Abstract
There are substantial declines in behavioral measures of cognitive function with age, including decreased function of executive processes and long-term memory. There is also evidence that, with age, there is a decrease in brain volume, particularly in the frontal cortex. When young and older adults perform cognitive tasks that depend heavily on frontal function, neuroimaging evidence indicates that older adults recruit additional brain regions in order to perform the tasks. This additional neural recruitment is termed “dedifferentiation,” and can take multiple forms. This recruitment of additional neural tissue with age to perform cognitive tasks was not reflected in the behavioral literature, and suggests that there is more plasticity in the ability to organize brain function than was previously suspected. We review both behavioral and neuroscience perspectives on cognitive aging, and then connect the findings in the two areas. From this integration, we suggest important unresolved questions and directions for future research.
Con la edad hay disminuciones sustanciales de las mediciones conductuales de la función cognitiva, incluyendo una disminución de la función de procesos de ejecución y memoria a largo plazo. También existen evidencias que con la edad hay una reducción en el volumen cerebral, particularmente en la corteza frontal. Cuando jóvenes y adultos mayores ejecutan pruebas cognitivas que dependen en gran medida de la función frontal, la evidencia de las neuroimágenes indica que los adultos mayores reclutan en forma adicional regiones cerebrales para poder realizar las pruebas. Este reclutamiento neural adicional se ha denominado “dediferenciación” y puede tomar múltiples formas. Este reclutamiento de tejido neural adicional con la edad para ejecutar pruebas cognitivas no se ha reflejado en la literatura conductual y se sugiere que existe una mayor plasticidad en la capacidad para organizar la función cerebral de lo que previamente se sospechaba. Nosotros revisamos tanto la perspectiva conductual como la de las neurociencias en el envejecimiento cognitivo y luego conectamos los hallazgos en las dos áreas. A partir de esta integración nosotros sugerimos importantes preguntas no resueltas y orientaciones para futuras investigaciones.
Un déclin important, lié à l'âge, est constaté lors des mesures des fonctions cognitives, avec en particulier une diminution fonctionnelle des processus d'exécution et de la mémoire à long terme. Une diminution du volume cérébral avec l'âge, concernant principalement le cortex frontal, a également été mise en évidence. La neuro-imagerie montre que, lors de l'accomplissement de tâches cognitives dépendant étroitement de la fonction frontale, les personnes âgées recrutent des régions cérébrales supplémentaires par rapport aux adultes jeunes. Ce recrutement neural supplémentaire, désigné sous le terme de « dédifférenciation », peut prendre de multiples formes. Ce phénomène, non pris en compte par la littérature comportementale, suggère qu'il existe plus de plasticité dans l'organisation de la fonction cérébrale qu'il n'avait été soupçonné jusqu'à présent. Cet article fait le point sur les acquis concernant le vieillissement cognitif, tant sur le plan comportemental que sur celui des neurosciences, afin de les mettre en liaison. A partir de cette réflexion, nous soulevons d'importantes questions restées sans réponse et ouvrons des voies pour la recherche future.
There is considerable folk wisdom about cognitive aging in our culture. One familiar but disheartening proverb suggests that “You can't teach an old dog new tricks,” while a more optimistic one states that “Older is wiser.” A third proverb, “Use it or lose it” seems to be a restatement of the American dream, as it implies that anyone can earn or maintain their “cognitive fortune” into late adulthood if only one works hard enough. Perhaps this third proverb explains the paradox represented in the first two. These proverbs encompass issues that have been of considerable concern to cognitive aging researchers for the past 25 years. Do older adults learn new information as effectively as the young? Does knowledge (a basis for wisdom) increase with age? What is the role of experience and cognitive “exercise” in protecting the cognitive system from age effects? Can using the mind actually protect it from cognitive loss or compensate for cognitive losses already sustained, much like exercise and diet can prevent or repair heart disease? Behavioral scientists have learned a great deal about these issues, but their knowledge is still incomplete. Recently, the ability to look into the mind with functional imaging techniques has provided scientists with new tools to address these questions.
The goal of the present paper is to provide one view of the relationship between behavioral findings about cognitive aging and their neural underpinnings, drawing primarily from structural and functional data about the aging brain. We first briefly review some of the cognitive mechanisms that have been isolated by behavioral studies as central to understanding agerelated cognitive decline and follow this with an overview of new findings about the aging brain from the structural and functional imaging literature. We also consider some methodological issues associated with imaging that constrain our ability to integrate brain and behavioral data. We then provide integration between the behavioral and neural findings, discussing findings where the brain and behavioral data are consistent, and other areas where they are inconsistent or simply unconnected. This integrative analysis provides us with a window into what questions and hypotheses are most in need of exploration in future work in understanding cerebral aging.
Aging, cognition, and behavior: an overview
There is a wealth of evidence suggesting that older adults have more trouble learning new information, exhibit less efficient reasoning skills, are slower to respond on all types of cognitive tasks, and are more susceptible to disruption from interfering information than younger adults (see reviews on these topics in edited volumes by Park and Schwarz,Citation1 and Craik and SalthouseCitation2). Below we review some of the major findings.
The nature of decline
There are a variety of mechanisms that have been suggested as fundamental causes for the decreased memory and information-processing abilities of older adults and they fall into two general categories. One is a global, undifferentiated, single-mechanism view. For example, SalthouseCitation3,Citation4 has suggested that age-related declines in the speed at which information is processed account for age differences on essentially all cognitive tasks. Speed of processing is measured by how rapidly young and old adults can make simple same/different judgments when presented with two figures or strings of letters or digits side by side. This simple task shows consistent declines across the life span () and predicts most, if not all, age-related variance on a broad array of cognitive tasks. In a related vein, Baltes and LindenbergerCitation5 have suggested that crude measures of sensory function (visual and auditory acuity) are even more fundamental than speed of processing in explaining age differences. They found that these sensory measures explained 49% of overall task variance on 14 different tasks in a sample of adults aged 69 to 105. They argue that this relationship is so strong because sensory function provides a crude overall measure of declining neuronal integrity in the older adult. The view that all types of cognitive decline with age are caused by a single mechanism has been labeled the “common cause” hypothesis.Citation6
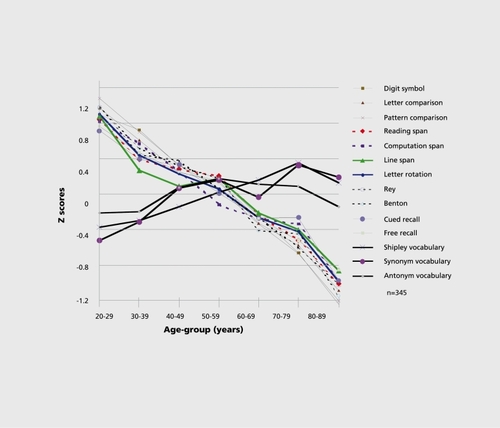
An alternative approach assumes that age-related declines are due to problems with specific cognitive mechanisms. For example, there is evidence that executive functions (eg, working memory processes, inhibitory function, and the ability to switch among tasks) decline with age (see ParkCitation7 and Zacks et al,Citation8 for reviews). Executive functions are used in service of many cognitive tasks, including reasoning, strategic encoding, and retrieval of information in long-term memory, and many everyday or work-related tasks that require learning or responding to novel information. We will consider working memory first, as it is the best understood of the executive functions. Working memory deficits can be thought of as decreased on-line capacity, or limited ability to store, process, and manipulate information. Working memory is typically assumed to consist of a central executive system, which processes information drawn from two passive storage subsystems that are domain-specific - verbal and visiospatial stores.Citation9 Virtually all cognitive tasks that require effortful processing are thought to require working memory. There are a range of models of working memory,Citation10,Citation11 but most are variants of the Baddeley model.Citation9 Regardless of the view one adopts, there is no question that working memory declines with age, particularly the processing or central executive component, as evidenced by the data presented in Figure 1.Citation12,Citation13
Hasher and ZacksCitation14 have emphasized the role of inhibition, rather than working memory, in age-related cognitive declines. They proposed that older adults are less effective at inhibiting irrelevant information than young adults. They argue that working memory capacity is not limited with age, but rather is filled with “mental clutter” so that capacity appears to be diminished. Older adults are assumed to be particularly deficient in a deletion operation in working memory whereby irrelevant information is efficiently discarded when it is no longer needed. There is considerable evidence that older adults have difficulty inhibiting irrelevant information in working memory.Citation15-Citation18
There is also evidence that older adults have difficulty in flexibly deploying mental resources and/or switching among different tasks. Two good everyday examples that require task switching are day-trading stocks and piloting an airplane. Both the day trader and airline pilot must constantly shift attention among various indicators and adjust their behavior accordingly (eg, trade a stock or adjust altitude). There is considerable evidence that older adults have difficulty switching from one operation to another relative to young adults,Citation19-Citation21 with older adults showing larger time costs for switching between tasks (or metaphorically, reloading mental software) than young adults, when compared with receiving repeated trials on the same task.
