Abstract
The deleterious and sometimes fatal outcomes of bacterial infectious diseases are the net result of the interactions between the pathogen and the host, and the genetically tractable fruit fly, Drosophila melanogaster, has emerged as a valuable tool for modeling the pathogen–host interactions of a wide variety of bacteria. These studies have revealed that there is a remarkable conservation of bacterial pathogenesis and host defence mechanisms between higher host organisms and Drosophila. This review presents an in-depth discussion of the Drosophila immune response, the Drosophila killing model, and the use of the model to examine bacterial–host interactions. The recent introduction of the Drosophila model into the oral microbiology field is discussed, specifically the use of the model to examine Porphyromonas gingivalis–host interactions, and finally the potential uses of this powerful model system to further elucidate oral bacterial-host interactions are addressed.
Introduction
The use of invertebrate animal models has provided tremendous insight into the pathogen–host interactions of many human pathogens, and has revealed that many aspects of these interactions in higher host organisms are conserved in invertebrates. One of these animal models, the genetically tractable fruit fly, Drosophila melanogaster (Drosophila), has been well established as a model for studying the host–pathogen interactions of bacterial Citation1 Citation2 Citation3 Citation4 Citation5 Citation6 Citation7 Citation8 Citation9 , fungal Citation10 Citation11 Citation12 Citation13 Citation14 Citation15 Citation16 Citation17 and viral pathogens, and prior to the sequencing of the mosquito genome and the subsequent development of genetic tools, the malaria parasite Citation23 Citation24 Citation25 Citation26 ; the model has also been used to probe the host defense response to infection.
This article will first provide an overview of the Drosophila model and address the advantages and drawbacks of the model for studying pathogen–host interactions. Next, the Drosophila immune response will be described in detail, and the various ways that the model has been used to study bacterial–host interactions will be addressed. Finally, the studies that used the Drosophila model to investigate Porphyromonas gingivalis–host interactions will be reviewed, followed by a discussion of the potential contributions that can be made using this model, toward a better understanding of oral bacteria–host interactions.
D. melanogaster as a model for studying host–pathogen interactions
Numerous studies have revealed that there is significant homology between the Drosophila immune response and the mammalian innate immune response (discussed in depth below). The absence of an adaptive immune response permits the study of the interactions between pathogens and the host's innate immune response in isolation. Drosophila are affordable to breed, are easy to handle, have a short generation time (10–14 days depending on the ambient temperature), have a clear endpoint (death), and can be used in quantities large enough to permit statistical analysis of the data. The Drosophila genome is fully sequenced Citation27 Citation28 Citation29 , and numerous well-developed genetic tools are available for the manipulation and analysis of Drosophila responses [reviewed in Citation30 Citation31]. Transposon mutagenesis has been used to successfully create loss-of-function mutants of at least 53% of Drosophila genes, with the ultimate goal of inactivating all genes Citation32 Citation33. Mutations are usually linked to visually identifiable markers, e.g. eye color or wing morphology, to allow for easy identification of mutant animals. Technologies for transgenic expression of genes in Drosophila are well developed Citation34 Citation35 Citation36 Citation37 and have been enhanced by the development of expression systems like upstream activation sequence (UAS)/GAL4 (yeast transcription activator) Citation38 Citation39 that allow for temporally and spatially regulated expression; linking transgenes with reporter genes like lacZ (β galactosidase) or gfp (green fluorescent protein) allows gene expression to be monitored. Mutant and transgenic Drosophila lines are readily available at stock centers and have been extensively used to probe the interactions between pathogens and the Drosophila host. Drosophila loss-of-function immune response gene mutants have been used to examine the roles of the genes in the response to infection with various pathogens [Citation11 Citation16 Citation19 Citation40 Citation41 Citation42 Citation43 and ], and transgenic Drosophila have been used to monitor the activation of immune response pathways upon infection Citation44 Citation45 and to examine the effects of transgenically expressed pathogen proteins on the host Citation21 Citation26 Citation46. Microarray and proteomic platforms as well as RNA interference (RNAi) lines and libraries have been developed and used to perform genome-wide analyses of Drosophila responses, in whole animals Citation41 Citation47 Citation48 Citation49 Citation50 Citation51 Citation52 Citation53 Citation54 Citation55 Citation56 and in the well-established Drosophila cell culture lines, Schneider-2 (S2, embryonic-derived phagocytic cell) and malignant blood neoplasm (mbn-2) Citation56 Citation57 Citation58 Citation59 . As an estimated 50% of Drosophila genes have mammalian homologs Citation60, results from these and other studies are relevant to mammals. A comprehensive collection of Drosophila information can be found online at FlyBase (http://www.flybase.net) Citation61, including but not limited to gene annotation information, stock availability, images, references, and investigator contact information. Other databases containing Drosophila information include Berkeley Drosophila Genome Project (BDGP), Drosophila Interactions Database (DroID), and FlyView.
It is important to note that as Drosophila are not natural hosts for most of the human pathogens studied with this model and lack homologs for many mammalian immune response features, e.g. adaptive immunity, care should be taken when translating findings to mammals. Drosophila are usually reared at room temperature (25°C), and they can be incubated at temperatures of up to 30°C during infection experiments without affecting survival, although males become sterile at this temperature. Above 30°C, Drosophila physiological processes begin to deteriorate, and they are rapidly killed at 37°C Citation62, which is the optimal growth temperature for most human pathogens. In addition, some pathogen virulence genes are selectively expressed at 37°C, which could result in the absence of a virulence phenotype in the Drosophila model. Although the Drosophila hemolymph (blood) is not involved in respiration, it receives oxygen via the trachea, which likely makes it an inhospitable environment for obligate anaerobes. The Drosophila model may be more suitable for studying aerobic and facultatively anaerobic microbes; however, the model has been successfully used to examine the interactions between the obligately anaerobic oral bacterium P. gingivalis and the host. Even though P. gingivalis did not multiply in the flies, the bacterium viably persisted in them for up to 60 h after infection, likely aided by its high degree of aerotolerance Citation63.
In summary, the availability of a large number of well-developed tools for manipulating Drosophila genetics and analyzing Drosophila immune responses make it a powerful model for studying pathogen–host interactions, and numerous studies have convincingly demonstrated that strong correlation exists between microbial pathogenesis in mammals and in Drosophila.
The Drosophila immune system
Drosophila rely solely on an innate immune system to combat infecting microbes, and like mammals, they detect the presence of invading microbes using pattern recognition receptors, which recognize conserved microbial motifs and activate a response that is specific for the type of invading microbe [see Citation64 for a recent review].
Humoral immune response
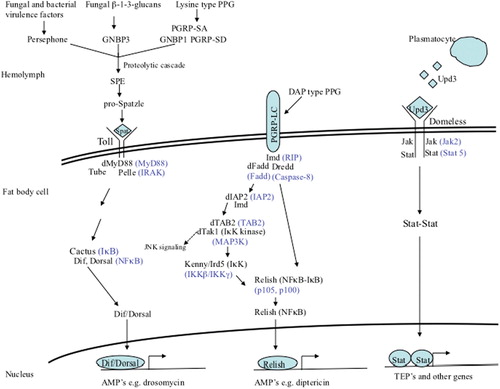
The Drosophila humoral response is mediated by three signaling pathways: the Imd (immune deficiency), Toll, and JAK/STAT (janus kinase/signal transducer and activator of transcription) pathways (). The hallmark of this response is the transient synthesis and release of antimicrobial peptides (AMPs) by the fat body – the major immune responsive tissue in Drosophila and the functional equivalent of the mammalian liver – directly into the hemolymph, where they accumulate to their effective concentrations. The Toll and Imd pathways are major regulators of Drosophila immune response genes and have been very well studied [see Citation65 for a recent in depth review].
The Imd pathway
The Imd signaling pathway () is homologous to the mammalian tumor necrosis factor receptor 1 signaling pathway, though they differ at the level of detection/activation. This pathway regulates both systemic and local (in response to natural infection) AMP expression, by fat body cells and gut epithelial cells, respectively, in response to primarily Gram-negative bacterial infections [ Citation66 Citation67 Citation68 also see ].
Activation and signal transduction
The pathway is activated by the direct detection of monomeric or polymeric meso-diaminopimelic (DAP)-type (Gram-negative type) peptidoglycan (PPG) by the transmembrane receptor, peptidoglycan recognition protein-LC (PGRP-LC) Citation69 Citation70. Binding of PPG by PGRP-LC leads to the intracellular recruitment of the adaptor molecule Imd Citation71 Citation72 Citation73 (receptor interacting protein, RIP, homolog), and the subsequent recruitment of Drosophila Fas-associated death domain protein (dFADD, FADD homolog) and the caspsase-8 homolog DREDD to the Imd/PGRP-LC complex Citation74. Drosophila inhibitor of apoptosis-2 (dIAP2) Citation75 and transforming-growth-factor-β activating kinase (dTAK1, IκB-kinase-kinase) Citation68 are also recruited, and dTAK1 becomes activated by TAK1-binding protein 2 (TAB2). dTAK1 phosphorylates the IκB-kinase complex (IKKβ/Ird5 and IKKγ/Kenny) Citation76, which in turn phosphorylates the dual domain nuclear factor kappa B (NFκB) family protein Relish Citation77 Citation78 Citation79 . Relish is cleaved, possibly by DREDD, which separates the inhibitory ankyrin domain from the Rel-homology domain Citation80. The phosphorylated Rel domain then translocates into the nucleus where it activates the transcription of Imd regulated genes Citation81 (discussed below).
Downregulation
To prevent the deleterious effects of an unchecked inflammatory response, the Imd pathway is regulated on multiple levels. Several PGRPs function as N-acetylmuramoyl-l-alanine amidases that scavenge and degrade PPG, converting it into non-immunostimulatory fragments. These include PGRP-SC1, -SC2 Citation82, -SB1 Citation83, and –LB, which is itself upregulated by the pathway creating a negative feedback loop Citation84. PGRP-LF is a membrane-bound receptor that sequesters DAP-type PPG and prevents it from binding to PGRP-LC and activating Imd signaling Citation85. Two proteins Caspar (Fas-associated factor 1 homolog) and Dnr1 block DREDD mediated cleavage of Relish Citation86 Citation87. Pirk/Rudra/PIMS (PGRP-LC-interacting inhibitor of Imd signaling) functions in a negative feedback loop to disrupt Imd/PGRP-LC binding and inhibit Imd signaling Citation88 Citation89 Citation90 , and the Ras/MAP kinase [mitogen-activated protein kinase (MAPK)] pathway negatively regulates Imd pathway activation by modulating the expression of Pirk/Rudra/PIMS Citation91. Finally, a ubiquitin-proteasome complex represses Imd signaling by degrading one of the pathway components, likely Relish Citation92.
The Toll pathway
The Toll signaling pathway () is homologous to the mammalian Toll/IL-1 receptor-signaling pathway, although unlike the mammalian receptors Drosophila Toll does not directly recognize bacterial components. The pathway regulates systemic AMP expression by the cells of the fat body, primarily in response to fungal and Gram-positive bacterial infections [ Citation93 Citation94 Citation95 Citation96 Citation97 , also see ].
Activation and signal transduction
The pathway is activated in response to the detection of lysine-type (Gram-positive type) PPG (LPPG) by circulating recognition molecules PGRP-SA Citation95 and Gram-negative binding protein 1 (GNBP1) Citation98, which function in a complex Citation99, and PGRP-SD Citation100. The Toll pathway is also activated in response to the detection of fungal cell wall β-1-3-glucans by GNBP3 or by the activation of the Drosophila protease Persephone by fungal- and bacterial-secreted virulence factors Citation101 Citation102 Citation103 Citation104 . A common proteolytic cascade is activated downstream of the recognition molecules Citation105 that culminates in the activation of Spatzle processing enzyme (SPE), which cleaves and activates the cytokine Spatzle Citation106. Activated Spatzle binds to the cell transmembrane receptor, Toll, and induces the receptor's dimerization, which recruits the adaptors, Drosophila myeloid differentiation factor 88 (dMyD88, homolog of mammalian MyD88), Tube (serine-threonine kinase), and Pelle (homolog of mammalian IL-1R associated kinase) intracellularly Citation96 Citation107. Subsequent to Pelle activation Cactus (inhibitor of kappa B, IκB) becomes phosphorylated and is then degraded, freeing the (NFκB) family proteins Dif and Dorsal to translocate into the nucleus and activate the expression of Toll-regulated genes Citation108 Citation109 Citation110 Citation111 . A functional redundancy exists between Dif and Dorsal in the control of immune response gene expression (as measured by drosomycin induction) in Drosophila larvae Citation112, but not in adults where Dif alone is sufficient for the induction of defensin and drosomycin Citation113, and the presence of either protein is sufficient for the induction of cecropin Citation94. Also, deformed epidermal autoregulatory growth factor 1 (Deaf1) is required downstream of Dorsal and Dif to activate genes (e.g. drosomycin) in response to fungal infections Citation114.
Downregulation
Like the Imd pathway, the Toll pathway is negatively regulated on multiple levels. Toll pathway activation induces the expression of Cactus (NFκB inhibitor) Citation115 and WntD (Wnt inhibitor of Dorsal) Citation116, creating negative feedback loops. Drosophila Ubc9 (dUbc9) also inhibits Toll pathway signaling at the level of Dif and Dorsal Citation117. Finally, a serine protease inhibitor (Spn1) acts upstream of SPE to downregulate Toll signaling in response to fungal infections, in a negative feedback manner Citation118.
Toll- and Imd-regulated genes
The Toll and Imd pathways regulate the expression of a subset of Drosophila genes that are induced upon septic injury Citation52 Citation53 Citation56. There is clearly some functional overlap and synergy in gene regulation between the pathways, as revealed by microarray- Citation53 and RNAi-analyses Citation119, the detection of Dif-Relish heterodimers in fly extracts Citation120, and the observation that Pseudomonas aeruginosa, Staphylococcus aureus, Listeria monocytogenes and Ehrlichia chafeensis activate both signaling pathways Citation4 Citation121 Citation122 Citation123 . The most well characterized Drosophila immune effectors regulated by the pathways are the AMPs, of which there are seven classes. Cecropin (antibacterial) Citation124 and Defensin (antibacterial) Citation125 form ion channels in the cytoplasmic membrane Citation126 Citation127, diptericin (antibacterial) Citation128 and drosomycin (antifungal) Citation129 Citation130 act on the cytoplasmic membrane causing lysis Citation131 Citation132, Attacin (antibacterial) Citation133 interferes with outer membrane synthesis Citation133, Drosocin (antibacterial) Citation134 interferes with the activities of DnaK Citation135, and the mode of action of Metchnikowin (anti-bacterial and –fungal) Citation136 has not yet been determined. The Toll and Imd signaling pathways together regulate most of the AMPs, but diptericin and drosomysin are primarily regulated by Imd and Toll, respectively, and are, therefore, used to monitor pathway activation Citation53 Citation66 Citation137. Microarray analysis showed that 283 of 400 previously identified Citation52 Drosophila immune response genes are regulated by either one or both pathways. In addition to the AMPs and pathway regulators, the proteins encoded by these genes are involved in iron metabolism, opsonization, melanization, iron sequestration, coagulation, reactive oxygen species (ROS) production, Jun kinase (JNK) pathway signaling, stress response, and many proteases Citation53.
JAK/STAT pathway
The Drosophila JAK/STAT signaling pathway () comprises the same components as the mammalian pathway, although they differ in the number of JAKs (one in Drosophila, four in mammals) and STATs (one in Drosophila, seven in mammals) they possess.
Figure 1. Drosophila signaling pathways that regulate humoral responses. The Toll pathway regulates the expression of genes in response to the detection of Gram-positive type PPG or fungal β-1-3-glucans or activation by some fungal and bacterial proteases. The Imd pathway regulates the expression of genes in response to the detection of Gram-negative type PPG. The JAK/STAT pathway regulates the expression of genes in response to Upd3 signaling by plasmatocytes. Mammalian homologs of pathway components are shown in blue. Negative regulators of the pathways are not shown.
