Abstract
Caffeine is not considered addictive, and in animals it does not trigger metabolic increases or dopamine release in brain areas involved in reinforcement and reward. Our objective was to measure caffeine effects on cerebral perfusion in humans using single photon emission computed tomography, with a specific focus on areas of reinforcement and reward. Two groups of nonsmoking subjects were studied, one with a low (8 subjects) and one with a high (6 subjects) daily coffee consumption. The subjects ingested 3 mg/kg caffeine or placebo in a raspberry-tasting drink, and scans were performed 45 min after ingestion. A control group of 12 healthy volunteers receiving no drink was also studied. Caffeine consumption led to a generalized, statistically nonsignificant perfusion decrease of 6% to 8%, comparable in low and high consumers. Compared with controls, low consumers displayed neuronal activation bilaterally in inferior frontal gyrusanterior insular cortex and uncus, left internal parietal cortex, right lingual gyrus, and cerebellum. In high consumers, brain activation occurred bilaterally only in hypothalamus. Thus, on a background of widespread low-amplitude perfusion decrease, caffeine activates a few regions mainly involved in the control of vigilance, anxiety, and cardiovascular regulation, but does not affect areas involved in reinforcing and reward.
No se considera que la cafeína sea adictiva y en animales no provoca aumento del metabolismo ni liberación de dopamina en las áreas que participan en los mecanismos de refuerzo y recompensa. Nuestros objetivos fueron medir los efectos de la cafeína en la perfusión cerebral utilizando la tomografía computada de emisión de fotón único con un foco específico en las áreas de refuerzo y recompensa. Se estudiaron dos grupos de sujetos no fumadores, uno con bajo consumo diario de café (8 sujetos) y otro con alto consumo (6 sujetos). Los sujetos ingirieron 3mg/kg de cafeína o placebo en una bebida con sabor a frambuesa y los escáners se realizaron 45 minutos post-ingesta. También se estudió un grupo control de 12 voluntarios sanos que no recibió ninguna bebida. El consumo de cafeína determinó una disminución generalizada del 6% al 8% de la perfusión, sin significación estadística, comparable tanto en los sujetos con bajo como con alto consumo. En comparación con los controles, los sujetos de bajo consumo mostraron actividad neuronal bilateral en el giro frontal inferiorcorteza insular anterior y en el uncus, la corteza parietal interna izquierda, el giro lingual derecho y el cerebelo. En los sujetos de alto consumo la activación cerebral ocurrió bilateralmente sólo en el hipotálamo. Por lo tanto, la cafeína -aunque provoca de fondo una extensa disminución de la perfusión de baja amplitud- activa unas pocas regiones involucradas principalmente en el control de la vigilancia, la ansiedad y la regulación cardiovascular, sin afectar áreas que participan en el refuerzo y la recompensa.
La caféine n'engendre pas de dépendance et ne déclenche pas d'augmentation du métabolisme ou de libération de dopamine dans les aires cérébrales impliquées dans le renforcement ou la récompense chez les animaux. Notre but était de mesurer les effets de la caféine sur la perfusion cérébrale humaine en utilisant la tomographie par émission monophotonique, en insistant sur les aires du renforcement et de la récompense. Deux groupes de sujets non fumeurs ont été étudiés, l'un ayant une faible (8 sujets) et l'autre une forte (6 sujets) consommation quotidienne de café. Les sujets ont pris 3 mg/kg de caféine ou un placebo dans une boisson au goût de framboise et des scanners ont été réalisés 45 min après l'ingestion, un groupe témoin de 12 volontaires sains ne prenant aucune boisson étant également étudié. La consommation de caféine a conduit à une réduction généralisée du débit statistiquement non significative de 6 % à 8 %, comparable chez les petits ou gros consommateurs. Les petits consommateurs, par rapport aux témoins, ont présenté une activation neuronale bilatérale dans le gyrus frontal inférieur, le cortex insulaire antérieur et l'uncus, dans le cortex pariétal interne gauche, le gyrus lingual droit et le cervelet. Chez les gros consommateurs, l'activation cérébrale est apparue de façon bilatérale seulement dans l'hypothalamus. Ainsi, sur un fond de diminution étendue du débit de faible amplitude, la caféine activerait quelques régions principalement impliquées dans le contrôle de la vigilance, de l'anxiété et de la régulation cardiovasculaire, mais n'affecterait pas les aires impliquées dans le renforcement et la récompense.