Another process that is important for many cognitive tasks, but particularly long-term memory tasks, is the ability to connect a memory event to a context. For example, an older adult may remember that he was told to take shark cartilage to improve his/her arthritis. What the older adult may not remember is whether they were instructed to do this by a personal physician or were told to do this by a friend who had just read it in the popular press. Thus, the older adult may find they remember a fact, but not the source of the information.Citation22,Citation23 This decreased ability to bind target information to source or context is a problem for older adults in long-term memory tasksCitation24-Citation26 as well as working memory.Citation15 Finally, there is no question that long-term memory declines with age (Figure 1). Problems with decreases in speed, working memory, switching, inhibition, and binding could all contribute to poor long-term memory. One problem with the behavioral literature in cognitive aging is that hypotheses about the mechanisms of age-related decline on cognitive tasks have frequently been presented as though evidence for age-sensitivity in one mechanism is evidence against another. This type of thinking is naive. It is likely that all of the different executive processes, as well as speed of processing, decline with age and collectively contribute to difficulties in reasoning, memory, and other higher order cognitive functions. Park et alCitation12 examined the joint contributions of speed and working memory to long-term memory function across the life span using structural equation models, and showed that both were important in explaining long-term memory function. This study thus verifies that global mechanisms like speed, as well as specific mechanisms like working memory, are fundamental in explaining variance in higher-order cognitive function. In a later study, Park et alCitation13 reiterated this finding for both visiospatial and verbal working and long-term memory. It is also important to recognize that these mechanisms may not all decline at the same rate within subjects and that deterioration of different mechanisms might be controlling similar levels of decline in different individuals. Cognitive aging researchers have devoted relatively little attention to understanding how the different mechanisms work separately or together in specific individuals to explain declines in cognitive function, and this is an important direction for future research.
Other important behavioral issues
Three other important issues with respect to behavioral changes in cognitive performance with age relate to rates of change across the life span, spared cognitive processes, and the potential to improve cognitive function.
Decline is continuous across the life span
Besides demonstrating that knowledge may increase with age while performance on processing-intensive tasks decreases, Figure 1 also illustrates one poorly understood and little-recognized phenomenon of some importance: both cross-sectional and longitudinal data suggest that decline is relatively continuous across the life span, beginning in young adulthood. We believe that symptoms of cognitive decline that begin occurring in the second decade of life become more noticeable in late adulthood only because they have accrued to the point where they result in noticeable declines in function.Citation27 As a crude metaphor, consider that an individual with a million dollars probably would not be very concerned initially about the loss of $2000 per year, but would be much more likely to notice and worry 50 years later when $100 000 in losses had accrued. So it seems to be with cognitive aging. It is also important to consider that losses of executive function and long-term memory in middle-aged adults may not be too apparent to the individual because they are offset by increases in expertise and skill. Little is understood about the interplay among increasing knowledge and declining executive function and memory across the life span. This is a tremendously important issue that we will have more to say about later.
Spared processes and growth of knowledge
We should also note that memory decline does not occur on tasks that arc low in their requirements for executive processes, ie, tasks where little effortful and strategic processing is required. Memory tasks where age invariance has been demonstrated include picture recognitionCitation28 and implicit memoryCitation29-Citation31 - both passive, nonstratcgic tasks. Also, there is some evidence that age differences are more pronounced for encoding new material, compared with retrieving information,Citation32,Citation33 due to the obligatory or automatic nature of retrieval.Citation34 Finally, knowledge organization and semantic memory remain relatively unchanged with age.Citation35,Citation36 Figure 1 demonstrates that world knowledge, as measured by vocabulary, is not subjected to age-related declines, and may even show growth across the life span. As Figure 1 illustrates, it is only tasks that require considerable mental effort or deliberate recollection that show age declines.Citation37
Repairing declining cognition
The primary view of how to “repair” declining cognition among cognitive aging psychologists has been to provide older adults with memory supports that bypass the need to encode items actively or deliberately, thus lessening the executive processing requirements of the tasks.Citation38 Specific examples in which environmental supports have effectively repaired the memory of older adults include (i) using matching pictures to support memory for sentences, resulting in dramatically better memory in older adults (probably due to dual coding of the material, which provides two routes to recallCitation39); and (ii) providing older adults with memory cues that are conceptually related to target words, resulting in larger memory improvements for the words in the old compared with the young.Citation40 The conceptually related cues built upon existing world knowledge possessed by older adults, and relied on activation of existing information rather than only on engagement of executive processes (primarily working memory).
Neural underpinnings of cognitive aging
Recently, the fields of neurosciencc and cognitive aging have begun to merge, and we have already learned a number of things that were not previously considered in a systematic way in the cognitive aging literature. In the following sections, we discuss what is known about neural decline and reorganization, as it relates to aging.