Activation and signal transduction
In response to septic injury, Drosophila plasmatocytes (macrophage like cells) secrete the cytokine, unpaired-3 (Upd3) Citation138 Citation139, which binds to the fat body cell transmembrane receptor, Domeless (homolog of mammalian class 1 cytokine receptor) Citation140 Citation141. Upd3 binding induces Domeless dimerization and the intracellular recruitment and phosphorylation of two Hopscotch (JAK) molecules, followed by two STAT92E (homolog of mammalian STAT5) molecules. Activated STAT92E molecules dimerize and translocate into the nucleus where they activate the expression of target genes Citation139.
Regulated genes
The JAK/STAT pathway regulates numerous physiological processes [reviewed in Citation142 Citation143 including immune responses; however, the contribution of this pathway to immune regulation has not been as well studied as the Toll and Imd pathways. In response to septic injury, the JAK/STAT pathway induces the expression of CG11501, which remains functionally uncharacterized Citation56 and, in combination with several other pathways Citation144, induces the expression of Turandots, which are stress-response proteins currently found only in Drosophila Citation145. The pathway is involved in the immune response in the Drosophila gut, where it helps maintain epithelial cell homeostasis, by regulating stem cell proliferation Citation51 Citation146. Activation of the JAK/STAT pathway in response to an intestinal Serratia marcescens infection negatively impacts Drosophila survival Citation51.
Downregulation
The JAK/STAT pathway is regulated on multiple levels like Imd and Toll. Drosophila protein inhibitor of activated STATs (dPIAS) inhibits STAT92E-dependent transcription by physically interacting with and blocking its DNA binding ability Citation147. Suppressor of cytokine signaling 36E (SOCS36E, mammalian soc5 homolog) expression is induced by JAK/STAT signaling and inhibits the pathway in a negative feedback loop Citation148 Citation149; a truncated form of STAT92E, which is transcribed from an alternate promoter, acts as a dominant negative regulator of JAK/STAT signaling Citation150; eye transformer, a receptor that is structurally related to Domeless, negatively regulates JAK/STAT signaling by interfering with STAT92E phosphorylation and activation Citation151.
Although the Drosophila antiviral immune response is not well characterized, a role for the Imd, Toll, and JAK-STAT pathways has been demonstrated Citation156 Citation157 Citation158 Citation159 .
Cellular immune response
The Drosophila cellular immune response is mediated by three sets of hemocytes: plasmatocytes, crystal cells, and lamellocytes, and while all three are found in larvae, only plasmatocytes are found in adults Citation160.
Plasmatocytes
These cells are similar to mammalian macrophages in that they express scavenger receptors Citation70 Citation161 Citation162 Citation163 Citation164 Citation165 , secrete cytokines Citation139 Citation166, and are responsible for the phagocytosis of microbes Citation45 Citation167 Citation168 Citation169 Citation170 and apoptotic cells Citation167 Citation171. Drosophila plasmatocytes also contribute to hemolymph coagulation by secreting the major clotting factor hemolectin Citation172, promote tolerance to intracellular bacterial infection (see below) Citation173, and utilize autophagy to control some intracellular pathogens (see below).
Phagocytosis
Phagocytosis involves several receptors many of which have mammalian homologs [see Citation174 for a recent in-depth review of Drosophila phagocytosis]. These include EGF repeat containing proteins Nimrod Citation162, Eater Citation161, and Draper Citation121, the scavenger receptors Croquemort (CD36 family), Peste (CD36 family) Citation165 Citation175, and Scavenger receptor CI (dSR-CI) Citation163, PGRP-LC Citation70, and Down syndrome cell adhesion molecule (Dscam, homolog of mammalian DSCAM) Citation164. Thus far, convincing in vivo evidence for the role of receptors in phagocytosis has only been obtained for Eater Citation161 and Nimrod Citation162. Phagocytosis may also be enhanced by the thiolester proteins (TEPs), which are members of the complement C3 (C3)/α2-macroglobulin (α2M) superfamily. The Drosophila genome encodes four Teps (I–IV) that are expressed at basal levels by fat body cells and larval hemocytes but strongly upregulated after a bacterial challenge (I, II, and IV in larvae and II and IV in adults); a fifth gene Tep V is believed to be a pseudogene Citation176. In vitro RNAi treatment of Drosophila S2 cells revealed that Teps II and III enhance the phagocytosis of Escherichia coli and S. aureus, respectively, likely by functioning as opsonins, while a sixth protein Tep VI (also known as macroglobulin complement related, Mcr) binds to and enhances the phagocytosis of Candida albicans Citation177. It has been suggested that Tep II may function as an α2M by virtue of its five splice isoforms that only differ in a domain that is similar to the bait region of α2Ms Citation176; however, this function has not been demonstrated. Although an in vivo role for the Teps (II and IV) in combating P. gingivalis infection has recently been demonstrated Citation178, Teps I–IV have been found to be dispensable for Drosophila survival after infection with a variety of other bacterial and fungal pathogens Citation179. RNAi studies have demonstrated that phagocytosis involves actin rearrangement Citation7 Citation177 Citation180, and phagocytosed microbes are taken up into a phagosome Citation57 and destroyed by mechanisms that are not yet clearly understood.
Autophagy
Autophagy is an evolutionarily conserved process by which cells envelope their cytoplasmic contents into double-membrane vesicles and, subsequently, deliver them to the lysosome for degradation. It is involved in the control of intracellular pathogen infections in mammals and was first reported as a Drosophila immune response by Yano et al., who observed that in vivo RNAi targeting an essential autophagy factor (Atg5) rendered Drosophila more susceptible to infection by the intracellular pathogen L. monocytogenes Citation181. They subsequently observed L. monocytogenes containing autophagosomes in Drosophila hemocytes using electron microscopy and identified the intracellular form of PGRP-LE (which detects DAP-type PPG) as the receptor responsible for the induction of autophagy. Autophagy is a part of the Drosophila antiviral immune response; however, the initiating receptor remains unidentified Citation182. Autophagy can also promote infection by pathogens such as the intracellular fungus Cryptococcus neoformans, which co-opts the autophagic system to maintain its lifestyle within Drosophila plasmatocytes Citation180.
Lamellocytes and crystal cells
Only present in Drosophila larvae, these cells are involved in encapsulation (lamellocytes) and melanization (crystal cells), immune responses that have no mammalian counterparts. Parasites that are too large to be phagocytosed, e.g. wasp eggs, are surrounded by lamellocytes and encapsulated, walling them off from the rest of the hemocoel (body cavity) Citation183 Citation184. The encapsulated particle is then destroyed by the local production of ROSs Citation185 Citation186, RNSs Citation187, and melanization, which is the synthesis and deposition of toxic melanin Citation188 Citation189. Melanization also occurs at the site of a cuticle breach and in Drosophila adults.
Coagulation
Coagulation is an immediate reaction that occurs at the site of a wound and prevents hemolymph loss, promotes wound healing, and acts as a secondary barrier to infection by immobilizing microbes Citation190 Citation191. Studies on hemolymph clots from Drosophila larvae revealed that the most abundant protein in the clot is hemolectin (homolog of mammalian clotting factor von Willebrand factor), which is secreted by plasmatocytes Citation172 Citation190 Citation192. Once the primary clot fibers have been formed, they are cross linked to each other and to other clot proteins like Fondue Citation193, by the enzyme transglutaminase (homolog of mammalian factor XIIIa) Citation194. Transglutaminase also accumulates on the surface of microbes and targets them to the clot, a role also observed for factor XIIIa in mammals Citation195. Drosophila clotting factor mutants have coagulation defects and are more sensitive to microbial infection Citation172 Citation190 Citation195.
Additional immune response features
MAPK pathways
Two MAPK pathways, the p38 MAPK pathway and the JNK pathway, contribute to the Drosophila immune response and are homologous to the mammalian pathways.
p38 MAPK pathway
p38 signaling in Drosophila has been shown to be involved in mediating stress responses Citation196 Citation197 Citation198 and immunity Citation40 Citation173 Citation199. Two MAPKs, p38a and p38b, are encoded by the Drosophila genome Citation196 Citation199, and they are activated in response to infection by the upstream MAPK-kinase (MKK) Licorne (homolog of mammalian MKK3) Citation199, which itself is activated by the MKK-kinase MEKK1 (homolog of mammalian MEKK4) Citation196 Citation198 Citation199 Citation200 . Once activated, p38 can then regulate the activity of transcription factors, such as ATF2 and d-Jun to control gene expression Citation199 Citation201. p38 null Drosophila are more susceptible than wildtype animals to killing by bacterial (40, and Igboin et al., unpublished) and fungal pathogens Citation40. Also overexpression of p38b conferred increased survival to Drosophila infected with intracellular bacteria (e.g. Salmonella typhimurium) without a corresponding decrease in bacterial load, and this was due to a p38b-dependent sequestering of the bacteria within plasmatocyte phagosomes Citation173 (see below for a discussion of tolerance). p38 signaling is negatively regulated by Alphabet, a phosphatase that antagonizes MEKK1 phosphorylation Citation202.
JNK pathway
In Drosophila, immune activation of JNK pathway signaling can occur via the binding of the cytokine Eiger (mammalian tumor necrosis factor homolog) to its receptor Wengen (mammalian TNF receptor homolog) Citation203 Citation204 Citation205 , also via the Imd pathway at the level of dTak1 Citation76, and mediates apoptosis, wound healing, morphogenesis, and immunity Citation206 Citation207 Citation208 . Downstream of Eiger/Wengen binding, one or more JNK kinase kinases (JNKKK) are activated by currently poorly understood mechanisms. The JNKKK dTak1, a shared component with the Imd pathway Citation76, activates two JNKKs, Hemipterous (mammalian MKK7 homolog) and Mkk4 (mammalian MKK4 homolog) Citation209, which in turn activate Basket (mammalian JNK homolog) Citation199 Citation209 Citation210. Activated JNK can then turn on the expression of transcription factors like AP1 Citation211 and other target genes. JNK signaling regulates a subset of immune response genes induced after septic injury Citation56 and also regulates AMP gene expression by the Imd pathway, downstream of Relish Citation212 Citation213. It has been reported that Drosophila JNK signaling in response to infection can either be beneficial or detrimental, as evidenced by the observation that Eiger null mutants are more susceptible than wildtype animals to killing by some pathogens, e.g. P. gingivalis and S. aureus Citation178 Citation214, but are more resistant than wildtype animals to killing by some intracellular pathogens, e.g. S. typhimurium Citation1 Citation214. It has also been reported that Eiger null mutants are no more susceptible to Gram-positive cocci, including S. aureus, infections than wildtype animals Citation215. Several negative regulators of JNK signaling have been identified, including Alphabet, a phosphatase that antagonizes dTAK1 phosphorylation Citation202, Pva (receptor tyrosine kinase) and Peroxiredoxin (redox sensing enzyme), which antagonize dTAK1 phosphorylation of JNK Citation216 Citation217, Puc, a phosphatase that antagonizes JNK activity Citation218, and Relish, whose activation results in proteosomal degradation of dTAK1 Citation219.
Tolerance
Tolerance is a process by which plants and animals endure a microbial infection and is one of the more recently described immune responses of Drosophila. Tolerance differs from resistance, in which the host reduces the fitness of the pathogen, thereby reducing the pathogen load. Tolerance in Drosophila has only been observed in response to bacterial infections, and the hallmarks of this response are increased fly survival without a corresponding decrease in the bacterial load Citation1 Citation173 Citation220 Citation221 or an increase in bacterial load without a corresponding decrease in fly survival Citation222. As examples of the first scenario, S. typhimurium-infected eiger null Drosophila survived 3 days longer than wt animals even though their bacterial loads were similar Citation1, and Drosophila overexpressing p38b survived S. typhimurium, L. monocytogenes, and Legionella pneumophilia infections longer than wildtype animals without a reduction in bacterial load Citation173. The mechanism of tolerance to S. typhimurium was shown to be via the sequestration of the bacteria within Drosophila hemocytes. As an example of the second scenario, wildtype and CG3066 (signaling protease involved in melanization) null Drosophila were killed at the same rate by Burkholderia cepacia, even though up to 25 times more bacteria were present in the mutant animals Citation222.
Reactive intermediate production
As mentioned previously, ROSs, e.g. superoxide anion, and RNSs, e.g. nitric oxide, are produced in Drosophila larvae during the melanotic encapsulation of particles that are too large to be phagocytosed Citation185 Citation186 Citation223. They also play a role in the immune response in the Drosophila gut where they are induced in response to natural infection (i.e. ingestion) Citation224 Citation225.
Drosophila dual oxidase (dDuox) is present on gut epithelial cells and contains a nicotinamide-adenine-dinucleotide-phosphate (NADPH)-oxidase domain and a peroxidase domain. The NADPH oxidase domain generates superoxide anion, which dismutates to hydrogen peroxide (H2O2), and the peroxidase domain uses the H2O2 to produce hypochlorous acid (HOCl) Citation224. Excessive ROS production is prevented by immune responsive catalase, which converts the H2O2 to water and oxygen Citation226.
NO participates in signaling in the gut of Drosophila larvae, which upregulate nitric oxide synthase expression upon natural infection with some Gram-negative bacteria. NO contributes to the induction of the AMP, diptericin, in the gut and hemocytes of naturally infected Drosophila larvae, and pharmacological inhibition of NOS has been shown to impair the ability of Drosophila larvae to survive septic and natural infections Citation225. The availability of a NO synthase null Drosophila mutant should shed additional light on the role of NO in the immune response.
Iron sequestration
The Drosophila genome encodes conserved iron-binding proteins transferrin Citation227 and ferritin Citation228, which can limit iron availability to invading microbes. Transferrin, ferritin, and iron transporters are upregulated in response to microbial challenges, which suggests a role for iron sequestration in controlling microbial infections in Drosophila Citation49 Citation50 Citation52 Citation227 Citation229.
RNA interference (RNAi)
RNAi involves the sequence-specific degradation of mRNA and is involved in the Drosophila antiviral immune response Citation19 Citation22 Citation230 Citation231. The role of RNAi in the Drosophila immune response was first reported by Li et al., who observed the accumulation of Flockhouse virus (FHV)-specific small interfering RNAs (siRNAs) in infected S2 cells Citation232. Galiana-Arnoux et al. Citation233, Zambon et al. Citation234, and van Rij et al Citation230 demonstrated an in vivo role for RNAi in combating viral infections. Drosophila also possess dsRNA uptake machinery Citation235 Citation236, which is important for the systemic RNAi response to extracellularly released (by controlled export or from lysed host cells) viral RNA Citation237.
In summary, three distinct pathways, Imd, Toll, and JAK/STAT, mediate the Drosophila humoral response, and the main humoral effectors are the AMPs, which are synthesized and secreted by the fat body. Two MAPK signaling pathways, JNK and p38, also regulate Drosophila immune responses, and all five signaling pathways are homologous to the pathways in mammals. The cellular immune response in Drosophila adults is mediated by plasmatocytes, macrophage-like cells that detect the presence of invading microbes via scavenger receptors, and phagocytose and subsequently destroy said microbes. Phagocytosis may be aided by the Teps, C3/α2M family proteins that function as opsonins. Additional Drosophila immune defense strategies include the production of reactive intermediates (oxygen and nitrogen), RNAi (to combat viral pathogens), the sequestration of iron, autophagy (to combat intracellular pathogens), and endurance of infections.
Drosophila infection models
Drosophila cultured cell lines (S2 and mbn-2), embryos, larvae, and adults (male and female) have been used to study host–pathogen interactions; however, adults and cultured cells are the most widely used. Drosophila embryos, larvae, and adults offer the advantage of whole animal models to examine pathogen–host interactions, and in all three models, the readout for pathogen virulence is fly mortality. A Drosophila embryo model was developed to study Photorhabdus asymbiotica infection, as it offered the added advantage of allowing the interaction between the bacterium and actively migrating Drosophila hemocytes to be followed in vivo, and in real time, using confocal microscopy Citation238. Drosophila larvae and adults have been used to identify and characterize pathogen and host factors that promote or inhibit infection by pathogens like P. gingivalis Citation63 Citation178, Pseudomonas fluorescens Citation3 Citation239, P. aeruginosa Citation40 Citation122 Citation240, Vibrio cholera Citation241 Citation242 Citation243 , S. aureus Citation244 Citation245, Klebsiella pneumonia Citation246, Streptococcus agalactiae Citation247, West Nile virus Citation19, Aspergillus fumigatus Citation13 Citation17, and Rhizopus oryzae Citation41. Drosophila adults have also been used to test the efficacy of antifungal Citation248 Citation249 Citation250 Citation251 and antibacterial drugs Citation252 Citation253 Citation254 and other antibacterial therapies Citation255 Citation256. It is important to take into account the immunological differences between the models when choosing one. For example, Drosophila larvae possess all three types of hemocytes but adults possess only plasmatocytes. Also, larval hemocytes do not actively migrate to a site of infection but rather arrive there by direct capture from the open circulatory system, in which they are passively pumped around Citation257.
