Abstract
Panic disorder is a common and disabling illness for which treatments are too frequently ineffective. Greater knowledge of the underlying biology could aid the discovery of better therapies. Although panic attacks occur unpredictably, the ability to provoke them in the laboratory with challenge protocols provides an opportunity for crucial insight into the neurobiology of panic. Two of the most well-studied panic provocation challenges are CO2 inhalation and lactate infusion. Although it remains unclear how these challenges provoke panic animal models of CO2 and lactate action are beginning to emerge, and offer unprecedented opportunities to probe the molecules and circuits underlying panic attacks. Both CO2 and lactate alter pH balance and may generate acidosis that can influence neuron function through a growing list of pH-sensitive receptors. These observations suggest that a key to better understanding of panic disorder may lie in more knowledge of brain pH regulation and pH-sensitive receptors.
El trastorno de pánico es una enfermedad común e incapacitante para la cual los tratamientos con gran frecuencia resultan ineficaces. El mayor conocimiento de las bases biológicas podría ayudar al descubrimiento de mejores terapias. Aunque los ataques de pánico son impredecibles, la capacidad de provocarlos en el laboratorio con protocolos de estimulación permite una gran oportunidad para introducirse en la neurobiología del pánico. La inhalación de CO2 y la infusión de lactato son dos de las pruebas de provocación de pánico más estudiadas. Aunque aun no está aclarado cómo provocan pánico estos estímulos, han comenzado a aparecer modelos animales de la acción de CO2 y de lactato, los que permiten oportunidades insospechadas para investigar las moléculas y circuitos que están a la base de los ataques de pánico. Tanto el CO2 como el lactato afectan el balance del pH y pueden generar acidosis, la que puede alterar el funcionamiento neuronal medíante una cantidad creciente de receptores sensibles al pH. Estas observaciones sugieren que una clave para una mejor comprensión del trastorno de pánico puede encontrarse en un mayor conocimiento de la regulación del pH cerebral y en los receptores sensibles al pH.
Le trouble panique est une pathologie courante et invalidante aux traitements trop souvent inefficaces. Une meilleure connaissance de la biologie sous-jacente pourrait faciliter la découverte de traitements plus efficaces. Malgré l'imprévisibilité des attaques de panique, la possibilité de les reproduire au laboratoire par des protocoles de provocation offre une opportunité de compréhension de la neurobiologie de la panique. Deux des épreuves de provocation de panique les mieux étudiées sont l'inhalation de CO2 et la perfusion de lactate. Bien que la façon dont ces épreuves provoquent la panique reste obscure, des modèles animaux de l'action du CO2 et du lactate commencent à émerger et offrent des opportunités sans précédent pour explorer les molécules et les circuits sous-tendant les attaques de panique. Le CO2 et le lactate changent tous les deux l'équilibre du pH et peuvent provoquer une acidose pouvant influer sur la fonction neuronale par l'intermédiaire d'une liste croissante de récepteurs sensibles au pH. Ces observations suggèrent que la clé d'une meilleure compréhension du trouble panique pourrait reposer sur une connaissance plus approfondie de la régulation cérébrale du pH et des récepteurs sensibles au pH.
Introduction
Panic disorder is a common psychiatric illness with a lifetime prevalence of about 4.5 %.Citation1 The hallmark of the disorder is recurring panic attacks, which can appear suddenly and unexpectedly. Panic symptoms include shortness of breath, palpitations, shaking, sweating, and fear of losing control.Citation2 These symptoms resemble those of other serious medical problems and lead some sufferers to think they are having a heart attack or a stroke. One of the most debilitating features of the illness is agoraphobia, a condition in which patients begin to avoid situations and places where a panic attack and the associated discomfort and embarrassment might occur. Consequently, many sufferers learn to avoid daily activities, greatly limiting their productivity and quality of life. Major depression often co-occurs.Citation3 When severe, these symptoms can be debilitating, particularly for the large number of patients who are refractory to current therapies. Identifying new therapies may require understanding of why panic attacks occur and what triggers them, knowledge that is currently lacking (see Box below).
Theories of panic etiology
Although the mechanisms for panic remain obscure, several theories provide useful foundations for conceptualizing the bases of the attacks: (i) Some theories focus on cognitive distortions and misinterpretations of somatic experiences. For example, sensing one's heartbeat may be misinterpreted as an impending heart attack, triggering uncontrolled fear (reviewed in ref 4). (ii) Other theories focus on ventilation. Klein's false suffocation alarm theory highlights the similarities between panic attacks and the powerful fear that suffocation evokes; this theory posits that a “suffocation alarm” is falsely triggered, thus inadvertently producing panic.Citation5 Interestingly, patients with a history of respiratory disease have a greater risk of panic disorder than the general population.Citation6-Citation8 Similarly, panic disorder patients with prominent respiratory symptoms were more likely to have a prior history of respiratory insult.Citation9 Thus, previous experience and adaptive plasticity and/or conditioning might play a role in panic.Citation13 (iii) Growing knowledge of the anatomy underlying fear conditioning led Gorman and others to speculate that a supercharged fear circuit could produce panic in response to a wide variety of arousing stimuli.Citation14,Citation15 This fear circuit is thought to include at least 5 components: (i) Sensory input from viscera via the nucleus of the solitary tract and sensory thalamus, (ii) Processing and conscious control via the prefrontal cortex, cingulate cortex, and insula. (iii) Processing context and fear through the hippocampus and amygdala, (iv) Coordinated output of behavioral, autonomic, and neuroendocrine manifestations from the amygdala via the hypothalamus, periacqueductal gray, locus coeruleus, and parabrachial nucleus.Citation14,Citation16 (v) modulation by monoamines including serotonin and the raphe nuclei.Citation14 Supporting this final component is the well-established benefit of selective serotonin reuptake inhibitors.
