant enzymes, catalase (CAT), peroxidise (POD), and superoxide dismutase (SOD) in testicular tissues were found significantly lower in the Cd alone treated animals as compared to the control (). This reduction in antioxidant enzyme concentration was significantly protected in the antioxidant treated groups compared to the Cd alone treated group. In the case of CAT, no significant difference was found when the Cd alone treated group was compared to the Cd + sulforaphane treated group ().
Table 1. Specific activities of catalase, peroxidise, and superoxide dismutase in the testis and prostate of control and rats exposed for 15 days to cadmium (Cd) alone, Cd + sulforaphane, Cd + vitamin E, and Cd + plant extract.
In prostate tissue, Cd treatment resulted in a significant reduction in the concentration of CAT, POD, and SOD as compared to the control group. The concentration of CAT in the vitamin E and sulforaphane treated animals did not increase significantly when compared to the Cd-alone treated group. The PE treated group showed a significant increase in CAT concentration. The POD concentration was found to be significantly high in all antioxidant treated groups as compared to the Cd alone treated group, however, SOD levels were increased only in the sulforaphane treated group as compared to the Cd-alone treated group ().
A significant increase in the level of lipid peroxidation (TBARS) was found in the Cd alone treated group when compared to the control group. Lipid peroxidation levels were significantly lower in the animals co-treated with antioxidants (sulforaphane, vitamin E, and PE) (). In comparison, the concentration of glutathione (GSR) decreased significantly in the Cd alone treated group compared to the control group in both testicular and prostate tissues. In the Cd + sulforaphane treated group, the concentration of GSR was significantly significantly higher compared to the Cd alone treated group. The concentration of GSR was also significantly higher in the group treated with vitamin E and PE ().
Table 2. Thiobarbituric acid reactive substance and glutathione reductase activity in testis and prostate of control and rats exposed for 15 days to cadmium, cadmium + sulforaphane, cadmium + vitamin E, and cadmium + plant extract.
Histological results
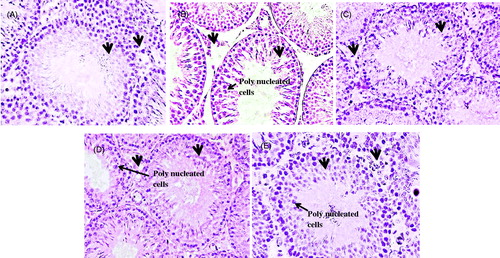
Microscopic evaluation of the control animals showed the typical features of tubular lumen, germinal epithelium, and interstitial space (). Treatment with Cd alone resulted in increased tubuler lumen, sloughing of the epithelial layer with scattered Sertoli cells, and degeneration of interstitial space (). The spermatids present in the lumen were scattered and reduced in number. Compared to the testis sections of the Cd treated animals, testicular sections of groups treated with Cd along with (sulforaphane, vitamin E, and PE) showed less histological change (). The effect of vitamin E was more prominent as compared to PE and sulforaphane.
Figure 1. Photomicrograph of seminiferous tubules in (A) Control; showing filled lumen with elongated spermatids, arranged spermatogonia, Sertoli cells, and Leydig cells with compact germinal epithelium (arrow) and small interstitial space (arrow) (B) Cd alone; showing tubules with empty lumen, sloughing of the epithelial layer, and polynucleated cells (arrow) with scattered Sertoli cells and degenerated interstitial space (arrow) (C) Cd + sulforaphane; showing round seminiferous tubule with elongated spermatids with no interstitial space degeneration (arrow) and less sloughing in epithelial layer of cells (arrow) (D) Cd + vitamin E; showing filled lumen and compact seminiferous tubules with less interstitial space and (E) Cd + PE treated group; with loose interstitial space (arrow) but with regular arranged cells in germinal epithelium and with elongated spermatids and least sloughing. Arrow heads indicating the loosening in the interstitial space and sloughing in the germinal epithelium, presence of polynucleated cells showing degeneration in the tubules. X40.