The methylxanthine, caffeine, is the most widely used psychoactive substance in the world. Most of the caffeine consumed comes from dietary sources such as coffee, tea, soft drinks, and chocolate. The content of caffeine of these food items ranges from 70 to 220 mg/150 mL for coffee to 30 to 40 mg/150 mL for tea, 15-35 mg/150 mL for cola drinks, and 4 mg/150 mL for chocolate drinks.Citation1 World caffeine consumption from all sources can be estimated as 76 mg/person/day, but reaches about 220 mg/day in the United States and Canada, and more than 400 mg/person/day in Sweden and Finland.Citation1,Citation2
Caffeine acts as a psychostimulant and exerts numerous effects on the brain. These include stimulant effects on motor behavior, modulation of mood states and levels of anxiety, effects on vigilance and sleep, on information processing and performance.Citation3 In the periphery, the effects of coffee/caffeine have been studied, but at moderate doses, they do not appear to exert harmful effects on cardiovascular function.Citation4
The issue of a possible dependence on caffeine has been debated for many years.Citation5-Citation8 Caffeine acts as a mild reinforcer (ie, maintaining its self-administration or being preferentially chosen over placebo), although not consistently in both humans and animals.Citation6 In humans, the widely recognized behavioral stimulant and mildly reinforcing properties of caffeine are probably responsible for the maintenance of caffeine self-administration.Citation7,Citation9 The possible physical dependence to the methyxanthine has been considered for about two decades,Citation5,Citation9,Citation10 but appears to be quite low compared with common drugs of abuse, such as cocaine, amphetamine, morphine, and nicotine.
The critical role of the mesolimbic dopamine system has been emphasized as underlying drug dependence.Citation11,Citation12 This system consists of the dopaminergic neurons originating in the ventral tegmental area, projecting to the nucleus accumbens, and ending in the frontal and prefrontal cortex. Drugs of abuse selectively activate the shell of the nucleus accumbens, which belongs to the mesolimbic dopaminergic system and is currently recognized as a critical target of drugs of abuse.Citation13-Citation15 The shell of the nucleus accumbens plays a role in emotion, motivation, and reward functions. The laterodorsal core part of the nucleus accumbens regulates somatomotor functions. The drugs of abuse specifically increase dopamine release and functional activity (glucose utilization and blood flow) in the shell of the nucleus accumbens without affecting the core of the nucleus.Citation13,Citation14 These druginduced changes in the shell of the nucleus accumbens have been hypothesized to relate to the general abuse liability of these drugs independently from their specific mechanism of action.Citation12
In a previous study, we investigated the effects of 1 to 10 mg/kg caffeine on local cerebral glucose utilization in rats. We showed that 1 to 5 mg/kg caffeine in the rat (70 to 350 mg for a 70-kg individual) which are in the range of normal human daily consumptionCitation1,Citation2 failed to increase metabolic levels in the shell of the nucleus accumbens.Citation15 Likewise, caffeine did not induce a release of dopamine in the shell of the nucleus accumbens when injected over a large spectrum of doses ranging from 0.5 to 30.0 mg/kg.Citation16,Citation17 However, the high dose of 10 mg/kg caffeine in rats, no longer representative of human consumption - 7 large cups of coffee in one sitting - led to unspecific metabolic increases in the shell and the core of the nucleus accumbens, together with the activation of most motor, limbic, thalamic, and cortical regions.Citation15
The objective of the present study was to extend these approaches to the human situation and to measure the effects of caffeine on cerebral perfusion in human subjects using single photon emission computed tomography (SPECT). We measured caffeine-induced perfusion changes in a large number of brain areas, including the areas involved in the circuit of dependence and reward, mainly the nucleus accumbens and prefrontal cortex. Moreover, two groups of subjects were studied, one with a low daily coffee consumption and one with a high daily coffee consumption. They were compared with a control group not exposed to any drink to account for the intraindividual variations of perfusion between two consecutive scans.