Drosophila S2- and mbn2-cell lines are used as models for plasmatocytes because they are immunocompetent, phagocytic, and express many plasmatocyte-specific scavenger receptors. They are very useful for studying the phagocytosis, trafficking, proliferation, and cell-to-cell spread of pathogens Citation2 Citation121 Citation177 Citation180 Citation258 and for large scale screening by RNAi to identify host factors that interact with pathogens Citation20 Citation51 Citation259 Citation260 Citation261 Citation262 .
Routes of infection
Three routes of infection have been used to introduce pathogens into Drosophila: injection/septic injury, ingestion/natural infection, and rolling (in fungal spores). The injection method introduces the pathogen directly into the Drosophila hemocoel and involves either pricking the body cavity (usually the thorax in adults) with a needle dipped into a concentrated culture or the injection of a precise dose of the pathogen using a nanoinjector. Ingestion is also referred to as natural infection and involves introducing the animals into vials containing filter disks soaked with media containing the bacterium of interest or vials containing a pregrown lawn of fungal spores. Some ingestion models require that the animals be starved prior to their introduction into food containing vials, which drives subsequently ingested food to the crop. The rolling method of infection has only been used with fungal pathogens and involves rolling the flies over a lawn of spores growing on a plate. The rolling and injection methods of infection require anesthetization, which is usually done with carbon dioxide, and the injection and rolling methods require subsequent transfer of the animals into vials with food. After infection, the animals are incubated at 25–30°C (embryos are covered with oil), and their survival is monitored. It is important to note that the route of infection affects the interaction of the pathogen with the host and can give different results. For example, the alb1 gene is involved in A. fumigatus virulence when the fungus is ingested by or introduced onto the cuticle of Drosophila but not when the fungus is injected into the animals Citation17.
Transgenic expression (non-infection based)
In situations where the Drosophila model is not permissive for infection, it has been possible to introduce pathogen virulence factors into the flies by transgenically expressing the virulence factors in the animals. Pathogen genes can be expressed ubiquitously Citation26 or in specific fly tissues such as the fat body Citation21, wings Citation263, and eyes Citation264 by placing the transgene under the control of tissue-specific promoters. This has facilitated the identification of host cell targets of Plasmodium sporozite proteins Citation26, anthrax lethal factor and edema factor Citation263 Citation265, pertussis toxin Citation266, HIV-1 viral protein U Citation21, and Epstein-Barr virus immediate-early protein BZLF1 Citation264 ().
Drosophila–pathogen interactions
Drosophila as a model for analyzing bacterial virulence
Numerous studies have shown that bacterial virulence factors that are important for the successful infection of mammals are also involved in infecting Drosophila Citation1 Citation63 Citation170 Citation238 Citation240 Citation246 Citation247 Citation252 Citation254 Citation263 Citation267. Bacterial virulence factors that promote or inhibit infection in Drosophila, many of which are also involved in mammalian infection, are listed in . These include F. novicida oxidative stress transcriptional regulator, OxyR Citation268, L. monocytogenes actin polymerization protein, actA Citation4, and V. cholera toxin Citation252. P. aeruginosa has been extensively studied using the Drosophila model [see Citation64 for a recent review], and in the interest of brevity, this section of the review will address the use of the model to study Francisella tularensis virulence.
Table 1. Human bacterial pathogen genes that are involved in the infection of Drosophila
F. tularensis is a facultative intracellular pathogen that is the etiological agent of tularemia, a disease that affects humans and small vertebrates. Three subspecies, F. holartica, F. tularensis, and F. novicida, cause disease in humans and are vectored by blood-sucking insects. F. tularensis is able to infect and propagate in Drosophila mbn2 Citation170 and S2 cells Citation269 in vitro, killing the cells in the process. The bacterium also infects and kills adult flies in a dose-dependent manner Citation170 Citation269. Injection of GFP-expressing F. tularensis enabled the systemic spread of the bacteria and their localization to Drosophila hemocytes (about 55% of the bacteria were intracellular at the time of death) to be observed. Similar to infections in human cells, the bacteria are taken up into phagosomes in Drosophila S2 cells, which do not fuse with lysosomes, and from which the bacteria can escape within 30–60 min postinfection Citation269. The intracellular growth locus (igl) and macrophage growth locus (mgl) are involved in F. tularensis virulence in mice Citation270, and when several genes from these loci were mutated and the null mutants tested in Drosophila, they were found to be less virulent than the wildtype parental strain Citation170 Citation269.
Two hundred and forty-nine F. novicida transposon insertion mutants were individually evaluated in a Drosophila killing model, and 49 genes that were required for normal virulence in the flies were identified () Citation271. The majority of the identified genes were novel F. novicida virulence factors; 43 mutants had attenuated killing, and the mutated loci in these strains included most of the Francisella pathogenicity island (FPI) genes; six mutants were hypervirulent in the model and included mutants in pilus assembly proteins, an outer membrane protein and a kinase. Most of the attenuated strains also had low proliferation abilities in mammalian macrophages (murine J774 cells), and many of them also demonstrated reduced cytotoxicity; the hypervirulent mutants were significantly more cytotoxic to the murine macrophages than the wildtype strains. The strong correlation between F. novicida mutant phenotypes in Drosophila and in mouse macrophages suggested that the survival of F. novicida-infected flies is a good indicator of the bacterium's pathogenesis in mammals.
A F. novicida transposon mutant library was screened to identify genes that were essential for growth and survival in Drosophila, and 149 negatively selected genes (56 confirmed, ) were identified Citation268. Sixty of the genes had previously been identified as virulence factors in murine models. Seven of the negatively selected genes encoded proteins that are important for the bacterium's ability to resist oxidative damage, and the mutants showed increased sensitivity to reactive-oxygen producing agents in subsequent in vitro experiments. Screening the library in wt and Imd pathway-null Drosophila identified a subset of seven F. novicida genes that are required to resist Drosophila Imd-regulated defenses (genes that were negatively selected in wt flies but rescued in the mutant flies). Five of the seven mutants showed increased sensitivity to AMPs, modeled by polymyxin B, which are the main Imd-regulated immune effectors. The two mutants that showed no phenotype in the AMP killing assay were attenuated due to other Imd-dependent mechanisms, as similar levels of AMPs were induced in response to infection with these mutants as with the wt strain. Five genes, recB, pilA, pilB, pyrF, and manB, identified in this study as being essential for F. novicida growth and survival within the fly, were also identified by Ahlund et al. as being involved in F. tularensis virulence in Drosophila Citation271.
Another F. novicida transposon mutant library (3,050 alleles representing 1,448 genes) was first screened in cultured S2 cells to identify genes required for the intracellular proliferation of the bacterium, and then, a subset of the mutants was tested for their lethality in adult flies () Citation58. Three hundred and ninety-four genes that when mutated resulted in a significant reduction in intracellular growth of the bacterium were identified, and 80 of the 168 subsequently tested mutants exhibited reduced lethality and proliferation in the adult animals. The mutated loci encoded FPI proteins and proteins involved in metabolism, type IV pili biogenesis, transport, and DNA modification. One hundred and thirty-five genes were also required for F. novicida replication in human macrophages. Genes that were newly identified as playing a role in F. novicida pathogenesis may not be identifiable by currently available mammalian models, or could be specifically required for the insect phase of the bacterium's life cycle. Four genes, minD, iglC, iglD, and FTN_0109, identified in this study as being impaired in growth and/or lethality in Drosophila, were also identified by Ahlund et al. as being involved in F. tularensis virulence in Drosophila Citation271. Seven genes, ruvC, glpD, mdaB, ilvE, kdpC, pilQ, and FTN_1014, identified in this study, were also identified by Moule et al. as being essential for F. novicida growth and survival within the fly Citation268.
The studies described above and referenced in show that bacterial pathogens utilize many of the same virulence mechanisms to infect mammals and Drosophila, and that the Drosophila model is useful for identifying novel virulence factors.
Drosophila as a model for analyzing the host response to bacterial infection
The host response activated upon a bacterial infection can fight and resolve the infection or contribute to the pathology caused by the infection, and the vast array of Drosophila mutant and transgenic lines, and microarray, RNAi, and proteomics platforms has facilitated in-depth analyses of the host response to infection by a variety of pathogens. The results of studies in which several different Drosophila tools (immune response gene mutants, transgenic expression, whole genome microarray, and RNAi) were exploited to examine the host response to infection are discussed in this section of the review. Drosophila genes that are involved in the response to human bacterial pathogens are shown in .
Table 2.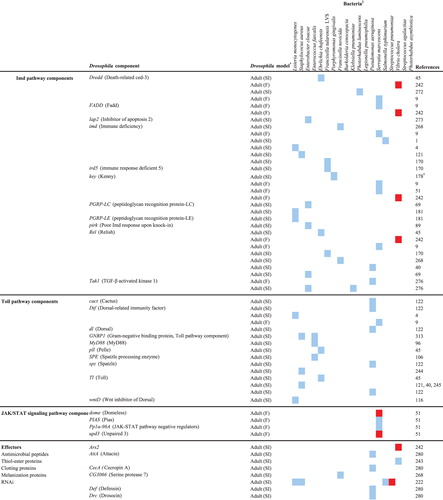

Loss-of-function immune response gene mutants
A large collection of loss-of-function Drosophila mutants have been generated by numerous investigators in the field and are readily available for use to identify Drosophila immune response components that play a role during infection. The use of knockout mutants of pathway components demonstrated that the well-characterized Toll- and Imd-signaling pathways are involved in the response to a variety of bacterial infections Citation4 Citation45 Citation49 Citation122 Citation170 Citation272 Citation273 Citation274 Citation275 . For example, Drosophila Dif (NFκB)-null mutants were more susceptible to killing than wt animals by L. monocytogenes, demonstrating a role for the Toll pathway in the defense against the bacterium Citation4. Imd pathway mutants were more susceptible than wildtype animals to killing by F. tularensis LVS Citation170, and both Imd and Toll mutants were more susceptible to killing by Erlichia chaffeensis Citation45. Once a Drosophila–pathogen model has been established, mutants that are deficient in specific host processes can be used to examine the interaction between those processes and the microbe. For example, Phg1 (nonaspanin)-null Drosophila were found to be highly susceptible to Gram-negative (P. aeruginosa, E. cloacae, and K. pneumoniae) bacterial infections, and it was demonstrated that this enhanced susceptibility was due to the inefficient phagocytic abilities of the fly hemocytes Citation276.
A large library of 1,231 transposon insertion mutants representing 8% of the Drosophila genome was screened to identify host genes required to survive a L. monocytogenes septic injury infection Citation277. Eighteen Drosophila mutants with increased susceptibility to killing by the bacterium were identified (), and the mutated loci included those involved in ubiquitination, RNA processing, and transcription activation. A comparison of the growth rates of L. monocytogenes in sensitive mutants and wildtype animals identified two classes of mutants: those in which bacterial growth was elevated relative to wildtype and those in which the bacterial levels remained unchanged relative to wildtype. This suggested that the first class of mutants is immunocompromised, i.e. deficient in genes required to resist the L.monocytogenes infection, while the second class of mutants has reduced endurance-characterized by lowered survival without a corresponding increase in bacterial proliferation-to the bacterial infection. Surprisingly, no overlap was observed between the Drosophila genes identified in this study and genes identified in two previous RNAi screens, likely because the use of S2 cells for the RNAi screens limited the observations to the processes involved in the interaction between L.monocytogenes and just one type of cell, whereas the whole animal model allowed for a much wider variety of processes to be examined.
An even larger library of 6,200 Drosophila transposon mutants was screened for altered susceptibility to a V. cholera natural infection, and 16 mutants were identified that contained disruptions in genes encoding proteins with homology to conserved domain proteins or mammalian proteins of know function Citation242 (). Seven of the Drosophila mutants demonstrated enhanced resistance, while nine demonstrated lowered resistance relative to the wildtype strain. Interestingly, the Imd pathway was found to contribute to pathology under normal circumstances (i.e. pathway mutants were more resistant to infection). It was determined that the mechanism of Imd involvement in causing pathology was due in part to an enhancement of cholera toxin activity, as imd pathway mutants were more resistant than wildtype animals to killing by a cholera toxin null V. cholera mutant. A significantly higher number of apoptotic intestinal epithelial cells was observed in infected imd pathway mutants than in wildtype animals, which suggested that under normal circumstances programmed cell death- a defense mechanism against intracellular pathogens in eukaryotes- is repressed by the Imd pathway in response to V. cholera infection.
Transgenic Drosophila
As discussed above (Routes of infection), transgenic Drosophila expressing pathogen virulence factors have been used to identify host targets. Host genes that are absent from Drosophila have also been transgenically expressed in the flies to study their interactions with pathogens. For example, the paraoxonase (PON) family of enzymes can degrade the P. aeruginosa quorum sensing (QS) signaling molecule N-3-oxododecanoyl homoserine lactone, and although they are conserved in mammals and Caenorhabditis elegans, the enzymes are absent from Drosophila. A QS deficient P. aeruginosa strain, synthetic acyl-homoserine lactones (AHLs), and Drosophila overexpressing human PON1 were used to examine the effect of the enzyme on the signaling molecule and on P. aeruginosa virulence in vivo Citation278. Drosophila that were infected with the P. aeruginosa QS-deficient mutant and ingested a control sucrose solution survived significantly better that flies that ingested synthetic AHLs, showing that QS is important for P. aeruginosa virulence in vivo. When human PON1 was overexpressed in the Drosophila, the animals were protected from the lethality of a wildtype P. aeruginosa infection. When AHLs were administered to P. aeruginosa QS mutant-infected Drosophila, PON1 overexpressing Drosophila but not wildtype animals were protected from killing, which showed that the basis of the observed protection was the inactivation of the AHLs by PON1. Therefore, the manipulation of PON expression could be explored as a treatment for some infectious diseases. In another study, it was observed that Drosophila overexpressing rhodanase, a bovine enzyme that detoxifies cyanide, survived longer than wildtype animals when infected with cyanogenic P. aeruginosa strains Citation279.
Whole genome microarray analysis
Whole-genome transcriptional profiling of whole animals has been used to assess the Drosophila global immune response to various infections Citation280 Citation281. The transcriptional profiles of Drosophila septically infected with virulent (PA14) and avirulent (CF5) strains of P. aeruginosa were compared, in order to identify defence-specific genes (expression altered after CF5 infection) and pathogenesis-specific genes (expression altered after PA14 infection) Citation280. Based on these criteria, 213 defence-specific genes (133 upregulated, 80 downregulated) and 28 pathogenesis-specific genes (16 upregulated, 12 downregulated) were identified. Interestingly, while infection with strain CF5 significantly upregulated AMP gene expression, infection with strain PA14 did not. Microarray analysis of AMP transcript levels following infection with a PA14 avirulent isogenic mutant revealed similar levels of expression as CF5, suggesting that the ability to reduce Drosophila AMP expression is a virulence trait of PA14 and likely other virulent P. aeruginosa strains.
Although transcriptional profiling provides a large amount of data, a subset of genes can be selected for more in depth examination. For example, a follow-up study examined the defense-specific genes that were non-immunity related, in a bid to understand the susceptibility to infection that occurs following trauma Citation282. It was observed that skeletal muscle genes (SMGs) were upregulated upon CF5 infection but not upon PA14 infection, suggesting that PA14 may suppress SMG induction in response to infection. It was determined that the JNK pathway is involved in the regulation of SMGs, as the transcriptional profile of a JNK pathway hypomorph infected with CF5 showed almost no induction of SMGs. Also, bacterial counts were higher in the thoraces of the JNK hypomorph than the wildtype strain but similar in the abdomens, suggesting that the JNK pathway protects against P. aeruginosa infection by promoting local tissue reconstruction after an infection. Interestingly, this role for the JNK pathway in the local defense against P. aeruginosa in tissue was conserved in a mouse open wound trauma model.