Brain atrophy
Structural imaging of brains indicate that, although the brain shrinks or shows volumetric changes with age, these changes are not equal across brain structures. There is more shrinkage with age in the frontal cortex, intermediate shrinkage in mediotemporal areas, and little decrease in volume in the occipital cortex.Citation41 There are decreases in both gray matter and white matter, and evidence for demyclination.Citation41,Citation42 Increases in white matter hyperintensities occur with age.Citation43 These are small decreases in white matter tissue irregularly distributed across regions. These lesions are viewed as pathological, but their cause is poorly understood and may be related to transient cerebral ischemia or hypertension. At the cellular level, dopamine receptors,Citation44,Citation45 transporters,Citation45 and storage vesiclesCitation46 all decline with aging, with rate estimates at 2% to 8% per decade. Volkow et alCitation47 found that this loss correlated with decreased activity in frontal and cingulate metabolism.
Dedifferentiation of brain activations with age
Types of dedifferentiation
There is considerable evidence from positron emission tomography (PET) and functional magnetic resonance imaging (fMRI) studies that older adults show less specificity or differentiation in brain recruitment while performing an array of cognitive tasks. We term this more diffuse pattern of brain activation with age “dedifferentiation” (along with other authors, including CabezaCitation48), but believe that the concept of dedifferentiation has multiple meanings that have not been systematically defined. Broadly put, by dedifferentiation, we mean that the number of brain sites recruited to perform a task increases with age or is different from sites used by young adults. We propose to classify dedifferentiation of neural function into three types.
One form of dedifferentiation is contralateral recruitment, in which young adults perform a cognitive task in a single hemisphere (eg, prefrontal cortex in the left hemisphere) and older adults recruit the homologous site in the other hemisphere (eg, prefrontal cortex in the left and right hemispheres), thus using two specific and homologous sites to perform a task. In this case, the dedifferentiation is characterized by a decrease in lateralization of function, but is nevertheless site-specific. In a seminal study in older adults using PET, Reuter-Lorenz et alCitation49 provided strong evidence for contralateral recruitment of dorsolateral prefrontal cortex in old adults on visual and verbal working memory tasks. In contrast, young adults showed typical patterns of left lateralization for verbal materials (letters) and right lateralization for judgments about the spatial location of the letters.
There is also some evidence for a second form of dedifferentiation, which we will call unique recruitment. In this case, older adults recruit additional brain areas that are not homologous to any sites activated in the young. Mcintosh et alCitation50 reported evidence for unique recruitment of sites in a working memory tasks in which participants were required to maintain memory for abstract stimuli that varied in texture. Participants held the stimuli in memory for either 500 or 4000 ms. Older adults recruited unique areas in the dorsolateral prefrontal cortex and mediotemporal areas, compared with young adults, and the areas recruited related to length of stimulus maintenance and task performance, whereas there was no relationship to recruitment and task performance in young adults.
A third form of dedifferentiation is substitution, in which a site that is used to perform a specific cognitive operation in young adults is not significantly activated by older adults, but different structures show activation. Hazlett et alCitation51 reported that old adults who performed like young adults on a verbal memory study showed activation of occipital cortex to perform the verbal memory task, whereas young adults showed primarily dorsolateral prefrontal cortex activation.
CabezaCitation48 distinguishes between dedifferentiation and compensation, suggesting that dedifferentiation involves a “difficulty in engaging specialized neural mechanisms,” whereas compensation involves the recruitment of additional neural tissue from specific sites to “counteract neurocognitive deficits,” such as in instances of bilateral recruitment. We would argue that any additional recruitment of brain areas, even homologous areas, is a form of dedifferentiation, and that dedifferentiation with age may be neutral (has no effect on performance), compensatory (improves performance), or neuropathologi cal (worsens performance or predicts later neurological disease). One important task of cognitive aging researchers is to understand these patterns and their implications for cognitive health and function, as well as whether they define a future cognitive trajectory.
Dedifferentiation of executive functions and long-term memory
Nearly all studies of the neuroscience of cognitive aging have provided evidence for one of the dedifferentiation mechanisms described above. Executive function and long-term memory have been the focus of most neuroimaging studies on aging. A summary of some selected and representative findings from this voluminous literature appear below. More detailed reviews of the literature on aging, imaging, and cognitive processes arc available.Citation48-Citation52,Citation56
Working memory and dedifferentiation. Perhaps the simplest statement about working memory and aging is that older and younger adults show different patterns of brain activations on these tasks. Before discussing differences, it would be useful to briefly review our present understanding of the neural organization of working memory. There is general agreement that stimuli that have high processing demands are processed bilaterally in the dorsolateral prefrontal cortex.Citation57-Citation60 Storage or maintenance functions in the working memory subsystems are lateralizcd for content and reside in the ventral lateral prefrontal cortex. Thus, when one presents tasks that are primarily storage-based, young adults will show left frontal activations for verbal materials and right frontal activations for visiospatial materials.
The study by Reuter-Lorenz et alCitation49 mentioned earlier provided evidence for contralateral recruitment of neural tissue for visiospatial and verbal working memory. In a related finding, Rypma and D'EspositoCitation61 used an event-related fMRI design to study working memory and aging. Event-related designs permit the investigator to examine activation across the different phases of stimulus presentation, storage, and response. Using this design, Rypma and D'Esposito found evidence for differential recruitment of dorsolateral prefrontal cortex in young and old, but only at retrieval and when the load was high. Both of these studies suggest that older adults may recruit brain tissue beyond the ventral storage areas at lower working memory loads than young adults do, thus making demands on the dorsolateral prefrontal cortex (where executive processes reside) earlier than young adults do. The Rypma and D'EspositoCitation61 study is also important in suggesting that the locus of the effect they observed is at the response (retrieval) stage, rather than at the encoding stage.
Inhibition and task-switching and dedifferentiation. The data on inhibition and task switching are relatively sparse. In one study on the imaging of interference effects, Jonides et alCitation62 demonstrated larger interference effects behaviorally for older adults compared with younger adults by creating response conflict in a verbal working memory task. Young adults showed more activation than older adults in the left lateral prefrontal cortex, and this appears to be an important site for mediating response conflicts in the young, since they showed smaller interference effects than the old. There were no other differences in activations between young and old, and so the study primarily provides information about how the young deal with response interference. In a study on task -switching in working memory, Smith et alCitation59 found age and performance to be important variables related to activation patterns. Both old adults and poorly performing young adults recruited left prefrontal cortex during a dual-task condition involving simple computations and storage, whereas young adults who performed well did not show these frontal activations. In both of these studies, patterns of unique recruitment were observed. For inhibition, young adults showed unique recruitment compared with older adults, whereas for a switching task, older adults and poor young adults showed unique recruitment of frontal regions.
Binding and dedifferentiation. Mitchell et alCitation63 investigated the ability of young and older adults to bind object with feature information (color, spatial) in a working memory task. They found evidence for a disturbed prefrontal/hippocampal circuit in older adults for performance of the binding operation. Older adults showed less activation than the young of anterior hippocampal structures in a binding condition compared with an object-only or feature-only condition, and also showed evidence of less activation in right prefrontal Brodmann area 10 for the binding condition. This important work provides an excellent bridge to understanding the relationship of working memory to long-term memory function, as it is primarily in long-term memory that source and binding operations have been demonstrated to be deficient.