Methods
Subjects
A total number of 26 normal human subjects (10 men and 16 women), ranging in age from 19 to 47 (mean age, 29.9 + 7.9 years; median, 28 years) with no history or clinical evidence of medical, neurological, or psychiatric disease participated in this study. The subjects were recruited among the healthy nonsmoking population, and met the following additional criteria: no night shiftwork, no use of any medication except for birth control, and no report of any history of alcohol or drug abuse. To exclude any morphological abnormality, cerebral magnetic resonance imaging (MRI) was performed in all cases. All subjects gave their informed written consent before the study, which was approved by the local ethical committee.
Caffeine groups
Within the caffeine groups, the first subgroup of eight subjects consisted of a population of very low caffeine consumers or abstainers (0 to 1 cup of coffee per day, ie, less than 100 mg/day, low-consumption, LC group); the second one included six subjects who consumed elevated quantities of coffee (more than 4 cups per day, ie, over 500 mg/day) and reported feeling “dependent” on coffee (high-consumption, HC group). This was only based on the subjects' own feelings and not on any DSM-IV criteria. The subjects were told that they were entering a study on the effects of caffeine on cerebral circulation, but were not informed about the exact purpose of the study, ie, the study of the specific effect of caffeine on the brain areas involved in drug dependence.
The subjects were asked to observe a 12-hour abstinence from caffeine-containing foods and beverages prior to the measurement of cerebral blood flow. Blood samples were taken at arrival at the hospital to reinforce compliance. The subjects ingested 3 mg/kg body weight caffeine or a placebo in a raspberry-tasting drink. The drinks were prepared by the pharmacy of the University Hospitals and were administered in a double -blind, randomized, counterbalanced design. The blood pressure and heart rate were measured and the mood and anxiety profiles of the subjects assessed with a specific questionnaire before and after caffeine ingestion. The questionnaire used was the State-Trait Anxiety Inventory (STAI) for adults which is the most widely used selfreport measure of tension and anxiety.
Control group
To allow group analysis and evaluation of the inter- and intraindividual variations of cerebral perfusion, a control group of 12 healthy volunteers was included in the study. Within this group of twelve healthy volunteers (low consumers of 0 to 2 cups of coffee daily) not receiving any drink before the two SPECT examinations, eight and six were randomly selected for comparison with the LC and HC caffeine group, respectively, while the whole caffeine-consuming group was compared with the totality of the control group.
SPECT procedure
The caffeine groups subjects were subjected to two separate morning examinations upon arrival at the hospital: (i) one SPECT study after the placebo beverage; and (ii) one SPECT study after the caffeine containing beverage. The two beverages were given on two different days at 7-day interval, in a double-blind randomized counterbalanced design. Upon arrival at the clinic, the subjects were invited to relax in a comfortable armchair in a quiet and pleasant room. A venous catheter for tracer injection was immediately inserted into the left arm, and a first blood sample was taken to measure caffeine levels to check for compliance to 12 h caffeine abstinence. Heart rate and blood pressure measurements were then performed and the subjects filled in the STAI questionnaire. Then, subjects received the caffeine or placebo drink and were asked to rest in the same surrounding for 45 min. This time was chosen since caffeine reaches peak values in the brain between 45 and 60 min postingestion.Citation3 Thereafter, the subjects underwent the same procedure for the measurement of plasma caffeine levels, cardiovascular parameters, and filled in the STAI questionnaire again. Immediately afterwards, the tracer, 640-925 MBq99mTc-ethyl cysteinate dimer (ECD, Neurolite, Bristol-Myers Squibb Medical Imaging), was injected into the already inserted venous catheter. The subjects were not allowed to read, write, or talk for 5 min, including the fixation period of the radiotracer.
