Abstract
In humans, influenza A viruses cause yearly outbreaks with high morbidity and excess fatality rates as a direct effect. Placed in its ecological niche, however – in dabbling ducks – avian influenza virus (AIV) induces quite a mild disease. It is when the virus crosses the species barrier that pathogenic traits are attributed to infection. When infecting phylogenetically more distant species (i.e. chicken and turkeys), the AIV can cause high morbidity and may in some cases change the virus into a highly pathogenic variant with nearly 100% fatality rate. Being a very adaptable virus, these spill-over events are frequent and numerous species are susceptible to influenza virus. When a subtype of AIV that has not previously infected humans crosses the species barrier, adapts to humans, and spreads easily, a pandemic event is imminent. There is no cure for influenza infection and vaccination is a cumbersome endeavor so, currently, the strategy when a pandemic strikes is damage control. The interest in AIV ecology has increased dramatically since the beginning of the millennium as a key factor for preventive work for future pandemics. This review gives a broad overview of influenza A virus ecology: in the natural host, accidental hosts, new endemic hosts, and humans.
Background
Novel introductions of influenza viruses into the human population from the animal kingdom continue to be a major health problem worldwide. During the last decade, influenza research has intensified to previously unseen magnitudes and together with this, also the interest in the inclusive virology research that focuses on influenza not only as a human pathogen but as an entity that is part of an ecosystem. With an increased knowledge of the dynamics of the ecosystem and the interplay of the parts therein, the goal is for human science to be able to more accurately predict new pandemics and take appropriate preventive countermeasures.
Introduction
Placed in its ecological niche – the dabbling ducks – influenza A virus is a benign disease Citation1. However, it is a very adaptable virus and it has been able to infect and adapt to a wide range of hosts () Citation2Citation3. The disease associated with infection shows a broad range of symptoms, depending in part on the genetic properties of the virus but also on which species of host is infected Citation4Citation5Citation6. In the natural host, no signs of infection can be identified by ocular inspection, while other bird species and mammals are more severely affected with symptoms ranging from very mild to very severe and ultimately death. It was first identified as an animal disease in 1878, when Eduardo Perroncito described a disease affecting poultry in northern Italy. Observations from this study describe an easily transmitted, initially mild disease, which increased in pathogenicity over time and in the end killed virtually all domestic fowl in the area. ‘Fowl plague’, as the disease would be called, was proved to be a viral disease in 1901 but not identified as influenza virus until 1955 Citation7. Since it was first described in 1878, the highly pathogenic avian influenza (HPAI) virus has caused repeated outbreaks of fowl plague at irregular intervals Citation8.
Fig. 1. Illustration of the host range of influenza A virus with the natural reservoir of influenza A virus, accidental hosts, and the subtypes that have been identified in the different groups. Illustration by Rebecca Rönnmark and Eric Gisaeus.
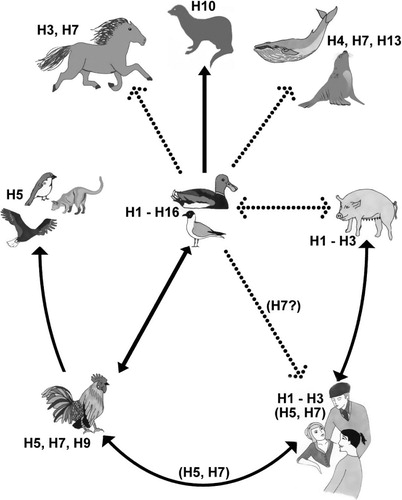
Despite thorough research where the molecular functions of the virus have been investigated in detail, influenza A virus continues to be a common human pathogen and each year the endemic seasonal flu results in mortality peaks and wide-spread morbidity with vast economic consequences Citation9. Rather than having been able to control this infection, we have adapted ourselves and our behavior to minimize its damage when the flu season strikes Citation10Citation11. Random introductions of novel viruses from the animal kingdom also continue to be a major health and economical problem for the human population, and several pandemic events have occurred during the 19th and 20th centuries including the notorious ‘Spanish Flu’ Citation12Citation13Citation14. It was long believed that this first recorded pandemic was the result of a direct transmission of a highly pathogenic avian virus to humans without intermediate hosts, though this belief has recently been called to question Citation15Citation16. Later pandemics did not occur in a direct fashion but used pigs, which are permissive to both avian and human adapted viruses, as mixing vessels () Citation17.