Long-term memory and dedifferentiation. The study of long-term memory is particularly interesting because there are well-defined stages of encoding and retrieval. One might expect that different patterns of activation would occur in these two phases in young adults when compared with older adults. There is a large and complex literature on aging, neuroimaging, and long-term memory that can only be presented at a cursor}' level in this paper. The short story for young adults is that there is evidence for left frontal activation at encoding and right frontal activation at retrieval.Citation64 Buckner and LoganCitation52 speculate that the left frontal region associated with encoding dissociates functionally in young adults, with dorsal regions being more important for lower level processing such as lexical and phonemic access, and the ventral region more important for elaborative, semantic processing. There is also clear evidence that remembered stimuli show activation at encoding in mediotemporal structures, whereas stimuli that are forgotten do not show evidence of mediotemporal involvement.Citation65,Citation66
The aging story is more complex with different patterns of recruitment compared with the young evidenced at both encoding and retrieval. In studies of encoding, there has generally been evidence for less activation of left prefrontal cortex in old adultsCitation67,Citation68 and also less engagement of mediotemporal structures.Citation68,Citation69 At retrieval, there is evidence for bilateral recruitment of frontal cortex,Citation67 whereas the young typically activate only right frontal cortex. Madden et alCitation70 also found evidence for bilateral recruitment at retrieval in dorsolateral prefrontal cortex. There are a range of opinions about the meaning of the recruitment patterns for long-term memory observed in older adults. Madden et alCitation71 found no evidence for deactivations in any areas with age, and so they argued that the increased activations observed were not compensatory for deactivations in other areas, but rather evidence for more distributed neural networks with age (unique recruitment). Madden et alCitation70,Citation71 used stepwise regressions to determine how well activated regions predicted reaction time (RT) performance. They reported that activations in a number of structures predicted RT performance in old but not young and reiterated this finding in a more precise analysis of gaussian components of reaction time. CabezaCitation48 suggests that the breadth of evidence for more distributed processes in older adults in long-term memory could reflect either healthy compensatory mechanisms or a general decline in brain efficiency (dedifferentiation) that is pathological. CabezaCitation48 views compensation as an optimistic and distinct account of age-related increases in recruitment from a pessimistic dedifferentiation view. Rather than viewing compensation and dedifferentiation as conflicting mechanisms accounting for a phenomenon, we suggest that dedifferentiation is a description of activation patterns, and may be compensatory or pathological. It is important to note, however, that CabezaCitation48 docs present evidence in a review of the literature suggesting that it is generally good performers who show bilateral recruitment, whereas poor old performers show hemispheric asymmetry like the young. Finally, Buckner and LoganCitation52 suggest that it is important to distinguish between selective and nonselective recruitment, because nonselective recruitment reflects dysfunction in performing cognitive tasks. They argue that older adults are less able to recruit appropriate frontal regions for task performance and the areas they do recruit are less appropriate for the memory task they must perform. Thus, Buckner and Logan see dedifferentiated patterns of recruitment as dysfunctional.
Plasticity
The notion that the brain is plastic and can reorganize with age was quite startling to behavioral researchers in cognitive aging, who had focused primarily on (i) demonstrating memory system changes versus invariance in cognitive function with age; and (ii) learning how to “repair” cognition through providing external cognitive cues, aids, or supports. We now have compelling evidence that the brain docs not just reflect passive decline, but is a dynamic structure that is reactive to experience, evidencing both gains and losses with age. Indeed, Kempermann et alCitation72 have demonstrated growth of new neurons in response to complex environments in aged rats, suggesting that stimulating experiences may enhance the organization and function of the brain. At this point, the neuroimaging literature on aging has just begun to uncover some of the activation patterns that characterize aging. This issue is a most exciting and challenging area of research - to understand how a process such as dedifferentiation, a term descriptively applied to imaging results, might reflect changes in neuronal plasticity.
Methodological issues in integrating functional brain data with behavioral data on aging
Before we discuss integration of brain and behavioural data in cognitive aging, we should note that there are many technical difficulties involved in relating patterns of cognitive behaviors across different age groups to patterns of brain activations. We do not have space to go into detail about these problems, but here are some important issues to keep in mind when interpreting results from neuroimaging studies on aging. First, old and young adults may differ in the strength of signal that is detectable from PET and fMRI studies. The signal detected by both PET and fMRI depends upon a vascular change in the small arterioles of the cerebral circulation. It is unclear to what extent aging impairs that hemodynamic reactivity and affects the activation signal. Furthermore, pathological changes, such as small, clinically silent infarcts of cerebral tissue, could have significant effects on the ability to detect activation in the aging brain. The atrophy of cerebral tissue with aging presents additional problems for functional neuroimaging, sometimes referred to as “partial volume” effects. In short, the amount of gray matter in a given volume of an acquired image will be less with aging, which could cause an apparent loss of signal. Although a study by Johnson and colleaguesCitation73 demonstrated that activation magnitude did not correlate with cerebral atrophy, loss of tissue may still obscure compensatory increases in activation for aging brains.
Second, the patterns of dedifferentiation described earlier can make brain activations associated with aging hard to interpret. If one thinks of the pattern of brain activations for each subject as a color transparency, and a final group brain image as a combined stack of the transparencies overlaid on one another, then an obvious problem emerges. For group analyses to yield significant results, brain signals need to be fairly robust and focused so that when individual subject's activations are “stacked” on top of one another, bright focal activations are readily apparent. If older adults' brains are organized somewhat idiosyncratically compared with young brains due to differences in experiences, disease states, and other neural insults, older adults will evidence more varied patterns of activation than young adults when performing cognitive tasks. Thus, when brain “transparencies” from older subjects are aggregated or stacked to form a single set of group activations, the resulting image will show many different activation sites of relatively low intensity due to the individual differences in activation patterns. Frequently, many of those sites will not reach significance and a low signal, will appear with diffuse activations occurring as a function of age, when, in fact, it may be that each older adult is showing a different pattern of robust activations. In general, group averages may be more misleading for older adults compared with young adults.
A third important issue is the interpretation of differences in activations between young and old adults. If one finds some type of dedifferentiation for old compared with young adults (eg, bilateral activation, or substitution in old), is this pattern evidence of neural health or pathology? Finally, it is hard to interpret brain activations if the performance on the cognitive tasks differs between young and old. If older adults are performing more poorly, perhaps that, is why there is evidence for less activation, and the cause/effect relationships between poor performance and unusual activations can be difficult to untangle. CabezaCitation48 provides a complete discussion of the many methodological issues to consider in understanding the implications of different patterns of brain activations between young and old adults.
Brain-behavior integration
In this final section, we provide a brief integration of the behavioral and imaging findings that we have discussed thus far, along with unanswered questions that occur when these domains are connected. We hope that this integrative exercise will point to areas where strong linkage between behavioral and imaging work exists, and other areas where development is needed, or even where behavioral and brain studies are inconsistent with one another. Table I connects major findings in cognitive aging to possible neural underpinnings, and Table II summarizes major cognitive neuroscience findings associated with aging and provides potential linkage to the behavioral literature in cognitive aging.
Table I Proposed and known connections between cognitive aging phenomena and neural mechanisms.
Table II Connections between neural findings and behavioral data in cognitive aging.