The control group subjects were also subjected to two separate morning SPECT examinations at 7-day intervals, in the same conditions, without any beverage. This procedure was used to evaluate the intrasubject variability between two examinations and to avoid the consequences of variable spontaneous mental activity and/or possible perfusion changes induced by the stressful environment related to SPECT examination.
SPECT imaging studies were realized with a low-energy, high-resolution, double-head camera (Helix, Elscint). The camera was operated in the “stop and shoot” mode, with acquisitions at 3° intervals and a total acquisition time of 25 min (120 projections, 642 matrix). The total number of counts was superior to 6 million. Slices were reconstructed by filtered back-projection using a Metz filter (FWMH of 8 mm). Slices were acquired 30 min after the injection of ECD.
Image analysis and statistics
Images were realigned with each other using affine algorithm in MEDIMAX software,Citation18 transformed into the standard anatomic space corresponding to the atlas of Talairach, and normalized in statistical parametric mapping software (SPM2, Wellcome Department of Imaging Neuroscience, London) using MATLAB (Mathworks Inc, Massachusetts) for calculations and image matrix manipulations.
After the preprocessing of SPECT images, all the statistical analysis was performed by multigroup analysis with SPM2. A voxel-by-voxel comparison according to the general linear model and I statistics was used to calculate the differences in cerebral perfusion between the two conditions (caffeine vs placebo) of caffeine groups and the differences between two conditions (rest vs rest) of the control group (LC group vs control group, HC group vs control group, and LC + HC groups vs control group). The resulting statistical parametric map was subsequently used to assign probability values (to voxels and clusters), which were corrected for multiple comparisons applied for the whole brain. Significant differences were defined at a corrected value of P<=0.001 and size >=10 voxels (80 mm3).
Results
Clinical data
Two groups of subjects were studied, including eight subjects (seven females and one male) and seven subjects (two females and five males) belonging to the LC and HC groups, respectively (Table I). All clinical data including age, arterial blood pressure, heart rate, electrocardiographic recording, and other current physiological parameters (plasma potassium level and hepatic enzyme activities) were in the physiological range and identical in both groups. None of these parameters was significantly affected by caffeine or placebo. In addition, the clinical surroundings had no significant influence on anxiety levels, as assessed upon arrival, and neither did the dose of caffeine ingested and the insertion of the catheter for ECD injection, as assessed 45 min later. Subjects had a tendency to feel more alert after the ingestion of the caffeine-containing drink. The low level of caffeine before SPECT analysis confirmed that the patients had omitted the methylxanthine from their food and beverage for 12 h. Circulating levels of caffeine were increased by a factor of 8 to 13 at about 1 h after caffeine ingestion.
Table I Clinical data of the subjects undergoing SPECT examinations for the effects of caffeine on cerebral blood flow
SPECT data
The consumption of caffeine led to a generalized decrease in perfusion levels of 6% to 8% which was of similar amplitude in both LC and HC groups (Table II). Compared with intraindividual variations of control subjects, these changes were not statistically significant.
Table II Caffeine-induced changes in cerebral perfusion calculated as follows : (SPECT CAFFEINE - SPECT PLACEBO) / SPECT PLACEBO using SISCOM
On the other hand, discrete perfusion increases corresponding to specific neuronal activation were found in specific regions. Indeed, when the LC caffeine group was compared with the control group ( ), increases in perfusion occurred bilaterally in the inferior frontal gyrus-anterior insular cortex (predominantly on the right side) and in the uncus, on the left side in the internal parietal cortex, on the right side in the lingual gyrus and cerebellum. In the HC group compared with the control group, perfusion increases were located bilaterally in hypothalamus. When both caffeine groups were pooled and compared with the whole control group, significant perfusion increases occurred bilaterally in the inferior frontal gyrus-anterior insula, hypothalamus, right cerebellum, and left uncus ( ).