Crucial advances might be made if panic attacks could be evoked in the laboratory so that the underlying mechanisms might be deconstructed. This review discusses progress based on this approach, which raises the possibility that brain pH and pH-sensitive receptors may contribute to the pathophysiology of panic disorder.
Panic provocation
Provocation challenges offer potential for unique insights into panic
Panic disorder is relatively unique among psychiatric illnesses, in that symptoms resembling the illness can be provoked by a number of chemicals called panicogens. Because naturally occurring panic attacks are unpredictable, the ability to induce an attack becomes a powerful tool for research. Moreover, the biological mechanisms of the panicogens themselves might tell us a lot about the neurobiology of the illness. Therefore, it has long been hoped that provocation challenges might shed light on the mechanisms underlying panic. Examples of agents with the potential ability to evoke panic attacks include carbon dioxide (CO2),Citation10,Citation11 sodium lactate,Citation12 doxapram,Citation17 cholecyslokinin (CCK-4) and related agonists,Citation18,Citation19 flumazenil,Citation20 caffeine,Citation21 adrenergic agonists (isoproterenol, yohimbine, epinephrine),Citation19 serotonin receptor activators (d-fenfluramine, metachlorophenylpiperazine (m-CPP),Citation19 and perhaps opioid receptor antagonists.Citation22,Citation23 The symptoms provoked by these agents can closely resemble naturally occurring attacks.Citation19,Citation24 Unfortunately, many of these provocation challenges have been studied too little to ascertain their mechanisms of action and relative potency.Citation19,Citation25 Interestingly, one twin study found a high concordance rate for CO2 sensitivity,Citation26 suggesting a genetic etiology, although much remains to be learned about the heritability of panicogen sensitivity.
Rodent models of panic provocation challenges
The technology revolutionizing neurobiological research in rodents is rapidly expanding knowledge of mechanisms underlying behavior. The ability to explore panicogens in animal models provides a powerful research opportunity. However, progress in this area has been limited in comparison with the advancements made in other behavioral models. Yet, several examples are notable. Shekhar and colleagues have developed rodent models of lactate-evoked panicCitation27,Citation28 and have found that orexin-expressing neurons in the hypothalamus play a critical role.Citation29 In another example, doxapram potentiated fear and anxiety-related behaviors in rats and induced expression of the immediate early gene c-Fos in the amygdala.Citation30 A few investigators have also begun to explore the effects of CO2 on fear and anxiety in rodents.Citation31-Citation33 For example, Mongeluzi et al found that high CO2 concentrations can serve as an unconditioned stimulus in Pavlovian fear conditioning.Citation31 Johnson et al observed that CO2 inhalation can induce c-Fos expression in fear circuit structures and may thus activate brain regions thought to be responsible for panic.Citation33 Despite these examples, the mechanisms underlying panicogen action and panic attacks remain largely unknown.
Clinical clues about panicogen action
Perhaps the most well-studied panicogens are CO2 and lactate. CO2 provocation challenges vary between investigators, but generally consist of breathing single or multiple breaths of CO2 at concentrations ranging from 5% to 35%.Citation24 Protocols for lactate provocation challenges typically include intravenous infusion of 0.5 M sodium D,L-lactate up to 10 mg/kg body weight over 20 minutes or until panic occurs.Citation12,Citation19,Citation34 Several observations led investigators to suggest that CO2 and lactate may share mechanisms of action.Citation35 For example, most CO2-sensitive panickers are also lactate-sensitive.Citation36 In addition, CO2 and lactate produce stereotypic responses. In particular both induce prominent ventilatory symptoms, suggesting a degree of neuro anatomical or physiological overlap.Citation37,Citation38 Interestingly, both CO2 and lactate may be more likely to affect panic disorder patients who report strong respiratory symptoms during their naturally occurring attacks.Citation38,Citation39 CO2 and lactate may also induce less hypothalamus-pituitary-adrenal (IIPA) axis activation than other panicogens,Citation40-Citation42 suggesting that their effects may be cortisol-independent. Consistent with this observation, inhibiting Cortisol synthesis failed to prevent CO2 -evoked panic.Citation43 Furthermore, both CO2 , and lactate play prominent roles in metabolism and share the potential to alter systemic acid-base balance.
Panic and acid-base balance
CO2 and brain acidosis
CO2 is constantly produced in the brain and throughout the body as a final product of carbohydrate metabolism. CO2 readily crosses cell membranes and the blood-brain barrier. In a reaction catalyzed by carbonic anhydrase, CO2 is hydrolyzed to carbonic acid (H2CO3), which readily dissociates into HCO3-and H+. The resulting acidosis is thought to be responsible for most of the physiological effects of CO2,Citation44 including stimulating acid-activated respiratory chemoreceptors in the brain stem.Citation45 These chemoreceptors stimulate breathing to expel CO2 and thus correctively raise systemic pH. Inhaling CO2 increases the partial pressure of CO2 in the blood and lowers pH throughout the body. Thus, the CO2 provocation challenges used in psychiatric research are likely to acutely and transiently acidify brain pH.