Behavioral findings
Single-mechanism views of decline
Speed of processing can account, for nearly all agerelated variance on cognitive tasks, and so it is important, to understand its neural connection. The neural substrate for age-related slowing, however, is not well specified. There has been some suggestion that slowing is due to a decrease in dopamine receptors (sec a review by Prull et alCitation55), demyelination, and white matter loss (see RazCitation41 for a review) or to increased dendritic projections that result in circuitous neural processing.Citation74 There is some evidence suggesting that dopamine receptors may play an important role in accounting for agerelated declines in perceptual speed.Citation75,Citation76 Both Backman et alCitation75 and Volkow et alCitation76 reported very substantial correlations between speed and dopamine receptor binding. Virtually no additional variance in speed could be explained when age was added into this relationship, suggesting that dopamine receptor binding is a substantially better predictor of slowing than age. These studies are correlational, and have small numbers of subjects with large age distributions, and so much larger numbers of subjects must be tested to address the reliability of this potentially important causal relationship.
Besides the dopamine receptor studies, there is an additional study that examined the relationship of slowing to activation patterns. Rypma and D'EspositoCitation61 found that, on a working memory task, the fastest old adults showed the most brain activation, whereas the fastest young adults showed the least, brain activity in the dorsolateral prefrontal cortex. They speculate that the reasons for this finding could be a shifting of the relationship between neural activation and optimal response discriminability for the old, or that age-related cortical atrophy mediated the high activation for faster older adults and that time on task may have mediated activations for slower young adults.
Ultimately, a convincing demonstration of the causes of age-related decline in speed of processing may require a large individual difference study where extreme ends of a distribution of fast, and slow older adults are studied with age tightly controlled (eg, study a single decade from 60 to 69 or 70 to 79 years, so that age is not an additional source of variance), and multiple measures of neural functioning using many techniques (including dopamine D2 receptors) are collected. Hopefully, from this approach, some important dissociations will be found that will permit an understanding of the neurobio logical basis for the ubiquitous finding that speed of processing declines with age. It is the authors' guess that behavioral speed represents an aggregate of many different cellular, structural, and functional changes that, occur in the brain with age, and that the particular nature of the changes that have aggregated to result, in slowing could vary considerably from individual to individual. We believe that a focus on a single mechanism to find the underlying cause of age-related decline in speed will be less successful than a “multiple cause” approach to understanding the indices of neural health with age. The “common cause” view that, sensory function provides a quick measure of global neural healthCitation6 may be even more difficult to isolate. There is evidence that there is a decreased amplitude of the hemodynamic response in visual cortex with age, although the summation properties of the hemodynamic response do not differ across age.Citation77,Citation78 On the other hand, the sensory cortex appears to be more resistant to the age-related volumetric decreases compared with other areas of the brain,Citation41 which is hard to reconcile with the sensory function hypothesis. It, is also difficult to reconcile the simplicity of the behavioral hypothesis with the complexity and distribution of the neuronal correlates of sensory function.
In sum, there is a great need for large studies that examine the neurobiological underpinnings of single-mechanism accounts of cognitive aging. It, is likely that, a multitude of accrued cerebral insults that differ widely among individuals could account, for this relationship.
Age-related declines in executive function and long-term memory
Given the volumetric decreases that occur reliably with age in prefrontal cortex, and the finding that shrinkage in this area predicts poor cognitive performance,Citation41 the linkage between behavioral declines in working memory, inhibition, task-switching, and long-term memory and prefrontal dysfunction is well established. The rather straightforward picture becomes more confusing, however, when one examines the relationship of activation patterns in prefrontal cortex to executive function, as there is evidence for increases and decreases in activation in frontal areas with age and evidence for various age-related forms of dedifferentiation, such as contralateral recruitment, unique recruitment, and substitution. Moreover, there is considerable variability across studies on specific areas of frontal activation as a function of age. Nevertheless, there can be little doubt that, much of the decline in executive processes observed in older adults is due to frontal dysfunction.
Another important, distinction between behavioral and ncuroscience research on aging and memory is that cognitive aging researchers tend to refer to speed of processing and working memory as “cognitive resources” accounting for age-related decline.Citation7,Citation27 In contrast, neuroscientists tend to distinguish executive processes that reside in the dorsolateral prefrontal cortexCitation49,Citation79 from the storage aspects of working memory, which have a separate, content-specific status in ventral lateral regions of the frontal cortex.Citation60,Citation79 The segmentation of the frontal cortex by neuroscientists into specific sites associated with inhibition, storage, and conflict, resolution have resulted in changes in the focus of behavioral work away from general explanatory constructs into more focused, operationbased views of age-related changes in cognitive function. Age-related declines in long-term memory have the added complexity of involvement of mediotemporal areas. There is only a weak relationship between mediotemporal volume and cognitive performanceCitation41 and relatively little functional evidence relating hippocampal dysfunction to memory dysfunction in normal older adults - although there arc few studies of this with age.Citation55 Moreover, the entorhinal cortex appears to be important for understanding both normal and pathological aging.Citation80 In general, most evidence accounting for age-related declines in long-term memory function have focused on encoding and retrieval operations residing in the frontal cortex, although there is clear evidence that, mediotemporal function is important for long-term memory function in young adults.
There is certainly sufficient evidence to suggest that prefrontal cortex plays a causal role in contributing to age-related changes in behavioral tasks of executive function and long-term memory. What is poorly understood is the specific relationship of neural activation patterns to cognitive aging, an issue taken up later in this article. Behavioral researchers may find it productive to focus their work in cognitive aging on specific executive processes and operations to understand changes in memory and higher order functions, as these operations better reflect neural architecture.
Continuous cognitive decline across life span
Dopamine receptors decline continuously across the life span and may play a role in the gradual decline of cognitive ability, along with volumetric declines in frontal cortex. Although there is considerable confusion about, patterns of neural recruitment in old and young, there are some broad generalizations that are reasonably well substantiated about the asymmetry of hemispheric recruitment in young compared with older adults.Citation48 What, remains entirely unexplored is how young adults make the transition from highly focused, latcralized activations in performing a cognitive operation, to the qualitatively dedifferentiated patterns of recruitment observed in older adults. The disconnect between continuous behavioral decline and qualitatively different recruitment patterns with age can only be understood by conducting large life span neuroimaging studies with attention to individual differences in both behavioral and neural domains as well as age. To say the least, this is an expensive and daunting task, but fruitful work needs to begin to understand the extant dissociated relationship between life span behaviors and neural activation patterns.
Increase in world knowledge and expertise
While basic cognitive mechanisms are declining, expertise and knowledge show growth. At present, we know very little about whether increased knowledge and experience results in brain reorganization and nothing about how the neural representation of knowledge might change across the life span. Polk and FarahCitation81 demonstrated neural segregation of letters and digits in young adults using fMRI, by showing selective activation for these two stimulus types. Moreover, in a behavioral study, they reported findings strongly suggestive of the notion that experience appears to result in functional reorganization of the brain.Citation82 They found that Canadian postal workers showed a smaller alphanumeric category effect on a visual detection task compared with control subjects. Canadian postal workers regularly sec a cooccurrence of alphanumeric characters together, given that Canadian zip codes contain both letters and numbers, whereas control subjects do not. These findings suggest that extended practice and experience may change neural organization. The understanding of the boundary conditions for how expertise and practice literally change brain organization is minimally understood and thus far, no connections have been made among experience, neural organization, and aging. This represents an important area for future research.