RNAi analysis
RNAi has been used extensively to examine the Drosophila host response to microbial infection in recent years due to the establishment of RNAi lines Citation47, dsRNA libraries representing the majority of Drosophila genes Citation283 Citation284, and well-developed, relatively easy methodology for use with bacterial Citation259, fungal Citation59, and viral pathogens Citation285.
The generation of over 22,000 transgenic Drosophila RNAi lines representing 12,800 genes allow for the knock down of host genes in specific tissues of the whole animal Citation47. Thirteen thousand and fifty-three RNAi lines representing 78% of the genome were screened to identify host factors that affected Drosophila survival after an ingestion infection with S. marcescens Citation51. Eight hundred and eighty-five lines with altered survival were identified, of which 790 were susceptible (RNAi of the genes increased susceptibility to killing by S. marcescens), and 95 were resistant (RNAi of the genes decreased susceptibility to killing by S. marcescens). Subsequent RNAi screening of the genes in the Drosophila gut epithelium identified 166 genes that affected Drosophila survival, most notably JAK/STAT signaling pathway components. Further study of the role of JAK/STAT signaling revealed that the pathway negatively regulates Drosophila survival in response to a S. marcescens infection, due to a disruption of intestinal cell homeostasis.
Drosophila S2 cells readily take up dsRNA added to their culture medium, and use it to silence genes Citation286, and the Drosophila RNAi Screening Center developed an RNAi library representing 13,900 Drosophila genes Citation284. This library has been used to perform genome-wide screens in S2 cells that aimed to identify host factors involved in infection by several pathogens Citation20 Citation165 Citation260 Citation261 Citation262 . A smaller targeted RNAi library representing 7,216 Drosophila genes that are conserved in metazoans Citation283 has been used to look specifically at host genes that may be relevant during human infection Citation177 Citation287 Citation288.
Drosophila as a model for examining polymicrobe–host interactions
Some human infectious diseases like periodontitis and cystic fibrosis are caused or exacerbated (in the case of CF) by polymicrobial communities; however, there is a dearth of in vivo models for studying the interactions among these microbes and between the microbes and the host. The polymicrobial community that colonizes the cyctic fibrosis airway results in persistent inflammation that ultimately destroys the lung, and P. aeruginosa is one of the primary CF pathogens. A Drosophila natural infection model was developed to examine the interaction between P. aeruginosa and other oropharyngeal (OF) species (including Streptococci, Neisseria, and Actinomyces) isolated from cystic fibrosis patients Citation289. A comparison of the survival of Drosophila infected with P. aeruginosa alone, each of the 40 OF species alone, and a combination of P. aeruginosa and the OF species identified three classes of microbes. The virulent class consisted of species that alone are able to kill Drosophila and enhanced the killing of Drosophila in combination with P. aeruginosa. The synergistic class represented species that alone are not pathogenic to Drosophila but, in combination with P. aeruginosa, significantly enhanced Drosophila killing. The avirulent group represented species that alone are not pathogenic to Drosophila and do not enhance the killing of Drosophila in combination with P. aeruginosa. A novel luminescence assay was used to monitor the expression of individual P. aeruginosa virulence genes in individual flies in real-time, during a coinfection with representatives of the synergistic class of OF species. P. aeruginosa virulence gene expression was altered in the presence of these OF species with half of the genes being upregulated, including several QS genes that are responsive to the interspecies signaling molecule autoinducer-2. This suggested that interspecies communication is important for modulating P. aeruginosa gene expression in a mixed microbial infection.
Using AMP (diptericin, cecropin, and drosomycin) expression as a readout for the immune response, three different host responses to a coinfection with P. aeruginosa and another OF species were observed: increased AMP expression as a result of the additive effect of both species, a suppression of AMP expression, and a synergistic activation of AMP expression to levels greater than would be achieved by the additive effect of both species. It was hypothesized that the hyperactivation of the immune response seen in the third instance could be detrimental to the animals. The results of this study demonstrate the complexity of polymicrobial- and polymicrobe–host-interactions and the power of the Drosophila model for deciphering these interactions.
Drosophila as a model for drug therapy and antibacterial screening
Drosophila provides the benefit of a whole animal context for drug and antibacterial testing, although the model is currently not as powerful as C. elegans for which high-throughput techniques have been developed Citation290. Drosophila can be used as an in vivo screen for antibiotics against bacteria like S. aureus Citation254 and V. cholera Citation252. Drosophila fed tetracycline or methicillin – which are used clinically to treat S. aureus infections – resisted a S. aureus infection that killed all flies fed a control sucrose solution Citation254. V. cholera infection in a Drosophila natural infection model mimics human cholera, and when the potassium ion channel blocker, clotrimazole, was co-administered with the bacterium the flies were resistant to the otherwise lethal infection Citation252.
Alternate antibacterial treatments such as phage therapy have been tested using Drosophila and have shown some promise. The lytic bacteriophage Caudovirales strains MPK1 and MPK6 were isolated based on their abilities to form plaques on lawns of P. aeruginosa Citation256. These phage also killed P. aeruginosa grown in broth culture, reducing the levels of the bacterium from 107 to 102 colony-forming units within 2 h. The efficacy of the phage against a P. aeruginosa infection in vivo was determined using a Drosophila septic injury model, and it was observed that when the phage were fed to the Drosophila, P. aeruginosa proliferation was inhibited and fly survival was enhanced. The phages were also protective against a P. aeruginosa infection in a mouse intraperitoneal model of infection.
D. melanogaster in oral microbiology
P. gingivalis
P. gingivalis is the first oral microbe that has been studied using the D. melanogaster model. More importantly, it is the first obligately anaerobic bacterium to be successfully studied using the model Citation63, despite the inhospitable oxygenated environment of the Drosophila hemolymph.
Using a septic injury route of infection, P. gingivalis was pathogenic to Drosophila, in a dose-dependent manner, with an intermediate level of pathogenicity between the non-pathogenic E. coil DH5-alpha and the highly pathogenic P. aeruginosa PA01 (). A comparison of clinically prevalent heteroduplex type strains of P. gingivalis revealed that all of the strains were virulent to some degree in Drosophila and that the highly disease-associated type strain, W83, was also the most pathogenic in the flies. P. gingivalis colony-forming unit levels did not increase in the Drosophila; however, the bacterium was able to persist in the flies up to 60 h postinfection. The relatively low temperature at which the infected flies were incubated (30°C), in addition to the oxygen rich environment of the hemolymph likely contribute to the inability of P. gingivalis to multiply in the host. However, P. gingivalis is aerotolerant and can survive exposure to air for up to 5 h without any loss in viability, which likely accounted for the bacterium's ability to persist in the Drosophila postinfection. P. gingivalis killing of Drosophila was not due to overt destruction of the fly tissues or a high bacterial burden as was observed with other pathogens, rather the observation that both live and heat-killed P. gingivalis effectively killed Drosophila suggested that the pathology may be due primarily to the host's own exaggerated immune response. Futher experiments are warranted to determine the exact cause of death of P. gingivalis-infected Drosophila, for example, in vivo RNAi could be used to dampen Drosophila immune responses and assess whether fly survival of P. gingivalis infection is enhanced as a result.
Figure 2. Survival curves of adult, female Drosophila infected with P. gingivalis and other bacterial species. E. coli DH5α (pink curve), P. aeruginosa strain PA01 (blue curve), P. gingivalis strain W83 (red curve), and vehicular control (yellow curve) Citation63.

Five P. gingivalis virulence gene mutants (arginine gingipains, lysine gingipain, major fimbriae, minor fimbriae, and capsule) that have been shown to be attenuated in rodent models of infection were also attenuated in the Drosophila model, demonstrating that P. gingivalis uses similar mechanisms of virulence in Drosophila and in mammals and that the bacterium's killing of Drosophila is multifactorial. It was hypothesized that as P. gingivalis spreads systemically (FITC-labeled bacteria were observed systemically using confocal microscopy), bacterial surface-associated components induce a systemic hyperactivation of the Drosophila immune response, which is not only bad for the bacterium but also harmful to the host.
To examine the Drosophila immune response to P. gingivalis infection Citation178, six immune response components were selected, and null mutants of these components were screened for altered susceptibility to P. gingvalis induced killing. Drosophila thiol-ester proteins, Tep II, Tep IV, and the JNK pathway ligand, Eiger (TNF homolog) were involved in the immune response against P. gingivalis infection, while the scavenger receptors Eater and Croquemort were dispensable for the response to P. gingivalis infection. Interestingly, the Imd pathway was initially found to be dispensable for the immune response against P. gingivalis, and because the strain that was used to infect the Drosophila (strain W83) is encapsulated, it was reasoned that the capsule may nullify any Imd pathway effects. This was supported by the observation that an unencapsulated P. gingivalis strain, strain 381, was significantly more pathogenic in Imd pathway-null flies than in wildtype flies. In a subsequent experiment, P. gingivalis strain W50UK, which is highly similar if not identical to W83, behaved like strain W83, while its isogenic capsule-null mutant, GPC, behaved like strain 381, when used to infect wildtype and Imd pathway-null flies, confirming that the bacterial capsule was responsible for the observed nullification of the Imd pathway.
To determine whether the P. gingivalis capsule prevented the activation of the Imd pathway by shielding the bacterium's peptidoglycan from detection by PGRP-LC, a dipt-LacZ reporter Drosophila strain was used to monitor activation of the pathway in response to W50UK and GPC infections. Beta galactosidase expression was detected in both cases, which demonstrated that the capsule of strain W50UK did not shield the bacterium's PPG from detection. To determine whether the P. gingivalis capsule could protect the bacterium from killing by Imd pathway regulated AMPs, the survival of strains W50UK and GPC after exposure to cecropin A and drosocin was compared. Strain GPC was significantly more susceptible to killing by the AMPs than W50UK, which showed that the P. gingivalis capsule mediates resistance to killing by Drosophila Imd-regulated AMPs. The results are also relevant to humans as the capsule also protected W50UK from killing by human beta-defensin-3. This was the first report to demonstrate that the P. gingivalis capsule is protective against host AMPs.
The future of the Drosophila model in the oral microbiology arena
Although the use of the Drosophila model for the study of oral microbiology is in its infancy, a wealth of information about oral bacteria–host interactions can be gleaned using this model. The major diseases of the oral cavity (periodontitis and caries) are polymicrobial in nature, and many putative periodontal Citation291 Citation292 Citation293 and caries Citation294 Citation295 pathogens have been identified and are yet to be characterized in terms of their interactions with the host. The introduction of the high-throughput Drosophila model will facilitate the screening of these potential pathogens, and the further characterization of species that are found to be virulent, as is being done with P. gingivalis. The Drosophila model is useful for studying P. gingivalis because this bacterium is aerotolerant, whereas a lot of oral bacteria will be killed by exposure to even a small amount of air and are, thus, unlikely to have virulent phenotypes in this model. Additional oral species have been screened for virulence using the Drosophila model, and a range of virulence has been observed (unpublished).
Further characterization of oral bacteria that are found to be pathogenic in the Drosophila model should involve screening for pathogen virulence factors, as well as host components that play a role in the response to infection. A large number of P. gingivalis mutants with in vitro phenotypes such as non-pigmentation, Citation296 Citation297 Citation298 Citation299 , enhanced biofilm forming ability, and loss of capsule synthesis Citation300 have been generated by transposon mutagenesis; however, likely due to the impracticality of using rodent models for large-scale screening, very few of these mutants have been tested for changes in virulence in vivo. The Drosophila model offers a high-throughput option to perform large-scale screening of these mutants to look for changes in virulence, which can then be further characterized. Also, the small size of Drosophila will make it relatively easy to profile its systemic response to infection, say with P. gingivalis, in a whole animal context, and could lead to the identification of novel host components that interact with the bacterium. Interesting candidates could be followed up on using null Drosophila mutants and/or in vivo RNAi.
The mixed microbial nature of periodontitis and caries warrants the study of polymicrobe interactions as well as polymicrobe–host interactions, and the work by Sibley et al Citation289 has laid the groundwork for the use of Drosophila to study these interactions in vivo. Studies are ongoing to examine the virulence of oral microbe mixtures comprised of P. gingivalis and other oral species, using Drosophila. The model could potentially be used to examine mixtures of microbes that are even more complex, possibly as complex as oral plaque.
Finally, Drosophila could potentially be used to test the efficacy of antimicrobials against oral bacteria in a whole-animal context; however, the model has not yet been developed for high-throughput drug testing.
It is important to keep in mind that Drosophila is not a natural host for oral microbes and that, due to anatomical and some host response – e.g. lack of an adaptive immune response – differences, the model is not directly comparable to humans. However, the large body of evidence obtained from studies in which the model was used to evaluate the host–microbe interplay has demonstrated that many mechanisms of microbial pathogenesis and the host response have been conserved between mammals and Drosophila. It is our belief that the addition of the Drosophila model to the repertoire of tools with which to study oral microbiology will facilitate infection research in this field considerably.
References
- Brandt SM, Dionne MS, Khush RS, Pham LN, Vigdal TJ, Schneider DS. Secreted bacterial effectors and host-produced eiger/TNF drive death in a Salmonella-infected fruit fly. PLoS Biol. 2004; 2: e418.
- Cheng LW, Portnoy DA. Drosophila S2 cells: an alternative infection model for Listeria monocytogenes. Cell Microbiol. 2003; 5: 875–85.
- de Lima Pimenta A, Di Martino P, Le Bouder E, Hulen C, Blight MA. In vitro identification of two adherence factors required for in vivo virulence of Pseudomonas fluorescens. Microbes Infect. 2003; 5: 1177–87.
- Mansfield BE, Dionne MS, Schneider DS, Freitag NE. Exploration of host-pathogen interactions using Listeria monocytogenes and Drosophila melanogaster. Cell Microbiol. 2003; 5: 901–11.
- Chugani SA, Whiteley M, Lee KM, D'Argenio D, Manoil C, Greenberg EP. QscR, a modulator of quorum-sensing signal synthesis and virulence in Pseudomonas aeruginosa. Proc Natl Acad Sci U S A. 2001; 98: 2752–7.
- Dionne MS, Ghori N, Schneider DS. Drosophila melanogaster is a genetically tractable model host for Mycobacterium marinum. Infect Immun. 2003; 71: 3540–50.
- Elwell C, Engel JN. Drosophila melanogaster S2 cells: a model system to study Chlamydia interaction with host cells. Cell Microbiol. 2005; 7: 725–39.
- Fauvarque MO, Bergeret E, Chabert J, Dacheux D, Satre M, Attree I. Role and activation of type III secretion system genes in Pseudomonas aeruginosa-induced Drosophila killing. Microb Pathog. 2002; 32: 287–95.
- Nehme NT, Liegeois S, Kele B, Giammarinaro P, Pradel E, Hoffmann JA, et al.. A model of bacterial intestinal infections in Drosophila melanogaster. PLoS Pathog. 2007; 3: e173.
- Alarco AM, Marcil A, Chen J, Suter B, Thomas D, Whiteway M. Immune-deficient Drosophila melanogaster: a model for the innate immune response to human fungal pathogens. J Immunol. 2004; 172: 5622–8.
- Apidianakis Y, Rahme LG, Heitman J, Ausubel FM, Calderwood SB, Mylonakis E. Challenge of Drosophila melanogaster with Cryptococcus neoformans and role of the innate immune response. Eukaryot Cell. 2004; 3: 413–9.
- Bhabhra R, Miley MD, Mylonakis E, Boettner D, Fortwendel J, Panepinto JC, et al.. Disruption of the Aspergillus fumigatus gene encoding nucleolar protein CgrA impairs thermotolerant growth and reduces virulence. Infect Immun. 2004; 72: 4731–40.
- Chamilos G, Bignell EM, Schrettl M, Lewis RE, Leventakos K, May GS, et al.. Exploring the concordance of Aspergillus fumigatus pathogenicity in mice and Toll-deficient flies. Med Mycol. 2010; 48: 506–10.