Influenza A virus in wild birds
It is widely accepted that all influenza virus strains infecting mammalian species originate from wild birds Citation2Citation3. All subtypes have been detected in isolates from avian species, and the evidence for the existence of a wild bird reservoir is strong throughout the world Citation18Citation19. Low pathogenic influenza A virus strains have been isolated from more than 105 species from 26 different families of birds and almost all isolates come from the families Anseriformes and to a lesser extent Charadriformes and Laridae Citation20. These families include birds such as ducks, geese, swans, waders, and gulls; although different species evolutionarily speaking, they share the trait of being adapted to life in an aquatic environment. Isolations of low pathogenic virus strains from pure land-dwelling birds are, on the contrary, rare. Further evidence that wild birds constitute a reservoir for influenza A virus comes from studies on viral evolution, which have shown limited evolution in wild ducks over time. It has therefore been suggested that influenza A virus exists in an evolutionary stasis in the reservoir species Citation2Citation21. This suggestion is supported by analysis of strains recovered from wild ducks that have been preserved in museums since the early 20th century, which show almost no antigenic drift when compared to modern avian strains Citation22. However, genetic studies on sequences from the contemporary gene pool show a mutation rate that is in contrast to the historical findings Citation23.
Propagation
Dabbling ducks are very susceptible to, and easily become infected with, avian strains of influenza A virus through the intake of contaminated food and water. During the period of infection, a large amount of virus is shed in the duck feces usually for about 7 days, but shedding has been recorded for as long as 21 days Citation24Citation25. Tracheal shedding also occurs, though this route is probably more relevant where the social behavior of certain species makes fecal–oral transmission difficult Citation26. Influenza A virus of different subtypes has been isolated in concentrations of up to 102,8 EID50/ml of water from unconcentrated lake water in lakes where wild ducks congregate Citation27Citation28. The species preference of influenza A virus is likely to be determined by the mode of transmission (i.e. by the fecal–oral route via water). Since virus is shed by infected birds in high quantities into an environment where it can survive for an extended time, the feeding and social behavior of the reservoir species is an important determinant for virus propagation. Species that feed in shallow calm waters where influenza A virus is found in the highest concentrations run the highest risk of becoming infected. Other species belonging to the Anseriformes, like swans and geese, graze to a larger extent on land in pastures and agricultural fields, which may lead to less efficient transmission though they are equally susceptible to the virus as their dabbling cousins. Scavenger species, such as raptors that may feed on diseased birds, are also susceptible but not to take part in efficient transmission as they do not dwell in water. In mallards, about one third of the population is replaced each year Citation29Citation30. As prevalence of influenza A virus has been shown to be higher in juvenile than in adult birds, the input of juvenile and, thus, immunologically naïve birds is most likely of key importance for upholding the number of susceptible birds in the population. However, one of the enigmas of influenza A virus ecology is how so many subtypes can circulate in the wild bird populations and persist from year to year when some of these subtypes are isolated rarely, and since the prevalence in the studied bird populations differ greatly between studied species, place, and time of year. While some subtypes are frequently isolated, others have been isolated only rarely in specific places or in specific species such as the H13 and H16 subtypes that have almost exclusively been isolated from gulls.