Morphometric results
Treating rats with Cd for 15 days caused a reduction in the thickness of the tunica albuginea. The thickness of the tunica albuginea was significantly smaller (48.27 ± 1.23) in the Cd alone treated animals as compared to the control (76.48 ± 2.41) (). Seminiferous epithelial height was decreased in comparison to that of the control. The mean seminiferous epithelial height was 222.30 ± 4.75 μm and 156.33 ± 2.47 μm in the controls and Cd groups, respectively (). Similarly seminiferous tubule diameter in Cd alone treated animals (67.9 ± 31.42) was significantly (p < 0.001) reduced than in the control (95.20 ± 2.80) (). Compared with Cd treated animals, those treated with Cd and plant extract (PE) showed an improvement in the mean tubular diameter and germ cell height. In animals receiving sulforaphane, vitamin E, and PE the tubular diameter and epithelial height of seminiferous tubules were normal and significantly high as compared to Cd alone treated animals ().
Table 3. Mean ± S.E.M tunica albugenia thickness, seminiferous tubule height and diameter (μm) in the treated and control animals.
Cell count
Compared to the control group there was a significant reduction in the mean number of spermatogonia, primary spermatocytes, secondary spermatocytes, and spermatids in the Cd alone treated animals group (). Co-treatment with vitamin E, sulforaphane, and PE resulted in an enumerative effect in the number of cells, when compared to Cd alone treated animals. However, no difference was noticed in the number of spermatids in the Cd alone treated animals and Cd plus PE treated animals ().
Table 4. Mean ± S.E.M number of different cell type in the seminiferous tubule in the treated and control animals.
Discussion
Exposure to Cd results in reproductive toxicity within the testicular tissue. These toxic effects include morphological changes resulting from the degeneration of cells under pathological conditions. In this experiment, Cd exposure resulted in the reduction of protein content and an increase in TBARS in both testis and prostate. These results are considered to echo the pathological changes within the tissue. Conversely, this reduction in protein content and increase in TBARS, were not prominent in the antioxidant treated groups after fifteen days of treatment. The biochemical changes observed reflect the increase in ROS following Cd exposure as described previously [Prithiviraj et al. Citation2013]. However, Cd itself is not involved in the generation of ROS, instead, it alters the levels of antioxidants and Zn available to the tissue by replacing Zn [Casalino et al. Citation2002]. Cadmium has also been reported to induce the release of other deposited heavy metals resulting in the increase of LPO [Casalino et al. Citation1997; Dorta et al. Citation2003]. The majority of antioxidant enzymes are deactivated because Cd binds to the active sites of the enzymes containing –SH groups thus blocking enzyme activity [Quig Citation1998].
The activity of antioxidant enzymes like CAT, POD, SOD, and GSR within the tissue were found to be reduced in the Cd treated group as compared to the antioxidant treated groups and control group. It has been reported previously that Cd exposure decreases CAT, SOD, POD, and GSR and increases TBARS [Awoniyi et al. 2011; Chandana et al. 2010; Lamia et al. Citation2010] resulting in the alteration of the tissue defense mechanism. Similarly, Cd induces the generation of free radicals within cardiac tissues, resulting in the abolition in the antioxidant enzymes and causing cardiovascular damage [Mukherjee et al. Citation2011]. A sufficient level of CAT is important for the reduction of ROS to reduce heavy metal toxicity [Gu and Hecht Citation1996]. Similarly SOD levels are important for the maintenance of H2O2 and O2 within the cell. If there is an imbalance in the H2O2 and O2, there is a reduction in CAT activity, resulting in the increase in free radicals [Aitken and Roman. Citation2008]. This increase in reactive OH– radicals may be due to the release of iron and sulfur from the proteins [Li and Shah 2004]. Cadmium has also been reported as a toxicant involved in the alteration of glutathione levels [Ikediobi et al. Citation2003]. The same results were observed in the present study, the glutathione levels were found depleted in the Cd alone treated group and were restored in the antioxidant treated groups.