Lactate and brain acidosis
Endogenous lactate is generated by glucose and glycogen metabolism. In the brain, astrocytes are thought to convert glucose and stored glycogen into lactate, which is exported to the interstitial space where it can be taken up by neurons to produce energy via oxidative metabolism.Citation46 Recent experiments suggest that shuttling lactate to neurons may be crucial for learning and memory.Citation47 The effects of intravenous lactate on systemic pH are more complex than those of CO2. One reason is that intravenously administered lactate can be metabolized by the liver to HCO3-, which might raise blood pH. In addition, in order to cross membranes such as the bloodbrain barrier, lactate requires monocarboxylate transporters (MCTs). Because these MCTs cotransport H+, they effectively move lactic acid, thus providing a mechanism that might lower pH in specific compartments, for example in the central nervous system (CNS). A recent review covers these complex effects of lactate on systemic pH.Citation35 Importantly, intravenous lactate causes hyperventilation,Citation48 suggesting that, like CO2 inhalation, lactate likely reduces pH at ventilatory chemoreceptors and perhaps elsewhere in the brain.Citation49 These observations suggest that CO2 and lactate may share an ability to acidify interstitial pH in the brain. Interestingly, another panicogen, doxapram, may act by a related mechanism. Both doxapram and protons stimulate ventilation and both can inhibit two-pore domain K+ channels (TWIK)-related acid-sensitive K+ (TASK) channels.Citation50,Citation51
pH abnormalities in panic disorder?
Increasing evidence suggests that pH may be abnormally regulated in panic disorder.Citation35,Citation44 Brain pH is largely controlled by the CO2/HCO3- buffering system, which is acutely regulated by breathing. Multiple investigators have reported irregular breathing in panic disorder,Citation52-Citation55 including greater tidal volume variability, which may be due to more frequent sighing.Citation52 Consistent with a persistent breathing irregularity, panic disorder patients exhibit a chronically low end-tidal CO2 Citation56-Citation59 and a compensatory decrease in serum bicarbonate.Citation2,Citation59 Those who exhibit breathing irregularities may also be more likely to have respiratory symptoms during an attack.Citation38,Citation60 Symptom cluster analyses have identified a subtype of panic disorder, in which respiratory symptoms appear to predominate.Citation38,Citation60 Interestingly, the respiratory subtype may be the most sensitive to CO2 Citation38 and lactate.Citation39 This subtype may also respond best to the antidepressant imipramine,Citation60 and may be more likely to be associated with a family history of panic disorder.Citation38 Supporting a role for pH in panic pathophysiology, correcting blood gas abnormalities through breathing control or pharmacology has been suggested to produce clinical improvement.Citation57,Citation59
Endogenous lactate and pH abnormalities in panic disorder
Lactate is a weak acid that can be an independent determinant of pH in biological systems.Citation61 Several studies using 1H-magnetic resonance spectroscopy suggest endogenous lactate levels may be elevated in panic disorder patients. Panic disorder patients had higher lactate levels than controls in response to visual cortex activation,Citation62 following hyperventilation,Citation63 and during lactate-induced panic.Citation49 Fiberoptic biosensor measurements of pH in primates suggest that intravenous lactate infusion reduces brain pH.Citation64 Phosphorus spectroscopy further suggests that the elevated brain lactate in panic disorder patients may change pH buffering capacity.Citation65 It was suggested that a vascular or metabolic abnormality might be responsible for the lactate elevation.Citation49,Citation53,Citation62 Consistent with this view, probands who had a family history of panic and an atypical CO2 ventilatory response were more likely to carry a polymorphism in a gene encoding lactate dehydrogenase, which catalyzes the conversion of lactate to pyruvate.Citation53
CNS chemosensitivity
CO2 and acid chemosensitivity in the CNS
The potential associations between panic disorder, the action of panicogens, and brain pH begs the question of how the brain normally senses and responds to pH change. The majority of research on chemosensitivity in the CNS has focused on respiratory control. Thus, understanding how pH regulates breathing could provide critical insights into panic disorder. Breathing rate and volume are exquisitely sensitive to CO2 in the blood, largely through interstitial pH and activation of pH-sensitive chemoreceptors.Citation45,Citation66 Although the precise sites of CO2-mediated ventilatory control are uncertain, they are thought to lie in the brain stemCitation67 (medulla and pons). Neurons in multiple brain stem sites can be activated by CO2 and low pH, suggesting the relevant chemosensitivity might reside at multiple locations.Citation67-Citation69 The retrotrapezoid nucleus (RTN) of the medulla has been implicated in respiratory chemosensation, particularly the cells expressing Phox2b, which is mutated in congenital hypoventilation syndrome.Citation67,Citation70 Other brainstem nuclei are sensitive to pH and have been implicated in pll-mediated ventilatory control; these regions include the medullary raphe nuclei, nucleus of the tractus solitarius, and locus coeruleus.Citation67,Citation69,Citation71 Thus, multiple chemosensitive sites are possible. The CO2 sensitivity in panic patients, and the associations between panic and ventilation, make it tantalizing to speculate that abnormalities in these chemosensitive neurons and receptors might contribute to panic attacks. Knowledge of pH-sensitive molecules in the brain and their physiological roles is rapidly growing, but much remains to be learned.