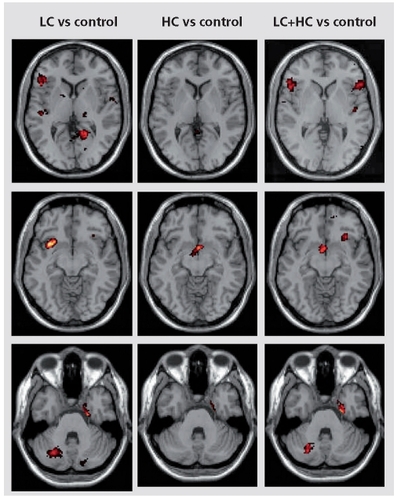
Discussion
The main findings of this study were the lack of significant differences in perfusion between caffeine-exposed subjects and controls, whether they were HC or LC, the lack of effects of the methylxanthine on the areas of reinforcing and reward and only very discrete changes in perfusion in areas mediating mainly anxiety, attention and vigilance, and cardiovascular function.
The vasoconstrictive properties of caffeine in the brain have been known for a long time, and caffeine has been shown to decrease cerebral blood flow in humans.Citation19-Citation23 Previous studies used the 133Xe-xenon inhalation technique,Citation22 positron emission tomography,Citation19 inversion recovery perfusion MR technique [20] and blood oxygenation level-dependent (BOLD) signal intensity changes in functional MRI (fMRI).Citation22,Citation23 Recent papers studied the effects of caffeine on cerebral circulation since caffeine ingestion might be a source of errors in functional brain imaging experiments.Citation20,Citation21,Citation23
The present study showed a 6% to 8% statistically nonsignificant caffeine-induced decrease in perfusion. Several other studies reported caffeine-induced cerebral blood flow decreases ranging from 3.4% to 18%Citation19,Citation20,Citation22,Citation24,Citation25 but not consistently in all subjects.Citation22 The reasons for the discrepancies may have various origins. First, the hemodynamic response measured by different techniques (cerebral blood flow, BOLD contrast, or perfusion changes) is not directly comparable. Second, in most if not all studies, the same dose of caffeine was given to the subjects independently of body weight. Conversely, in the present study, the dose of caffeine ingested was adjusted to body weight, ie, 3 mg/kg. The third factor differing amongst the studies is the period of abstinence from caffeine. The latter was similar to the one applied here, ie, about 12 h in several studies,Citation19,Citation23 very short, 2 to 3 hours in other studies,Citation22,Citation24 or much longer, ie, 30 hours.Citation20,Citation21 The period of abstinence may be an important factor in the effect of caffeine on cerebral blood flow, and has been a matter of concern in imaging studies over the last few years.Citation20,Citation21 Indeed, the cerebral blood flow rates and velocities are increased during the withdrawal state, mainly in high usersCitation20,Citation26 and go back to baseline values after about 2 h.Citation26 Therefore the widespread lack of significance in the perfusion values recorded in the present study with and without caffeine may partly reflect the withdrawal state induced by the overnight caffeine deprivation imposed on the subjects. On the other hand, the discrete changes recorded in some brain areas after caffeine indicate the specific changes due to the methylxanthine.