In the antioxidant treated groups, the histological damage within the tissues was less as compared to the Cd alone treated groups. The damage within the tissue section was also evident by the presence of polynucleated cells within the sections as the number of polynucleated cells is linked to the presence of damage. The concentration of polynucleated cells were high in the Cd alone treated group as compared to the antioxidant treated group. The tissue is considered abnormal when there are polynucleated cells present per section (polynucleated or giant cells are often seen as spermatids in a common membrane, however other cells may be involved) [Bailey et al. Citation1981]. Similarly, the activity of antioxidant enzymes was also high as compared to the Cd alone treated group. This increase in the antioxidant enzyme activity may have protected the tissues from the damage induced by Cd. It has been well-known that vitamin E is the potent antioxidant that restores the antioxidant enzyme content of the tissue and protects histological changes within the tissue. Cadmium exposure leads to a reduction of non-enzymatic antioxidants like vitamin E and C levels within the tissue [Prithiviraj et al. Citation2013]. These non-enzymatic antioxidants are important for boosting the body defense mechanism against metal toxicity and are important for spermatogenesis [Johnson Citation1979; Paolicchi et al. Citation1996]. Co-administration of cadmium with either sulforaphane or Ficus religiosa offset the cadmium induced decrease in antioxidant enzymes which can be ascribed to the possible role of the protectant in preventing the surge in free radicals or by possibly boosting the level of non-enzymic antioxidants.
Ficus religiosa has antioxidant, anticancerous, and anti-inflamatoy effects [Arora et al. Citation1999; Blois Citation1958; De-Feudis et al. Citation2003; Panda Citation2005] and induces glutathione peroxidase (GPX), glutathione S-transferase (GST), glutathione reductase (GRD), superoxide dismutase (SOD), and catalase (CAT) activity and decreases the level of lipid peroxidation (LPO) against CCl4 induced liver toxicity [Gupta et al. Citation2004]. Similarly, sulforaphane is a natural enzymatic antioxidant found in vegetables like broccoli and brussels sprouts [Dinkova-Kostova and Talaya 2008; Fahey and Talalay Citation1999]. All parts are used in diseases of the blood, vagina, and uterus. The root-extract has been used as a treatment for sexual debility, fruit in diseases of the blood and heart, root in gout, root-bark in leucorrhoea, aphrodisiac, seed in urinary discharges, leaves as antiemetic, and fruits as purgative [Guha et al. Citation2001; Kirtikar and Basu, Citation2005]. The present findings are in accordance with the previous findings in which it was reported that in the cell, sulforaphane is involved in the translocation of Nrf2 into the nucleus which binds to the electrophile response element (EpRE) of the gene. This EpRE is associated with the transcription of different antioxidant enzymes like CAT, GST, GRD, and glutathione reductases [Dinkova-Kostova and Talalay, Citation2008].
These effects of sulforaphane and PE may be the same as vitamin E, acting as free radical hunters. These findings can be supported by the work of Kirana et al. (Citation2009), who reported that Ficus religiosa plant extract reduces the oxidative stress in diabetic conditions. In previous studies it has been reported that the effect of Cd on tissues is because of its absorbance or through binding different substances [Gunn and Gould Citation1970]. Antioxidant treatment reduced the tissue effects of Cd which likely reflect the increase in Zn in the tissue as observed previously [Jahan et al. Citation2014]. Cadmium uptake by the nuclear fraction is reduced by pretreatment with Zn and Se. Similarly, an increase in Se causes a decrease in the availability of the metals [Parizek Citation1971]. The increase in Zn increases the production of metallothionein like proteins that reduce the amount of free cadmium [Webb Citation1972]. The protection provided by these antioxidants may also be due to the metal chelating activity of these substances. It was found that the amount of Cd in the tissues as well as in the serum was low in the groups treated with vitamin E, sulforaphane, and PE of Ficus religiosa [Jahan et al. Citation2014].