pH-sensitive receptors and respiratory chemosensation
Understanding the molecules that underlie pH effects on ventilatory control could pave the way for understanding pH sensitivity in the brain in general. Thus far no single molecule has been found to be responsible for respiratory chemosensation. A number of molecules have the potential to detect falling pH and stimulate breathing.Citation72 Members of the TWIK family are pH-sensitiveCitation73; a subset, the TASK channels, have garnered attention as potential respiratory chemoreceptors. Because TASK channels help maintain membrane voltage near the resting potential, inhibiting these channels increases excitability and the likelihood of generating action potentials. TASK channels can be inhibited by small reductions in extracellular pH. For example, reducing pH by just 1/10th of a unit from pH 7.4 to pH 7.3 inhibits TASK-1.Citation73 TASK-1 and TASK-3 are widely expressed in brain,Citation74 while TASK-2 expression in brain is limited to a few brain stem nuclei, including the retrotrapezoid nucleus (RTN), which has been implicated in pH control of ventilation. Nevertheless, disrupting the genes encoding TASK-1, TASK-2, or TASK-3 in mice failed to eliminate the centrally mediated hypercapnic ventilatory response,Citation74-Citation76 suggesting that the TASK channels are not required. However, some pH-sensitive responses were affected. Loss of TASK-1, TASK-3, or both reduced the pH sensitivity of cultured raphe neurons, but not that of RTN neurons.Citation74 TASK-1 disruption also reduced peripheral chemosensitivity to hypercapnia in the carotid body.Citation75 Additionally, TASK-2 disruption in mice increased the respiratory response to mild hypercapnia (1.5 and 2% CO2), suggesting a modulatory role.Citation76.
pH-sensitive ion channels, G-protein coupled receptors, and intracellular signaling molecules
Besides the TASK channels, a wide number of additional molecules might sense pH in the brain. Examples of pH-sensitive ion channels include transient receptor potential (TRP) channels,Citation77 P2X receptors,Citation78,Citation79 voltage-dependent Ca2+ channels,Citation80 N-methyl-D aspartate (NMDA) receptors,Citation81 acid-sensing ion channels (ASICs),Citation82-Citation84 and inward rectifier K channels.Citation85 Examples of pH-sensitive G-protein coupled receptors include OGR1, GPR4, TDAG8, adenosine Al receptors, and metabotropic P2Y receptors.Citation72,Citation78,Citation86 pH-sensitive intracellular signaling molecules include Pyk2 and soluble adenlyl cyclase (sAC).Citation72 All of these molecules are sensitive enough to detect pH changes that occur during physiology or pathophysiology. Further, all of these molecules have been suggested as candidates for pH chemosensitivity.Citation72,Citation86 Though more investigation is needed, some of these molecules have already been implicated in pH sensing. For example, voltage-dependent Ca2+ channels and NMDA receptors modulate synaptic plasticity in response to changes in extracellular pH.Citation80,Citation81 Adenosine Al receptors, adenosine triphosphate (ATP) receptors (P2X and P2Y), and ASIC1a have been implicated in the ability of CO2 and low pH to inhibit seizure activity.Citation32,Citation78 Recent studies also investigated the potential role in the inward rectifier K+ channel Kir5.1, which is highly sensitive to extracellular pH when heteromerically coupled to Kir4.1. Disrupting Kir5.1 produced abnormal respiration and metabolic acidosis in mice, however central hypercapnic ventilatory responses remained intact. Instead, impaired sensory afferent nerve conduction was thought to be responsible for the abnormal respiratory phenotype.Citation85
Effects of chemosensation on arousal and emotion circuits
pH-sensitive respiratory chemosensors in the brain stem medulla and pons comprise a powerful mechanism for controlling systemic CO2 and pH. Slow or shallow breathing acidifies systemic pH, while fast or deeper breathing raises systemic pH, making it more alkaline. There may also be a need for higher level (more rostral) brain structures to monitor pH, for example to produce appropriate cognitive or behavioral responses to rising CO2. Rising CO2 heralds the potential threat of suffocation, a terrifying situation that demands immediate detection and action to ensure survival. The clusters of pH-sensitive neurons in the medulla and pons that stimulate breathing might communicate this need for action to higher level structures. Alternatively, it might be advantageous if sites above the medulla and pons sensed pH more directly.Citation68,Citation69 A prominent example is midbrain serotonergic neurons. Midbrain raphe neurons are highly pH-sensitive and increase firing when CO2 rises and pH falls.Citation87 These neurons are well positioned to deliver serotonin (5-HT) to forebrain, cortical, and subcortical structures and thus alter mood and cognition in response to CO2 and low pH. In sleep, a rising CO2 and falling pH might signal the need to reposition the airway or to relieve an obstruction. During sleep CO2 inhalation causes wild-type mice to wake up, whereas CO2 fails to wake mice lacking pH-sensitive serotonin neurons.Citation88 Thus, dysfunction of these neurons might play a critical role in sudden infant death syndrome,Citation89 where a failure to wake may lead to suffocation. Neurons in even higher order brain areas are also activated by low pH, including orexin-expressing neurons in the hypothalamus.Citation90 These orexin-expressing neurons have been implicated in narcolepsy and arousal, and are positioned to influence diverse physiological functions including adaptive behaviors, metabolism, respiration, and panic.Citation29 Recently the amygdala was also implicated in CO2 and acid chemosensation and CO2-evoked fear.Citation32