In the present study, on a background of widespread statistically nonsignificant perfusion decrease, discrete increases in perfusion corresponding to specific neuronal activation could be identified. Brain activation was mostly seen in the LC group. In this group, significant activation was recorded in regions known to mediate anxiety like the inferior frontal gyrus-anterior insular cortex, the uncus, the lingual gyrus, and the cerebellum.Citation27,Citation28
Simultaneously, many other regions involved in the regulation of anxiety levels, such as the amygdala, cingulate and orbitofrontal cortex, thalamus, and striatum, were not activated by caffeine. The inferior frontal gyrus-anterior insular cortex seems to play a role in anticipating aversive stimuli and in anxiety and emotion regulation.Citation29 Its activation was observed in different anticipatory anxiety induction protocols,Citation30-Citation32 and was totally different from the claustrum-posterior insular cortex activation observed in pharmacologically induced panic attacks with cardiovascular and visceral symptomatologyCitation32-Citation34 Caffeine is known to be anxiogenic, at low doses in a subset of individuals and at quite large doses in most of the population.Citation35 The activation recorded only in a limited number of areas may reflect the fact that the subjects did not report increased anxiety after ingestion of the caffeinated drink. They could also imply that caffeine may specifically act at some given steps of the anxiety process, for example, at the anterior insular cortex for integration of internal state, parietal cortex for spatially specific associations, but does not reach, at this dose, the sensory-motor integration in thalamus and the initiation of action - since there is no defensive action required - depending on the anterior cingulate cortex.Citation28,Citation36
Brain activation was observed in the internal parietal cortex of LC subjects and in the hypothalamus of HC subjects. These activations relate to changes in vigilance and attention. The parietal cortex is critical for attention and spatial updating. It is involved in visual representations of space in an eye-centered coordinate frame, and in providing a signal for directing the eyes towards these objects.Citation28,Citation37 The hypothalamus mediates many vegetative functions as well as attention and vigilance. In our experience, hypothalamic activations were often associated with changes in attention and vigilance. These activations are in line with the known effects of caffeine on vigilance. Indeed, the pattern of consumption of caffeine throughout the day shows that caffeine is mostly consumed to increase the level of vigilance,Citation35 and caffeine is well known to impair sleep.Citation3,Citation38 Likewise, caffeine focuses available attention and energy on the task to complete, mostly by limiting distracting external stimuli.Citation39
Finally, the anterior insular cortex, which was activated by caffeine, regulates cardiovascular function and respiratory rhythms. Numerous epidemiologic studies have focused on the effects of coffee and caffeine on cardiovascular risk, cholesterol, and blood pressure (for review see ref 40). The data currently available indicate that a
moderate caffeine intake does not adversely affect cardiovascular function. However, a recent meta-analysis on the relationship between coffee, caffeine, and blood pressure reported that caffeine consumption increases blood pressure by a mean value of 4.2 mm for systolic and 2.0 mm Hg for diastolic blood pressure.Citation41 In the present study, the values of systolic blood pressure slightly increased after caffeine, especially in the HC group, but because of the large interindividual variability, this slight change was not significant. In the present study, the main difference in caffeine-induced brain activation between LC and HC subjects was the involvement of hypothalamus which was the single region affected in HC, while perfusion was not affected in hypothalamus when the same amount of caffeine was given to LC. When both groups were pooled, the caffeine-induced brain activation was significant in all areas involved in the two groups.
In the present study, we did not record any brain activation or inhibition in the different components of the brain circuit of dependence. In the presurgical followup of a 20-year-old male epileptic patient with right temporal lobe epilepsy and seizures induced by compulsive smoking, we were incidentally able to show that nicotine induced clear focal brain activation in the nucleus accumbens, the key area involved in addiction and reward, while caffeine (3.5 mg/kg) did not induce change in brain activation in the nucleus accumbens (Marescaux, Namer, and Nehlig, unpublished data). Therefore, these earlier data plus the present data reflect that caffeine at doses representing about two cups of coffee in one sitting does not activate the circuit of dependence and reward and especially not the main target area, the nucleus accumbens.Citation10-Citation12 This lack of effect is present both in light and heavy coffee drinkers who had claimed that they felt “dependent” on coffee. This data is in agreement with our previous data on rats in which the doses of 2.5 and 5.0 mg/kg also failed to activate the circuit of dependence and reward.Citation15 It is also in good agreement with recent reviews reporting that caffeine acts as a mild stimulant that is able to restore mental alertness and wakefulness, mainly in situations with low alertness level.Citation8,Citation42 Therefore, caffeine appears to be different from drugs of dependence like cocaine, amphetamine, morphine, and nicotine, and does not fulfil the common criteria or the scientific definitions to be considered an addictive substance.Citation42
This study was supported by a grant from PEC-ISIC, Vevey (Switzerland), the Hôpitaux Universitaires de Strasbourg and University Louis Pasteur and was approved by the Alsace ethics committee (n° 00/80, 12.09.2000 and n° 02/35, 09.04.2002). All subjects gave their written informed consent prior to participation.