Cadmium mediated testis pathological events are multifactorial and reflect the alteration of the antioxidant defense mechanism and immunity. In addition, the resulting damage to vascular bundles, blood - testis barrier ends in cellular necrosis. Supplementation of sulforaphane and Ficus Religiosa brings remarkable recovery. However, supplementation of vitamin E showed an improvement in the levels of antioxidants and enumerative effect on reproductive tissue. Ficus religiosa and sulforaphane may prove useful in the treatment of oxidative stress related to the toxicity of heavy metals and degeneration. This work supports the use of sulforaphane and Ficus religiosa in the protection against toxicity caused by exposure to heavy metals in reproductive tissues.
Materials and Methods
Plant collection and extract preparation
Plants of Ficus religiosa at maturity were collected during the month of May. Leaves were shade dried at room temperature for two w, chopped, and ground mechanically of mesh size 1 mm.
Methanolic leaf extract of Ficus religiosa was prepared by simple maceration process according to the method of Handa et al. [Citation2008], with slight modification. During experiment the crude extract was diluted with distilled water just before treatment. Methanolic extract was used because all the organic solvents are soluble in methanol and is easily extracted. Sulforaphane in the form of powder was obtained locally. All other chemicals including Cd in the form of CdCl2 and vitamin E in the form of tocopherol (97%) were purchased from Sigma-Aldrich (Germany).
Animals
Adult male Sprague-Dawley rats weighing 180 ± 20 g having age of 90–95 d used for this study were obtained from the rat colony maintained at the animal house in Quaid-i-Azam University, Islamabad. All the animals were kept at 12 h light/12 h dark period having free access to standard laboratory food and tap water was available ad libitum. Animals were restrained for one week prior to commencement of experiment. All the animal handling and sacrifications were approved by the Department committee of Animal Sciences Quaid-i-Azam University.
Experimental design
Animals were divided into five groups. The control group was injected subcutaneously with saline while the Cd alone treated group received subcutaneous injection of 0.2 mg/kg CdCl2. This dose of Cd was selected on the basis of previously used dose by Yang et al. [Citation2000]. Other groups were treated with sulphoraphane (25 µg/rat) [Cho et al. Citation2006], vitamin E (75 mg/kg) [Adaikpoh and Obi 2009], and PE (100 mg/kg) [Kirana et al. Citation2009] orally along with subcutaneous injection of 0.2 mg/kg CdCl2 for 15 d. Vitamin E was used as a standard antioxidant, to compare the effects of sulforaphane and PE with the vitamin treated group. At the end of treatment, animals were anesthetized with chloroform. Paired testes and prostate gland were dissected out quickly and washed in 0.9% cold normal saline and were weighed. One testis was fixed in sera for histological evaluation. The other testis and prostate gland were used for the enzyme assays.
Assessment of antioxidant enzymes
Testicular and prostatic tissue (90 mg) were homogenized in 10 volume of 100 mM potassium hydrogen phosphate (KH2PO4) buffer containing 1 mM EDTA, pH 7.4 and centrifuged at 12,000 × g for 30 min at 4°C. The supernatant was collected and used for the following assays.
The total protein of the homogenate and supernatant of tissue were determined by the method of Lowry et al. [Citation1951]. The concentration of soluble protein was calculated using Bovine’s serum albumin as a standard.
Catalase activities were determined by the method of Chance and Maehly [Citation1955] with some modification. The reaction solution of CAT activities contained 2.5 ml of 50 mM phosphate buffer (pH 5.0), 0.4 ml of 5.9 mM H2O2, and 0.1 ml enzyme extract. Changes in absorbance of the reaction solution at 240 nm were determined after one min. One unit of CAT activity was defined as an absorbance change of 0.01 as units/min.
Activities of POD were determined by the method of Chance and Maehly (Citation1955) with some modification. The POD reaction solution 3 ml contained 2.5 ml of 50 mM phosphate buffer (pH 5.0), 0.1 ml of 20 mM guaiacol, 0.3 ml of 40 mM H2O2, and 0.1 ml enzyme extract. Changes in absorbance of the reaction solution at 470 nm were determined after one min. One unit of POD activity was defined as an absorbance change of 0.01 as units/min.