Pathogenesis
It is commonly believed that all birds are susceptible to AIV infection, although some species are more resistant than others. Infections by low pathogenic strains in ducks have traditionally been considered benign, as there are no evident clinical signs of disease. Though there is indication of coupling infection to a decreased body mass, aquatic birds do not appear to be severely affected by the disease and the infection does not seem to limit an infected bird's interaction with other birds or the environment Citation31. Nor does infection with low pathogenic strains seem to limit mallard capability for migration flights that could transport the virus long distances to new susceptible flocks, though the contrary has been shown for swans Citation32. However, it is difficult to evaluate if the birds are completely unaffected or actually become sick in a subtle way. Few studies have been conducted in this area, but histopathological signs of mild pneumonia have been shown in ducks even though no other signs of disease were evident Citation33. The HPAI viruses behave differently, however, and strains that are highly pathogenic for chickens may cause milder to no disease and different signs of disease in other species Citation5Citation33Citation34Citation35.
Studies using experimental infection have also shown that ducks may be re-infected with the same strain after 2 months indicating that the protection of acquired immunity is poor Citation25. Using a recapture scheme for wild ducks and sampling the same duck at regular intervals has shown re-infection at even shorter intervals Citation31. Other bird species such as chicken, pheasant, turkey, and quail mount a humoral response with high levels of IgM and IgY production Citation36. However, in large-scale studies, juvenile ducks are found to be infected with influenza A virus more frequently than adult birds, indicating some sort of acquired immunity or improved immune response Citation2.
Influenza A virus in domestic fowl
As previously discussed, AIV may enter into domestic bird populations as low pathogenic strains that only cause mild disease. Subtypes H5 and H7, however, may evolve into highly pathogenic strains. The fast mutation rate that is displayed in domestic fowl is probably due to the extremely high propagation rates in dense flocks.
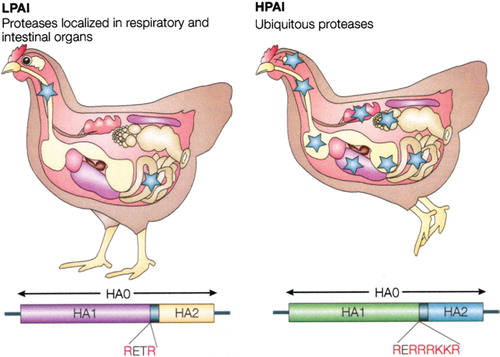
Influenza A virus causes a wide spectrum of symptoms in reared birds, from mild illness to a highly contagious and fatal disease resulting in severe epidemics. Highly pathogenic avian influenza is characterized by severe illness, rapid death, and a mortality in the affected populations that approaches 100% within 72 hours. Many different species of domestic birds including chickens, turkeys, quail, and ostriches are susceptible to epidemics of rapidly fatal influenza Citation7Citation37. The main difference between infection with highly pathogenic virus strains and low pathogenic virus strains is systemic contra-localized infection, the cleavage of the hemagglutinin (HA) by ubiquitous proteases that expunges the restriction to cells in the respiratory tract Citation36. Several mutations may add to the pathogenicity of strains causing HPAI but the accumulation of basic amino acids (AAs) at the cleavage site is diagnostic for highly pathogenic disease (). Outbreaks of HPAI are often difficult to control since the virus can persist and remain active for some time in the environment and because it is highly transmissible. In areas with dense poultry populations and/or limited resources for surveillance and control such as Mexico, South-East Asia, and Africa, outbreaks are even harder to contain.
Fig. 2. Illustration of localized low pathogenic avian influenza (LPAI) infection versus systemic HPAI infection. Adapted by permission from Macmillan Publishers Ltd: [Nature Reviews Microbiology] ‘Influenza: lessons from past pandemics, warnings from current incidents’ copyright 2005.
The human host
Only subtypes H1N1 and H3N2 of influenza A virus follow an epidemiological pattern in humans and are considered endemic, though a H2N2 persisted for a long time. Influenza A viruses can be isolated somewhere in the world every month and the infection is sustained and perpetuated in the human population Citation38. The virus strains that circulate in humans mainly cause respiratory disease and preferably infect the epithelium lining the airways. A spread of a subtype of influenza A virus, which the human population has not experienced before and, thus, has no immunity against, may be rapid and cause concurrent outbreaks around the globe resulting in a pandemic. The severity of a pandemic may vary depending on the strain, and pandemic strains may behave differently to the seasonal epidemic strains.