Passive environmental support improves memory
There is compelling behavioral evidence that passive environmental supports (such as inducing deep encoding) can mitigate age differences in memory compared with intentional, self-initiated conditions,Citation83 and there are compelling neural connections to this important, behavioral finding. For example, there is evidence that, although older adults show less recruitment of frontal cortex when intentionally encoding words, activation for older subjects increases relative to young subjects under incidental, deep encoding conditions.Citation84-Citation86
Neural findings
Neural atrophy with age
The finding that neural atrophy is greatest, in the frontal lobes is consistent, with decreases in executive function and long-term memory, but, as mentioned earlier, it is less clear how preserved occipital volume links to the common-cause sensory view of aging. Adding to the mystery is the finding that, despite preserved occipital volume, the amplitude of the hemodynamic response in sensory cortex is decreased for old relative to young, although the hemodynamic response in motor cortex is not as age-sensitive. White matter loss and white matter hyperintensities can be measured with careful analysis of structural images. At present, measuring atrophy in a site-specific manner and relating it to behaviors and neural activation is a labor-intensive task, but, it is an important technical approach in studying cerebral aging.
Neural dedifferentiation with age
The stunning finding that older adults use different parts of their brains to perform cognitive tasks compared with young adults was something that was barely suggested by the behavioral literature on aging. Indeed, neuroimaging cognitive processes in young and older adults has literally changed our understanding of the aging mind. It has been known for decades that, as cognitive tasks become more complex, performance between old and young adults becomes increasingly divergent, but speculation about, this finding typically focused on speed of processing as the underlying mechanism.Citation4 One sentient study on interhemispheric connectivity by Reuter-Lorenz et alCitation87 provided behavioral evidence that the use of two hemispheres to perform a letter-matching task was facilitative for old, but not young. More behavioral studies of this type that focus on linking the recent, neural findings to behavioral phenomena are clearly warranted.
There are a number of issues that, arise in interpreting the different, patterns of brain recruitment between old and young. We do not, know whether the different, forms of dedifferentiation observed in older subjects are compensatory and adaptive, suggesting successful aging, or are a harbinger of neuropathological changes (although there are certainly hints that, they are compensatory, see CabezaCitation48 for a summary). We cannot be sure whether the different patterns reflect strategy differences or functional differences. We do not actually know what young adults are doing with the unrecruited areas activated only by the old. Do young adults have cognitive reserves that they may draw upon when they are ill or stressed, evidencing recruitment, patterns like older adults? We do not understand the role of the environment, nutrition, and toxic substances in altering neurodevelopmental trajectories that may result, in dedifferentiation.
Despite the many unresolved questions as well as problems associated with interpretation of findings, a corpus of work is emerging that, strongly suggests that older adults recruit more symmetrically from the two hemispheres than young adults in frontal areas to perform a range of cognitive tasks. little is known about recruitment patterns in other areas of the brain and this is another important new frontier. Given the infancy of the field, we know quite a lot and there is little question that neural findings are changing the way we think about the behavioral aspects of cognitive aging. Again, to better understand the phenomena of dedifferentiation, large studies, across multiple laboratories, with standardized imaging protocols, life span samples, extensive behavioral testing, and demographic and medical information appear to be the only way to answer many of the questions about brain-behavior relationships in aging.
A second methodological route to better understanding brain-behavior relationships in older adults would involve studying only large samples of elderly, carefully characterizing the samples neuropsychologically, and looking at changes in activations on cognitive tasks as a function of out-of-the-scanner performance. Studying large groups of elderly minimizes some of the issues associated with agerelated differences in hemodynamic responses.
Plasticity
The fact that the brain reorganizes with age and the finding that old rats in enriched environments sprout, neurons suggest that the aging brain can be characterized by plasticity and the potential for experience to enhance function. The prospect for enhancement of brain function in response to experience in late adulthood contrasts considerably with recent conclusions that there are few behavioral gains in cognition that occur as we age.Citation88 There is very substantial evidence that cognitive decline is an unavoidable concomitant of normal aging.Citation5,Citation13 However, although cognitive declines occur for all adults with increasing age, the declines may be steeper when not modulated across the life span by stimulating experience. Once again, neural findings suggest the importance of studying the role of training and rich life experiences on not only maintenance of cognitive function, but, also brain reorganization. This is again a case where a focused study of only older adults could yield important and interpretable findings.
Conclusion
A strong linkage of neural and behavioral research on aging will ultimately lead to an understanding of what is needed to be neurally fit. Only 40 years ago, we had little understanding of the relationship of smoking and cholesterol levels to cardiovascular health. It is likely that, just as there are behavioral routes to healthy bodies, there are behavioral routes to healthy minds. The linkage of studies of brain and behavior across the life span will result, in critical knowledge that will allow individuals to take control of their cognitive future and alter the neurobiological age of their minds. Such an outcome offers huge rewards to both individuals and to our society. A tremendous national investment in the neuroscience of cognitive aging combined with committed effort, and generous cooperation among scholars is needed.
The authors gratefully acknowledge the support of the National Institute on Aging Neuroscience and Neuropsychology Program in preparation of this paper (R01AGO6265-15) awarded to all authors (D. Park, Principal Investigator). Additional support was provided by the National Institute of Deafness and Other Communication Disorders (DC04205) to T. Polk, and by the National Institute of Mental Health (MH01258) to S. Taylor.
REFERENCES
- ParkDC.SchwarzN.edsCognitive Aging: A Primer. Philadelphia, Pa: Psychology Press;2000
- CraikFIM.SalthouseTA.edsThe Handbook of Aging and Cognition. 2nd ed. Mahwah, NJ: Lawrence Erlbaum Associates;2000
- SalthouseTA.Theoretical Perspectives on Cognitive Aging. Hillsdale, NJ: Erlbaum;1991
- SalthouseTA.The processing-speed theory of adult age differences in cognition.Psychol Rev.19961034034288759042
- BaltesPB.LindenbergerU.Emergence of a powerful connection between sensory and cognitive functions across the adult life span: a new window to the study of cognitive aging?Psychol Aging.19971212219100264
- LindenbergerU.BaltesPB.Sensory functioning and intelligence in old age: a strong connection.Psychol Aging.199493393557999320
- Park DC The basic mechanisms accounting for age-related decline in cognitive function. In: Park DC, Schwarz N, eds.Cognitive Aging: A Primer. Philadelphia, Pa: Psychology Press;2000322
- ZacksRT.HasherL.LiKZH.Human memory. In: Craik FIM, Salthouse TA, eds.The Handbook of Aging and Cognition. 2nd ed. Mahwah, NJ: Lawrence Erlbaum Associates;2000293358
- BaddeleyA.Working Memory. Oxford, UK: Clarendon Press;1986
- EngleRW.TuholskiSW.LaughlinJE.ConwayARA.Working memory, short-term memory, and general fluid intelligence: a latent-variable approach.J Exp Psychol Gen.199912830933110513398
- ShahP.MiyakeA.The separability of working memory resources for spatial thinking and language processing: an individual differences approach.J Exp Psychol Gen.19961254278851737
- ParkDC.SmithAD.LautenschlagerG.et al.Mediators of long-term memory performance across the life span.Psychol Aging.1996116216379000294
- ParkDC.DavidsonNS.LautensclagerG.SmithAD.SmithPK.HeddenT.Models of visuospatial and verbal working memory across the adult life span.Psychol Aging. In press.