Superoxide dismutase activity was estimated by the method of Kakkar et al. [Citation1984]. Reaction mixture contained 0.1 ml of phenazine methosulphate (186 μM), 1.2 ml of sodium pyrophosphate buffer (0.052 mM, pH 7.0), 0.3 ml of supernatant after centrifugation (1,500 × g, 10 min followed by 10,000 × g, 15 min) of 10% homogenate was added to the reaction mixture. Enzyme reaction was initiated by adding 0.2 ml of NADH (780 μM) and stopped after 1 min by adding 1 ml of glacial acetic acid. Amount of chromogen formed was measured by recording color intensity at 560 nm. Results are expressed in units/mg protein.
Glutathione reductase activity was determined by the method of Carlberg and Mannervik [1975]. The reaction solution composed of 1.65 ml phosphate buffer: (0.1 M, pH 7.6), 0.1 ml EDTA (0.5 mM), 0.1 ml NADPH (0.1 mM), 0.05 ml oxidized glutathione (1 mM), and 0.1 ml 10% homogenate in a total volume of 2 ml. Enzyme activity was quantitated at 25°C by measuring disappearance of NADPH at 340 nm and was calculated as nM NADPH oxidized/min/mg protein using molar extinction coefficient of 6.22 × 103/M cm.
The assay for lipid peroxidation was carried out following the method of Wright et al. [Citation1981] as modified by Iqbal et al. [Citation1996]. The reaction mixture in a total volume of 1.0 ml contained 0.58 ml phosphate buffer (0.1 M pH 7.4), 0.2 ml homogenate sample, 0.2 ml ascorbic acid (100 mM), and 0.02 ml ferric chloride (100 mM). The reaction mixture was incubation at 37°C for 1 h in a shaking water bath. The reaction was stopped by the addition of 1.0 ml 10% trichloroacetic acid, after the addition of 1.0 ml 0.67% thiobarbituric acid, all tubes were boiled in a water bath for 20 min and then shifted to a crushed ice bath before centrifuging at 2500 × g for 10 min. The amount of TBARS formed in each of the sample was assessed by measuring optical density of the supernatant at 535 nm using spectrophotometer 37°C using molar extinction coefficient of 1.56 × 105 M−1 cm−1.
Morphometrical analysis
For microscopic evaluation, tissues were immersed in fixative sera. Following dehydration in the descending grades of ethyl alcohol, tissues were clarified in cedar wood oil and then embedded in paraffin. Five μm thick sections were cut out of the paraffin block by using Reichert Microtome. Sections were then affixed to pre-cleaned albumenized glass slides and stretched at 60°C on Fisher slide warmer. Slides were then transferred to a paraffin oven for the next 12 h for the complete deparaffinization. Hematoxylin and Eosin staining was carried out and the slides were then examined under a Nikon optishot research microscope (Germany) with fixed automatic micro photographic system (Cannon, Japan). Histomorphometry was carried out by measuring tunica albuginea thickness, seminiferous tubule diameter, and epithelial height. Mean number of spermatogonia, primary spermatocytes, secondary spermatocytes, and spermatids in the control and treated groups was reported.
Statistical analysis
Mean ± SEM of morphological and histological data of control, CdCl2 treated and various antioxidants treated groups were determined, analyzed, and compared by ANOVA (analysis of variance) and Tukey’s test, using SPSS. A difference was considered significant at the p < 0.05 level.
Declaration of interest
This piece of work was supported and funded by the Department of Animal Sciences, Quaid-i-Azam University, Islamabad. Pakistan. The authors report no conflict of interest. The authors alone are responsible for the content and writing of the paper.
