Abstract
Recent research has overcome the old paradigms of the brain as an immunologically privileged organ, and of the exclusive role of neurotransmitters and neuropeptides as signal transducers in the central nervous system. Growing evidence suggests that the signal proteins of the immune system - the cytokines - are also involved in modulation of behavior and induction of psychiatric symptoms. This article gives an overview on the nature of cytokines and the proposed mechanisms of immune-to-brain interaction. The role of cytokines in psychiatric symptoms, syndromes, and disorders like sickness behavior, major depression, and schizophrenia are discussed together with recent immunogenetic findings.
La investigación reciente ha sobrepasado los antiguos paradigmas que consideraban que el cerebro era un órgano inmunológicamente privilegiado y que los neurotransmisores y neuropéptidos tenían un papel exclusivo en la transducción de señales en el sistema nervioso central. Existe creciente evidencia que sugiere que las proteínas de señales del sistema inmune - las citoquinas - también participan en la modulación de la conducta y en la inducción de síntomas psiquiátricos. Estre artículo entrega una visión panorámica acerca de la naturaleza de las citoquinas y los mecanismos propuestos de la interacción entre la inmunidad y el cerebro. Se discute el papel de las citoquinas en síntomas, síndromes y trastornos psiquiátricos como la conducta de enfermedad, la depresión mayor y la esquizofrenia, como también recientes hallazgos inmunogenéticos.
La recherche récente a dépassé les anciens paradigmes considérant le cerveau comme un organe immunologiquement privilégié et le rôle exclusif des neurotransmetteurs et des neuropeptides comme transducteurs de signaux dans le système nerveux central. De plus en plus d'arguments suggèrent que les protéines de signal du système immunitaire - les cytokines - sont aussi impliquées dans la modulation du comportement et le développement des symptômes psychiatriques. Cet article tente de donner un rapide aperçu de la nature des cytokines et des mécanismes supposés mis en jeu dans l'interaction entre immunité et cerveau. Sont présentés le rôle des cytokines dans les symptômes psychiatriques, syndromes et troubles tels que le «comportement de maladie», la dépression majeure et la schizophrénie ainsi que les découvertes récentes en immunogénétique.
A short history of PNI
The first evidence suggesting an interaction between the brain and the immune system came from studies performed 80 years ago by Russian investigators. Derived directly from a Pavlovian perspective on the conditioning of behavioral and physiological responses, a conditioned stimulus (CS) was repeatedly paired with injections of foreign proteins. Subsequent exposure to the CS, alone, was purported to induce antibody production in addition to a conditioned increase in a variety of non-specific defense responses.Citation1 As this research attracted very little attention outside the Soviet Union, the commonly accepted beginning of research in the field of psycho-neuroimmunology (PNI) is associated with the experiments of Ader, who was studying taste aversion conditioning in rats in the 1970s. Conditioned animals that were reexposed to a CS, previously paired with the immunosuppressive effects of cyclophosphamide showed an attenuated antibody response to sheep red blood cells.Citation2 The results of these initial experiments demonstrated that the immune system was subject to classical conditioning.
A more complex research direction in the field of PNI was the study of behavioral influences on immunity, starting in the 1950s with the research on stress and infectious disease.Citation3 During the 1970s, Besedovsky was beginning to systematically investigate the neuroendocrine-immune system network with his studies on the effects of immune responses on neural and endocrine function.Citation4 Felten described the direct contact of noradrenergic sympathetic nerve fibers with lymphocytes and macrophages.Citation5 He showed that these nerve fibers were localized in specific compartments of lymphoid organs, forming close, synaptic-like neuroeffector junctions with T lymphocytes and macrophages.Citation6 These “hard-wired” connections between the brain and the immune system have since been shown to be a major route for behavioral and central cytokine influences on immune function. They are, thus, a cornerstone for a mechanistic understanding of the signaling between the nervous system and immune system.
All these investigations demonstrated the influence of the central nervous system (CNS) on the immune function. However, this is only half the truth, as the brain-immune interaction is bidirectional. The old paradigm of the brain as an immunologically privileged organ may have inhibited the research of the immune system's action on brain and behavior. Meanwhile, it is commonly accepted that immune cells enter the brain even under normal, nonpathological conditions, and that all kinds of brain cells - neurons, glial and endothelial cells - are sensitive to the transmitters of the immune system: the cytokines.
Direct evidence for the neural activities of cytokines was first obtained after injections of various cytokines systemically or into the cerebral ventricles (intracerebroventricular [ICV]). These studies established that cytokines can activate the hypothalamus-pituitary-adrenal (HPA) axis,Citation7,Citation9 induce fever,Citation10 prolong slow-wave sleep,Citation11 reduce foodCitation12 and water intake,Citation13 and decrease motilityCitation14 These effects were evident not only in experimental animal, but also in humans who received cytokine injections for cancer treatment.Citation15,Citation16 The most tested cytokine with regard to brain-immune interactions is interleukin-1 (IL-1), although other cytokines such as the tumor necrosis factor alpha (TNF-α),Citation17,Citation18 interferon (IFN),Citation19,Citation20 IL-2,Citation21, IL-6,Citation22 and IL-12Citation23 can all induce one or several of the above responses.
A large number of studies have investigated the ways in which the cytokines influence brain function. Although there are still a lot of open questions, the following sections will try to give a short overview of the current knowledge of this part of PNI.
Cytokines
Nomenclature
The term cytokine defines a large group of nonenzymic proteins that act as hormones. Their actions, as well as their target cell populations, are both diverse and overlapping. Once released into the environment, cytokines traverse small distances to ligate their high-affinity receptors via either autocrine or paracrine fashion. Upon ligation of the cytokine receptor, a signaling cascade is triggered resulting in an alteration in gene transcription by the target cell.
The pleiotropic features of cytokines have led to a confusing nomenclature because they were frequently named for their biological activity. It therefore repeatedly happened that a single cytokine was described by several names. Another problem in the nomenclature of cytokines emerged from their redundancy of action. In the 1970s, two terms were introduced: the term cytokine defined the large group of protein transmitters of the immune system. The term interleukin tried to list all known cytokines in numerical order. At the time this article was written, the list of ILs ended at IL-27.Citation24 Unfortunately, some historical groups like the IFNs, TNF, lymphotoxins (LT), transforming growth factor beta (TGFβ), leukocyte inhibitory factor (LIF), and most of the colony stimulating factors (CSFs) remained to be known by their old names.Citation25 This is particularly confusing, for example, the IFNs are divided into two totally different groups: type I interferons (IFN-α, IFN-β and others) and type II interferon IFN-γ. On the other hand, the new group of chemokines was defined after IL-8 and several related cytokines were determined to form a distinct cytokine family.
According to the complexity and diversity of the cytokines, there are several possibilities for grouping them. They can be grouped by structural similarities, clustered chromosomal localization of their encoding genes, or principally similar functional features.
Structure
The typical cytokine is a glycosylated monomeric peptide of about 150 amino acids. Others are homodimers (eg, IL-5, M-CSF) or homotrimers (TNF-α and LT-α), or heterodimers (IL-12) or heterotrimers (LT-α/β).Citation26 Three-dimensional structure studies have shown that many otherwise nonhomologous cytokines adopt similar conformations. These structural features of some cytokines permit their grouping into families. Members of the large IL-2/IL-4 family (including IL-2 to IL-7, IL-9, IL-11, IL-12 p35, IL-13, IL-15, type I and type II IFNs, and CSFs) share a common tertiary architecture characterized by bundles of four antiparallel α-helices in a spatially similar arrangement.Citation26-Citation28
Two important cytokine families show distinct structures:
The IL-1 family, consisting of IL-1α, IL-1β, IL-1 receptor antagonist (IL-1ra), and IL-18, is characterized by a β-trefoil structure.Citation26 IL-1α and IL-1β exert identical actions via binding to a single 80-kDa cell surface receptor (IL-1RI) and an accessory protein (AcP).Citation29 IL-1β and IL-18 are formed as biologically inactive precursors that are cleaved by the enzyme ICE (caspase 1).Citation30 IL-1ra is a highly specific, competitive antagonist of IL-1RI, blocking all actions of IL-1 by inhibiting the association between IL-1RI and AcPCitation30 Four other members of this family have recently been identified, but their biological activity - especially with regard to their actions on the CNS - remains to be elucidated.Citation31
Another structurally similar cytokine family is that of the TNFs including TNF-α, TNF-β, LT-β, Fas ligand (CD90L), CD40 ligand, TNF-related apoptosis-inducing ligand (TRAIL), and several other TNF ligand superf amily members.Citation32 The characteristic structure of this family is a β-jellyrollCitation26 Members of the TNF family act as trimers, most of which are membrane-bound and so are quite distinct in their properties from the other cytokines.Citation33
Clustered chromosomal localization of cytokines
The cytokines are not members of a single gene superfamily. Remarkably few similarities have been noted in their primary nucleotide or amino acid sequences, and their genes are, for the most part, scattered throughout the genome. However, some chromosomal regions where cytokine coding genes are clustered are known. Most interestingly, some of these chromosomal regions seem to be associated with psychiatric disorders, especially schizophrenia.