The amygdala is a chemosensor that detects CO2 and acidosis to elicit fear
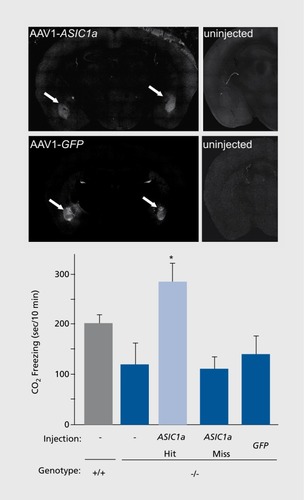
It is well established that the amygdala integrates sensory input from other brain structures to orchestrate fear behavior; however, the amygdala itself was not previously known to act as a pH sensor. Ziemann et al suspected this possibility after observing that the acid sensing ion channel-1a (ASIC1a) was abundantly expressed in the basolateral amygdala and other fear circuit structures,Citation91,Citation92 and it was found that breathing 10% CO2 lowered pH to levels sufficiently to activate ASIC1a in amygdala neurons.Citation32 To test CO2 triggered fear in mice, four behavioral paradigms were developed: (i) CO2-evoked freezing; (ii) CO2-potentiated center avoidance in the open field; (iii) CO2 aversion; and (iv) CO2-enhanced fear conditioning.Citation32 Genetically disrupting or pharmacologically inhibiting ASIC1a reduced fear-like behavior in these paradigms.Citation32 Particularly striking was the freezing behavior, which is often used as a correlate of fear and panic in mice. Like other fear-evoking stimuli, breathing 10% CO2 induced a dramatic freezing response in wild-type mice. Disrupting or inhibiting ASIC1a significantly blunted this response.Citation32 To test whether the amygdala itself might sense pH, acidic artificial cerebrospinal fluid was microinfused into the amygdala to lower pH to ~6.8 from normal pH 7.35. Acidifying the amygdala produced freezing behavior in wild-type mice that resembled the freezing evoked by CO2 inhalation. Interestingly, in the ASIC1a knockout mice amygdala acidosis induced little or no freezing. The freezing deficit was likely specific to low pH because the ASIC1a knockouts froze normally when the amygdala was electrically stimulated. Finally, the authors tested whether ASIC1a in the amygdala might be sufficient to produce CO2-evoked freezing. Restoring ASIC1a expression to the amygdala of ASIC1a-null mice with an ASIC1a-expressing adeno-associated virus corrected the CO2-evoked freezing deficit ( .)Together these findings suggest that the amygdala itself can act as a chemosensor. These experiments further identify ASIC1a as key molecular mediator of this chemosensitive response.
Interoception and false alarms
It is intriguing that a brain structure that mediates fear has a chemosensory role. The ability to sense or monitor internal bodily states (interoception) is a common human experience, ranging from vague sensations to powerful and uncontrollable emotions. Often the language needed to communicate these sensations seems inadequate. Yet, interoceptive sensations may be critical for survival. pH might be one of a variety of signals that could produce interoceptive sensations by activating pH-sensitive receptors in the brain to evoke adaptive responses. The survival value of rapidly detecting CO2 to prevent suffocation seems clear. Nearly 20 years ago Donald Klein drew from this observation to hypothesize that the suffocation detection system might be falsely triggered to produce panic attacks.Citation5 Conceivably, heightened pH sensitivity could constitute such a false alarm.
Summary
We don't yet know why panic attacks occur. Nor do we completely understand why those who suffer panic attacks are hypersensitive to panicogens. However, the potential ability of CO2 and lactate, the two most well-studied panicogens, to alter brain pH suggests that pH chemosensation could be instrumental. Acid-sensitive molecules are widely distributed in fear circuit structures and elsewhere in the brain. Consistent with this observation, a variety of brain sites have been implicated in pH chemosensation including brain stem respiratory nuclei, midbrain raphe neurons, hypothalamus, and amygdala. However, a number of questions remain. For example, what specific role(s) do each of these pH-sensitive sites and pH-sensitive molecules play? Could there be additional sources of acidosis and pH fluctuation besides CO2 or lactate that might activate these chemosensory pathways? Finally, might genetic or epigenetic variability in chemosensation lead to panic disorder or other psychiatric and neurological illnesses? That we are now in a position to ask these questions is in itself a significant advance. As we continue to learn more about CO2 and pH chemosensation in the brain, the answers to these questions may be within reach. Moreover, an improved understanding of pH signaling and dysregulation might very well lead to an entirely new avenue of therapeutic intervention.
The author thanks Drs Michael Welsh, George Richerson, Margaret Price, William Coryell, Jess Fiedorowicz, and Tom Brashers-Krug for discussions and helpful comments. Dr Wemmie's work is supported by NIMH 1R01MH085724-01, NINDS 1R01NS0641 59-01 A109, Department of Veteran's Affairs Merit Review Program, McKnight Endowment Fund for Neuroscience, and the DANA Foundation.