Author contributions
Led the design, conceived the study, performed the experimental work, and analyzed the results: SJ; Conceived an initial part of the study and performed the experiment: AZ, UI; Performed the histology and helped in compiling the results: NI; Wrote the paper with input from all other authors: HU.
| Abbreviations | ||
| Cd | = | cadmium |
| CAT | = | catalase |
| POD | = | peroxidise |
| SOD | = | superoxide dismutase |
| GSR | = | glutathione reductase |
| TBARS | = | lipid peroxidation |
| ROS | = | reactive oxygen species |
| LPO | = | lipid peroxidation |
| PE | = | plant extract |
| GPX | = | glutathione peroxidise |
| GST | = | glutathione S-transferase |
| GRD | = | glutathione reductase |
| EpRE | = | electrophile response element |
References
- Aitken, R.J., and Roman, S.D. (2008) Antioxidant system and oxidative stress in the testes. Oxid Med Cell Longev 1:15–24
- Arora, D., Dubey, S.D., and Ojha, J.K. (1999) Role of Rasayana in preventing the complications of diabetes mellitus. J Diabet Associa of India 39:47–50
- Awoniyi, D.O., Aboual, Y.G., Marnewick, J.L., Plesis, S.S., and Brooks, N.L. (2012) The effects of rooibos (Aspalathus linearis), green tea (Camellia sinensis) and commercial rooibos and green tea supplement on epididymal sperm in oxidative stress-induced rats. Phytother Res 26:1231–9
- Azzi, A., Boscoboinik, D., Chatelain, E., Ozer, N.K., and Stauble, B. (1993) d-alpha-tocopherol control of cell proliferation. Mol Aspects Med 14:265–71
- Bailey, K.I., O’Brain, W.D., and Dunn, F. (1981) Ultrasonically induced, in vivo morphological damage in mouse testicular tissue. Arch Androl 6:301–6
- Blois, M.S. (1958) Antioxidant determinations by the use of a stable free radical. Nature 181:1199–200
- Bonda, E., Wlostowski, T., and Krasowska, A. (2004) Testicular toxicity induced by dietary cadmium is associated with decreased testicular zinc and increased hepatic and renal metallothionein and zinc in the bank vole. Biometals 17:615–24
- Carlrberg, I., and Mannervik, B. (1975) Purification and characterization of the flavoenzyme glutathione reductase from rat liver. J Biol Chem 250:5475--80
- Casalino, E., Calzaretti, G., Sblano, C., and Landriscina, C. (2002) Molecular inhibitory mechanisms of antioxidant enzymes in rat liver and kidney by cadmium. Toxicology 179:37–50
- Casalino, E., Sblano, C., and Clemente, L. (1997) Enzyme activity alteration by cadmium administration to rats: The possibility of iron involvement in the lipid peroxidation. Arch Biochem and Biophysic 346:171–9
- Chance, B., and Maehly, A.C. (1955) Assay of catalase and peroxidase. Methods Enzymol 2:764–75
- Chanda, M., Tushar, K.B., Kazi, M.A., Kausik, C., and Debidas, G. (2010) Diabetes-induced testicular disorders vis-a-vis germ cell apoptosis in albino rat: Remedial effect of hexane fraction of root of musa paradisiaca and leaf of coccinia indica. J Health Sci 56:641–54
- Cho, E.J., Lee, Y.A., Yoo, H.H., and Yokozawa, T. (2006) Protective effect of broccoli (brassica oleracea) against oxidative damage in vitro and in vivo. J Nutr Sci Vitaminol 52:437–44
- De-Feudis, F.V., Papadopoulos, V., Drieu, K., and Fundam, C. (2003) Ginkgo hiloha extracts and cancer: A research area in its infancy. Fund Clin Pharmacol 17:405–17
- Dinkova-Kostova, A.T., and Talalay, P. (2008) Direct and indirect antioxidant properties of inducers of cytoprotective proteins. Mol Nutr Food Res 52:S128–38