- Chamilos G, Lionakis MS, Lewis RE, Lopez-Ribot JL, Saville SP, Albert ND, et al.. Drosophila melanogaster as a facile model for large-scale studies of virulence mechanisms and antifungal drug efficacy in Candida species. J Infect Dis. 2006; 193: 1014–22.
- Glittenberg MT, Silas S, Maccallum DM, Gow NA, Ligoxygakis P. Wild-type Drosophila melanogaster as an alternative model system for investigating the pathogenicity of Candida albicans. Dis Model Mech. 2011 Jul; 4(4): 504–14.
- Lamaris GA, Chamilos G, Lewis RE, Kontoyiannis DP. Virulence studies of Scedosporium and Fusarium species in Drosophila melanogaster. J Infect Dis. 2007; 196: 1860–4.
- Lionakis MS, Kontoyiannis DP. The growing promise of Toll-deficient Drosophila melanogaster as a model for studying Aspergillus pathogenesis and treatment. Virulence. 2010; 1: 488–99.
- Battaglia PA, Zito S, Macchini A, Gigliani F. A Drosophila model of HIV-Tat-related pathogenicity. J Cell Sci. 2001; 114: 2787–94.
- Chotkowski HL, Ciota AT, Jia Y, Puig-Basagoiti F, Kramer LD, Shi PY, et al.. West Nile virus infection of Drosophila melanogaster induces a protective RNAi response. Virology. 2008; 377: 197–206.
- Hao L, Sakurai A, Watanabe T, Sorensen E, Nidom CA, Newton MA, et al.. Drosophila RNAi screen identifies host genes important for influenza virus replication. Nature. 2008; 454: 890–3.
- Leulier F, Marchal C, Miletich I, Limbourg-Bouchon B, Benarous R, Lemaitre B. Directed expression of the HIV-1 accessory protein Vpu in Drosophila fat-body cells inhibits Toll-dependent immune responses. EMBO Rep. 2003; 4: 976–81.
- Mukherjee S, Hanley KA. RNA interference modulates replication of dengue virus in Drosophila melanogaster cells. BMC Microbiol. 2010; 10: 127.
- Schneider D, Shahabuddin M. Malaria parasite development in a Drosophila model. Science. 2000; 288: 2376–9.
- Kotsyfakis M, Ehret-Sabatier L, Siden-Kiamos I, Mendoza J, Sinden RE, Louis C. Plasmodium berghei ookinetes bind to Anopheles gambiae and Drosophila melanogaster annexins. Mol Microbiol. 2005; 57: 171–9.
- Brandt SM, Jaramillo-Gutierrez G, Kumar S, Barillas-Mury C, Schneider DS. Use of a Drosophila model to identify genes regulating Plasmodium growth in the mosquito. Genetics. 2008; 180: 1671–8.
- Yan J, Yang X, Mortin MA. Shahabuddin M. Malaria sporozoite antigen-directed genome-wide response in transgenic Drosophila. Genesis. 2009; 47: 196–203.
- Adams MD, Celniker SE, Holt RA, Evans CA, Gocayne JD, et al.. The genome sequence of Drosophila melanogaster. Science. 2000; 287: 2185–95.
- Celniker SE, Wheeler DA, Kronmiller B, Carlson JW, Halpern A, Patel S, et al.. Finishing a whole-genome shotgun: release 3 of the Drosophila melanogaster euchromatic genome sequence. Genome Biol. 2002; 3: RESEARCH0079.
- Hoskins RA, Carlson JW, Kennedy C, Acevedo D, Evans-Holm M, Frise E, et al.. Sequence finishing and mapping of Drosophila melanogaster heterochromatin. Science. 2007; 316: 1625–8.
- Adams MD, Sekelsky JJ. From sequence to phenotype: reverse genetics in Drosophila melanogaster. Nat Rev Genet. 2002; 3: 189–98.
- Venken KJ, Bellen HJ. Emerging technologies for gene manipulation in Drosophila melanogaster. Nat Rev Genet. 2005; 6: 167–78.
- Bellen HJ, Levis RW, Liao G, He Y, Carlson JW, Tsang G, et al.. The BDGP gene disruption project: single transposon insertions associated with 40% of Drosophila genes. Genetics. 2004; 167: 761–81.
- Thibault ST, Singer MA, Miyazaki WY, Milash B, Dompe NA, Singh CM, et al.. A complementary transposon tool kit for Drosophila melanogaster using P and piggyBac. Nat Genet. 2004; 36: 283–7.
- Rubin GM, Spradling AC. Vectors for P element-mediated gene transfer in Drosophila. Nucleic Acids Res. 1983; 11: 6341–51.
- Golic MM, Rong YS, Petersen RB, Lindquist SL, Golic KG. FLP-mediated DNA mobilization to specific target sites in Drosophila chromosomes. Nucleic Acids Res. 1997; 25: 3665–71.
- Groth AC, Fish M, Nusse R, Calos MP. Construction of transgenic Drosophila by using the site-specific integrase from phage phiC31. Genetics. 2004; 166: 1775–82.
- Venken KJ, He Y, Hoskins RA, Bellen HJ. P[acman]: a BAC transgenic platform for targeted insertion of large DNA fragments in D. melanogaster. Science. 2006; 314: 1747–51.
- Fischer JA, Giniger E, Maniatis T, Ptashne M. GAL4 activates transcription in Drosophila. Nature. 1988; 332: 853–6.
- Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993; 118: 401–15.
- Chen J, Xie C, Tian L, Hong L, Wu X, Han J. Participation of the p38 pathway in Drosophila host defense against pathogenic bacteria and fungi. Proc Natl Acad Sci U S A. 2010; 107: 20774–9.
- Chamilos G, Lewis RE, Hu J, Xiao L, Zal T, Gilliet M, et al.. Drosophila melanogaster as a model host to dissect the immunopathogenesis of zygomycosis. Proc Natl Acad Sci U S A. 2008; 105: 9367–72.
- Levitin A, Marcil A, Tettweiler G, Laforest MJ, Oberholzer U, Alarco AM, et al.. Drosophila melanogaster Thor and response to Candida albicans infection. Eukaryot Cell. 2007; 6: 658–63.
- Mueller S, Gausson V, Vodovar N, Deddouche S, Troxler L, Perot J, et al.. RNAi-mediated immunity provides strong protection against the negative-strand RNA vesicular stomatitis virus in Drosophila. Proc Natl Acad Sci U S A. 2010; 107: 19390–5.
- Ferrandon D, Jung AC, Criqui M, Lemaitre B, Uttenweiler-Joseph S, Michaut L, et al.. A drosomycin-GFP reporter transgene reveals a local immune response in Drosophila that is not dependent on the Toll pathway. EMBO J. 1998; 17: 1217–27.
- Luce-Fedrow A, Von Ohlen T, Chapes SK. Ehrlichia chaffeensis infections in Drosophila melanogaster. Infect Immun. 2009; 77: 4815–26.
- Avet-Rochex A, Bergeret E, Attree I, Meister M, Fauvarque MO. Suppression of Drosophila cellular immunity by directed expression of the ExoS toxin GAP domain of Pseudomonas aeruginosa. Cell Microbiol. 2005; 7: 799–810.
- Dietzl G, Chen D, Schnorrer F, Su KC, Barinova Y, Fellner M, et al.. A genome-wide transgenic RNAi library for conditional gene inactivation in Drosophila. Nature. 2007; 448: 151–6.
- Karlsson C, Korayem AM, Scherfer C, Loseva O, Dushay MS, Theopold U. Proteomic analysis of the Drosophila larval hemolymph clot. J Biol Chem. 2004; 279: 52033–41.
- Levy F, Rabel D, Charlet M, Bulet P, Hoffmann JA, Ehret-Sabatier L. Peptidomic and proteomic analyses of the systemic immune response of Drosophila. Biochimie. 2004; 86: 607–16.
- Vierstraete E, Verleyen P, Sas F, Van den Bergh G, De Loof A, Arckens L, et al.. The instantly released Drosophila immune proteome is infection-specific. Biochem Biophys Res Commun. 2004; 317: 1052–60.
- Cronin SJ, Nehme NT, Limmer S, Liegeois S, Pospisilik JA, Schramek D, et al.. Genome-wide RNAi screen identifies genes involved in intestinal pathogenic bacterial infection. Science. 2009; 325: 340–3.
- De Gregorio E, Spellman PT, Rubin GM, Lemaitre B. Genome-wide analysis of the Drosophila immune response by using oligonucleotide microarrays. Proc Natl Acad Sci U S A. 2001; 98: 12590–5.
- De Gregorio E, Spellman PT, Tzou P, Rubin GM, Lemaitre B. The Toll and Imd pathways are the major regulators of the immune response in Drosophila. EMBO J. 2002; 21: 2568–79.
- Irving P, Troxler L, Heuer TS, Belvin M, Kopczynski C, Reichhart JM, et al.. A genome-wide analysis of immune responses in Drosophila. Proc Natl Acad Sci U S A. 2001; 98: 15119–24.
- Irving P, Ubeda JM, Doucet D, Troxler L, Lagueux M, Zachary D, et al.. New insights into Drosophila larval haemocyte functions through genome-wide analysis. Cell Microbiol. 2005; 7: 335–50.
- Boutros M, Agaisse H, Perrimon N. Sequential activation of signaling pathways during innate immune responses in Drosophila. Dev Cell. 2002; 3: 711–22.
- Stuart LM, Boulais J, Charriere GM, Hennessy EJ, Brunet S, Jutras I, et al.. A systems biology analysis of the Drosophila phagosome. Nature. 2007; 445: 95–101.
- Asare R, Akimana C, Jones S, Abu Kwaik Y. Molecular bases of proliferation of Francisella tularensis in arthropod vectors. Environ Microbiol. 2010; 12: 2587–612.
- Stroschein-Stevenson SL, Foley E, O'Farrell PH, Johnson AD. Phagocytosis of Candida albicans by RNAi-treated Drosophila S2 cells. Methods Mol Biol. 2009; 470: 347–58.
- Bernards A, Hariharan IK. Of flies and men—studying human disease in Drosophila. Curr Opin Genet Dev. 2001; 11: 274–8.
- Tweedie S, Ashburner M, Falls K, Leyland P, McQuilton P, Marygold S, et al.. FlyBase: enhancing Drosophila gene ontology annotations. Nucleic Acids Res. 2009; 37: D555–9.
- Krebs RA, Feder ME. Hsp70 and larval thermotolerance in Drosophila melanogaster: how much is enough and when is more too much?. J Insect Physiol. 1998; 44: 1091–101.
- Igboin CO, Moeschberger ML, Griffen AL, Leys EJ. Porphyromonas gingivalis virulence in a Drosophila melanogaster model. Infect Immun. 2011; 79: 439–48.
- Limmer S, Quintin J, Hetru C, Ferrandon D. Virulence on the fly: Drosophila melanogaster as a model genetic organism to decipher host-pathogen interactions. Curr Drug Targets. 2011; 12: 978–99.
- Ganesan S, Aggarwal K, Paquette N, Silverman N. NF-kappaB/Rel proteins and the humoral immune responses of Drosophila melanogaster. Curr Top Microbiol Immunol. 2011; 349: 25–60.
- Leulier F, Rodriguez A, Khush RS, Abrams JM, Lemaitre B. The Drosophila caspase Dredd is required to resist gram-negative bacterial infection. EMBO Rep. 2000; 1: 353–8.
- Rutschmann S, Jung AC, Zhou R, Silverman N, Hoffmann JA, Ferrandon D. Role of Drosophila IKK gamma in a toll-independent antibacterial immune response. Nat Immunol. 2000; 1: 342–7.
- Vidal S, Khush RS, Leulier F, Tzou P, Nakamura M, Lemaitre B. Mutations in the Drosophila dTAK1 gene reveal a conserved function for MAPKKKs in the control of rel/NF-kappaB-dependent innate immune responses. Genes Dev. 2001; 15: 1900–12.
- Gottar M, Gobert V, Michel T, Belvin M, Duyk G, Hoffmann JA, et al.. The Drosophila immune response against Gram-negative bacteria is mediated by a peptidoglycan recognition protein. Nature. 2002; 416: 640–4.
- Ramet M, Manfruelli P, Pearson A, Mathey-Prevot B, Ezekowitz RA. Functional genomic analysis of phagocytosis and identification of a Drosophila receptor for E. coli. Nature. 2002; 416: 644–8.
- Choe KM, Lee H, Anderson KV. Drosophila peptidoglycan recognition protein LC (PGRP-LC) acts as a signal-transducing innate immune receptor. Proc Natl Acad Sci U S A. 2005; 102: 1122–6.
- Lemaitre B, Kromer-Metzger E, Michaut L, Nicolas E, Meister M, Georgel P, et al.. A recessive mutation, immune deficiency (imd), defines two distinct control pathways in the Drosophila host defense. Proc Natl Acad Sci U S A. 1995; 92: 9465–9.
- Georgel P, Naitza S, Kappler C, Ferrandon D, Zachary D, Swimmer C, et al.. Drosophila immune deficiency (IMD) is a death domain protein that activates antibacterial defense and can promote apoptosis. Dev Cell. 2001; 1: 503–14.
- Hu S, Yang X. dFADD, a novel death domain-containing adapter protein for the Drosophila caspase DREDD. J Biol Chem. 2000; 275: 30761–4.
- Leulier F, Lhocine N, Lemaitre B, Meier P. The Drosophila inhibitor of apoptosis protein DIAP2 functions in innate immunity and is essential to resist gram-negative bacterial infection. Mol Cell Biol. 2006; 26: 7821–31.
- Silverman N, Zhou R, Erlich RL, Hunter M, Bernstein E, Schneider D, et al.. Immune activation of NF-kappaB and JNK requires Drosophila TAK1. J Biol Chem. 2003; 278: 48928–34.
- Hedengren M, Asling B, Dushay MS, Ando I, Ekengren S, Wihlborg M, et al.. Relish, a central factor in the control of humoral but not cellular immunity in Drosophila. Mol Cell. 1999; 4: 827–37.
- Silverman N, Zhou R, Stoven S, Pandey N, Hultmark D, Maniatis T. A Drosophila IkappaB kinase complex required for Relish cleavage and antibacterial immunity. Genes Dev. 2000; 14: 2461–71.
- Lu Y, Wu LP, Anderson KV. The antibacterial arm of the Drosophila innate immune response requires an IkappaB kinase. Genes Dev. 2001; 15: 104–10.
- Stoven S, Silverman N, Junell A, Hedengren-Olcott M, Erturk D, Engstrom Y, et al.. Caspase-mediated processing of the Drosophila NF-kappaB factor Relish. Proc Natl Acad Sci U S A. 2003; 100: 5991–6.
- Erturk-Hasdemir D, Broemer M, Leulier F, Lane WS, Paquette N, Hwang D, et al.. Two roles for the Drosophila IKK complex in the activation of Relish and the induction of antimicrobial peptide genes. Proc Natl Acad Sci U S A. 2009; 106: 9779–84.
- Bischoff V, Vignal C, Duvic B, Boneca IG, Hoffmann JA, Royet J. Downregulation of the Drosophila immune response by peptidoglycan-recognition proteins SC1 and SC2. PLoS Pathog. 2006; 2: e14.
- Mellroth P, Karlsson J, Steiner H. A scavenger function for a Drosophila peptidoglycan recognition protein. J Biol Chem. 2003; 278: 7059–64.
- Zaidman-Remy A, Herve M, Poidevin M, Pili-Floury S, Kim MS, Blanot D, et al.. The Drosophila amidase PGRP-LB modulates the immune response to bacterial infection. Immunity. 2006; 24: 463–73.
- Maillet F, Bischoff V, Vignal C, Hoffmann J, Royet J. The Drosophila peptidoglycan recognition protein PGRP-LF blocks PGRP-LC and IMD/JNK pathway activation. Cell Host Microbe. 2008; 3: 293–303.
- Guntermann S, Primrose DA, Foley E. Dnr1-dependent regulation of the Drosophila immune deficiency signaling pathway. Dev Comp Immunol. 2009; 33: 127–34.
- Kim M, Lee JH, Lee SY, Kim E, Chung J. Caspar, a suppressor of antibacterial immunity in Drosophila. Proc Natl Acad Sci U S A. 2006; 103: 16358–63.