The assistance of the Centre d'Investigations Cliniques (CIC) and pharmacy staff of the Hôpitaux Universitaires de Strasbourg is gratefully acknowledged. We also wish to thank A. Mathis and G. Erb for valuable advice.
REFERENCES
- DebryGedCoffee and Health London, UK: John Libbey 1994
- BaroneJJRobertsHRCaffeine consumptionFood Chem Toxicol1996341191268603790
- FredholmBBBättigKHolmenJNehligAZvartauEActions of caffeine in the brain with special reference to factors that contribute to its widespread usePharmacol Rev.1999518313310049999
- HigdonJVFreiBCoffee and health: a review of recent human researchCrit Rev Food Sci Nutr. 20064610112316507475
- GriffithsRRWoodsonPPCaffeine physical dependence, a review of human and laboratory animal studiesPsychopharmacology1988944374513131789
- JulianoL. MGriffithsRRA critical review of caffeine withdrawal: empirical validation of symptoms and signs, incidence, severity, and associated featuresPsychopharmacology 200417612915448977
- NehligAAre we dependent on coffee and caffeine? A review on human and animal data Neurosci Biobehav Rev.19992356357610073894
- NehligAAre we dependent on coffee and caffeine: an update. in Nehlig A, edCoffee, Tea, Chocolate and the Brain Boca Raton, FL: CRC Press 2004133146
- GriffithsRRMumfordGKCaffeine reinforcement, discrimination, tolerance and physical dependence in laboratory animals and humans. In: Schuster, CR, Kuhar MJ, edsHandbook of Experimental Pharmacology Heidelberg, Germany: Springer Verlag 1996118315341
- KoobGFDrugs of abuse: anatomy, pharmacology and function of reward pathwaysTrends Pharmacol Sci1992131771841604710
- SelfDWNestlerEJMolecular mechanisms of drug reinforcement and addictionAnnu Rev Neurosci1995184634957605071
- Di ChiaraGThe role of dopamine in drug abuse viewed from the perspective of its role in motivationDrug Alcohol Depend199538951377671769
- PontieriFETandaGDi ChiaraGIntravenous cocaine, morphine, and amphetamine preferentially increase extracellular dopamine in the "shell" as compared with the "core" of the rat nucleus accumbensProc Natl Acad Sci USA19959212304123088618890
- PontieriFETandaGOrziFDi ChiaraGEffects of nicotine on the nucleus accumbens and similarity to those of addictive drugsNature19963822552578717040
- NehligABoyetSDose-response study of caffeine effects on cerebral functional activity with a specific focus on dependenceBrain Res.2000858717710700599
- AcquasETandaGDi ChiaraGDifferential effects of caffeine on dopamine and acetylcholine transmission in brain areas of drug-naive and caffeine-pretreated ratsNeuropsychopharmacology20022718219312093592
- De LucaMABassareoVBauerADi ChiaraGCaffeine and accumbens shell dopamineJ Neurochem200710315716317655660
- NikouCHeitzFArmspachJPNamerIJGruckerDRegistration of MR/MR and MR/SPECT brain images by fast stochastic optimization of robust voxel similarity measuresNeuroImage1998830439698573
- CameronOGModellJGHariharanMCaffeine and human cerebral blood flow: a positron emission tomography studyLife Sci.199047114111462122148