- Dorta, D.J., Leite, S., DeMarco, K.C., Prado, I.M.R., Rodrigues, T., Mingatto, F.E., et al. (2003) A proposed sequence of events for cadmium-induced mitochondrial impairment. J Inorg Biochem 97:251–7
- El-Demerdash, F.M., Youself, M.I., Kedwany, F.S., and Baghidadi, H.H. (2004) Cadmium induced changes in lipid peroxidation blood hematology, biochemical parameters and semen quality of male rats: Protective role of vitamin E and beta-carotene. Food Chem Toxicol 42:1563--71
- El-Shahat, A.E.R., Gabr, A., Meki, A.R., and Mehana, E.S. (2009) Altered testicular morphology and oxidative stress induced by cadmium in experimental rats and protective effect of simultaneous green tea extract. Int J Morphol 27:757--64
- Fahey, J.W., and Talalay, P. (1999) Antioxidant functions of sulforaphane: A potent inducer of phase II detoxication enzymes. Food Chem Toxicol 37:973–9
- Gu, W., and Hecht, B. (1996) Developmental expression of glutathione peroxidase, catalase, and manganese superoxidase dismutase mRNAs during spermatogenesis in the mouse. J Androl 17:256–62
- Guerrero-Beltran, C.E., Calderon-Oliver, M., Tapia, E., Omar, N., Campos. M., Javier, D., et al. (2010) Sulforaphane protects against cisplatin-induced nephrotoxicity. Toxicol Lett 192(3):278--85
- Guha, B.D.N., Sensarma, P., and Pal, D.C. (2001) A Lexicon of Medicinal Plants in India. Naya Prokash, Bidhan Sarani, Calcutta, India, pp. 201–4
- Gunn, S.A., and Gould, T.C. (1970) Cadmium and other mineral elements. In The Testis vol. IIT. Johnson, A.D., Gomes, W.R., and Van Demark, N.L., ed. Academic Press. New York. pp. 377
- Gupta, R., Gupta, E., Dhakal, K., Thakur, R., and Ahnn, J. (2004) Vitamin C and vitamin E protect the rat testes from cadmium-induced reactive oxygen species. Mole Cells 17:132–9
- Handa, S.S., Khanuja, S.P.S., Longo, G., and Rakesh, D.D. (2008) Extraction Technologies for Medicinal and Aromatic Plants. International centre for Science and High Technology, Trieste, Italy, pp. 21–5
- Ikediobi, C.O., Badisa, V.D.L., Ayuk-Takem, L.T., Latinwo, L.M., and West, J. (2004) Response of antioxidant enzymes and redox metabolites to cadmium-induced oxidative stress in CRL-1439 normal rat liver cells. Int J Mol Med 14:87–92
- Iqbal, Z., Prince-Danso, C.S. Hayat, M., and Nisar, K. (1996) Epidemiology of Hydatid Disease. Echinococcosis in dogs and jackals in Faisalabad (Pakistan). Indian Vet J 73:620–2
- Jahan, S., Khan, M., Ahmed, S., and Ullah H. (2014) Comparative analysis of antioxidants against cadmium induced reproductive toxicity in adult male rats. Syst Biol Reprod Med 60:28–34
- Johnson, F.C. (1979) The antioxidant vitamins. CRC Crit Rev Food Sci Nutr 11:217–309
- Kakkar, P., Das, B., and Viswanathan, P.N. (1984) A modify spectrometric assay of super oxide dismutase. Indian J Biochem Biophys 21:130–2
- Kini, R.D., Tripathi, Y., Raghuveer, C.V., Sheila, R.P., Ramswamy, C., Nayanatara, A.K., et al. (2009) Protective role of vitamin E against Cadmium chloride induced testicular damage in rats. JPBS 22(2):12--16
- Kirana, H., Agrawal, S.S., and Srinivasan, B.P. (2009) Aqueous extract of Ficus religiosa Linn reduces oxidative stress in experimentally induced type 2 diabetic rats. Indian J Exp Biol 47:822–6
- Kirtikar, K.R., and Basu, B.D. (2005) Indian Medicinal Plants. Volume III, International Book Distributors, Dehradun (Uttaranchal), India, pp. 2327–9