- Aggarwal K, Rus F, Vriesema-Magnuson C, Erturk-Hasdemir D, Paquette N, Silverman N. Rudra interrupts receptor signaling complexes to negatively regulate the IMD pathway. PLoS Pathog. 2008; 4: e1000120.
- Kleino A, Myllymaki H, Kallio J, Vanha-aho LM, Oksanen K, Ulvila J, et al.. Pirk is a negative regulator of the Drosophila Imd pathway. J Immunol. 2008; 180: 5413–22.
- Lhocine N, Ribeiro PS, Buchon N, Wepf A, Wilson R, Tenev T, et al.. PIMS modulates immune tolerance by negatively regulating Drosophila innate immune signaling. Cell Host Microbe. 2008; 4: 147–58.
- Ragab A, Buechling T, Gesellchen V, Spirohn K, Boettcher AL, Boutros M. Drosophila Ras/MAPK signalling regulates innate immune responses in immune and intestinal stem cells. EMBO J. 2011; 30: 1123–36.
- Khush RS, Cornwell WD, Uram JN, Lemaitre B. A ubiquitin-proteasome pathway represses the Drosophila immune deficiency signaling cascade. Curr Biol. 2002; 12: 1728–37.
- Lemaitre B, Nicolas E, Michaut L, Reichhart JM, Hoffmann JA. The dorsoventral regulatory gene cassette Spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell. 1996; 86: 973–83.
- Rutschmann S, Jung AC, Hetru C, Reichhart JM, Hoffmann JA, Ferrandon D. The Rel protein DIF mediates the antifungal but not the antibacterial host defense in Drosophila. Immunity. 2000; 12: 569–80.
- Michel T, Reichhart JM, Hoffmann JA, Royet J. Drosophila Toll is activated by Gram-positive bacteria through a circulating peptidoglycan recognition protein. Nature. 2001; 414: 756–9.
- Tauszig-Delamasure S, Bilak H, Capovilla M, Hoffmann JA, Imler JL. Drosophila MyD88 is required for the response to fungal and Gram-positive bacterial infections. Nat Immunol. 2002; 3: 91–7.
- Rutschmann S, Kilinc A, Ferrandon D. Cutting edge: the toll pathway is required for resistance to gram-positive bacterial infections in Drosophila. J Immunol. 2002; 168: 1542–6.
- Gobert V, Gottar M, Matskevich AA, Rutschmann S, Royet J, Belvin M, et al.. Dual activation of the Drosophila toll pathway by two pattern recognition receptors. Science. 2003; 302: 2126–30.
- Wang L, Weber AN, Atilano ML, Filipe SR, Gay NJ, Ligoxygakis P. Sensing of Gram-positive bacteria in Drosophila: GNBP1 is needed to process and present peptidoglycan to PGRP-SA. EMBO J. 2006; 25: 5005–14.
- Bischoff V, Vignal C, Boneca IG, Michel T, Hoffmann JA, Royet J. Function of the Drosophila pattern-recognition receptor PGRP-SD in the detection of Gram-positive bacteria. Nat Immunol. 2004; 5: 1175–80.
- Ligoxygakis P, Pelte N, Hoffmann JA, Reichhart JM. Activation of Drosophila Toll during fungal infection by a blood serine protease. Science. 2002; 297: 114–6.
- Kim YS, Ryu JH, Han SJ, Choi KH, Nam KB, Jang IH, et al.. Gram-negative bacteria-binding protein, a pattern recognition receptor for lipopolysaccharide and beta-1,3-glucan that mediates the signaling for the induction of innate immune genes in Drosophila melanogaster cells. J Biol Chem. 2000; 275: 32721–7.
- Gottar M, Gobert V, Matskevich AA, Reichhart JM, Wang C, Butt TM, et al.. Dual detection of fungal infections in Drosophila via recognition of glucans and sensing of virulence factors. Cell. 2006; 127: 1425–37.
- El Chamy L, Leclerc V, Caldelari I, Reichhart JM. Sensing of danger signals and pathogen-associated molecular patterns defines binary signaling pathways upstream of Toll. Nat Immunol. 2008; 9: 1165–70.
- Kambris Z, Brun S, Jang IH, Nam HJ, Romeo Y, Takahashi K, et al.. Drosophila immunity: a large-scale in vivo RNAi screen identifies five serine proteases required for Toll activation. Curr Biol. 2006; 16: 808–13.
- Jang IH, Chosa N, Kim SH, Nam HJ, Lemaitre B, Ochiai M, et al.. A Spatzle-processing enzyme required for toll signaling activation in Drosophila innate immunity. Dev Cell. 2006; 10: 45–55.
- Sun H, Bristow BN, Qu G, Wasserman SA. A heterotrimeric death domain complex in Toll signaling. Proc Natl Acad Sci U S A. 2002; 99: 12871–6.
- Reichhart JM, Georgel P, Meister M, Lemaitre B, Kappler C, Hoffmann JA. Expression and nuclear translocation of the rel/NF-kappa B-related morphogen dorsal during the immune response of Drosophila. C R Acad Sci III. 1993; 316: 1218–24.
- Ip YT, Reach M, Engstrom Y, Kadalayil L, Cai H, Gonzalez-Crespo S, et al.. Dif, a dorsal-related gene that mediates an immune response in Drosophila. Cell. 1993; 75: 753–63.
- Lemaitre B, Meister M, Govind S, Georgel P, Steward R, Reichhart JM, et al.. Functional analysis and regulation of nuclear import of dorsal during the immune response in Drosophila. EMBO J. 1995; 14: 536–45.
- Wu LP, Anderson KV. Regulated nuclear import of Rel proteins in the Drosophila immune response. Nature. 1998; 392: 93–7.
- Manfruelli P, Reichhart JM, Steward R, Hoffmann JA, Lemaitre B. A mosaic analysis in Drosophila fat body cells of the control of antimicrobial peptide genes by the Rel proteins Dorsal and DIF. EMBO J. 1999; 18: 3380–91.
- Meng X, Khanuja BS, Ip YT. Toll receptor-mediated Drosophila immune response requires Dif, an NF-kappaB factor. Genes Dev. 1999; 13: 792–7.
- Kuttenkeuler D, Pelte N, Ragab A, Gesellchen V, Schneider L, Blass C, et al.. A large-scale RNAi screen identifies Deaf1 as a regulator of innate immune responses in Drosophila. J Innate Immun. 2010; 2: 181–94.
- Nicolas E, Reichhart JM, Hoffmann JA, Lemaitre B. In vivo regulation of the IkappaB homologue cactus during the immune response of Drosophila. J Biol Chem. 1998; 273: 10463–9.
- Gordon MD, Dionne MS, Schneider DS, Nusse R. WntD is a feedback inhibitor of Dorsal/NF-kappaB in Drosophila development and immunity. Nature. 2005; 437: 746–9.
- Chiu H, Ring BC, Sorrentino RP, Kalamarz M, Garza D, Govind S. dUbc9 negatively regulates the Toll-NF-kappa B pathways in larval hematopoiesis and drosomycin activation in Drosophila. Dev Biol. 2005; 288: 60–72.
- Fullaondo A, Garcia-Sanchez S, Sanz-Parra A, Recio E, Lee SY, Gubb D. Spn1 regulates the GNBP3-dependent Toll-signalling pathway in Drosophila melanogaster. Mol Cell Biol. 2011.
- Tanji T, Hu X, Weber AN, Ip YT. Toll and IMD pathways synergistically activate an innate immune response in Drosophila melanogaster. Mol Cell Biol. 2007; 27: 4578–88.
- Tanji T, Yun EY, Ip YT. Heterodimers of NF-kappaB transcription factors DIF and Relish regulate antimicrobial peptide genes in Drosophila. Proc Natl Acad Sci U S A. 2010; 107: 14715–20.
- Hashimoto Y, Tabuchi Y, Sakurai K, Kutsuna M, Kurokawa K, Awasaki T, et al.. Identification of lipoteichoic acid as a ligand for draper in the phagocytosis of Staphylococcus aureus by Drosophila hemocytes. J Immunol. 2009; 183: 7451–60.
- Lau GW, Goumnerov BC, Walendziewicz CL, Hewitson J, Xiao W, Mahajan-Miklos S, et al.. The Drosophila melanogaster toll pathway participates in resistance to infection by the gram-negative human pathogen Pseudomonas aeruginosa. Infect Immun. 2003; 71: 4059–66.
- Luce-Fedrow A, Von Ohlen T, Boyle D, Ganta RR, Chapes SK. Use of Drosophila S2 cells as a model for studying Ehrlichia chaffeensis infections. Appl Environ Microbiol. 2008; 74: 1886–91.
- Kylsten P, Samakovlis C, Hultmark D. The cecropin locus in Drosophila; a compact gene cluster involved in the response to infection. EMBO J. 1990; 9: 217–24.
- Dimarcq JL, Hoffmann D, Meister M, Bulet P, Lanot R, Reichhart JM, et al.. Characterization and transcriptional profiles of a Drosophila gene encoding an insect defensin. A study in insect immunity. Eur J Biochem. 1994; 221: 201–9.
- Hultmark D, Steiner H, Rasmuson T, Boman HG. Insect immunity. Purification and properties of three inducible bactericidal proteins from hemolymph of immunized pupae of Hyalophora cecropia. Eur J Biochem. 1980; 106: 7–16.
- Cociancich S, Ghazi A, Hetru C, Hoffmann JA, Letellier L. Insect defensin, an inducible antibacterial peptide, forms voltage-dependent channels in Micrococcus luteus. J Biol Chem. 1993; 268: 19239–45.
- Wicker C, Reichhart JM, Hoffmann D, Hultmark D, Samakovlis C, Hoffmann JA. Insect immunity. Characterization of a Drosophila cDNA encoding a novel member of the diptericin family of immune peptides. J Biol Chem. 1990; 265: 22493–8.
- Fehlbaum P, Bulet P, Michaut L, Lagueux M, Broekaert WF, Hetru C, et al.. Insect immunity. Septic injury of Drosophila induces the synthesis of a potent antifungal peptide with sequence homology to plant antifungal peptides. J Biol Chem. 1994; 269: 33159–63.
- Zhang ZT, Zhu SY. Drosomycin, an essential component of antifungal defence in Drosophila. Insect Mol Biol. 2009; 18: 549–56.
- Keppi E, Pugsley P, Lambert J, Wicker C, Dimarcq JL, Hoffmann J, et al.. Mode of action of diptericin A, a bactericidal peptide induced in the hemolymph of Phormia terranovae larvae. Arch Insect Biochem Physiol. 1989; 10: 229–39.
- Tian C, Gao B, Rodriguez Mdel C, Lanz-Mendoza H, Ma B, Zhu S. Gene expression, antiparasitic activity, and functional evolution of the drosomycin family. Mol Immunol. 2008; 45: 3909–16.
- Asling B, Dushay MS, Hultmark D. Identification of early genes in the Drosophila immune response by PCR-based differential display: the Attacin A gene and the evolution of attacin-like proteins. Insect Biochem Mol Biol. 1995; 25: 511–8.
- Bulet P, Dimarcq JL, Hetru C, Lagueux M, Charlet M, Hegy G, et al.. A novel inducible antibacterial peptide of Drosophila carries an O-glycosylated substitution. J Biol Chem. 1993; 268: 14893–7.
- Otvos L, Jr O I, Rogers ME, Consolvo PJ, Condie BA, Lovas S, et al.. Interaction between heat shock proteins and antimicrobial peptides. Biochemistry. 2000; 39: 14150–9.
- Levashina EA, Ohresser S, Bulet P, Reichhart JM, Hetru C, Hoffmann JA. Metchnikowin, a novel immune-inducible proline-rich peptide from Drosophila with antibacterial and antifungal properties. Eur J Biochem. 1995; 233: 694–700.
- Romeo Y, Lemaitre B. Drosophila immunity: methods for monitoring the activity of Toll and Imd signaling pathways. Methods Mol Biol. 2008; 415: 379–94.
- Harrison DA, McCoon PE, Binari R, Gilman M, Perrimon N. Drosophila unpaired encodes a secreted protein that activates the JAK signaling pathway. Genes Dev. 1998; 12: 3252–63.
- Agaisse H, Petersen UM, Boutros M, Mathey-Prevot B, Perrimon N. Signaling role of hemocytes in Drosophila JAK/STAT-dependent response to septic injury. Dev Cell. 2003; 5: 441–50.
- Brown S, Hu N, Hombria JC. Identification of the first invertebrate interleukin JAK/STAT receptor, the Drosophila gene domeless. Curr Biol. 2001; 11: 1700–5.
- Chen HW, Chen X, Oh SW, Marinissen MJ, Gutkind JS, Hou SX. mom identifies a receptor for the Drosophila JAK/STAT signal transduction pathway and encodes a protein distantly related to the mammalian cytokine receptor family. Genes Dev. 2002; 16: 388–98.
- Agaisse H, Perrimon N. The roles of JAK/STAT signaling in Drosophila immune responses. Immunol Rev. 2004; 198: 72–82.
- Arbouzova NI, Zeidler MP. JAK/STAT signalling in Drosophila: insights into conserved regulatory and cellular functions. Development. 2006; 133: 2605–16.
- Brun S, Vidal S, Spellman P, Takahashi K, Tricoire H, Lemaitre B. The MAPKKK Mekk1 regulates the expression of Turandot stress genes in response to septic injury in Drosophila. Genes Cells. 2006; 11: 397–407.
- Ekengren S, Hultmark D. A family of Turandot-related genes in the humoral stress response of Drosophila. Biochem Biophys Res Commun. 2001; 284: 998–1003.
- Liu W, Singh SR, Hou SX. JAK-STAT is restrained by Notch to control cell proliferation of the Drosophila intestinal stem cells. J Cell Biochem. 2010; 109: 992–9.
- Betz A, Lampen N, Martinek S, Young MW, Darnell JE Jr. A Drosophila PIAS homologue negatively regulates stat92E. Proc Natl Acad Sci U S A. 2001; 98: 9563–8.
- Karsten P, Hader S, Zeidler MP. Cloning and expression of Drosophila SOCS36E and its potential regulation by the JAK/STAT pathway. Mech Dev. 2002; 117: 343–6.
- Callus BA, Mathey-Prevot B. SOCS36E, a novel Drosophila SOCS protein, suppresses JAK/STAT and EGF-R signalling in the imaginal wing disc. Oncogene. 2002; 21: 4812–21.
- Henriksen MA, Betz A, Fuccillo MV, Darnell JE Jr. Negative regulation of STAT92E by an N-terminally truncated STAT protein derived from an alternative promoter site. Genes Dev. 2002; 16: 2379–89.
- Kallio J, Myllymaki H, Gronholm J, Armstrong M, Vanha-aho LM, Makinen L, et al.. Eye transformer is a negative regulator of Drosophila JAK/STAT signaling. FASEB J. 2010; 24: 4467–79.
- Makki R, Meister M, Pennetier D, Ubeda JM, Braun A, Daburon V, et al.. A short receptor downregulates JAK/STAT signalling to control the Drosophila cellular immune response. PLoS Biol. 2010; 8: e1000441.
- Baeg GH, Zhou R, Perrimon N. Genome-wide RNAi analysis of JAK/STAT signaling components in Drosophila. Genes Dev. 2005; 19: 1861–70.
- Muller P, Kuttenkeuler D, Gesellchen V, Zeidler MP, Boutros M. Identification of JAK/STAT signalling components by genome-wide RNA interference. Nature. 2005; 436: 871–5.
- Bach EA, Vincent S, Zeidler MP, Perrimon N. A sensitized genetic screen to identify novel regulators and components of the Drosophila janus kinase/signal transducer and activator of transcription pathway. Genetics. 2003; 165: 1149–66.
- Avadhanula V, Weasner BP, Hardy GG, Kumar JP, Hardy RW. A novel system for the launch of alphavirus RNA synthesis reveals a role for the Imd pathway in arthropod antiviral response. PLoS Pathog. 2009; 5: e1000582.
- Costa A, Jan E, Sarnow P, Schneider D. The Imd pathway is involved in antiviral immune responses in Drosophila. PLoS One. 2009; 4: e7436.