- HasherL.ZacksRT.Working memory, comprehension, and aging: a review and a new view. In: Bower GH, ed.The Psychology of Learning and Motivation. San Diego, Calif: Academic Press;1988193225
- HeddenT.ParkDC.Aging and interference in verbal working memory.Psychol Aging. In press.
- MayCP.HasherL.KaneMJ.The role of interference in memory span.Mem Cognit.199927759767
- ZacksR.HasherL.Directed ignoring: Inhibitory regulation of working memory. In: Dagenback D, Carr TH, eds.inhibitory Processes in Attention, Memory and Language. San Diego, Calif: Academic Press;1994241264
- ZacksR.HasherL.Cognitive gerontology and attentional inhibition: a reply to Burke and McDowd.J Gerontol B Psychol Sci Soc Sci.199752P274P2839403516
- KramerAF.HahnS.GopherD.Task coordination and aging: explorations of executive control processes in the task switching paradigm.Acta Psychol.1999101339378
- KrayJ.LindenbergerU.Adult age differences in task switching.Psychol Aging.20001512614710755295
- MayrU.Age differences in the selection of mental sets: the role of inhibition, stimulus ambiguity, and response-set overlap.Psychol Aging.2001169610911302371
- JohnsonMK.DeLeonardisDM.HashtroudiS.FergusonSA.Aging and single versus multiple cues in source monitoring.Psychol Aging.1995105075178749578
- SpencerWD.RazN.Memory for facts, source, and context: can frontal lobe dysfunction explain age-related differences?Psychol Aging.199491491598185862
- ChalfonteBL.JohnsonMK.Feature memory and binding in young and older adults.Mem Cognit.199624403416
- ParkDC.PuglisiJT.Older adults' memory for the color of pictures and words.J Gerontol.1985401982043973361
- ParkDC.PuglisiJT.LutzR.Spatial memory in older adults: effects of intentionality.J Gerontol.1982373303357069157
- ParkDC.HeddenT.Working memory and aging. In: Naveh-Benjamin M, Moscovitch M, Roediger HL III, eds.Perspectives on Human Memory and Cognitive Aging: Essays in Honour of Fergus Craik. East Sussex, UK: Psychology Press. In press.
- ParkDC.PuglisiJT.SmithAD.Memory for pictures: does an age-related decline exist?Psychol Aging.1986111173267373
- LaVoieD.LightLL.Adult age differences in repetition priming. A metaanalysis.Psychol Aging.19944539553
- LightLL.LaVoieD.Direct and indirect measures of memory in old age. In: Graf P, Masson MEJ, eds.Implicit Memory: New Directions in Cognition, Development, and Neuropsychology. Hillsdale, NJ: Lawrence Earlbaum Associates;1993207230
- ParkDC.ShawRJ.Effect of environmental support on implicit and explicit memory in younger and older adults.Psychol Aging.199276326421466832
- ParkDC.SmithAD.DudleyWN.LafronzaV.The effects of age and a divided attention task presented during encoding and retrieval on memory.J Exp Psychol Learn Mem Cognit.198915118511912530311
- AndersonND.CraikFIM.Naveh-BenjaminM.The attentional demands of encoding and retrieval in younger and older adults: I. Evidence from divided attention costs.Psychol Aging.1998134054239793117
- CraikFIM.GovoniR.Naveh-BenjaminM.AndersonND.The effects of divided attention on encoding and retrieval processes in human memory.J Exp Psychol Gen.19961251591808683192
- LightL.The organization of memory in old age. In: Craik FIM, Salthouse TA, eds.The Handbook of Aging and Cognition. Hillsdale, NJ: Lawrence Erlbaum Associates;1992111165
- HowardDV.McAndrewsMP.LasagaMl.Semantic priming of lexical decisions in young and old adults.J Gerontol.1981367077147299087
- LightLL.Memory and aging: four hypotheses in search of data.Annu Rev Psychol.1991423333762018397
- CraikFIM.A functional account of age differences in memory. In: Klix F, Hagendorf H, eds.Human Memory and Cognitive Capabilities, Mechanisms, and Performances. Amsterdam, The Netherlands: Elsevier;1986409422
- CherryKE.ParkDC.FrieskeDA.SmithAD.Verbal and pictorial elaborations enhance memory in young and older adults.Aging, Neuropsychol Cognit.199631529
- ParkDC.SmithAD.MorrellRW.PuglisiJT.DudleyWN.Effects of contextual integration on recall of pictures by older adults.J Gerontol B Psychol Sci Soc Sci.1990455257
- RazN.Aging of the brain and its impact on cognitive performance: Integration of structural and functional findings. In: Craik FIM, Salthouse TA, eds.The Handbook of Aging and Cognition. 2nd ed. Mahwah, N: Lawrence Erlbaum Associates;2000190
- KemperTL.Neuroanatomical and neuropathogical changes during aging and in dementia. In: Albert ML, Knoepfel EJE, eds.Clinical Neurology of Aging. 2nd ed. New York, NY: Oxford University Press;1994367
- Gunning-DixonFM.RazN.The cognitive correlates of white matter abnormalities in normal aging: a quantitative review.Neuropsychology.20001422423210791862
- SeernanP.BzowejNH.GuanHC.et al.Human brain dopamine receptors in children and aging adults.Synapse.198713994043505371
- VolkowND.WangGJ.FowlerJS.et al.Measuring age-related changes in dopamine D2 receptors with 11C-raclopride and 18F-N-methylspiroperidol.Psychiatry Res.19966711168797238
- FreyKA.BohnenNI.MeyerP.et al.Presynaptic monoaminergic vesicles in normal aging studied by (+)-[C-1 1]DTBZ PET.J Nucl Med.199839S15
- VolkowND.LoganJ.FowlerJS.et al.Association between age-related decline in brain dopamine activity and impairment in frontal and cingulate metabolism.AM J Psychiatry.2000157758010618016
- CabezaR.Functional neuroimaging of cognitive aging. In: Cabeza R, Kingstone A, eds.Handbook of Functional Neuroimaging of Cognition. Cambridge, Mass: MIT Press;2001
- Reuter-LorenzPA.JonidesJ.SmithEE.et al.Age differences in the frontal lateralization of verbal and spatial working memory revealed by P ET.J Cognit Neurosci.200017418710769314
- McintoshAR.SekulerAB.PenpeciC.et al.Recruitment of unique neural systems to support visual memory in normal aging.Curr Biol.199991275127810556091
- HazlettEA.BuchsbaumMS.MohsRC.et al.Age-related shift in brain region activity during successful memory performance.Neurobiol Aging.1998194374459880046
- BucknerRL.LoganJM.Frontal cotributions to episodic memory encoding in the young and elderly. In: Parker AE, Wilding AE, Bussey T, eds.The Cognitive Neuroscience of Memory Encoding and Retrieval. Philadelphia, Pa: Psychology Press. In press.