Chromosome 1
One cluster of genes coding for members of the IL-10 family is located on chromosomal regions 1q32. These are the cytokines IL-10, IL-19, IL-20, and IL-24.Citation34 This region is of major interest in genetic schizophrenia research, as several linkage studies identified a susceptibility locus for schizophrenia there.Citation35-Citation37 Indeed, a recent study points to the IL-10 gene itself as a susceptibility gene for schizophrenia.Citation38
Chromosome 2
With exception of IL-18, the members of the IL-1 family are encoded by closely linked genes on the long arm of chromosome 2.Citation31 An association of polymorphisms in the genes coding for IL-1α, IL-1β, and IL-1ra with schizophrenia was reported by Katila and colleagues.Citation39 Although this finding was not replicated, another group described bifrontal temporal gray matter volume deficits and generalized white matter tissue deficits in schizophrenia patients who were carriers of a distinct polymorphism in the IL-1β gene.Citation40
Chromosome 5
The cytokine gene cluster on chromosomal region 5q23-35 contains genes coding for IL-3, IL-4, IL-5, IL-9, IL-12 p40, IL-13, GM-CSF, and others.Citation26 This is of particular interest, as linkage studies pointed to a possible susceptibility locus for schizophrenia in this chromosomal region.Citation41 The same region was recently identified as containing the susceptibility gene for Crohn's disease.Citation42
Chromosome 6
The genes coding for three members of the TNF family, TNF-α, TNF-β, and LT-β, are located in an immunologically important region: the HLA-III region, which is embedded between the HLA-II and the HLA-I region on the short arm of chromosome 6. Again, genome scans have indicated a linkage of this chromosomal region with schizophrenia.Citation43 Boin and colleagues reported the association of a functional single nucleotide polymorphism in the TNF-α gene with schizophrenia,Citation44 but a replication study performed by our group could not confirm this finding.Citation45
Chromosome 9
The type I IFNs are encoded on chromosomal region 9p22. This cluster contains about 15 closely linked functional IFN-α and IFN-ω genes in addition to a single IFN-β gene.Citation46 All members of the IFN-α/β family (IFN-α, IFN-β, IFN-ω, and IFN-τ) show at least 30% homology in their amino acid sequence.Citation25 No conclusive data are available indicating a susceptibility gene for a psychiatric disorder in this chromosomal region.
Functional concepts
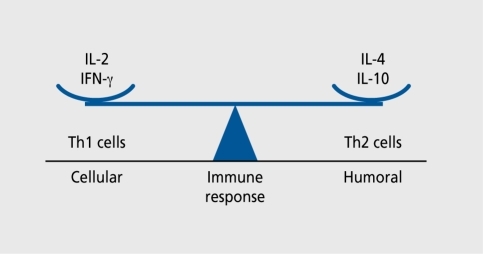
Cytokines are most commonly grouped by their functional similarities, although this kind of categorization is highly arbitrary with regard to their pleiotropy. One of the most prominent concepts used to discriminate two distinct ways that the specific immune system can react on environmental stimuli is the classification of T helper 1 (Th1) and T helper 2 (Th2) cell diversity. This classification is based on the cytokine production patterns of T helper cells and reflects the polarization of the immune answer to either a cell-mediated (Thl) or a humoral (Th2) immune response.Citation33
Th1 cells mainly produce IFN-γ, IL-2, IL-12, IL-18, and TNF-β, while Th2 cells principally secrete IL-4, IL-5, IL-6, IL-10, IL-13, and TGFβ. TNF-α and IL-10 are commonly characterized as Th2-like cytokines, although they are synthesized by both Th1 and Th2 cells.Citation47,Citation48 IL-12 and IL-4 are essential for the development of Th1 and Th2, respectively.Citation49 The Th1 system promotes cell-mediated immune responses against intracellular pathogens, whereas the Th2 system helps B cell maturation and promotes humoral immune responses against extracellular pathogens. Th1 and Th2 cytokines antagonize each other in promoting their own type of response, while suppressing the other type of helper cell. Which system will dominate over the other depends on the relative timing and ratio of IL-4 to IFN-γ and IL-12.Citation50-Citation52 may help to understand the balance between Th1 and Th2.
Such a polarized development of T cells happens not only on the peripheral level, but also in the CNS. Although initiation of T-cell responses is unlikely to occur within the CNS, T cells and monocytes will be massively recruited if pathogens are placed into the cerebral ventricles.Citation53 Perivascular macrophages, owing to their location close to the blood-brain barrier (BBB), can stimulate T cells to proliferate and secrete Th1 cytokines.Citation54 Following extravasation into the CNS parenchyma, T cells also interact with intrinsic CNS cells, particularly microglia and astrocytes.Citation53 Microglia progressively acquire a clear-cut macrophage phenotype in response to CNS injuries,Citation55 and can induce the production of Th1 cytokine IL-12Citation56,Citation57 and of Th2 cytokines such as IL-10 and TGFβ.Citation53 Astrocytes are also potential sources of TGFβ, which inhibits MHC II (major histocompatibility complex II) and ICAM-1 (intercellular adhesion molecule 1) expression in macrophage/microglia.Citation58 Microglia and astrocytes also secrete chemokines that may affect the recruitment of Th1 and Th2 cells. In sum, a complex network between microglia, astrocytes, and T cells is involved in the balance between Th1 and Th2 systems, which in turn might have impact on immune responses within the CNS.
How do cytokines act on the CNS?
Five ways for cytokine signals to enter the brain
One of the major reasons why the brain has long been defined as an immunologically privileged organ is the presence of a tight barrier between the brain and the periphery: the BBB. To our current knowledge, there are five pathways via which cytokine signals may cross the BBB.
Transport across the BBB
There is evidence for an active, saturable, and specific transport system for certain cytokines across the BBB. By the use of radiolabeled cytokines in animal experiments, cytokines like IL-1, TNF-α, and IL-2 were demonstrated to be transported across the BBB.Citation59-Citation62 These experiments suggest that active transport plays a significant role in getting cytokines across the BBB. One limitation, however, is that the absorption of labeled cytokines into the brain tissue may not reflect the transport of cytokines across the BBB, but the binding of cytokines to the BBB. Some data show that the majority of intravenously (IV) infused radiolabeled IL-1α can be found on brain endothelial cells,Citation63 or on the surface and pinocytotic vesicles of the brain endothelia shortly after injection.Citation64
Passage of circumventricular organs
The second pathway is that cytokines may affect the C the circumventricular organs (CVOs), which possess a leaky BBB. The CVOs are midline structures bordering the 3rd and 4th ventricles and are the only areas of the brain at are outside the BBB. CVOs are characterized by the small size, high permeability, and fenestrated capillaries. These barrier-deficient areas are recognized as important sites for communicating with the cerebrospinal fluid and between the brain and peripheral organs via blood-borne products. CVOs include the following structuresCitation65,Citation66:
Pineal gland, which is known as the regulatory organ of the circadian rhythm because it produces the hormone melatonin from the amino acid tryptophan.
Median eminence of the hypothalamus, which arises behind the optic chiasma and is continuous with the pituitary stalk; it communicates with the cerebrospinal fluid.
Subfornical organ, which is positioned under the fornix and is one of the “sensory CVOs” responsible for maintaining body fluid balance.
Area postrema (AP), which is a CVO close to the nucleus of the solitary tract, part of the brain-stem bordering the fourth ventricle. The AP is another “sensory CVO” involved in body fluid homeostasis. It is also thought to play a role in emesis and vomiting.
Subcommissural organ, which contacts the third ventricle covering the posterior commissure. It comprises a complex of neurosecretory ependymal cells known to secrete various glycoproteins into the cerebrospinal fluid. The functional significance of these glycoproteins has not yet been determined.
Organum vasculosum of the lamina terminalis (OVLT), which is a CVO close to the hypothalamic thermoregulatory center.
The intermediate and neural lobes of the pituitary are sometimes also cited as CVOs.
Lesions of the OVLT suppressed intraperitoneal (IP) lipopolysaccharide (LPS)-induced feverCitation67,Citation68 and removal of AP-blocked IL-1-induced c-fos expression in the paraventricular nucleus,Citation69 indicating the important role of these CVOs in transmitting the peripheral cytokines into the brain. However, there are also controversial results, showing the opposite effect.Citation70,Citation71 The discrepant results may be attributable to the extent of the lesion and the different doses of LPS and IL-1 used in these studies. Altogether, it seems that low doses LPS and IL-1 may specifically affect the CVOs and high doses of LPS and IL-1 may gain access to CNS at other sites.Citation72
Transmission via the vagus nerve
The third pathway for cytokines to engage the CNS is the vagus nerve. Numerous studies have been published demonstrating the involvement of vagus nerve in peripheral cytokine-induced CNS responses. One of the first observations was that peripheral LPS-induced hyperalgesia can be blocked by vagotomy, indicating that afferent vagal pathways innervate specific regions of the brain as a key connection between peripheral cytokines and the CNS.Citation73 Others reported the role of the vagus nerve in inducing fever,Citation74 activating the HPA axis, depleting norepinephrine in the hypothalamus,Citation75 prolonging slow- wave sleep,Citation76 and suppressing food-motivated behavior.Citation77 Thus, there is major evidence that vagal afferents are important for conveying signals generated from IP injection of low doses of LPS. The role of vagal afferents is more important for the behavioral depression that develops in response to peripheral immune stimuli than for the induction of fever and activation of the HPA axis.Citation78
De novo synthesis by BBB cells
The fourth pathway is that peripheral immune stimuli may induce the production of cytokines by cells of the BBB, which then secret cytokines into the brain parenchyma. In situ hybridization studies showed that the cells of the BBB respond to peripheral immune stimulation by producing IL-1,Citation79 IL-6,Citation80 and TNF-α.Citation81 Thus, during systemic immune challenge, production of cytokines by cells of the BBB may result in widespread cytokine activity in the entire CNS. This is consistent with a report that the IL-1 bioactivity can be found in all brain regions after high-dose peripheral LPS injection.Citation82
Local action of cytokines at many brain sites may actually be mediated via the receptors on endothelial cells. This binding induces another important effect of peripheral cytokines on cells of the BBB: the induction of cyclooxygenase-2 (COX-2), a rate-limiting enzyme of prostaglandin synthesis. Predominantly IL-1 and TNF-α induce the expression of COX-2 in endothelial cells of the BBB.Citation83,Citation84
As many cytokine-induced CNS effects can be blocked by COX inhibitors,Citation85,Citation86 the cytokine-induced COX-2 activity in BBB cells may represent a central mechanism of cytokine-CNS interaction.