- Zambon RA, Nandakumar M, Vakharia VN, Wu LP. The Toll pathway is important for an antiviral response in Drosophila. Proc Natl Acad Sci U S A. 2005; 102: 7257–62.
- Dostert C, Jouanguy E, Irving P, Troxler L, Galiana-Arnoux D, Hetru C, et al.. The Jak-STAT signaling pathway is required but not sufficient for the antiviral response of Drosophila. Nat Immunol. 2005; 6: 946–53.
- Lanot R, Zachary D, Holder F, Meister M. Postembryonic hematopoiesis in Drosophila. Dev Biol. 2001; 230: 243–57.
- Kocks C, Cho JH, Nehme N, Ulvila J, Pearson AM, Meister M, et al.. Eater, a transmembrane protein mediating phagocytosis of bacterial pathogens in Drosophila. Cell. 2005; 123: 335–46.
- Kurucz E, Markus R, Zsamboki J, Folkl-Medzihradszky K, Darula Z, Vilmos P, et al.. Nimrod, a putative phagocytosis receptor with EGF repeats in Drosophila plasmatocytes. Curr Biol. 2007; 17: 649–54.
- Ramet M, Pearson A, Manfruelli P, Li X, Koziel H, Gobel V, et al.. Drosophila scavenger receptor CI is a pattern recognition receptor for bacteria. Immunity. 2001; 15: 1027–38.
- Watson FL, Puttmann-Holgado R, Thomas F, Lamar DL, Hughes M, Kondo M, et al.. Extensive diversity of Ig-superfamily proteins in the immune system of insects. Science. 2005; 309: 1874–8.
- Philips JA, Rubin EJ, Perrimon N. Drosophila RNAi screen reveals CD36 family member required for mycobacterial infection. Science. 2005; 309: 1251–3.
- Shia AK, Glittenberg M, Thompson G, Weber AN, Reichhart JM, Ligoxygakis P. Toll-dependent antimicrobial responses in Drosophila larval fat body require Spatzle secreted by haemocytes. J Cell Sci. 2009; 122: 4505–15.
- Charroux B, Royet J. Elimination of plasmatocytes by targeted apoptosis reveals their role in multiple aspects of the Drosophila immune response. Proc Natl Acad Sci U S A. 2009; 106: 9797–802.
- Nehme NT, Quintin J, Cho JH, Lee J, Lafarge MC, Kocks C, et al.. Relative roles of the cellular and humoral responses in the Drosophila host defense against three gram-positive bacterial infections. PLoS One. 2011; 6: e14743.
- Pham LN, Dionne MS, Shirasu-Hiza M, Schneider DS. A specific primed immune response in Drosophila is dependent on phagocytes. PLoS Pathog. 2007; 3: e26.
- Vonkavaara M, Telepnev MV, Ryden P, Sjostedt A, Stoven S. Drosophila melanogaster as a model for elucidating the pathogenicity of Francisella tularensis. Cell Microbiol. 2008; 10: 1327–38.
- Defaye A, Evans I, Crozatier M, Wood W, Lemaitre B, Leulier F. Genetic ablation of Drosophila phagocytes reveals their contribution to both development and resistance to bacterial infection. J Innate Immun. 2009; 1: 322–34.
- Lesch C, Goto A, Lindgren M, Bidla G, Dushay MS, Theopold U. A role for Hemolectin in coagulation and immunity in Drosophila melanogaster. Dev Comp Immunol. 2007; 31: 1255–63.
- Shinzawa N, Nelson B, Aonuma H, Okado K, Fukumoto S, Miura M, et al.. p38 MAPK-dependent phagocytic encapsulation confers infection tolerance in Drosophila. Cell Host Microbe. 2009; 6: 244–52.
- Ulvila J, Vanha-Aho LM, Ramet M. Drosophila phagocytosis—still many unknowns under the surface. APMIS. 2011; 119: 651–62.
- Franc NC, Dimarcq JL, Lagueux M, Hoffmann J, Ezekowitz RA. Croquemort, a novel Drosophila hemocyte/macrophage receptor that recognizes apoptotic cells. Immunity. 1996; 4: 431–43.
- Lagueux M, Perrodou E, Levashina EA, Capovilla M, Hoffmann JA. Constitutive expression of a complement-like protein in toll and JAK gain-of-function mutants of Drosophila. Proc Natl Acad Sci U S A. 2000; 97: 11427–32.
- Stroschein-Stevenson SL, Foley E, O'Farrell PH, Johnson AD. Identification of Drosophila gene products required for phagocytosis of Candida albicans. PLoS Biol. 2006; 4: e4.
- Igboin CO, Tordoff KP, Moeschberger ML, Griffen AL, Leys EJ. Porphyromonas gingivalis-host interactions in a Drosophila melanogaster model. Infect Immun. 2011; 79: 449–58.
- Bou Aoun R, Hetru C, Troxler L, Doucet D, Ferrandon D, Matt N. Analysis of thioester-containing proteins during the innate immune response of Drosophila melanogaster. J Innate Immun. 2011; 3: 52–64.
- Qin QM, Luo J, Lin X, Pei J, Li L, Ficht TA, et al.. Functional analysis of host factors that mediate the intracellular lifestyle of Cryptococcus neoformans. PLoS Pathog. 2011; 7: e1002078.
- Yano T, Mita S, Ohmori H, Oshima Y, Fujimoto Y, Ueda R, et al.. Autophagic control of listeria through intracellular innate immune recognition in Drosophila. Nat Immunol. 2008; 9: 908–16.
- Shelly S, Lukinova N, Bambina S, Berman A, Cherry S. Autophagy is an essential component of Drosophila immunity against vesicular stomatitis virus. Immunity. 2009; 30: 588–98.
- Rizki TM, Rizki RM. Lamellocyte differentiation in Drosophila larvae parasitized by Leptopilina. Dev Comp Immunol. 1992; 16: 103–10.
- Nappi AJ, Silvers M. Cell surface changes associated with cellular immune reactions in Drosophila. Science. 1984; 225: 1166–8.
- Nappi AJ, Vass E. Hydrogen peroxide production in immune-reactive Drosophila melanogaster. J Parasitol. 1998; 84: 1150–7.
- Nappi AJ, Vass E, Frey F, Carton Y. Superoxide anion generation in Drosophila during melanotic encapsulation of parasites. Eur J Cell Biol. 1995; 68: 450–6.
- Nappi AJ, Vass E, Frey F, Carton Y. Nitric oxide involvement in Drosophila immunity. Nitric Oxide. 2000; 4: 423–30.
- Ashida M. The prophenoloxidase cascade in insect immunity. Res Immunol. 1990; 141: 908–10.
- Nappi AJ, Vass E. Melanogenesis and the generation of cytotoxic molecules during insect cellular immune reactions. Pigment Cell Res. 1993; 6: 117–26.
- Scherfer C, Karlsson C, Loseva O, Bidla G, Goto A, Havemann J, et al.. Isolation and characterization of hemolymph clotting factors in Drosophila melanogaster by a pullout method. Curr Biol. 2004; 14: 625–9.
- Bidla G, Lindgren M, Theopold U, Dushay MS. Hemolymph coagulation and phenoloxidase in Drosophila larvae. Dev Comp Immunol. 2005; 29: 669–79.
- Goto A, Kumagai T, Kumagai C, Hirose J, Narita H, Mori H, et al.. A Drosophila haemocyte-specific protein, hemolectin, similar to human von Willebrand factor. Biochem J. 2001; 359: 99–108.
- Scherfer C, Qazi MR, Takahashi K, Ueda R, Dushay MS, Theopold U, et al.. The Toll immune-regulated Drosophila protein Fondue is involved in hemolymph clotting and puparium formation. Dev Biol. 2006; 295: 156–63.
- Lindgren M, Riazi R, Lesch C, Wilhelmsson C, Theopold U, Dushay MS. Fondue and transglutaminase in the Drosophila larval clot. J Insect Physiol. 2008; 54: 586–92.
- Wang Z, Wilhelmsson C, Hyrsl P, Loof TG, Dobes P, Klupp M, et al.. Pathogen entrapment by transglutaminase—a conserved early innate immune mechanism. PLoS Pathog. 2010; 6: e1000763.
- Han SJ, Choi KY, Brey PT, Lee WJ. Molecular cloning and characterization of a Drosophila p38 mitogen-activated protein kinase. J Biol Chem. 1998; 273: 369–74.
- Craig CR, Fink JL, Yagi Y, Ip YT, Cagan RL. A Drosophila p38 orthologue is required for environmental stress responses. EMBO Rep. 2004; 5: 1058–63.
- Inoue H, Tateno M, Fujimura-Kamada K, Takaesu G, Adachi-Yamada T, Ninomiya-Tsuji J, et al.. A Drosophila MAPKKK, D-MEKK1, mediates stress responses through activation of p38 MAPK. EMBO J. 2001; 20: 5421–30.
- Han ZS, Enslen H, Hu X, Meng X, Wu IH, Barrett T, et al.. A conserved p38 mitogen-activated protein kinase pathway regulates Drosophila immunity gene expression. Mol Cell Biol. 1998; 18: 3527–39.
- Zhuang ZH, Zhou Y, Yu MC, Silverman N, Ge BX. Regulation of Drosophila p38 activation by specific MAP2 kinase and MAP3 kinase in response to different stimuli. Cell Signal. 2006; 18: 441–8.
- Sano Y, Akimaru H, Okamura T, Nagao T, Okada M, Ishii S. Drosophila activating transcription factor-2 is involved in stress response via activation by p38, but not c-Jun NH(2)-terminal kinase. Mol Biol Cell. 2005; 16: 2934–46.
- Baril C, Sahmi M, Ashton-Beaucage D, Stronach B, Therrien M. The PP2C Alphabet is a negative regulator of stress-activated protein kinase signaling in Drosophila. Genetics. 2009; 181: 567–79.
- Igaki T, Kanda H, Yamamoto-Goto Y, Kanuka H, Kuranaga E, Aigaki T, et al.. Eiger, a TNF superfamily ligand that triggers the Drosophila JNK pathway. EMBO J. 2002; 21: 3009–18.
- Kanda H, Igaki T, Kanuka H, Yagi T, Miura M. Wengen, a member of the Drosophila tumor necrosis factor receptor superfamily, is required for Eiger signaling. J Biol Chem. 2002; 277: 28372–5.
- Kauppila S, Maaty WS, Chen P, Tomar RS, Eby MT, Chapo J, et al.. Eiger and its receptor, Wengen, comprise a TNF-like system in Drosophila. Oncogene. 2003; 22: 4860–7.
- Moreno E, Yan M, Basler K. Evolution of TNF signaling mechanisms: JNK-dependent apoptosis triggered by Eiger, the Drosophila homolog of the TNF superfamily. Curr Biol. 2002; 12: 1263–8.
- Sluss HK, Han Z, Barrett T, Goberdhan DC, Wilson C, Davis RJ, et al.. A JNK signal transduction pathway that mediates morphogenesis and an immune response in Drosophila. Genes Dev. 1996; 10: 2745–58.
- Bidla G, Dushay MS, Theopold U. Crystal cell rupture after injury in Drosophila requires the JNK pathway, small GTPases and the TNF homolog Eiger. J Cell Sci. 2007; 120: 1209–15.
- Chen W, White MA, Cobb MH. Stimulus-specific requirements for MAP3 kinases in activating the JNK pathway. J Biol Chem. 2002; 277: 49105–10.
- Baril C, Therrien M. Alphabet, a Ser/Thr phosphatase of the protein phosphatase 2C family, negatively regulates RAS/MAPK signaling in Drosophila. Dev Biol. 2006; 294: 232–45.
- Kockel L, Homsy JG, Bohmann D. Drosophila AP-1: lessons from an invertebrate. Oncogene. 2001; 20: 2347–64.
- Delaney JR, Stoven S, Uvell H, Anderson KV, Engstrom Y, Mlodzik M. Cooperative control of Drosophila immune responses by the JNK and NF-kappaB signaling pathways. EMBO J. 2006; 25: 3068–77.
- Kallio J, Leinonen A, Ulvila J, Valanne S, Ezekowitz RA, Ramet M. Functional analysis of immune response genes in Drosophila identifies JNK pathway as a regulator of antimicrobial peptide gene expression in S2 cells. Microbes Infect. 2005; 7: 811–9.
- Schneider DS, Ayres JS, Brandt SM, Costa A, Dionne MS, Gordon MD, et al.. Drosophila eiger mutants are sensitive to extracellular pathogens. PLoS Pathog. 2007; 3: e41.
- Geuking P, Narasimamurthy R, Lemaitre B, Basler K, Leulier F. A non-redundant role for Drosophila Mkk4 and hemipterous/Mkk7 in TAK1-mediated activation of JNK. PLoS One. 2009; 4: e7709.
- Bond D, Foley E. A quantitative RNAi screen for JNK modifiers identifies Pvr as a novel regulator of Drosophila immune signaling. PLoS Pathog. 2009; 5: e1000655.
- Radyuk SN, Michalak K, Klichko VI, Benes J, Orr WC. Peroxiredoxin 5 modulates immune response in Drosophila. Biochim Biophys Acta. 2010; 1800: 1153–63.
- Martin-Blanco E, Gampel A, Ring J, Virdee K, Kirov N, Tolkovsky AM, et al.. puckered encodes a phosphatase that mediates a feedback loop regulating JNK activity during dorsal closure in Drosophila. Genes Dev. 1998; 12: 557–70.
- Park JM, Brady H, Ruocco MG, Sun H, Williams D, Lee SJ, et al.. Targeting of TAK1 by the NF-kappa B protein Relish regulates the JNK-mediated immune response in Drosophila. Genes Dev. 2004; 18: 584–94.
- Corby-Harris V, Habel KE, Ali FG, Promislow DE. Alternative measures of response to Pseudomonas aeruginosa infection in Drosophila melanogaster. J Evol Biol. 2007; 20: 526–33.
- Dionne MS, Pham LN, Shirasu-Hiza M, Schneider DS. Akt and FOXO dysregulation contribute to infection-induced wasting in Drosophila. Curr Biol. 2006; 16: 1977–85.
- Ayres JS, Schneider DS. A signaling protease required for melanization in Drosophila affects resistance and tolerance of infections. PLoS Biol. 2008; 6: 2764–73.
- Tirouvanziam R, Davidson CJ, Lipsick JS, Herzenberg LA. Fluorescence-activated cell sorting (FACS) of Drosophila hemocytes reveals important functional similarities to mammalian leukocytes. Proc Natl Acad Sci U S A. 2004; 101: 2912–7.
- Ha EM, Oh CT, Bae YS, Lee WJ. A direct role for dual oxidase in Drosophila gut immunity. Science. 2005; 310: 847–50.
- Foley E, O'Farrell PH. Nitric oxide contributes to induction of innate immune responses to gram-negative bacteria in Drosophila. Genes Dev. 2003; 17: 115–25.
- Ha EM, Oh CT, Ryu JH, Bae YS, Kang SW, Jang IH, et al.. An antioxidant system required for host protection against gut infection in Drosophila. Dev Cell. 2005; 8: 125–32.
- Yoshiga T, Georgieva T, Dunkov BC, Harizanova N, Ralchev K, Law JH. Drosophila melanogaster transferrin. Cloning, deduced protein sequence, expression during the life cycle, gene localization and up-regulation on bacterial infection. Eur J Biochem. 1999; 260: 414–20.
- Georgieva T, Dunkov BC, Dimov S, Ralchev K, Law JH. Drosophila melanogaster ferritin: cDNA encoding a light chain homologue, temporal and tissue specific expression of both subunit types. Insect Biochem Mol Biol. 2002; 32: 295–302.
- Vierstraete E, Verleyen P, Baggerman G, D'Hertog W, Van den Bergh G, Arckens L, et al.. A proteomic approach for the analysis of instantly released wound and immune proteins in Drosophila melanogaster hemolymph. Proc Natl Acad Sci U S A. 2004; 101: 470–5.
- van Rij RP, Saleh MC, Berry B, Foo C, Houk A, Antoniewski C, et al.. The RNA silencing endonuclease Argonaute 2 mediates specific antiviral immunity in Drosophila melanogaster. Genes Dev. 2006; 20: 2985–95.