Other mammalian hosts
Influenza A virus is able to infect several mammalian species and in some cases create endemic propagation. This has been shown both experimentally and in nature as described for the species below Citation39. Highly pathogenic virus strains, such as the currently circulating H5N1 virus that originates from South-East Asia, have shown an increased host range and are able to infect many species that had previously not been considered vulnerable. Thus, the range of species-infectivity is heavily dependent on strain type. Pigs are frequently infected by influenza A viruses and there are specific swine-adapted strains. However, pigs are also susceptible both to human- and avian-adapted virus strains, which can be explained by the fact that the respiratory epithelium of pigs express both α2,3 and α2,6-linked sialic acid (SA) Citation17. Avian virus strains of different subtypes have been found in pigs on a number of occasions. Influenza A virus strains in horses are thought to be of avian origin. Different subtypes have been found to infect horses and antigenic drift creates distinct lineages within the subtypes. At least two subtypes have created stable lineages: H7N7 and H3N8 Citation40Citation41Citation42Citation43. Some strains have been suggested to be recent introductions from wild birds Citation44. Historically, canine species have not been a significant carrier of influenza virus. However, in 2004, an outbreak in racing greyhounds was caused by a H3N8 influenza A virus and was found to be an equine influenza A virus variant that had adapted to spread in canines Citation45. This triggered further investigation among dog breeders in the United States and found serologic evidence of common influenza infection and also virus isolates Citation46. During the recent outbreak in South-East Asia, a surveillance investigation has isolated H5N1 influenza virus from dogs and has also found that antibodies to H5N1 are common in Thai dogs suggesting that they have previously been infected Citation47. Feline species were not considered particularly susceptible to influenza virus prior to the recent outbreak of avian influenza H5N1 that started in South-East Asia in 2003. However, after 50 captive tigers and leopards became ill and died after having been fed infected chicken carcasses several investigations were performed Citation48Citation49. It was shown that there was not only direct transmission from the contaminated food but also probable transmission between tigers Citation50. Experimental infection of domestic cats has shown that cats infected with the H5N1 highly pathogenic strain develop lethal systemic infection and excrete virus in both the respiratory and digestive tract secretions. The cats in the experiment could also infect each other Citation51Citation52. In Europe, cats have also been found to be infected by the H5N1 virus in areas where there have been outbreaks in wild birds Citation53. Mink and ferrets have been found to be susceptible to influenza A virus and have been used in experiments since 1933 Citation54Citation55. When infected with human-type influenza virus, the symptoms displayed are very similar to those of humans: respiratory symptoms like sneezing and coughing, decreased appetite with following weight loss, lethargy, and fever Citation56. As a disease model, ferrets can be argued to be the best to mimic human disease, though mice tend to be more commonly used simply because ferrets are much more difficult to handle Citation57. Infection of seals with influenza A virus has been reported on several occasions, and there is good reason to believe this is not an uncommon event. In 1979–1980, seals off Cape Cod in the eastern United States died of hemorrhagic pneumonia Citation58. The causative agent of disease was found to be influenza A virus of the subtype H7N7. The virus contained avian-like genes but behaved as a mammalian strain Citation59. During the autopsies and handling of experimentally infected seals, people handling the animals developed conjunctivitis from influenza infection Citation60. In a subsequent outbreak among seals during the season 1982–1983, another even more avian-like virus was recovered from seals suffering from pneumonia. This virus belonged to the H4N5 subtype Citation61. Further surveys of seals in the area have also found H3N3 virus strains to be present in seals Citation62Citation63. Seals have also been shown to be infected by influenza B virus of human origin Citation64.Whales have been found infected on several occasions, but difficulties in surveillance of these animals and high costs has made it difficult to determine the frequency of occurrence of these events, though it does not seem common Citation65Citation66Citation67. Analysis show the most probable route of introduction has been directly from birds to the aquatic mammals Citation66Citation68.