Infiltrating leukocytes
Finally, it has to be considered that cytokines may enter the brain via infiltrating leukocytes. It has long been known that leukocytes may enter the brain under both normal and pathological conditions.Citation87 In normal brain, scattered and random crossing of the BBB by leukocytes provide immune surveillance for the CNS.Citation88 Under pathological conditions such as bacterial meningitis, activated leukocytes expressing inflammatory cytokines may infiltrate the brain.Citation89,Citation90 Additionally, CNS action of cytokines may weaken the BBB, promoting an increased infiltration of cytokine producing leukocytes.
Summary
The above mechanisms for the entrance of cytokines into the brain highlight the limitations of measuring peripheral levels of cytokines in neuropsychiatrie disorders. Cytokine levels in the blood (ie, serum or plasma) may reflect the systemic immune status and have been established as useful clinical markers in septic shock, inflammatory disorders, or cancers,Citation91 but cannot conclusively clarify the cytokine expression within the CNS.
Cytokine and cytokine receptor expression in the brain
The specificity of the response to cytokines is provided by their receptors. Thus the expression of cytokine receptors is necessary for signal transmission of the cytokines entering the brain. Table 1 lists some selected cytokines and their receptors that are expressed within the CNS.
Table I Expression of some selected cytokines and their receptors on neurons and glial cells. Th, T helper cell; C, cytokine; R, receptor; IFN, interferon; TNF-α, tumor necrosis factor alpha; IL, interleukin; AIDS, acquired immune deficiency syndrome; CNS, central nervous system; HPA, hypothalamus-pituitary-adrenal; BBB, blood-brain barrier; ACTH, adrenocorticotropic hormone.
Cytokine-neurotransmitter interactions
As the known effects of cytokines on the brain physiology are extremely numerous and complex, we will just give a few examples.
The effect of IFNs on neurons starts very early during brain development, where they regulate neuronal migration and differentiation.Citation92 In vitro and in vivo studies showed the modulating effect of IFNs on the production of prolactinCitation93 and - of particular interest regarding psychopathology - on the catecholaminergic, dopaminergic, serotonergic, and glutamatergic neurotransmitter systems, eg, the induction of transcriptional activity of the serotonin (5-hydroxytryptamine [5-HT]) transporter.Citation94-Citation97
TNF-α regulates the secretion of norepinephrine in the brain.Citation98 Peripheral administration of TNF-α induces the cerebral tryptophan contentCitation99 and the synthesis of 5-HT and dopamine.Citation100
There is experimental evidence that IL-1 can activate the 5-HT transporter thereby increasing the reuptake of 5-HT from the synaptic cleft.Citation101 Furthermore, IL-1 enhances nonrapid eye movement (NREM) sleep and activates the serotonergic system.Citation102 It has also been reported that the NREM sleep enhancement induced by IL-1 is partially inhibited by brain 5-HT depletion,Citation103 suggesting that this IL-1 effect is partly mediated by the serotonergic system. Observations that the biological activities of IL-1 and 5-HT overlap to a large extent suggest that interactions between these two systems may be relevant to the manifestation of behavior under a variety of conditions. In contrast to IL-1, the Th2 cytokine IL-10 reduces NREM sleep.Citation104
IL-2 can affect gene expression, neuronal activity, and neurotransmitter release in brain regions subserving sleep, memory and cognition, locomotion, and neuroendocrine function. IL-2 modulates the neurotransmission of acetylcholine, dopamine, and norepinephrine in a biphasic manner.Citation105 It appears to be a potent and specific regulator of neurotransmission in frontal cortex, hippocampus, striatum, and hypothalamus.Citation106
IL-6 is produced by neurons, astrocytes, and microglia.Citation107 This cytokine promotes neuronal differentiation and survival,Citation108 and modulates the neurotransmitter systems summarized above.Citation109-Citation111 Several studies have investigated the influence of IL-6 on the production, release, and metabolism of 5-HT. Peripherally administered IL-6 increases the concentrations of tryptophan and the 5-HT metabolite 5-hydroxyindole acetic acid (5-HIAA) in the brain,Citation112-Citation114 and it has been proposed that the interaction between IL-6 and brain 5-HT is a complex process.Citation115
Recent studies have demonstrated a new type of neural activity of cytokines. IL-1 appears to act on neurons in hippocampus and amygdala to inhibit long-term potentiation and weaken synaptic strength.Citation116,Citation117 A contrary effect was demonstrated for TNF-α. Astrocytes continuously release TNF-α to control synaptic strength. The group led by Beattie has demonstrated that TNF-a induces the expression of glutamatergic receptors of the AMPA (α-amino-3-hydroxy-5-methyl-4-isoxazole pro prionate) type on neuronal axons.Citation118 Inhibition of the TNF-α signal by specific antibodies or soluble receptors lead to a reduced AMPA receptor expression. These cytokine activities may be involved in the ability of cytokines to alter the neural processes of learning.Citation119,Citation120
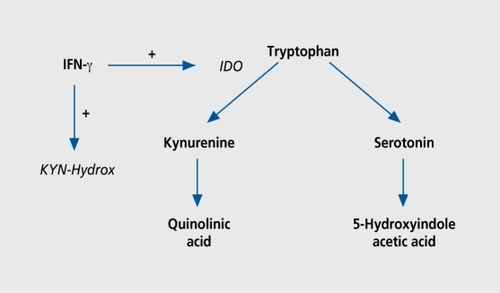
Besides the direct action of cytokines on brain cells, a biochemical link between cytokines and 5-HT is provided by the IFN-γ-controlled tryptophan metabolism. The essential amino acid tryptophan is the precursor of two distinct metabolism pathways, leading to the products 5-HT or kynurenine () The enzyme indoleamine-2,3-dioxygenase (IDO) metabolizes tryptophan to kynurenine, which is then converted to quinolinic acid by the enzyme kynurenine hydroxylase. Both IDO and kynurenine hydroxylase are induced by IFN-γ. The activity of IDO is an important regulatory component in the control of lymphocyte proliferation.Citation121 It induces a halt in the lymphocyte cell cycle due to the catabolism of tryptophan.Citation122 The Th2 cytokines IL-4 and IL-10 inhibit the IFN-β-induced tryptophan catabolism by IDO.Citation123 The enzyme IDO is located in several cell types including monocytes and microglial cells.Citation124 Thus, an IFN-yinduced, IDO-mediated decrease in CNS tryptophan availability may lead to a serotonergic deficiency.
Cytokines and psychiatric symptoms
Sickness behavior
The IL-1 receptor was the first cytokine-related structure detected in the brain.Citation125,Citation126 In 1988, the group led by Dantzer demonstrated the functional relevance of IL-1 and its receptor in the brain: IL-1 injected into the brain of rats induced conditioned taste aversion.Citation127 In the same year, the effect of another cytokine, TNF-α, was introduced, when the group of Plata-Salaman showed the suppressing effect of IL-1 and TNF-α on feeding behavior.Citation128 Later, it was shown that peripherally administered LPS induces the expression of IL-1α, IL-1β, TNF-α, and IL-6 in brain macrophages and microglia.Citation129-Citation131 This effect seems to be a key mechanism in the induction of non-specific sickness symptoms including fever, anorexia, hyperalgesia, and the so-called sickness behavior, which is characterized by weakness, malaise, Hstlessness, cognitive impairment, depressed mood, lethargy, and reduced feeding behavior.Citation132
Behavioral changes during cytokine therapy
The first indication of a role for cytokines in sickness behavior in humans came from clinical trials in which recombinant or purified cytokines were used to treat specific cancers or chronic viral infections such as hepatitis B or C. The syndrome produced by these inflammatory mediators resembled those seen in patients with major depression (MD).Citation133
The first report on the sickness behavior-inducing effect of cytokines was published by Smedley and colleagues, who treated patients with advanced locally recurrent breast cancer with a high dose (160 MU/week) of IFN-α.Citation15 Within 1 h of administration, they observed influenzalike symptoms, which 1 week later were superseded by lethargy, anorexia, and nausea, with a consequent loss of weight in most patients. Other side effects included profound somnolence, confusion, and paresthesia. Low-dose IFN-α therapy (3-5 MU three times a week) induces less severe psychiatric symptoms such as irritability and depression accompanied by impaired concentration, lack of motivation, sleep disturbances, and decreased libido.Citation134 Depressive symptoms induced by IFN-α or IL-2 therapy were described to be related to a decreased tryptophan availability.Citation135
Not only sickness behavior, but also schizophrenia-like symptoms including agitation, cognitive impairment, disorientation, delusions, and hallucinations are induced by IL-2 and IFN-α.Citation136,Citation137 Denicoff and colleagues were the first to report dose- and time-related psychiatric side effects in cancer patients treated with recombinant IL-2 that ranged from brief to severe agitation and combat iveness, requiring antipsychotic therapy.Citation138
Besides the observation in patients suffering from malignancies or chronic inflammatory diseases, experimental data in healthy humans confirmed that cytokines, particularly TNF-α and IL-6, induce depressed mood, anxiety, and memory impairment.Citation139
Major depression
The observations described above led to the hypothesis that sickness behavior may serve as a model for the immune-related pathophysiology of major depression (MD).Citation132 In fact, there is a large body of evidence for an altered immune response in depressed patients.