- Lamia, S., Mohamed, B., Abdelhamid, K., Khaled, S., and Imed, M. (2010) Influence of combined treatment with zinc and selenium on cadmium induce testicular pathophysiology in rat. Food and Chem Toxicol 48:2759–65
- Li, J.M., and Shah, A.M. (2003) ROS generation by nonphagocytic NADPH oxidase: Potential relevance in diabetic nephropathy. J Am Soc Nephrol 14:S221–S6
- Lowry, O.H., Rosenbrough, N.J., Farr, A.L., and Randall, R.J. (1951) Protein measurement with Folin phenol reagent. J Biol Chem 193:265–75
- Martin, J.J., Martin, R., Codesal, J., Fraile, B., Paniagua, R., and Santamaria, L. (2001) Cadmium chloride-induced dysplastic change in the ventral rat prostste: An immunohistochemical and quantitative study. Prostate 46:11–20
- Mukherjee. R., Banerjee, S., Joshi, N., Singh, P.K., Baxi, D., and Ramachandran, A.V. (2011) A combination of melatonin and alpha lipoic acid has greater cardioprotective effect than either of them singly against cadmium-induced oxidative damage. Cardiovasc Toxicol 11:78–88
- Nandi, D., Liu, X., Winter, J.L., Miall, R.C., Aziz, T.Z., and Stein, J.F. (2001) Frequency-dependent effects of chronic deep brain stimulation of the pedunculopontine region in a normal non-human primate. J Physiol 533:68
- Panda, H. (2000) Herb cultivation and medicinal uses. (2nd ed) National Institute of Industrial Research, Delhi India. pp. 302–3
- Paolicchi, A., Pezzini, A., and Saviozzi, M. (1996) Localization of a GSH-dependent dehydroascorbate reductase in rat tissues and subcellular fractions. Arch Biochem Biophys 333:489–95
- Parizek, J., Ostadalova, I., Kalouskova, J., Babicky, A., and Benes, J. (1971) The detoxifying effects of selenium. Interrelations between compounds of selenium and certain metals. In: Newer Trace Elements in Nutrition. Mertz, W. and Cornatzer, W.E. eds., Marcel Dekker, New York. pp. 85
- Prithiviraj, E., Suresh, S., Lakshmi, N.V., Ganesh, M.K., Ganesh, L., and Parkash, S. (2013) Protective effect of Eithania somnifera (Linn.) on cadmium-induced oxidative injury in rat testis. Phytopharmacol 4:269–90
- Prozialeck, W.C., Vaidya, V.S., Liu, J., Waalkes, M.P., Edward, J.R., Lamar, P.C., et al. (2007) Kidney injury molecule-1 is an early biomarker of cadmium nephrotoxicity. Kidney Int 72:985–93
- Quig, D. (1998) Cystein metabolism and metal toxicity. Altern Med Rev 3:262–70
- Sen-Gupta, R., Sen-Gupta, E., Dhakal, B.K., Thakur, A.R., and Ahnn, J. (2004) Vitamin C and vitamin E protect the rat testies from cadmium induced reactive oxygen species. Mol Cells 17:132–9
- Tremellen, K., Miari, G. and Froiland, R. (2008) oxidative stress and male infertility – a clinical perspective. Hum Reprod Update 14:243–58
- Webb, M. (1972) Protection by zinc against cadmium toxicity. Biochem Pharmacol 21:2767--71
- Wright, C., Nicholson, S., Angus, V., Sainsbury, R.J., Farndon, J., Cairns, J., et al. (1981) Relationship between c-erbB-2 protein product expression and response to endocrine therapy in advanced breast cancer. Br J Cancer 65:118–21
- Yang, H., Carter, C.E., Xu, C., Singh, P.K., Johns, M.M., Johnson, J.E., et al. (2000) Cadmium induced apoptosis in the urogenital organs of the male rat and its suppression by chelation. J Toxicol Environ Health 52:149–68
- Yu, H.N., Shen, S.R., and Yin, J.J. (2007) Effect of metal ions, catechins, and their interaction on prostate cancer. Crit Rev Food Sci Nutr 47:711–9