REFERENCES
- KesslerRC.ChiuWT.JinR.RuscioAM.ShearK.WaltersEE.The epidemiology of panic attacks, panic disorder, and agoraphobia in the National Comorbidity Survey Replication.Arch Gen Psychiatry.20066341542416585471
- American Psychiatric Association.Diagnostic and Statistical Manual of Mental Disorders. 4th ed, Text Revision. Washington, DC: American Psychiatric Association;2000
- KesslerRC.ChiuWT.DernierO.MerikangasKR.WaltersEE.Prevalence, severity, and comorbidity of 12-month DSM-IV disorders in the National Comorbidity Survey Replication.Arch Gen Psychiatry.20056261762715939839
- MoynihanJE.GevirtzRN.Respiratory and cognitive subtypes of panic. Preliminary validation of Ley's model.Behav Modif.20012555558311530716
- KleinDF.False suffocation alarms, spontaneous panics, and related conditions. An integrative hypothesis.Arch Gen Psychiatry.1993503063178466392
- SmollerJW.PollackMH.OttoMW.RosenbaumJF.KradinRL.Panic anxiety, dyspnea, and respiratory disease. Theoretical and clinical considerations.Am J Respir Crit Care Med.19961546178680700
- GiardinoND.CurtisJL.AbelsonJL.et al.The impact of panic disorder on interoception and dyspnea reports in chronic obstructive pulmonary disease.Biol Psychol.20108414214620176074
- LiverrnoreN.SharpeL.MckenzieD.Panic attacks and panic disorder in chronic obstructive pulmonary disease: a cognitive behavioral perspective.Resp Med.201010412461253
- BouwerC.SteinDJ.Association of panic disorder with a history of traumatic suffocation.Am J Psychiatry.1997154156615709356565
- DruryAN.The percentage of carbon dioxide in the alveolar air, and the tolerance to accumulating carbon dioxide in case of co-called “irritable heart”.Heart.19187165173
- CohenME.WhitePD.Life situations, emotions, and neurocirculatory asthenia (anxiety neurosis, neurasthenia, effort syndrome).Psychosom Med.19511333535714892184
- PittsFN.McClureJN.Lactate metabolism in anxiety neurosis.N Engl J Med.1967227132913366081131
- LeyR.The modification of breathing behavior. Paviovian and operant control in emotion and cognition.Behav Modif.19992344147910467892
- GormanJM.KentJM.SullivanGM.CoplanJD.Neuroanatomical hypothesis of panic disorder, revised.Am J Psychiatry.200015749350510739407
- NardiAE.FreireR.ZinWA.Panic disorder and control of breathing.Resp Physio Neurobiol.2009167133143
- MezzasalmaMA.ValençaAM.LopesFL.NascimentoI.ZinWA.NardiAE.[Neuroanatomy of panic disorder].Rev Bras Psiquiatr.20042620220615645066
- LeeYJ.CurtisGC.WegJG.AbelsonJL.ModellJG.CampbellKM.Panic attacks induced by doxapram.Biol Psychiatry.1993332952978471685
- de Montigny, C Cholecystokinin tetrapeptide induces panic-like attacks in healthy volunteers. Preliminary findings.Arch Gen Psychiatry.1989465115172730276
- EsquivelG.SchruersK.GriezE.Experimental models: panic and fear. In: Blanchard RJ, Griebel G, Nutt D, eds.Handbook of Anxiety and Fear. Amsterdam, the Netherlands: Elsevier2008413434
- NuttDJ.GlueP.LawsonC.Wilson S. Flumazenil provocation of panic attacks. Evidence for altered benzodiazepine receptor sensitivity in panic disorder.Arch Gen Psychiatry.1990479179252171449
- CharneyDS.HeningerGR.JatlowPI.Increased anxiogenic effects of caffeine in panic disorders.Arch Gen Psychiatry.1985422332432983630
- MaremmaniI.MariniG.FornaiF.Naltrexone-induced panic attacks.Am J Psychiatry.19981554479501763
- PrêterM.LeeSH.PetkovaE.VannucciM.KimS.KleinDF.Controlled cross-over study in normal subjects of naloxone-preceding-lactate infusions; respiratory and subjective responses: relationship to endogenous opioid system, suffocation false alarm theory and childhood parental loss.Psychol Med.20114138539320444308
- RassovskyY.KushnerMG.Carbon dioxide in the study of panic disorder: issues of definition, methodology, and outcome.J Anx Disord.200317132
- PeskindER.JensenCF.PascualyM.et al.Sodium lactate and hypertonic sodium chloride induce equivalent panic incidence, panic symptoms, and Hypernatremia in panic disorder.Biol Psychiatry.199844100710169821565
- BellodiL.PernaG.CaldirolaD.ArancioC.BertaniA.Di BellaD.CO2induced panic attacks: a twin study.Ami J Psychiatry.199815511841188
- SajdykTJ.ShekharA.Sodium lactate elicits anxiety in rats after repeated GABA receptor blockade in the basolateral amygdala.Eur J Pharmacol.200039426527310771292
- ShekharA.SajdykTJ.GehlertDR.RainnieDG.The amygdala, panic disorder, and cardiovascular responses.Ann N Y Acad Sci.200398530832512724167
- JohnsonP.TruittW.FitzSD.et al.A key role for orexin in panic anxiety.Nat Med.20101611111520037593
- SullivanGM.ApergisJ.GormanJM.LeDouxJE.Rodent doxapram model of panic: behavioral effects and c-Fos immunoreactivity in the amygdala.Biol Psychiatry.20035386387012742673
- MongeluziDL.RoselliniRA.LeyR.CaldaroneBJ.StockHS.The conditioning of dyspneic suffocation fear. Effects of carbon dioxide concentration on behavioral freezing and analgesia.Behav Modif.20032762063614531158