As described above, IFN-γ is a characteristic marker of Th1 cells. IFN-γ is produced in higher amounts by lymphocytes of patients with MD than by those of healthy controls,Citation140 and higher plasma levels of IFN-γ in depressed patients, accompanied by lower plasma tryptophan availability, were described.Citation141,Citation142 This gives additional evidence for a possible link between the Th1 -like cytokine IFN-γ and the IDO-related reduction in 5-HT availability in the CNS of depressed patients. Given a functional relationship among the Th1 -dominated immune system, the serotonergic system, and MD, antidepressant therapy should be adequate to induce a Th1 to Th2 shift. There are indeed some reports demonstrating the potency of antidepressants to significantly reduce the IFN-γ/IL-10 ratio in vitroCitation143 and to suppress the Th1 response in patients.Citation144
The most frequently investigated immune parameter in patients suffering from MD is IL-6. Most of the publications report a marked increase of in vitro IL-6 productionCitation145 or serum IL-6 levels in depressed patients.Citation146-Citation151 Since IL-6 is a prominent marker of monocyte activity, a predominant activation of the monocyte/macrophage system in MD was hypothesized.Citation152 IL-6 may be involved in the modulation of the HPA axis.Citation153 Activation of the HPA axis is one of the best-documented changes in MD.Citation154 Furthermore, the relationship between psychological or physical stress and an enhanced IL-6 secretion in the peripheral immune system seems to be well established.Citation155-Citation158 Impaired stress coping is often observed in depressed patients. Thus, the high number of reports of elevated peripheral IL-6 levels in MD patients may be related to psychological stress.
On the other hand, there is evidence for a relationship between high peripheral IL-6 levels and elevated CNS 5-HT availability. IV or IP administration of IL-6 induced not only an activation of the HPA axis, but also an increase in brain tryptophan and 5-HT metabolism, whereas the norepinephrine metabolism was unaffected.Citation113 Accordingly, IL-6 seems to mediate the activation of the HPA axis and the 5-HT CNS after administration of the endotoxin LPS.Citation112 Thus, elevated plasma levels of IL-6 do not fit with the hypothesis of a 5-HT deficiency in MD. Rather, it should be recognized that an inherent heterogeneity exists in the etiology of depression and different neurotransmitter systems may be disturbed.
On the basis of the commonly accepted idea that MD may be a heterogeneous group of disease entities, the group of Arolt and Rothermundt investigated the difference between melancholic and nonmelancholic MD regarding their cytokine expression patterns.Citation159 They detected profound differences between these diagnostic subgroups: nonmelancholic patients showed increased counts of leukocytes, lymphocytes, and natural killer (NK) cells in the acute stage of disease and after 2 and 4 weeks of treatment. However, their in vitro production of the cytokines IL-2, IL-10, and IFN-γ was unchanged compared with that of healthy controls. Melancholic patients on the other hand demonstrated normal cell counts, but a decreased in vitro production of IL-2, IFN-γ, and IL-10 during the acute stage of disease. Following clinical improvement, cytokine production patterns normalized in these patients.
Schizophrenia
A pathophysiological role of cytokines is also discussed in the other major psychiatric disorder, schizophrenia. The reports of the psychotic symptoms inducing effects of IL-2 in cancer patients attracted attention of this Th1-like cytokine to the immunopsychiatric schizophrenia research. Early studies reported elevated IL-2 levels in cerebrospinal fluid of schizophrenia patients,Citation160,Citation161 but others failed to replicate these intriguing findings.Citation162-Citation165 IL-2 levels in serum were reported to be either increasedCitation166 or decreased.Citation167
A significant decrease in the production of IL-2 by peripheral lymphocytes is one of the best-replicated immunological findings in schizophrenia.Citation168,Citation169 Some data suggested that decreased IL-2 production is associated with acute illness in patients who produce elevated amounts of autoantibodies, or in patients with later age at onset,Citation170-Citation172 though there are again some contradictory findings.Citation173,Citation174
The group led by Arolt repeatedly found a markedly decreased in vitro production of IFN-γ, but the association with psychopathological variables was not consistently replicable.Citation168,Citation175,Citation176
Mittleman et al examined the cerebrospinal fluid of juvenile first-onset schizophrenia patients in comparison to juvenile patients with obsessive-compulsive disorder or attention deficit hyperactivity, and reported a reduction to undetectable levels of IFN-γ in most of the schizophrenia patients, in contrast to the measured levels in the other neuropsychiatrie patients. On the other hand, levels of the Th2 cytokine IL-4 were only detectable in cerebrospinal fluid of juvenile patients with schizophrenia, but were too low to be detected in that of the control groups.Citation165
As stated above, the IL-10 gene was identified as a possible susceptibility gene for schizophrenia. it is remarkable that administration of clozapine at a 10-4 M concentration markedly reduces the in vitro production of IL-10 by stimulated lymphocytes.Citation177 This may indicate the possible relevance of IL-10 in the pathophysiology of schizophrenia. IL-10 was in fact reported to be increased in the serum of schizophrenia patients.Citation178 Two out of three in vitro stimulation studies showed no difference between schizophrenia patients and healthy controls,Citation174,Citation179 while one described a highly significant increase in IL-10 production of chronically ill schizophrenia patients, compared with healthy controls, with medium levels in the subgroup of paranoid schizophrenia patients.Citation180
Peripheral administration of IL-6 induces increased dopamine and 5-HT turnovers in the hippocampus and frontal cortex of rodents, without influencing the metabolism of norepinephrine.Citation114 Within the brain, IL-6 is produced not only by glial cells, but also by neurons.Citation181 Thus, IL-6 is of interest in schizophrenia research. A remarkable number of publications report significantly increased serum IL-6 levels in schizophrenia patients. Some of them additionally found an association with duration of illness, negative symptoms, and treatmentresistant schizophrenia.Citation150,Citation166,Citation182,Citation187 However, these data are limited by several confounding factors influencing serum IL-6 levels, such as smoking, gender, age, body mass index, and ongoing infections, as well as clozapine treatment.Citation188,Citation189
In summary, the whole body of data on cytokines in schizophrenia indicates a relatively reduced production of Th1-like cytokines and a more pronounced production of Th2-like cytokines. This cytokine profile, together with the numerous findings of increased levels of circulating antibodies and other immunological data, prompted us to the hypothesis of a Th1 to Th2 shift in schizophrenia.Citation190,Citation191 A cytokine dysbalance like a Th2 predominance may be related to a disturbed neurodevelopment and brain maturation, as it is proposed as pathomorphologic correlate of schizophrenia.Citation192
Concluding remarks
The interdisciplinary approach of PNI has led to an integrative view of the immune system and the nervous system. Meanwhile, it is commonly accepted that not only does the CNS influence the immune reaction, but also that the immune system, particularly via its hormones- the cytokines - acts on brain function and behavior. There is ample evidence for the contribution of cytokines in psychiatric symptoms, syndromes, and disorders, and the involvement of the immune system fits to other commonly accepted etiopathological concepts like the neuro-developmental hypothesis of schizophrenia.
Genetic research gives further evidence for the possible involvement of the cytokine system especially in schizophrenia. However, the exact mechanisms of (inter) action must be elucidated in further investigations. Immunopsychiatrists may learn from somatic disorders like the systemic lupus erythematosus (SLE), an inflammatory disease affecting many organ systems including the CNS. The CNS affection in SLE encompasses a wide spectrum of neurological and psychiatric features including dementia, anxiety, depression, and psychosis,Citation193 and the causative role of cytokines, predominantly TNF-α, for the neuropsychiatrie symptoms of SLE was proposed.Citation134 Another aspect for future research derives from first therapy approaches in psychiatric disorders based on immunological considerations. The report of the therapeutic efficacy of a COX-2 inhibitor in schizophreniaCitation194 has particularly demonstrated the importance of immunological research in psychiatric disorders.
Thus, the new paradigm of brain-immune interaction appears to evoke new research and treatment strategies.