Implications for future introductions of AIV
Low pathogenic influenza A virus strains do not seem to hinder dabbling ducks from migrating. Thus, these virus strains may be carried over large distances by the birds in a relay pattern where one bird carries the virus a short distance and another carries it further. Until the present outbreak of HPAI H5N1 that started in Asia, it was not believed that wild birds could be infected with highly pathogenic virus strains and still perform long distance migrations. As some species of ducks have shown a high resistance to these strains, this belief has had to be reviewed. After the HPAI H5N1 had somehow been exported to Russia, Europe, and Africa in 2006, sudden satellite outbreaks in wild birds showed that transport of highly pathogenic strains by wild birds is a reality. Knowledge of influenza ecology and epidemiology thus becomes a key factor in the pre-pandemic work.
Elementary strategy planning in any battle has one basic goal: Do not get caught off guard. The ability to foresee an outbreak allows pandemic control systems to focus containment on relevant areas in advance. Farm animals, for example, are the common interface between zoonotic diseases in the wild and humans. Primary introduction of influenza A virus into poultry and domestic animal holdings are likely due to fecal contamination by wild birds either directly by contamination of the holdings or indirectly through contaminated water supplies or feed. Holdings where wild birds and domestic birds share the same habitat due to agricultural practices are at the highest risk for outbreaks, suggesting that wild bird transmission is the most common route Citation69. When a subtype that can be deemed a risk for humans is detected, such as the highly pathogenic H5N1, knowledge of how it may travel with migratory birds as well as by human transport provides opportunities for quick outbreak control. If a problematic subtype is suspected to arrive at a specific area during a specific period, regulatory government authorities may take preventive action, for example, by issuing grazing restrictions for farm animals to limit their exposure to the expected pathogen.
Conflict of interest and funding
This study was funded by the Swedish Research Council and the Swedish Research Council FORMAS. Statistical input and AIV surveillance support were provided by Ottenby Birding Station.
References
- Jourdain E, Gunnarsson G, Wahlgren J, Latorre-Margalef N, Bröjer C, Sahlin S, et al.. Influenza virus in a natural host, the mallard: experimental infection data. PLoS ONE. 2010; 5: e8935.
- Webster RG, Bean WJ, Gorman OT, Chambers TM, Kawaoka Y. Evolution and ecology of influenza A viruses. Microbiol Rev. 1992; 56: 152–79.
- Olsen B, Munster VJ, Wallensten A, Waldenström J, Osterhaus ADME, Fouchier RAM. Global patterns of influenza A virus in wild birds. Science. 2006; 312: 384–88.
- Horimoto T, Kawaoka Y. Influenza: lessons from past pandemics, warnings from current incidents. Nat Rev Microbiol. 2005; 3: 591–600.
- Kishida N, Sakoda Y, Isoda N, Matsuda K, Eto M, Sunaga Y, et al.. Pathogenicity of H5 influenza viruses for ducks. Arch Virol. 2005; 150: 1383–92.
- Isoda N, Sakoda Y, Kishida N, Bai GR, Matsuda K, Umemura T. Pathogenicity of a highly pathogenic avian influenza virus, A/chicken/Yamaguchi/7/04 (H5N1) in different species of birds and mammals. Arch Virol. 2006; 151: 1267–79.
- Capua I, Mutinelli F. A colour atlas and text on avian influenza. Papi editore. Bologna, 2001
- WHO. Avian influenza A(H5N1)-update 31: situation (poultry) in Asia: need for a long-term response, comparison with previous outbreaks. 2004. available from: http://www.who.int/csr/don/2004_03_02/en/.
- Franco-Paredes C, Hernandez-Ramos I, Del Rio C, Alexander KT, Tapia-Conyer R, Santos-Preciado JI. H1N1 influenza pandemics: comparing the events of 2009 in Mexico with those of 1976 and 1918–1919. Arch Med Res. 2009; 40: 669–72.
- Robinson KR. The role of nursing in the influenza epidemic of 1918–1919. Nurs Forum. 1990; 25: 19–26.
- Webster RG. The importance of animal influenza for human disease. Vaccine. 2002; 20: S16–S20.