- ZiemannAE.AllenJE.DahdalehNS.et al.The amygdala is a chemosensor that detects carbon dioxide and acidosis to elicit fear behavior.Cell.20091391012102119945383
- JohnsonP.FitzSD.HollisJH.et al.Induction of c-Fos in 'panic/defence'related brain circuits following brief hypercarbic gas exposure.J Psychopharmacol (Oxford).201125263620080924
- GormanJ.LiebowitzM.FyerA.et al..Lactate infusions in obsessivecompulsive disorder.Am J Psychiatry.19851428644014509
- EsquivelG.SchruersKR.MaddockRJ.ColasantiA.GriezEJ.Acids in the brain: a factor in panic?J Psychopharmacol.20102463964719460873
- GormanJM.FyerMR.GoetzR.et al.Ventilatory physiology of patients with panic disorder.Arch Gen Psychiatry.19884531392827599
- GoetzRR.GormanJM.DillonDJ.et al.Do panic disorder patients indiscriminately endorse somatic complaints?Psychiatry Res.1989292072132798599
- FreireR.PernaG.NardiAE.Panic disorder respiratory subtype: psychopathology, laboratory challenge tests, and response to treatment.Harv Rev Psychiatry.20101822022920597592
- GoetzRR.KleinDF.GormanJM.Symptoms essential to the experience of sodium lactate-induced panic.Neuropsychopharmacology.1996143553668703304
- SinhaSS.CoplanJD.PineDS.MartinezJA.KleinDF.GormanJM.Panic induced by carbon dioxide inhalation and lack of hypothalamic-pituitary-adrenal axis activation.Psychiatry Res.199986939810397411
- van DuinenMA.SchruersKR.MaesM.GriezEJ.CO2 challenge induced HPA axis activation in panic.Int J Neuropsychopharmacol.20071079780417076937
- AbelsonJL.LiberzonI.Dose response of adrenocorticotropin and Cortisol to the CCK-B agonist pentagastrin.Neuropsychopharmacology.19992148549410481831
- BelgorodskyA.KnyazhanskyL.LoewenthalU.ArbelleJ.CohenH.BenjaminJ.Effects of the Cortisol synthesis inhibitor metyrapone on the response to carbon dioxide challenge in panic disorder.Depress Anxiety.20052114314816001448
- ZandbergenJ.PolsH.De LoofC.LousbergH.GriezE.Effect of hypercapnia and other disturbances in the acid-base-balance on panic disorder.Hillside J Clin Psychiatry.1989111851972488058
- RichersonGB.Cellular mechanisms of sensitivity to pH in the mammalian respiratory system. In: Kaila K, Ransom BR, eds.pH and Brain Function. New York, NY: Wiley- Liss, Inc;199850953319381963
- PellerinL.Bouzier-SoreAK.AubertA.et al.Activity-dependent regulation of energy metabolism by astrocytes: an update.Glia.2007551251126217659524
- SuzukiA.SternSA.BozdagiO.et al.Astrocyte-neuron lactate transport is required for long-term memory formation.Cell.201114481082321376239
- OlssonM.HoHP.AnnerbrinkK.ThyleforsJ.ErikssonE.Respiratory responses to intravenous infusion of sodium lactate in male and female Wistar rats.Neuropsychopharmacology.200227859112062909
- DagerSR.FriedmanSD.HeideA.et al.Two-dimensional proton echoplanar spectroscopic imaging of brain metabolic changes during lactateinduced panic.Arch Gen Psychiatry.19995670779892258
- GottenJF.KeshavaprasadB.LasterMJ.EgerEl.YostCS.The ventilatory stimulant doxapram inhibits TASK tandem pore (K2P) potassium channel function but does not affect minimum alveolar anesthetic concentration.Anesth Analg.200610277978516492828
- BaylissDA.SiroisJE.TalleyEM.The TASK family: two-pore domain background K+ channels.Moi Interv.2003320521914993448
- AbelsonJL.WegJG.NesseRM.CurtisGC.Persistent respiratory irregularity in patients with panic disorder.Biol Psychiatry.20014958859511297716
- PhilibertRA.NelsonJJ.SandhuHK.CroweRR.CoryellWH.Association of an exonic LDHA polymorphism with altered respiratory response in probands at high risk for panic disorder.Am J Med Genet.2003117B1 11712555227
- CoryellW.DindoL.FyerA.PineDS.Onset of spontaneous panic attacks: a prospective study of risk factors.Psychosom Med.20066875475717012529
- CoryellW.FyerA.PineD.MartinezJ.ArndtS.Aberrant respiratory sensitivity to CO(2) as a trait of familial panic disorder.Biol Psychiatry.20014958258711297715
- PappLA.MartinezJM.KleinDF.et al.Respiratory psychophysiology of panic disorder: three respiratory challenges in 98 subjects.Am J Psychiatry.1997154155715659356564
- MeuretAE.WilhelmFH.RitzT.RothWT.Feedback of end-tidal pCO(2) as a therapeutic approach for panic disorder.J Psychiatr Res.20074256056817681544
- HegelMT.FergusonRJ.Psychophysiological assessment of respiratory function in panic disorder: evidence for a hyperventilation subtype.Psychosom Med.1997592242309178332
- GormanJM.FyerAJ.RossDC.et al.Normalization of venous pH pCO2, and bicarbonate levels after blockade of panic attacks.Psychiatry Res.19851457653921997
- BriggsAC.StretchDD.BrandonS.Subtyping of panic disorder by symptom profile.Br J Psychiatry.19931632012098075912
- StewartPA.Independent and dependent variables of acid-base control.Respir Physiol.19783392627857