Selected abbreviations and acronyms
| BBB | = | blood-brain barrier |
| COX | = | cyclooxygenase-2 |
| CS | = | conditioned stimulus |
| CSF | = | colony-stimulating factor |
| CVO | = | circumventricular organ |
| HPA | = | hypothalamus-pituitary-adrenal (axis) |
| 5-HT | = | serotonin (5-hydroxytryptamine) |
| ICV | = | intracerebroventricular |
| IDO | = | indoleamine-2,3-dioxygenase |
| IFN | = | interferon |
| IL | = | interleukin |
| LPS | = | lipopolysaccharide |
| LT | = | lymphotoxin |
| MD | = | major depression |
| PNI | = | psychoneuroimmunology |
| TGFβ | = | transforming growth factor beta |
| Th | = | T helper (cell) |
| TNF-α | = | tumor necrosis factor alpha |
REFERENCES
- Metal'nikovS.ChorineV.Rôle des réflexes conditionnels dans l'immunité.Ann inst Pasteur.192640893900
- AderR.CohenN.Behaviorally conditioned immunosuppression.Psychosom Med.1975373333401162023
- RasmussenAF.MarshJT.BrillNQ.Increased susceptibility to herpes simplex in mice subjected to avoidance-learning stress or restraint.Proc Soc Exp Biol Med.19579618318913485051
- BesedovskyH.SorkinE.Network of immune-neuroendocrine interactions.Clin Exp Immunol.197727112849642
- WilliamsJM.PetersonRG.SheaPA.SchmedtjeJF.BauerDC.FeltenDL.Sympathetic innervation of murine thymus and spleen: evidence for a functional link between the nervous and immune systems.Brain Res Bull.1981683947193506
- FeltenDL.FeltenSY.BellingerDL.et al.Noradrenergic sympathetic neural interactions with the immune system: structure and function.Immunol Rev.19871002252603326822
- BerkenboschF.van OersJ.del ReyA.TildersF.BesedovskyH.Corticotropin-releasing factorproducing neurons in the rat activated by interleukin-1.Science19872385245262443979
- BesedovskyH.del ReyA.Neuroendocrine and metabolic responses induced by inter leukin-1.J Neurosci Res.1987181721782824799
- SapolskyR.RivierC.YamamotoG.PlotskyP.ValeW.Interleukin-1 stimulates the secretion of hypothalamic corticotropin-releasing factor.Science.19872385225242821621
- DuffGW.DurumSK.The pyrogenic and mitogenic actions of interleukin-1 are related.Nature.19833044494516603577
- KruegerJM.WalterJ.DinarelloCA.WolffSM.ChedidL.Sleep-promoting effects of endogenous pyrogen (interleukin-1).Am J Physiol.1984246(6, pt 2)R994R9996611091
- McCarthyDO.KlugerMJ.VanderAJ.Effect of centrally administered interleukin-1 and endotoxin on food intake of fasted rats.Physiol Behav.1986367457493520613
- ChanceWT.FischerJE.Aphagic and adipsic effects of interleukin-1.Brain Res.19915682612641814572
- CrestaniF.SeguyF.DantzerR.Behavioural effects of peripherally injected interleukin-1: role of prostaglandins.Brain Res.19915423303352029641
- SmedleyH.KatrakM.SikoraK.WheelerT.Neurological effects of recombinant human interferon.BMJ (Clin Res Ed).1983286262264
- SpriggsDR.ShermanML.FreiE III.KufeDW.Clinical studies with tumour necrosis factor.Ciba Found Symp.19871312062273330011
- KapasL.KruegerJM.Tumor necrosis factor-beta induces sleep, fever, and anorexia.Am J Physiol.1992263(3, pt 2)R703R7071415661
- KapasL.HongL.CadyAB.et al.Somnogenic, pyrogenic, and anorectic activities of tumor necrosis factor-alpha and TNF-alpha fragments.Am J Physiol.1992263(3, pt 2)R708R7151357984
- DinarelloCA.BernheimHA.DuffGW.et al.Mechanisms of fever induced by recombinant human interferon.J Clin Invest.1984749069136590569
- KimuraM.MajdeJA.TothLA.OppMR.KruegerJM.Somnogenic effects of rabbit and recombinant human interferons in rabbits.Am J Physiol.1994267(1, pt 2)R53R617519411
- RibeiroRC.RillD.RobersonPK.et al.Continuous infusion of interieukin-2 in children with refractory malignancies.Cancer.1993726236288319196
- LeMayLG.VanderAJ.KlugerMJ.Role of interleukin 6 in fever in rats.Am J Physiol.1990258(3, pt 2)R798R8032316725
- AtkinsMB.RobertsonMJ.GordonM.et al.Phase I evaluation of intravenous recombinant human interleukin 12 in patients with advanced malignancies.Clin Cancer Res.199734094179815699
- PflanzS.TimansJC.CheungJ.et al.IL-27, a heterodimeric cytokine composed of EBI3 and p28 protein, induces proliferation of naive CD4(+) T cells.Immunity.20021677979012121660
- VilcekJ.The cytokines: an overview. In: Thomson A, ed.The Cytokine Handbook. London, UK: Academic Press;1998120
- KelsoA.Cytokines: principles and prospects.Immunol Cell Biol.1998763003179723771
- BazanJF.Unraveling the structure of IL-2.Science.19922574104131631562
- BoulayJL.PaulWE.The interleukin-4 family of lymphokines.Curr Opin Immunol.199242942981418708
- RothwellNJ.LuheshiGN.Interleukin 1 in the brain: biology, pathology and therapeutic target.Trends Neurosci.20002361862511137152
- FantuzziG.DinarelloCA.Interleukin-18 and interleukin-1 beta: two cytokine substrates for ICE (caspase-1).J Clin Immunol.19991911110080100
- SmithDE.RenshawBR.KetchemRR.KubinM.GarkaKE.SimsJE.Four new members expand the interleukin-1 superfamily.J Biol Chem.20002751169117510625660
- BodmerJL.SchneiderP.TschoppJ.The molecular architecture of the TNF superfamily.Trends Biochem Sci.200227192611796220
- JanewayCA.TraversP.WalportM.ShlomchikM.Immunobiology. 5th ed. New York, NY: Garland Publishing;2001
- FickenscherH.HorS.KupersH.KnappeA.WittmannS.StichtH.The interleukin-10 family of cytokines.Trends Immunol.200223899611929132
- GurlingHM.KalsiG.BrynjolfsonJ.et al.Genomewide genetic linkage analysis confirms the presence of susceptibility loci for schizophrenia, on chromosomes 1q32.2, 5q33.2, and 8p21-22 and provides support for linkage to schizophrenia, on chromosomes 11q23.3-24 and 20q12.1-11.23.Am J Hum Genet20016866167311179014
- RosaA.FananasL.CuestaMJ.PeraltaV.ShamP.1q21-q22 locus is associated with susceptibility to the reality-distortion syndrome of schizophrenia spectrum disorders.Am J Med Genet200211451651812116186
- BrzustowiczLM.HodgkinsonKA.ChowEW.HonerWG.BassettAS.Location of a major susceptibility locus for familial schizophrenia on chromosome 1q21-q22.Science.200028867868210784452
- ChiavettoLB.BoinF.ZanardiniR.et al.Association between promoter polymorphic haplotypes of interleukin-10 gene and schizophrenia.Biol Psychiatry.20025148048411922883
- KatilaH.HanninenK.HurmeM.Polymorphisms of the interleukin-1 gene complex in schizophrenia.Mol Psychiatry.1999417918110208450
- MeisenzahlEM.RujescuD.KirnerA.et al.Association of an interleukin-1beta genetic polymorphism with altered brain structure in patients with schizophrenia.Am J Psychiatry.20011581316131911481169
- SchwabSG.HallmayerJ.AlbusM.et al.A genome-wide autosomal screen for schizophrenia susceptibility loci in 71 families with affected siblings: support for loci on chromosome 10p and 6.Mol Psychiatry.2000563864911126394
- RiouxJD.DalyMJ.SilverbergMS.et al.Genetic variation in the 5q31 cytokine gene cluster confers susceptibility to Crohn disease.Nat Genet.20012922322811586304
- LindholmE.EkholmB.BalciunieneJ.et al.Linkage analysis of a large Swedish kindred provides further support for a susceptibility locus for schizophrenia on chromosome 6p23.Am J Med Genet19998836937710402504
- BoinF.ZanardiniR.PioliR.AltamuraCA.MaesM.GennarelliM.Association between -G308A tumor necrosis factor alpha gene polymorphism and schizophrenia.Mol Psychiatry.20016798211244489
- RiedelM.KronigH.SchwarzMJ.et al.No association between the G308A polymorphism of the tumor necrosis factor-alpha gene and schizophrenia.Eur Arch Psychiatry Clin Neurosci.200225223223412451465
- KwiatkowskiDJ.DiazMO.Dinucleotide repeat polymorphism at the IFNA locus (9p22).Hum Mol Genet199216581301184
- MatuseviciusD.NavikasV.PalasikW.PirskanenR.FredriksonS.LinkH.Tumor necrosis factor-alpha, lymphotoxin, interleukin (IL)-6, IL-10, IL-12 and perforin mRNA expression in mononuclear cells in response to acetylcholine receptor is augmented in myasthenia gravis.J Neuroimmunol.1996711911988982119
- YsselH.de WaalMR.RoncaroloMG.et al.IL-10 is produced by subsets of human CD4+ T cell clones and peripheral blood T cells.J immunol.1992149237823841356125
- HayakawaS.FujikawaT.FukuokaH.et al.Murine fetal resorption and experimental pre-eclampsia are induced by both excessive Th1 and Th2 activation.J Reprod immunol.20004712113810924746
- SederRA.PaulWE.Acquisition of lymphokine-producing phenotype by CD4+ T cells.Annu Rev immunol.1994126356737912089
- RomagnaniS.Biology of human TH1 and TH2 cells.J Clin immunol.1995151211297559914