- Hope-Simpson RE, Golubev DB. A new concept of the epidemic process of influenza A virus. Epidemiol Infect. 1987; 99: 5–54.
- Del Rio C, Hernandez-Avila M. Lessons from previous influenza pandemics and from the Mexican response to the current influenza pandemic. Arch Med Res. 2009; 40: 677–80.
- Morens DM, Taubenberger JK, Jeffery K, Harvey HA, Memoli MJ. The 1918 influenza pandemic: lessons for 2009 and the future. Crit Care Med. 2010; 38: e10–e20.
- Reid AH, Fanning TG, Janczewski TA, Lourens RM, Taubenberger JK. Novel origin of the 1918 pandemic influenza virus nucleoprotein gene. J Virol. 2004; 78: 12462–70.
- Antonovics J, Hood ME, Howell Baker C. Molecular virology: was the 1918 flu avian in origin?. Nature. 2006; 440: E9.; discussion E9–E10.
- Ito T, Couceiro JNSS, Kelm S, Baum LG, Krauss S, Castrucci MR, et al.. Molecular basis for the generation in pigs of influenza A viruses with pandemic potential. J Virol. 1998; 72: 7367–73.
- Alexander DJ. A review of avian influenza in different bird species. Vet Microbiol. 2000; 74: 3–13.
- Fouchier RA, Munster V, Wallensten A, Bestebroer TM, Herfst S, Smith D, et al.. Characterization of a novel influenza A virus hemagglutinin subtype (H16) obtained from black-headed gulls. J Virol. 2005; 79: 2814–22.
- Stallknecht DE, Shane SM. Host range of avian influenza virus in free-living birds. Vet Res Commun. 1988; 12: 125–41.
- Bean WJ, Schell M, Katz J, Kawaoka Y, Naeve C, Gorman O, et al.. Evolution of the H3 influenza virus hemagglutinin from human and nonhuman hosts. J Virol. 1992; 66: 1129–38.
- Reid AH, Fanning TG, Slemons RD, Janczewski TA, Dean J, Taubenberger JK. Relationship of pre-1918 avian influenza HA and NP sequences to subsequent avian influenza strains. Avian Dis. 2003; 47: 921–25.
- Chen R, Holmes EC. Avian influenza virus exhibits rapid evolutionary dynamics. Mol Biol Evol. 2006; 23: 2336–41.
- Webster RG, Yakhno M, Hinshaw VS, Bean WJ, Murti KG. Intestinal influenza: replication and characterization of influenza viruses in ducks. Virology. 1978; 84: 268–78.
- Kida H, Yanagawa R, Matsuoka Y. Duck influenza lacking evidence of disease signs and immune response. Infect Immun. 1980; 30: 547–53.
- Ellstrom P, Latorre-Margalef N, Griekspoor P, Waldenström J, Olofsson J, Wahlgren J, et al.. Sampling for low-pathogenic avian influenza A virus in wild Mallard ducks: oropharyngeal versus cloacal swabbing. Vaccine. 2008; 26: 4414–16.
- Hinshaw VS, Webster RG, Turner B. Water-bone transmission of influenza A viruses?. Intervirology. 1979; 11: 66–68.
- Ito T, Okazaki K, Kawaoka Y, Takada A, Webster RG, Kida H. Perpetuation of influenza A viruses in Alaskan waterfowl reservoirs. Arch Virol. 1995; 140: 1163–72.
- Bentz P.-G. Studies on some urban Mallard Anas platyrhynchos populations in Scandinavia. Part I: causes of death, mortality and longevity among Malmö Mallards as shown by ringing recoveries. Fauna Norv. Ser. C, Cinclus. 1985; 8: 44–56.
- Gunnarsson G, Elmberg J, Dessborn L, Jonzén N, Pöysä H, Valkama J. Survival estimates, mortality patterns, and population growth of Fennoscandian mallards Anas platyrhynchos. Annales Zoologici Fennici. 2008; 45: 483–95.
- Latorre-Margalef N, Gunnarsson G, Munster VJ, Fouchier RAM, Osterhaus ADME, Elmberg J, et al.. Effects of influenza A virus infection on migrating Mallard ducks. Proceedings of the Royal Society B-Biological Sciences. 2009; 276: 1029–36.