- FieldASLaurientiPJYenYFBurdetteJHMoodyDMDietary caffeine consumption and withdrawal: confounding variables in quantitative cerebral perfusion studies?Radiology200322712913512616005
- LaurientiPJFieldASBurdetteJHMaldjianJAet al.Relationship between caffeine-induced changes in resting cerebral perfusion and blood oxygenation level-dependent signalAm J Neuroradiol2003241607161113679279
- MathewRJWilsonWHCaffeine induced changes in cerebral circulationStroke1985168148173901422
- MulderinkTAGitelmanDRMesulamMMParrishTBOn the use of caffeine as a contrast booster for BOLD fMRI studiesNeuroImage200215374411771972
- MathewRJWilsonWHBehavioral and cerebrovascular effects of caffeine in patients with anxiety disordersActa Psychiatr Scand19908217222399815
- RagabSLuntMBirchAThomasPJenkinsonDFCaffeine reduces cerebral blood flow in patients recovering from an ischaemic strokeAge Ageing20043329930315082437
- CouturierEG. MLamanDMvan DuijnM. AJvan DuijnHInfluence of caffeine and caffeine withdrawal on headache and cerebral blood flow velocitiesCephalalgiae199717188190
- KiltsCDKelseyJEKnightBElyTDet al.The neural correlates of social anxiety disorder and response to pharmacotherapyNeuropsychopharmacology2006312243225316525417
- PaulusMPSteinMBAn insular view of anxietyBiol Psychiatry 20066038338716780813
- AugustineJRCircuitry and functional aspects of the insular lobe in primates including humansBrain Res Rev1996222292448957561
- ChuaPKramsMToniIPassinghamRDolan RA functional anatomy of anticipatory anxietyNeuroImage1999956357110334900
- NitschkeJBSarinopoulosIMackiewiczKLSchaefer HS, Davidson RJ. Functional neuroanatomy of aversion and its anticipationNeuroImage20062910611616181793
- SchunckTErbGMathisAGillesCet al.Functional magnetic resonance imaging characterization of CCK-4-induced panic attack and subsequent anticipatory anxietyNeuroImage2006311197120816600640
- ReimanEMRaichleMERobinsEMintunMAet al.Neuroanatomical correlates of a lactate-induced anxiety attackArch Gen Psychiatry1989464935002786401
- CameronOGZubietaJKGrunhausLMinoshimaSEffects of yohimbine on cerebral blood flow, symptoms, and physiological functions in humansPsychosom Med.20006254955910949101
- BriceCFSmithAPEffects of caffeine on mood and performance: a study of realistic consumptionPsychopharmacology (Berlin) 200216418819212404081
- WardakCDuhamelJRLe rôle du cortex pariétal (Gaze control: role of the parietal cortex)Med Sci (Paris)200420899714770370
- HubbardEMPiazzaMPinelPDehaeneSInteractions between number and space in parietal cortexNat Rev Neurosci2005643544815928716
- JacobsonKAGaoZGAdenosine receptors as therapeutic targetsNat Rev Drug Discov.2006524726416518376
- RuijterJLoristMMSnelJDe RuiterMBThe influence of caffeine on sustained attention: an ERP studyPharmacol Biochem Behav200066293710837841
- NawrotPJordanSEastwoodJRotsteinJet al.Effects of caffeine on human healthFood Addit Contam20032013012519715
- NoordzijMUiterwaalCSArendsLRKokFJet al.Blood pressure response to chronic intake of coffee and caffeine: a meta-analysis of randomized controlled trialsJ Hypertens20052392192815834273
- SatelSIs caffeine addictive?- -a review of the literatureAm J Drug Alcohol Abuse20063249350217127537