- PaludanSR.lnterleukin-4 and interferon-gamma: the quintessence of a mutual antagonistic relationship.Scand J immunol.1998484594689822252
- AloisiF.RiaF.AdoriniL.Regulation of T-cell responses by CNS antigenpresenting cells: different roles for microglia and astrocytes.Immunol Today.20002114114710689302
- FordAL.FoulcherE.LemckertFA.SedgwickJD.Microglia induce CD4 T lymphocyte final effector function and death.J Exp Med.1996184173717458920862
- KreutzbergGW.Microglia: a sensor for pathological events in the CNS.Trends Neurosci.1996193123188843599
- KrakowskiML.OwensT.The central nervous system environment controls effector CD4+ T cell cytokine profile in experimental allergic encephalomyelitis.Eur J Immunol.199727284028479394808
- StalderAK.PagenstecherA.YuNC.et al.Lipopolysaccharide-induced IL-12 expression in the central nervous system and cultured astrocytes and microglia.J immunol.1997159134413519233631
- HailerNP.HeppnerFL.HaasD.NitschR.Astrocytic factors deactivate antigen presenting cells that invade the central nervous system.Brain Pathol.199884594749669697
- BanksWA.KastinAJ.DurhamDA.Bidirectional transport of interleukin1 alpha across the blood-brain barrier.Brain Res Bull.1989234334372611685
- BanksWA.OrtizL.PlotkinSR.KastinAJ.Human interleukin (IL) 1 alpha, murine IL-1 alpha and murine IL-1 beta are transported from blood to brain in the mouse by a shared saturable mechanism.J Pharmacol Exp Ther.19912599889961762091
- BanksWA.KastinAJ.BroadwellRD.Passage of cytokines across the blood-brain barrier.Neuroimmunomodulation.199522412488963753
- WaguespackPJ.BanksWA.KastinAJ.lnterleukin-2 does not cross the blood-brain barrier by a saturable transport system.Brain Res Bull.1994341031098044683
- ManessLM.KastinAJ.BanksWA.Relative contributions of a CVO and the microvascular bed to delivery of blood-borne IL-1 alpha to the brain.Am J Physiol.1998275(2, pt 1)E207E2129688620
- HashimotoM.IshikawaY.YokotaS.et al.Action site of circulating interleukin-1 on the rabbit brain.Brain Res.19915402172232054614
- DavsonH.SegalMB.Physiology of the CSF and Blood-Brain Barriers. Boca Raton, Fla: CRC Press;1996
- GanongWF.Circumventricular organs: definition and role in the regulation of endocrine and autonomic function.Clin Exp Pharmacol Physiol.20002742242710831247
- BlatteisCM.HalesJR.McKinleyMJ.FawcettAA.Role of the anteroventral third ventricle region in fever in sheep.Can J Physiol Pharmacol.198765125512603497700
- BlatteisCM.BealerSL.HunterWS.LlanosQ.AhokasRA.MashburnTA Jr.Suppression of fever after lesions of the anteroventral third ventricle in guinea pigs.Brain Res Bull.1983115195266365250
- LeeSC.DicksonDW.BrosnanCF.Interleukin-1, nitric oxide and reactive astrocytes.Brain Behav Immunol.19959345354
- StittJT.Evidence for the involvement of the organum vasculosum laminae terminalis in the febrile response of rabbits and rats.J Physiol.19853685015114078749
- EricssonA.AriasC.SawchenkoPE.Evidence for an intramedullary prostaglandin-dependent mechanism in the activation of stress-related neuroendocrine circuitry by intravenous interleukin-1.J Neurosci.199717716671799278551
- QuanN.HerkenhamM.Connecting cytokines and brain: a review of current issues.Histol Histopathol.20021727328811813877
- WatkinsLR.WiertelakEP.GoehlerLE.SmithKP.MartinD.MaierSF.Characterization of cytokine-induced hyperalgesia.Brain Res.199465415267982088
- SehicE.BlatteisCM.Blockade of lipopolysaccharide-induced fever by subdiaphragmatic vagotomy in guinea pigs.Brain Res.19967261601668836556
- FleshnerM.GoehlerLE.HermannJ.ReltonJK.MaierSF.WatkinsLR.Interleukin-1 beta induced corticosterone elevation and hypothalamic NE depletion is vagally mediated.Brain Res Bull.1995376056107670884
- OppMR.TothLA.Somnogenic and pyrogenic effects of interleukin1beta and lipopolysaccharide in intact and vagotomized rats.Life Sci.1998629239369496715
- Bret-DibatJL.BlutheRM.KentS.KelleyKW.DantzerR.Lipopolysaccharide and interleukin-1 depress food-motivated behavior in mice by a vagal-mediated mechanism.Brain Behav Immunol.19959242246
- KonsmanJP.ParnetP.DantzerR.Cytokine-induced sickness behaviour: mechanisms and implications.Trends Neurosci.20022515415911852148
- QuanN.WhitesideM.HerkenhamM.Time course and localization patterns of interleukin-1beta messenger RNA expression in brain and pituitary after peripheral administration of lipopolysaccharide.Neuroscience.1998832812939466417
- VallieresL.RivestS.Regulation of the genes encoding interleukin-6, its receptor, and gp130 in the rat brain in response to the immune activator lipopolysaccharide and the proinflammatory cytokine interleukin-1 beta.J Neurochem.199769166816839326296
- NadeauS.RivestS.Regulation of the gene encoding tumor necrosis factor alpha (TNF-alpha) in the rat brain and pituitary in response in different models of systemic immune challenge.J Neuropathol Exp Neurol.199958617710068315
- QuanN.SundarSK.WeissJM.Induction of interleukin-1 in various brain regions after peripheral and central injections of lipopolysaccharide.J Neuroimmunol.1994491251348294551
- EkM.EngblomD.SahaS.BlomqvistA.JakobssonPJ.Ericsson-DahlstrandA.Inflammatory response: pathway across the blood-brain barrier.Nature.200141043043111260702
- MarkKS.TrickierWJ.MillerDW.Tumor necrosis factor-alpha induces cyclooxygenase-2 expression and prostaglandin release in brain microvessel endothelial cells.J Pharmacol Exp Ther.20012971051105811356928
- JohnsonRW.von BorellE.Lipopolysaccharide-induced sickness behavior in pigs is inhibited by pretreatment with indomethacin.J Anim Sci.1994723093148157515
- DunnAJ.SwiergielAH.The role of cyclooxygenases in endotoxin- and interleukin-1-induced hypophagia.Brain Behav immunol.200014141152
- OehmichenM.DomaschD.WietholterH.Origin, proliferation, and fate of cerebrospinal fluid cells. A review on cerebrospinal fluid cell kinetics.J Neurol.19822271451506181221
- HickeyWF.Migration of hematogenous cells through the blood-brain barrier and the initiation of CNS inflammation.Brain Pathol.19911971051669702
- FreiK.PianiD.PfisterHW.FontanaA.Immune-mediated injury in bacterial meningitis.Int Rev Exp Pathol.199334(Pt B)1831928458712
- Del MaschioA.De LuigiA.Martin-PaduraI.et al.Leukocyte recruitment in the cerebrospinal fluid of mice with experimental meningitis is inhibited by an antibody to junctional adhesion molecule (JAM).J Exp Med.19991901351135610544206
- BienvenuJ.MonneretG.FabienN.RevillardJP.The clinical usefulness of the measurement of cytokines.Clin Chem Lab Med.20003826728510928646
- BakhietM.TjernlundA.MousaA.et al.RANTES promotes growth and survival of human first-trimester forebrain astrocytes.Nat Cell Biol.2001315015711175747
- VankelecomH.MatthysP.DenefC.Involvement of nitric oxide in the interferon-gamma-induced inhibition of growth hormone and prolactin secretion in anterior pituitary cell cultures.Mol Cell Endocrinol.19971291571679202399
- KamataM.HiguchiH.YoshimotoM.YoshidaK.ShimizuT.Effect of single intracerebroventricular injection of alpha-interferon on monoamine concentrations in the rat brain.Eur Neuropsychopharmacol.20001012913210706995
- MorikawaO.SakaiN.ObaraH.SaitoN.Effects of interferon-alpha, interferon-gamma and cAMP on the transcriptional regulation of the serotonin transporter.Eur J Pharmacol.19983493173249671113
- ShutoH.KataokaY.HorikawaT.FujiharaN.OishiR.Repeated interferon-alpha administration inhibits dopaminergic neural activity in the mouse brain.Brain Res.19977473483519046014
- KatafuchiT.TakeS.HoriT.Roles of cytokines in the neural-immune interactions: modulation of NMDA responses by IFN-alpha.Neurobiology (Bp).199533193278696300
- NickolaTJ.IgnatowskiTA.ReynoldsJL.SpenglerRN.Antidepressant drug-induced alterations in neuron-localized tumor necrosis factor-alpha mRNA and alpha(2)-adrenergic receptor sensitivity.J Pharmacol Exp Ther.200129768068711303058
- AndoT.DunnAJ.Mouse tumor necrosis factor-alpha increases brain tryptophan concentrations and norepinephrine metabolism while activating the HPA axis in mice.Neuroimmunomodulation.1999631932910474050
- HayleyS.WallP.AnismanH.Sensitization to the neuroendocrine, central monoamine and behavioural effects of murine tumor necrosis factoralpha: peripheral and central mechanisms.Eur J Neurosci.2002151061107611918665