- MaddockRJ.BuonocoreMH.CopelandLE.RichardsAL.Elevated brain lactate responses to neural activation in panic disorder: a dynamic 1H-MRS study.Mol Psychiatry.20091453754518180759
- DagerSR.StraussWL.MarroKl.RichardsTL.MetzgerGD.ArtruAA.Proton magnetic resonance spectroscopy investigation of hyperventilation in subjects with panic disorder and comparison subjects.Am J Psychiatry.19951526666727726305
- DagerSR.YimJB.KhalilGE.ArtruAA.BowdenDM.KennyMA.Application of a novel fiber-optic biosensor in situ to investigate the metabolic effect of lactate infusion.Neuropsychopharmacology.1995123073137576007
- FriedmanSD.MathisCM.HayesC.RenshawP.DagerSR.Brain pH response to hyperventilation in panic disorder: preliminary evidence for altered acid-base regulation.Am J Psychiatry.200616371071516585448
- NattieE.Central chemoreceptors, pH, and respiratory control. In: Kaila K, Ransom BR, eds.pH and Brain Function. New York, NY: Wiley-Liss, Inc.1998535560
- MarinaN.AbdalaAP.TrappS.et al.Essential role of Phox2b-expressing ventrolateral brainstem neurons in the chemosensory control of inspiration and expirât ion.J Neurosci.201030124661247320844141
- NattieG.LiA.Multiple central chemoreceptor sites: cell types and function in vivo.AdvExp Med Biol.2008605343347
- NattieE.LiA.Central chemoreception is a complex system function that involves multiple brain stem sites.J Appl Physiol.20091061464146618467549
- AmielJ.LaudierB.Attié-BitachT.et al.Polyalanine expansion and frameshift mutations of the paired-like homeobox gene PHOX2B in congenital central hypoventilation syndrome. Nat Genet.200333459461 12640453
- RichersonGB.WangW.HodgesMR.DohleCI.Diez-SampedroA.Homing in on the specific phenotype(s) of central respiratory chemoreceptors.Exp Physiol.200590259266 discussion 266-25915728134
- TresguerresM.BuckJ.LevinLR.Physiological carbon dioxide, bicarbonate, and pH sensing.PflugersArch.2010460953964
- BucklerKJ.Two-pore domain k(+) channels and their role in chemoreception.Adv Exp Med Biol.2010661153020204721
- MulkeyDK.TalleyEM.StornettaRL.et al.TASK channels determine pH sensitivity in select respiratory neurons but do not contribute to central respiratory chemosensitivity.J Neurosci.200727140491405818094244
- TrappS.AllerMl.WisdenW.GourineAV.A role for TASK-1 (KCNK3) channels in the chemosensory control of breathing.J Neurosci.2008288844885018753386
- GestreauC.HeitzmannD.ThomasJ.et al.Task2 potassium channels set central respiratory CO2 and 02 sensitivity.Proc Natl Acad Sci USA.20101072325233020133877
- CuiN.ZhangX.TadepalliJS.et al.Involvement of TRP channels in the CO2 chemosensitivity of locus coeruleus neurons.J Neurophysiol.20111052791280121430274
- DullaCG.DobelisP.PearsonT.FrenguelliBG.StaleyKJ.MasinoSA.Adenosine and ATP link PCO2 to cortical excitability via pH.Neuron.2005481011102316364904
- BirdsongWT.FierroL.WilliamsFG.et al.Sensing muscle ischemia: coincident detection of acid and ATP via interplay of two ion channels.Neuron.20106873974921092862
- DeVriesSH.Exocytosed protons feedback to suppress the Ca(2+) current in mammalian cone photoreceptors.Neuron.2001321107111711754841
- MakaniS.CheslerM.Endogenous alkaline transients boost postsynaptic NMDA receptor responses in hippocampal CA1 pyramidal neurons.J Neurosci.2007277438744617626204
- WaldmannR.LazdunskiM.H+-gated cation channels: neuronal acid sensors in the NaC/DEG family of ion channels.Curr Opin Neurobiol.199884184249687356
- KrishtalO.The ASICs: signaling molecules? Modulators?Trends Neurosci.20032647748312948658
- WemmieJA.PriceMP.WelshMJ.Acid-sensing ion channels: advances, questions and therapeutic opportunities.Trends Neurosci.20062957858616891000
- TrappS.TuckerSJ.GourineAV.Respiratory responses to hypercapnia and hypoxia in mice with genetic ablation of Kir5.1 (Kcnj16).Exp Physiol.20119645145921239463
- McGuireJ.HermanJP.GhosalS.EatonK.SalleeFR.SahR.Acid-sensing by the T cell death-associated gene 8 (TDAG8) receptor cloned from rat brain.Biochem Biophys Res Comm.200937842042519501050
- SeversonCA.WangW.PieriboneVA.DohleCI.RichersonGB.Midbrain serotonergic neurons are central pH chemoreceptors.Nat Neurosci.200361139114014517544
- BuchananG.RichersonG.Central serotonin neurons are required for arousal to CO2.Proc Natl Acad Sci US A.20101071635416359
- BuchananG.RichersonG.Role of chemoreceptors in mediating dyspnea.Respir Physiol Neurobiol.200916791919118647
- WilliamsRH.JensenLT.VerkhratskyA.FuggerL.BurdakovD.Control of hypothalamic orexin neurons by acid and CO2.Proc Natl Acad Sci USA.2007104106851069017563364
- WemmieJA.AskwithCC.LamaniE.CassellMD.FreemanJHJ.WelshMJ.Acid-sensing ion channel 1 is localized in brain regions with high synaptic density and contributes to fear conditioning.J Neurosci.2003235496550212843249
- CoryellM.ZiemannAE.WestmorelandPJ.et al.Targeting ASIC1a reduces innate fear and alters neuronal activity in the fear circuit.Biol Psychiatry.2007621140114817662962