- GradyCL.Brain imaging and age-related changes in cognition.Exp Gerontol.1998336616739951614
- GradyCL.CraikFIM.Changes in memory processing with age.Curr Opin Neurobioi.200010224231
- PrullMW.GabrieliJDE.BungeSA.Age-related changes in memory: a cognitive neuroscience perspective. In: Craik FIM, Salthouse TA, eds.The Handbook of Aging and Cognition. 2nd ed. Mahwah, NJ: Lawrence Erlbaum Associates;200091154
- Reuter-LorenzPA.MarshuetzC.JonidesJ.SmithEE.HartleyA.KoeppeR.Neurocognitive ageing of storage and executive processes.Eur J Cog Psychol.200113257278
- D'EspositoM.AguirreGK.ZarahnE.BallardD.ShinRK.LeaseJ.Functional MRI studies of spatial and nonspatial working memory.Cognit Brain Res.19987113
- OwenAM.The functional organization of working memory processes within human lateral frontal cortex: the contribution of functional neuroimaging.Eur J Neurosci.19979132913399240390
- SmithEE.GevaA.JonidesJ.MillerA.Reuter-LorenzP.KoeppeRA.The neural basis of task switching in working memory: effects of performance and aging.Proc Natl Acad Sci U S A.2001982095210011172081
- SmithEE.JonidesJ.Storage and executive processes in the frontal lobes.Science.19992831657166110073923
- RypmaB.D'EspositoM.Isolating the neural mechanisms of age-related changes in human working memory.Nat Neurosci.2000350951510769393
- JonidesJ.MarshuetzC.SmithEE.Reuter-LorenzPA.KoeppeRA.HartleyA.Age differences in behavior and PET activation reveal differences in interference resolution in verbal working memory.J Cognit Neurosci.20001218819610769315
- MitchellKJ.JohnsonMK.RayeCL.D'EspositoM.fMRI evidence of agerelated hippocampal dysfunction in feature binding in working memory.Cognit Brain Res.200010197206
- TulvingE.KapurS.CraikFIM.MoscovitchM.HouleS.Hemispheric encoding/retrieval in episodic memory: positron emission tomography findings.Proc Natl Acad Sci U S A.199491201620208134342
- BrewerJB.ZhaoZ.DesmondJE.GloverGH.GabrieliJD.Making memories: brain activity that predicts how well visual experience will be remembered.Science.1998281118511879712581
- WagnerAD.SchacterDL.RotteM.et al.Building memories: remembering and forgetting of verbal experiences as predicted by brain activity.Science.1998281118811919712582
- CabezaR.GradyCL.NybergL.et al.Age-related differences in neural activity during memory encoding and retrieval: a positron emission tomography study.J Neurosci.1997173914008987764
- GradyCL.McintoshAR.HorwitzB.et al.Age-related reductions in human recognition memory due to impaired encoding.Science.19952692182217618082
- SchacterDL.SavageCR.AlpertNM.RauchSL.AlbertMS.The role of hippocampus and frontal cortex in age-related memory changes: a PET study.Neuroreport.19967116511698817525
- MaddenDJ.TurkingtonTG.ProvenzaleJM.et al.Adult age differences in the functional neuroanatomy of verbal recognition memory.Hum Brain Mapp.199971151359950069
- MaddenDJ.TurkingtonTG.ProvenzaleJM.HawkTC.HoffmanJM.ColemanRE.Selective and divided visual attention: Age-related changes in regional cerebral blood flow measured by H2 15O PET.Hum Brain Mapp.1997538940920408243
- KempermannG.KuhnHG.GageFH.Experience-induced neurogenesis in the senescent dentate gyrus.J Neurosci.199818320632129547229
- JohnsonSC.SaykinAJ.BaxterLC.et al.The relationship between fMRI activation and cerebral atrophy: comparison of normal aging and Alzheimer's disease.Neuroimage.20001117918710694460
- SalthouseTA.Steps toward the explanation of adult age differences in cognition. In: Perfect T, Maylor E, eds.Theoretical Debate in Cognitive Aging. London, UK: Oxford University Press;2000
- BackmanL.GinovartN.DixonRA.et al.Age-related cognitive deficits mediated by changes in the striatal dopamine system.Am J Psychiatry.200015763563710739428
- VolkowND.GurRC.WangG.et al.Association between decline in brain dopamine activity with age and cognitive and motor impairment in healthy individuals.Am J Psychiatry.19981553443499501743
- BucknerRL.SnyderAL.RaichleME.MorrisJC.Functional brain imaging of young, nondemented, and demented older adults.J Cognit Neurosci.200012(suppl)243411506645
- RossMH.Yurgelun-ToddDA.RenshawPF.et al.Age-related reduction in functional MRI response to photic stimulation.Neurology1997481731769008514
- D'EspositoM.AguirreGK.ZarahnE.BallardD.ShinRK.LeaseJ.Functional MRI studies of spatial and nonspatial working memory.Cognit Brain Res.19987113
- MorrisonJH.Age-related shifts in neural circuit characteristics and their impact on age-related cognitive impairments. In: Stern PC, Carstensen LL, eds.The Aging Mind: Opportunities in Cognitive Research. Washington, DC: National Academy Press;200083113
- PolkTA.FarahMJ.The neural development and organization of letter recognition: evidence from functional neuroimaging, computational modeling, and behavioral studies.Proc Natl Acad Sci U S A.1998958478529448250
- PolkTA.FarahMJ.Late experience alters vision.Nature.19953766486497651514
- CraikFIM.JenningsJM.Human memory. In: Craik FIM, Salthouse TA, eds.The Handbook of Aging and Cognition. Hillsdale, NJ: Lawrence Erlbaum;199251110
- LoganJM.BucknerRL.Age-related changes in neural correlates of encoding. Cognitive Neuroscience Society Eighth Annual Meeting. New York, NY, April. Abstract. 2001
- GradyCL.McintoshAR.RajahMN.CraikFIM.Neural correlates of the episodic encoding of pictures and words.Proc Natl Acad Sci USA.199895270327089482951
- GradyCL.McintoshAR.RajahMN.BeigS.CraikFIM.The effects of age on the neural correlates of episodic encoding.Cereb Cortex.2000980581410600999
- Reuter-LorenzPA.StanczakL.MillerA.Neural recruitment and cognitive aging: Two hemispheres are better than one, especially as you age.Psychol Sci.199910494500
- DixonRA.The basic mechanisms accounting for age-related decline in cognitive function. In: Park DC, Schwarz N, eds.Cognitive Aging: A Primer. Philadelphia, Pa: Psychology Press;20002342