- RamamoorthyS.RamamoorthyJD.PrasadPD.et al.Regulation of the human serotonin transporter by interleukin-1 beta.Biochem Biophys Res Commun.19952165605677488148
- GemmaC.ImeriL.De SimoniMG.MandaM.Interleukin-1 induces changes in sleep, brain temperature, and serotonergic metabolism.Am J Physiol.1997272(2, Pt 2)R601R6069124484
- ImeriL.GemmaC.De SimoniMG.OppMR.MandaM.Hypothalamic serotonergic activity correlates better with brain temperature than with sleep-wake cycle and muscle tone in rats.Neuroscience.1999891241124610362311
- OppMR.SmithEM.HughesTK Jr.Interleukin-10 (cytokine synthesis inhibitory factor) acts in the central nervous system of rats to reduce sleep.J Neuroimmunol.1995601651687642744
- PetittoJM.McCarthyDB.RinkerCM.HuangZ.GettyT.Modulation of behavioral and neurochemical measures of forebrain dopamine function in mice by species-specific interleukin-2.J Neuroimmunol.1997731831909058775
- HanischUK.QuirionR.Interleukin-2 as a neuroregulatory cytokine.Brain Res Brain Res Rev.1995212462848806016
- Van WagonerNJ.BenvenisteEN.Interleukin-6 expression and regulation in astrocytes.J Neuroimmunol.199910012413910695723
- GadientRA.OttenUH.Interieukin-6 (IL-6)- a molecule with both beneficial and destructive potentials.Prog Neurobiol.1997523793909304698
- SongC.MeraliZ.AnismanH.Variations of nucleus accumbens dopamine and serotonin following systemic interleukin-1, interleukin-2 or interleukin-6 treatment.Neuroscience.19998882383610363820
- QiuZ.ParsonsKL.GruolDL.Interleukin-6 selectively enhances the intracellular calcium response to NMDA in developing CNS neurons.J Neurosci.199515668866997472429
- QiuZ.SweeneyDD.NetzebandJG.GruolDL.Chronic interleukin-6 alters NMDA receptor-mediated membrane responses and enhances neurotoxicity in developing CNS neurons.J Neurosci.19981810445104569852582
- WangJ.DunnAJ.The role of interleukin-6 in the activation of the hypothalamo-pituitary-adrenocortical axis and brain indoleamines by endotoxin and interleukin-1 beta.Brain Res.19998153373489878816
- WangJ.DunnAJ.Mouse interleukin-6 stimulates the HPA axis and increases brain tryptophan and serotonin metabolism.Neurochem int.1998331431549761458
- ZalcmanS.Green-JohnsonJM.MurrayL.et al.Cytokine-specific central monoamine alterations induced by interleukin-1, -2 and -6.Brain Res.199464340497518332
- BarkhudaryanN.DunnAJ.Molecular mechanisms of actions of interieukin-6 on the brain, with special reference to serotonin and the hypothalamo-pituitary-adrenocortical axis.Neurochem Res.1999241169118010485589
- KatsukiH.NakaiS.HiraiY.AkajiK.KisoY.SatohM.Interleukin-1 beta inhibits long-term potentiation in the CA3 region of mouse hippocampal slices.Eur J Pharmacol.19901813233262166677
- BellingerFP.MadambaS.SigginsGR.Interleukin 1 beta inhibits synaptic strength and long-term potentiation in the rat CA1 hippocampus.Brain Res.19936282272348313151
- BeattieEC.StellwagenD.MorishitaW.et al.Control of synaptic strength by glial TNFalpha.Science.20022952282228511910117
- GibertiniM.Cytokines and cognitive behavior.Neuroimmunomodulation.199851601659730681
- GibertiniM.NewtonC.FriedmanH.KleinTW.Spatial learning impairment in mice infected with Legionella pneumophila or administered exogenous interieukin-1-beta.Brain Behav immunol.19959113128
- MellorAL.MunnDH.Tryptophan catabolism and T-cell tolerance: immunosuppression by starvation?Immunol Today.19992046947310500295
- MunnDH.ShafizadehE.AttwoodJT.BondarevI.PashineA.MellorAL.Inhibition of T cell proliferation by macrophage tryptophan catabolism.J Exp Med.19991891363137210224276
- WeissG.MurrC.ZollerH.et al.Modulation of neopterin formation and tryptophan degradation by Th1- and Th2-derived cytokines in human monocytic cells.Clin Exp Immunol.199911643544010361231
- AlberatiGD.RicciardiCP.KohlerC.CesuraAM.Regulation of the kynurenine metabolic pathway by interferon-gamma in murine cloned macrophages and microglial cells.J Neurochem.19966699610048769859
- FarrarWL.KilianPL.RuffMR.HillJM.PertCB.Visualization and characterization of interleukin 1 receptors in brain.J Immunol.19871394594632955042
- BrederCD.DinarelloCA.SaperCB.Interleukin-1 immunoreactive innervation of the human hypothalamus.Science.19882403213243258444
- TaziA.DantzerR.CrestaniF.Le MoalM.Interleukin-1 induces conditioned taste aversion in rats: a possible explanation for its pituitary-adrenal stimulating activity.Brain Res.19884733693713266091
- Plata-SalamanCR.OomuraY.KaiY.Tumor necrosis factor and interleukin-1 beta: suppression of food intake by direct action in the central nervous system.Brain Res.19884481061143260533
- Van DamAM.BrounsM.LouisseS.BerkenboschF.Appearance of interleukin-1 in macrophages and in ramified microglia in the brain of endotoxin-treated rats: a pathway for the induction of non-specific symptoms of sickness?Brain Res.19925882912961393581
- GattiS.BartfaiT.Induction of tumor necrosis factor-alpha mRNA in the brain after peripheral endotoxin treatment: comparison with interleukin-1 family and interleukin-6.Brain Res.19936242912948252403
- LayeS.ParnetP.GoujonE.DantzerR.Peripheral administration of lipopolysaccharide induces the expression of cytokine transcripts in the brain and pituitary of mice.Brain Res Mol Brain Res.1994271571627877446
- DantzerR.Cytokine-induced sickness behavior: where do we stand?Brain Behav immunol.200115724
- PapanicolaouDA.WilderRL.ManolagasSC.ChrousosGP.The pathophysiologic roles of interleukin-6 in human disease.Ann Intern Med.19981281271379441573
- SchaeferM.EngelbrechtMA.GutO.et al.Interferon alpha (IFNalpha) and psychiatric syndromes: a review.Prog Neuropsychopharmacol Biol Psychiatry.20022673174612188106
- CapuronL.RavaudA.NeveuPJ.MillerAH.MaesM.DantzerR.Association between decreased serum tryptophan concentrations and depressive symptoms in cancer patients undergoing cytokine therapy.Mol Psychiatry.2002746847312082564
- LernerDM.StoudemireA.RosensteinDL.Neuropsychiatrie toxicity associated with cytokine therapies.Psychosomatics.19994042843510479948
- FattovichG.GiustinaG.FavaratoS.RuolA.A survey of adverse events in 11 241 patients with chronic viral hepatitis treated with alfa interferon.J Hepatol.19962438478834023
- DenicoffKD.RubinowDR.PapaMZ.et al.The neuropsychiatrie effects of treatment with interleukin-2 and lymphokine-activated killer cells.Ann Intern Med.19871072933003497595
- ReichenbergA.YirmiyaR.SchuldA.et al.Cytokine-associated emotional and cognitive disturbances in humans.Arch Gen Psychiatry.20015844545211343523
- SeidelA.AroltV.HunstigerM.RinkL.BehnischA.KirchnerH.Increased CD56+ natural killer cells and related cytokines in major depression.Clin Immunol Immunopathol.19967883858599889
- MaesM.ScharpeS.MeltzerHY.et al.Increased neopterin and interferon-gamma secretion and lower availability of L-tryptophan in major depression: further evidence for an immune response.Psychiatry Res.1994541431607761549
- MaesM.VerkerkR.VandoolaegheE.et al.Serotonin-immune interactions in major depression: lower serum tryptophan as a marker of an immune-inflammatory response.Eur Arch Psychiatry Clin Neurosci.19972471541619224908
- MaesM.SongC.LinAH.et al.Negative immunoregulatory effects of antidepressants: inhibition of interferon-gamma and stimulation of interieukin-10 secretion.Neuropsychopharmacology.19992037037910088138
- MohrDC.GoodkinDE.IslarJ.HauserSL.GenainCP.Treatment of depression is associated with suppression of nonspecific and antigen-specific T(H)1 responses in multiple sclerosis.Arch Neurol.2001581081108611448297
- MaesM.ScharpeS.MeltzerHY.et al.Relationships between interieukin-6 activity, acute phase proteins, and function of the hypothalamicpituitary-adrenal axis in severe depression.Psychiatry Res.19934911277511248
- MaesM.MeltzerHY.BosmansE.et al.Increased plasma concentrations of interleukin-6, soluble interleukin-6, soluble interleukin-2 and transferrin receptor in major depression.J Affect Disord.1995343013098550956
- SluzewskaA.RybakowskiJ.BosmansE.et al.Indicators of immune activation in major depression.Psychiatry Res.1996641611678944394
- BerkM.WadeeAA.KuschkeRH.O'NeillKA.Acute phase proteins in major depression.J Psychosom Res.1997435295349394269
- MaesM.BosmansE.De JonghR.KenisG.VandoolaegheE.NeelsH.Increased serum IL-6 and IL-1 receptor antagonist concentrations in major depression and treatment-resistant depression.Cytokine.199798538589367546