- van Gils JA, Munster VJ, Radersma R, Liefhebber D, Fouchier RAM, Klaassen M. Hampered foraging and migratory performance in swans infected with low-pathogenic avian influenza A virus. PLoS One. 2007; 2: e184.
- Cooley AJ, Van Campen H, Philpott MS, Easterday BC, Hinshaw VS. Pathological lesions in the lungs of ducks infected with influenza A viruses. Vet Pathol. 1989; 26: 1–5.
- Alexander DJ, Allan WH, Parsons DG, Parsons G. The pathogenicity of four avian influenza viruses for fowls, turkeys and ducks. Res Vet Sci. 1978; 24: 242–47.
- Perkins LE, Swayne DE. Susceptibility of laughing gulls (Larus atricilla) to H5N1 and H5N3 highly pathogenic avian influenza viruses. Avian Dis. 2002; 46: 877–85.
- Suarez DL, Schultz-Cherry S. Immunology of avian influenza virus: a review. Dev Comp Immunol. 2000; 24: 269–83.
- Perez DR, Webby RJ, Hoffmann E, Webster RG. Land-based birds as potential disseminators of avian mammalian reassortant influenza A viruses. Avian Dis. 2003; 47: 1114–17.
- Cox NJ, Subbarao K. Global epidemiology of influenza: past and present. Annu Rev Med. 2000; 51: 407–21.
- Hinshaw VS, Webster RG, Easterday BC, Bean WJ Jr. Replication of avian influenza A viruses in mammals. Infect Immun. 1981; 34: 354–61.
- Berg M, Desselberger U, Abusugra IA, Klingeborn B, Linné T. Genetic drift of equine 2 influenza A virus (H3N8), 1963–1988: analysis by oligonucleotide mapping. Vet Microbiol. 1990; 22: 225–36.
- Guo Y, Wang M, Zheng GS, Li WK, Kawaoka Y, Webster RG. Seroepidemiological and molecular evidence for the presence of two H3N8 equine influenza viruses in China in 1993–94. J Gen Virol. 1995; 76: 2009–14.
- Oxburgh L, Klingeborn B. Cocirculation of two distinct lineages of equine influenza virus subtype H3N8. J Clin Microbiol. 1999; 37: 3005–9.
- Ozaki H, Shimizu-Nei A, Sugita S, Sugiura T, Imagawa H, Kida H. Antigenic variation among equine H3N8 influenza virus hemagglutinins. Jpn J Vet Res. 2001; 48: 177–86.
- Guo Y, Wang M, Kawaoka Y, Gorman O, Ito T, Saito T, et al.. Characterization of a new avian-like influenza A virus from horses in China. Virology. 1992; 188: 245–55.
- Yoon KJ, Cooper VL, Schwartz KJ, Harmon KM, Kim WI, Janke BH, et al.. Influenza virus infection in racing greyhounds. Emerg Infect Dis. 2005; 11: 1974–76.
- Payungporn S, Crawford PC, Kouo TS, Chen L-M, Pompey J, Castleman WL, et al.. Influenza A virus (H3N8) in dogs with respiratory disease, Florida. Emerg Infect Dis. 2008; 14: 902–8.
- Butler D. Thai dogs carry bird-flu virus, but will they spread it?. Nature. 2006; 439: 773.
- Keawcharoen J, Oraveerakul K, Kuiken T, Fouchier RA, Amonsin A, Payungporn S. Avian influenza H5N1 in tigers and leopards. Emerg Infect Dis. 2004; 10: 2189–91.
- Amonsin A, Payungporn S, Theamboonlers A, Thanawongnuwech R, Suradhat S, Pariyothorn N, et al.. Genetic characterization of H5N1 influenza A viruses isolated from zoo tigers in Thailand. Virology. 2006; 344: 480–91.