- FrommbergerUH.BauerJ.HaselbauerP.FraulinA.RiemannD.BergerM.Interleukin-6 (IL-6) plasma levels in depression and schizophrenia: comparison between the acute state and after remission.Eur Arch Psychiatry Clin Neurosci.19972472282339332905
- SongC.LinA.BonaccorsoS.et al.The inflammatory response system and the availability of plasma tryptophan in patients with primary sleep disorders and major depression.J Affect Disord.1998492112199629951
- SmithRS.MaesM.The macrophage-T-lymphocyte theory of schizophrenia: additional evidence.Med Hypotheses.1995451351418531836
- Plata-SalamanCR.Immunoregulators in the nervous system.Neurosci Biobehav Rev.1991151852151852312
- RoyA.PickarD.PaulS.DoranA.ChrousosGP.GoldPW.CSF corticotropin-releasing hormone in depressed patients and normal control subjects.Am J Psychiatry.19871446416453495188
- SalasMA.EvansSW.LevellMJ.WhicherJT.Interleukin-6 and ACTH act synergistically to stimulate the release of corticosterone from adrenal gland cells.Clin Exp immunol.1990794704732156641
- LeMayLG.VanderAJ.KlugerMJ.The effects of psychological stress on plasma interleukin-6 activity in rats.Physiol Behav.1990479579612388952
- ZhouD.KusnecovAW.ShurinMR.DePaoliM.RabinBS.Exposure to physical and psychological stressors elevates plasma interleukin 6: relationship to the activation of hypothalamic-pituitary-adrenal axis.Endocrinology.1993133252325308243274
- MiyaharaS.KomoriT.FujiwaraR.et al.Effects of repeated stress on expression of interleukin-6 (IL-6) and IL-6 receptor mRNAs in rat hypothalamus and midbrain.Life Sci.200066L93L98
- RothermundtM.AroltV.FenkerJ.GutbrodtH.PetersM.KirchnerH.Different immune patterns in melancholic and non-melancholic major depression.Eur Arch Psychiatry Clin Neurosci.2001251909711407444
- LicinioJ.SeibyiJP.AltemusM.CharneyDS.KrystaiJH.Elevated CSF levels of interleukin-2 in neuroleptic-free schizophrenic patients.Am J Psychiatry.1993150140814108102512
- McAllisterCG.van KammenDP.RehnTJ.et al.Increases in CSF levels of interleukin-2 in schizophrenia: effects of recurrence of psychosis and medication status.Am J Psychiatry.1995152129112977653683
- RapaportMH.McAllisterCG.PickarD.TamarkinL.KirchDG.PaulSM.CSF IL-1 and IL-2 in medicated schizophrenic patients and normal volunteers.Schizophr Res.1997251231299187011
- BarakV.BarakY.LevineJ.NismanB.RoismanI.Changes in interleukin-1 beta and soluble interleukin-2 receptor levels in CSF and serum of schizophrenic patients.J Basic Clin Physiol Pharmacol.1995661698562579
- el MallakhRS.SuddathRL.WyattRJ.Interieukin-1 alpha and interieukin-2 in cerebrospinal fluid of schizophrenic subjects.Prog Neuropsychopharmacol Biol Psychiatry.1993173833918475320
- MittlemanBB.CastellanosFX.JacobsenLK.RapoportJL.SwedoSE.ShearerGM.Cerebrospinal fluid cytokines in pediatric neuropsychiatrie disease.J Immunol.1997159299429999300724
- KimYK.KimL.LeeMS.Relationships between interleukins, neurotransmitters and psychopathology in drug-free male schizophrenics.Schizophr Res.20004416517510962218
- TheodoropoulouSt.SpanakosG.BaxevanisCN.et al.Cytokine serum levels, autologous mixed lymphocyte reaction and surface marker analysis in never medicated and chronically medicated schizophrenic patients.Schizophr Res.2001471225
- AroltV.RothermundtM.WandingerKP.KirchnerH.Decreased in vitro production of interferon-gamma and interleukin-2 in whole blood of patients with schizophrenia during treatment.Mol Psychiatry.2000515015810822342
- KimYK.LeeMS.SuhKY.Decreased interleukin-2 production in Korean schizophrenic patients.Biol Psychiatry.1998437017049583005
- GanguliR.BrarJS.SolomonW.ChengappaKN.RabinBS.Altered interleukin-2 production in schizophrenia: association between clinical state and autoantibody production.Psychiatry Res.1992441131231480677
- YangZW.ChengappaKN.ShurinG.et al.An association between anti-hippocampal antibody concentration and lymphocyte production of IL-2 in patients with schizophrenia.Psychol Med.1994244494558084939
- GanguliR.BrarJS.ChengappaKR.et al.Mitogen-stimulated interleukin-2 production in never-medicated, first-episode schizophrenic patients. The influence of age at onset and negative symptoms [see comments].Arch Gen Psychiatry.1995526686727632120
- O'DonnellMC.CattsSV.WardPB.et al.Increased production of interieukin-2 (IL-2) but not soluble interleukin-2 receptors (slL-2R) in unmedicated patients with schizophrenia and schizophreniform disorder.Psychiatry Res.1996651711789029665
- CazzulloCL.SacchettiE.GalluzzoA.et al.Cytokine profiles in drugnaive schizophrenic patients.Schizophr Res.20014729329811278147
- HornbergM.AroltV.WilkeI.KruseA.KirchnerH.Production of interferons and lymphokines in leukocyte cultures of patients with schizophrenia.Schizophr Res.1995152372427543276
- WilkeI.AroltV.RothermundtM.WeitzschC.HornbergM.KirchnerH.Investigations of cytokine production in whole blood cultures of paranoid and residual schizophrenic patients.Eur Arch Psychiatry Clin Neurosci.19962462792848863007
- SongC.LinA.KenisG.BosmansE.MaesM.Immunosuppressive effects of clozapine and haloperidol: enhanced production of the interleukin-1 receptor antagonist.Schizophr Res.20004215716410742653
- MaesM.BocchioCL.BignottiS.et al.Increased serum interleukin-8 and interleukin-10 in schizophrenic patients resistant to treatment with neuroleptics and the stimulatory effects of clozapine on serum leukemia inhibitory factor receptor.Schizophr Res.20025428129111950553
- RothermundtM.AroltV.WeitzschC.EckhoffD.KirchnerH.Immunological dysfunction in schizophrenia: a systematic approach.Neuropsychobiology.1998371861939648126
- CazzulloCL.ScaroneS.GrassiB.et al.Cytokine production in chronic schizophrenia patients with or without paranoid behaviour.Prog Neuropsychopharmacol Biol Psychiatry.1998229479579789879
- RingheimGE.BurgherKL.HerouxJA.Interleukin-6 mRNA expression by cortical neurons in culture: evidence for neuronal sources of interleukin-6 production in the brain.J Neuroimmunol.1995631131238550808
- GanguliR.YangZ.ShurinG.et al.Serum interleukin-6 concentration in schizophrenia: elevation associated with duration of illness.Psychiatry Res.1994511107910974
- MaesM.BosmansE.CalabreseJ.SmithR.MeltzerHY.Interleukin-2 and interleukin-6 in schizophrenia and mania: effects of neuroleptics and mood stabilizers.J Psychiatr Res.1995291411527666381
- NaudinJ.MegeJL.AzorinJM.DassaD.Elevated circulating levels of IL-6 in schizophrenia.Schizophr Res.1996202692738827853
- LinA.KenisG.BignottiS.et al.The inflammatory response system in treatment-resistant schizophrenia: increased serum interleukin-6.Schizophr Res.1998329159690329
- AkiyamaK.Serum levels of soluble IL-2 receptor alpha, IL-6 and IL-1 receptor antagonist in schizophrenia before and during neuroleptic administration.Schizophr Res.1999379710610227112
- MaesM.BocchioCL.BignottiS.et al.Effects of atypical antipsychotics on the inflammatory response system in schizophrenic patients resistant to treatment with typical neuroleptics.Eur Neuropsychopharmacol.20001011912410706993
- HaackM.HinzeSD.FenzelT.et al.Plasma levels of cytokines and soluble cytokine receptors in psychiatric patients upon hospital admission: effects of confounding factors and diagnosis.J Psych iatr Res.199933407418
- MaesM.BosmansE.KenisG.De JongR.SmithRS.MeltzerHY.In vivo immunomodulatory effects of clozapine in schizophrenia.Schizophr Res.1997262212259323354
- SchwarzMJ.ChiangS.MüllerN.AckenheilM.T-heiper-1 and T-heiper-2 responses in psychiatric disorders.Brain Behav Immunol.200115340370
- SchwarzMJ.MullerN.RiedelM.AckenheilM.The Th2-hypothesis of schizophrenia: a strategy to identify a subgroup of schizophrenia caused by immune mechanisms.Med Hypotheses.20015648348611339852
- NawaH.TakahashiM.PattersonPH.Cytokine and growth factor involvement in schizophrenia - support for the developmental model.Mol Psychiatry.2000559460311126390
- The American College of Rheumatology nomenclature and case definitions for neuropsychiatrie lupus syndromes.Arthritis Rheum.19994259960810211873
- MüllerN.RiedelM.ScheppachC.et al.Beneficial antipsychotic effects of celecoxib add-on therapy compared to risperidone alone in schizophrenia.Am J Psychiatry.20021591029103412042193