- Thanawongnuwech R, Amonsin A, Tantilertcharoen R, Damrongwatanapokin S, Theamboonlers A, Payungporn S. Probable tiger-to-tiger transmission of avian influenza H5N1. Emerg Infect Dis. 2005; 11: 699–701.
- Kuiken T, Rimmelzwaan G, van Riel D, van Amerongen G, Baars M, Fouchier R. Avian H5N1 influenza in cats. Science. 2004; 306: 241.
- Rimmelzwaan GF, van Riel D, Baars M, Bestebroer TM, van Amerongen G, Fouchier RA, et al.. Influenza A virus (H5N1) infection in cats causes systemic disease with potential novel routes of virus spread within and between hosts. Am J Pathol. 2006; 168: 176–83.; quiz 364.
- ECDC. Editorial team. Further spread of avian influenza in Europe, detection in French farmed birds and German cat. Euro Surveill. 2006; 11: pii = 2911. Available from: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId = 2911.
- Smith W, Andrewes CH, Laidlaw PP. A virus obtained from influenza patients. Lancet. 1933: 66–68.
- Okazaki K, Yanagawa R, Kida H. Contact infection of mink with 5 subtypes of avian influenza virus. Brief report. Arch Virol. 1983; 77: 265–69.
- Bodewes R, Rimmelzwaan GF, Osterhaus ADME. Animal models for the preclinical evaluation of candidate influenza vaccines. Expert Rev Vaccines. 2010; 9: 59–72.
- Maher JA, DeStefano J. The ferret: an animal model to study influenza virus. Lab Anim (NY). 2004; 33: 50–53.
- Geraci JR, St Aubin DJ, Barker IK, Webster RG, Hinshaw VS, Bean WJ, et al.. Mass mortality of harbor seals: pneumonia associated with influenza A virus. Science. 1982; 215: 1129–31.
- Webster RG, Hinshaw VS, Bean WJ, Van Wyke KL, Geraci JR, St Aubin DJ, et al.. Characterization of an influenza A virus from seals. Virology. 1981; 113: 712–24.
- Webster RG, Geraci J, Petursson G, Skirnisson K. Conjunctivitis in human beings caused by influenza A virus of seals. N Engl J Med. 1981; 304: 911.
- Hinshaw VS, Bean WJ, Webster RG, Rehg JE, Fiorelli P, Early G, et al.. Are seals frequently infected with avian influenza viruses?. J Virol. 1984; 51: 863–65.
- Callan RJ, Early G, Kida H, Hinshaw VS. The appearance of H3 influenza viruses in seals. J Gen Virol. 1995; 76: 199–203.
- Ohishi K, Kishida N, Ninomiya A, Kida H, Takada Y, Miyazaki N, et al.. Antibodies to human-related H3 influenza A virus in Baikal seals (Phoca sibirica) and ringed seals (Phoca hispida) in Russia. Microbiol Immunol. 2004; 48: 905–9.
- Osterhaus AD, Rimmelzwaan GF, Martina BE, Bestebroer TM, Fouchier RA. Influenza B virus in seals. Science. 2000; 288: 1051–53.
- Ridgway SH. Reported causes of death of captive killer whales (Orcinus orca). J Wildl Dis. 1979; 15: 99–104.
- Hinshaw VS, Bean WJ, Geraci J, Fiorelli P, Early G, Webster RG. Characterization of two influenza A viruses from a pilot whale. J Virol. 1986; 58: 655–56.
- Nielsen O, Clavijo A, Boughen JA. Serologic evidence of influenza A infection in marine mammals of arctic Canada. J Wildl Dis. 2001; 37: 820–25.
- Mandler J, Gorman OT, Ludwig S, Schroeder E, Fitch WM, Webster RG, et al.. Derivation of the nucleoproteins (NP) of influenza A viruses isolated from marine mammals. Virology. 1990; 176: 255–61.
- Gilbert M, Chaitaweesub P, Parakamawongsa T, Premashthira S, Tiensin T, Kalpravidh W, et al.. Free-grazing ducks and highly pathogenic avian influenza, Thailand. Emerg Infect Dis. 2006; 12: 227–34.