- Wang XH, Aliyari R, Li WX, Li HW, Kim K, Carthew R, et al.. RNA interference directs innate immunity against viruses in adult Drosophila. Science. 2006; 312: 452–4.
- Li H, Li WX, Ding SW. Induction and suppression of RNA silencing by an animal virus. Science. 2002; 296: 1319–21.
- Galiana-Arnoux D, Dostert C, Schneemann A, Hoffmann JA, Imler JL. Essential function in vivo for Dicer-2 in host defense against RNA viruses in Drosophila. Nat Immunol. 2006; 7: 590–7.
- Zambon RA, Vakharia VN, Wu LP. RNAi is an antiviral immune response against a dsRNA virus in Drosophila melanogaster. Cell Microbiol. 2006; 8: 880–9.
- Saleh MC, van Rij RP, Hekele A, Gillis A, Foley E, O'Farrell PH, et al.. The endocytic pathway mediates cell entry of dsRNA to induce RNAi silencing. Nat Cell Biol. 2006; 8: 793–802.
- Ulvila J, Parikka M, Kleino A, Sormunen R, Ezekowitz RA, Kocks C, et al.. Double-stranded RNA is internalized by scavenger receptor-mediated endocytosis in Drosophila S2 cells. J Biol Chem. 2006; 281: 14370–5.
- Saleh MC, Tassetto M, van Rij RP, Goic B, Gausson V, Berry B, et al.. Antiviral immunity in Drosophila requires systemic RNA interference spread. Nature. 2009; 458: 346–50.
- Vlisidou I, Dowling AJ, Evans IR, Waterfield N, ffrench-Constant RH, Wood W. Drosophila embryos as model systems for monitoring bacterial infection in real time. PLoS Pathog. 2009; 5: e1000518.
- Olcott MH, Henkels MD, Rosen KL, Walker FL, Sneh B, Loper JE, et al.. Lethality and developmental delay in Drosophila melanogaster larvae after ingestion of selected Pseudomonas fluorescens strains. PLoS One. 2010; 5: e12504.
- Kim SH, Park SY, Heo YJ, Cho YH. Drosophila melanogaster-based screening for multihost virulence factors of Pseudomonas aeruginosa PA14 and identification of a virulence-attenuating factor, HudA. Infect Immun. 2008; 76: 4152–62.
- An D, Apidianakis Y, Boechat AL, Baldini RL, Goumnerov BC, Rahme LG. The pathogenic properties of a novel and conserved gene product, KerV, in proteobacteria. PLoS One. 2009; 4: e7167.
- Berkey CD, Blow N, Watnick PI. Genetic analysis of Drosophila melanogaster susceptibility to intestinal Vibrio cholerae infection. Cell Microbiol. 2009; 11: 461–74.
- Park SY, Heo YJ, Kim KS, Cho YH. Drosophila melanogaster is susceptible to Vibrio cholerae infection. Mol Cells. 2005; 20: 409–15.
- Jin LH, Shim J, Yoon JS, Kim B, Kim J, Kim-Ha J, et al.. Identification and functional analysis of antifungal immune response genes in Drosophila. PLoS Pathog. 2008; 4: e1000168.
- Tabuchi Y, Shiratsuchi A, Kurokawa K, Gong JH, Sekimizu K, Lee BL, et al.. Inhibitory role for D-alanylation of wall teichoic acid in activation of insect Toll pathway by peptidoglycan of Staphylococcus aureus. J Immunol. 2010; 185: 2424–31.
- Benghezal M, Fauvarque MO, Tournebize R, Froquet R, Marchetti A, Bergeret E, et al.. Specific host genes required for the killing of Klebsiella bacteria by phagocytes. Cell Microbiol. 2006; 8: 139–48.
- Baron MJ, Wong SL, Nybakken K, Carey VJ, Madoff LC. Host glycosaminoglycan confers susceptibility to bacterial infection in Drosophila melanogaster. Infect Immun. 2009; 77: 860–6.
- Chamilos G, Lewis RE, Kontoyiannis DP. Lovastatin has significant activity against zygomycetes and interacts synergistically with voriconazole. Antimicrob Agents Chemother. 2006; 50: 96–103.
- Lamaris GA, Ben-Ami R, Lewis RE, Chamilos G, Samonis G, Kontoyiannis DP. Increased virulence of Zygomycetes organisms following exposure to voriconazole: a study involving fly and murine models of zygomycosis. J Infect Dis. 2009; 199: 1399–406.
- Lamaris GA, Ben-Ami R, Lewis RE, Kontoyiannis DP. Does pre-exposure of Aspergillus fumigatus to voriconazole or posaconazole in vitro affect its virulence and the in vivo activity of subsequent posaconazole or voriconazole, respectively? A study in a fly model of aspergillosis. J Antimicrob Chemother. 2008; 62: 539–42.
- Lionakis MS, Lewis RE, May GS, Wiederhold NP, Albert ND, Halder G, et al.. Toll-deficient Drosophila flies as a fast, high-throughput model for the study of antifungal drug efficacy against invasive aspergillosis and Aspergillus virulence. J Infect Dis. 2005; 191: 1188–95.
- Blow NS, Salomon RN, Garrity K, Reveillaud I, Kopin A, Jackson FR, et al.. Vibrio cholerae infection of Drosophila melanogaster mimics the human disease cholera. PLoS Pathog. 2005; 1: e8.
- Broderick KE, Feala J, McCulloch A, Paternostro G, Sharma VS, Pilz RB, et al.. The nitric oxide scavenger cobinamide profoundly improves survival in a Drosophila melanogaster model of bacterial sepsis. FASEB J. 2006; 20: 1865–73.
- Needham AJ, Kibart M, Crossley H, Ingham PW, Foster SJ. Drosophila melanogaster as a model host for Staphylococcus aureus infection. Microbiology. 2004; 150: 2347–55.
- Estin ML, Stoltz DA, Zabner J. Paraoxonase 1, quorum sensing, and P. aeruginosa infection: a novel model. Adv Exp Med Biol. 2010; 660: 183–93.
- Heo YJ, Lee YR, Jung HH, Lee J, Ko G, Cho YH. Antibacterial efficacy of phages against Pseudomonas aeruginosa infections in mice and Drosophila melanogaster. Antimicrob Agents Chemother. 2009; 53: 2469–74.
- Babcock DT, Brock AR, Fish GS, Wang Y, Perrin L, Krasnow MA, et al.. Circulating blood cells function as a surveillance system for damaged tissue in Drosophila larvae. Proc Natl Acad Sci U S A. 2008; 105: 10017–22.
- Serio AW, Jeng RL, Haglund CM, Reed SC, Welch MD. Defining a core set of actin cytoskeletal proteins critical for actin-based motility of Rickettsia. Cell Host Microbe. 2010; 7: 388–98.
- Agaisse H. Investigating the involvement of host factors involved in intracellular pathogen infection by RNAi in Drosophila cells. Methods Mol Biol. 2008; 415: 395–402.
- Akimana C, Al-Khodor S, Abu Kwaik Y. Host factors required for modulation of phagosome biogenesis and proliferation of Francisella tularensis within the cytosol. PLoS One. 2010; 5: e11025.
- Agaisse H, Burrack LS, Philips JA, Rubin EJ, Perrimon N, Higgins DE. Genome-wide RNAi screen for host factors required for intracellular bacterial infection. Science. 2005; 309: 1248–51.
- Sessions OM, Barrows NJ, Souza-Neto JA, Robinson TJ, Hershey CL, Rodgers MA, et al.. Discovery of insect and human dengue virus host factors. Nature. 2009; 458: 1047–50.
- Guichard A, Park JM, Cruz-Moreno B, Karin M, Bier E. Anthrax lethal factor and edema factor act on conserved targets in Drosophila. Proc Natl Acad Sci U S A. 2006; 103: 3244–9.
- Adamson AL, Wright N, LaJeunesse DR. Modeling early Epstein-Barr virus infection in Drosophila melanogaster: the BZLF1 protein. Genetics. 2005; 171: 1125–35.
- Guichard A, McGillivray SM, Cruz-Moreno B, van Sorge NM, Nizet V, Bier E. Anthrax toxins cooperatively inhibit endocytic recycling by the Rab11/Sec15 exocyst. Nature. 2010; 467: 854–8.
- Fitch CL, de Sousa SM, O'Day PM, Neubert TA, Plantilla CM, Spencer M, et al.. Pertussis toxin expression in Drosophila alters the visual response and blocks eating behaviour. Cell Signal. 1993; 5: 187–207.
- Castonguay-Vanier J, Vial L, Tremblay J, Deziel E. Drosophila melanogaster as a model host for the Burkholderia cepacia complex. PLoS One ; : e. 2010; 5: 11467.
- Moule MG, Monack DM, Schneider DS. Reciprocal analysis of Francisella novicida infections of a Drosophila melanogaster model reveal host-pathogen conflicts mediated by reactive oxygen and imd-regulated innate immune response. PLoS Pathog. 2010; 6: e1001065.
- Santic M, Akimana C, Asare R, Kouokam JC, Atay S, Kwaik YA. Intracellular fate of Francisella tularensis within arthropod-derived cells. Environ Microbiol. 2009; 11: 1473–81.
- Lauriano CM, Barker JR, Yoon SS, Nano FE, Arulanandam BP, Hassett DJ, et al.. MglA regulates transcription of virulence factors necessary for Francisella tularensis intraamoebae and intramacrophage survival. Proc Natl Acad Sci U S A. 2004; 101: 4246–9.
- Ahlund MK, Ryden P, Sjostedt A, Stoven S. Directed screen of Francisella novicida virulence determinants using Drosophila melanogaster. Infect Immun. 2010; 78: 3118–28.
- Aymeric JL, Givaudan A, Duvic B. Imd pathway is involved in the interaction of Drosophila melanogaster with the entomopathogenic bacteria, Xenorhabdus nematophila and Photorhabdus luminescens. Mol Immunol. 2010; 47: 2342–8.
- Valanne S, Kleino A, Myllymaki H, Vuoristo J, Ramet M. Iap2 is required for a sustained response in the Drosophila Imd pathway. Dev Comp Immunol. 2007; 31: 991–1001.
- Avet-Rochex A, Perrin J, Bergeret E, Fauvarque MO. Rac2 is a major actor of Drosophila resistance to Pseudomonas aeruginosa acting in phagocytic cells. Genes Cells. 2007; 12: 1193–204.
- Garver LS, Wu J, Wu LP. The peptidoglycan recognition protein PGRP-SC1a is essential for Toll signaling and phagocytosis of Staphylococcus aureus in Drosophila. Proc Natl Acad Sci U S A. 2006; 103: 660–5.
- Bergeret E, Perrin J, Williams M, Grunwald D, Engel E, Thevenon D, et al.. TM9SF4 is required for Drosophila cellular immunity via cell adhesion and phagocytosis. J Cell Sci. 2008; 121: 3325–34.
- Ayres JS, Freitag N, Schneider DS. Identification of Drosophila mutants altering defense of and endurance to Listeria monocytogenes infection. Genetics. 2008; 178: 1807–15.
- Stoltz DA, Ozer EA, Taft PJ, Barry M, Liu L, Kiss PJ, et al.. Drosophila are protected from Pseudomonas aeruginosa lethality by transgenic expression of paraoxonase-1. J Clin Invest. 2008; 118: 3123–31.
- Broderick KE, Chan A, Balasubramanian M, Feala J, Reed SL, Panda M, et al.. Cyanide produced by human isolates of Pseudomonas aeruginosa contributes to lethality in Drosophila melanogaster. J Infect Dis. 2008; 197: 457–64.
- Apidianakis Y, Mindrinos MN, Xiao W, Lau GW, Baldini RL, Davis RW, et al.. Profiling early infection responses: Pseudomonas aeruginosa eludes host defenses by suppressing antimicrobial peptide gene expression. Proc Natl Acad Sci U S A. 2005; 102: 2573–8.
- Gordon MD, Ayres JS, Schneider DS, Nusse R. Pathogenesis of listeria-infected Drosophila wntD mutants is associated with elevated levels of the novel immunity gene edin. PLoS Pathog. 2008; 4: e1000111.
- Apidianakis Y, Mindrinos MN, Xiao W, Tegos GP, Papisov MI, Hamblin MR, et al.. Involvement of skeletal muscle gene regulatory network in susceptibility to wound infection following trauma. PLoS One. 2007; 2: e1356.
- Foley E, O'Farrell PH. Functional dissection of an innate immune response by a genome-wide RNAi screen. PLoS Biol. 2004; 2: E203.
- Flockhart I, Booker M, Kiger A, Boutros M, Armknecht S, Ramadan N, et al.. FlyRNAi: the Drosophila RNAi screening center database. Nucleic Acids Res. 2006; 34: D489–94.
- Cherry S. RNAi screening for host factors involved in viral infection using Drosophila cells. Methods Mol Biol. 2011; 721: 375–82.
- Clemens JC, Worby CA, Simonson-Leff N, Muda M, Maehama T, Hemmings BA, et al.. Use of double-stranded RNA interference in Drosophila cell lines to dissect signal transduction pathways. Proc Natl Acad Sci U S A. 2000; 97: 6499–503.
- Cheng LW, Viala JP, Stuurman N, Wiedemann U, Vale RD, Portnoy DA. Use of RNA interference in Drosophila S2 cells to identify host pathways controlling compartmentalization of an intracellular pathogen. Proc Natl Acad Sci U S A. 2005; 102: 13646–51.
- Elwell CA, Ceesay A, Kim JH, Kalman D, Engel JN. RNA interference screen identifies Abl kinase and PDGFR signaling in Chlamydia trachomatis entry. PLoS Pathog. 2008; 4: e1000021.
- Sibley CD, Duan K, Fischer C, Parkins MD, Storey DG, Rabin HR, Surette MG. Discerning the complexity of community interactions using a Drosophila model of polymicrobial infections. PLoS Pathog. 2008; 4: e1000184.
- Moy TI, Ball AR, Anklesaria Z, Casadei G, Lewis K, Ausubel FM. Identification of novel antimicrobials using a live-animal infection model. Proc Natl Acad Sci U S A. 2006; 103: 10414–9.
- Kumar PS, Leys EJ, Bryk JM, Martinez FJ, Moeschberger ML, Griffen AL. Changes in periodontal health status are associated with bacterial community shifts as assessed by quantitative 16S cloning and sequencing. J Clin Microbiol. 2006; 44: 3665–73.
- Kumar PS, Griffen AL, Moeschberger ML, Leys EJ. Identification of candidate periodontal pathogens and beneficial species by quantitative 16S clonal analysis. J Clin Microbiol. 2005; 43: 3944–55.
- Kumar PS, Griffen AL, Barton JA, Paster BJ, Moeschberger ML, Leys EJ. New bacterial species associated with chronic periodontitis. J Dent Res. 2003; 82: 338–44.
- Gross EL, Leys EJ, Gasparovich SR, Firestone ND, Schwartzbaum JA, Janies DA, et al.. Bacterial 16S sequence analysis of severe caries in young permanent teeth. J Clin Microbiol. 2010; 48: 4121–8.
- Aas JA, Griffen AL, Dardis SR, Lee AM, Olsen I, Dewhirst FE, et al.. Bacteria of dental caries in primary and permanent teeth in children and young adults. J Clin Microbiol. 2008; 46: 1407–17.
- Hoover CI, Yoshimura F. Transposon-induced pigment-deficient mutants of Porphyromonas gingivalis. FEMS Microbiol Lett. 1994; 124: 43–8.
- Chen T, Dong H, Tang YP, Dallas MM, Malamy MH, Duncan MJ. Identification and cloning of genes from Porphyromonas gingivalis after mutagenesis with a modified Tn4400 transposon from Bacteroides fragilis. Infect Immun. 2000; 68: 420–3.
- Shoji M, Ratnayake DB, Shi Y, Kadowaki T, Yamamoto K, Yoshimura F, et al.. Construction and characterization of a nonpigmented mutant of Porphyromonas gingivalis: cell surface polysaccharide as an anchorage for gingipains. Microbiology. 2002; 148: 1183–91.
- Sato K, Kido N, Murakami Y, Hoover CI, Nakayama K, Yoshimura F. Lipopolysaccharide biosynthesis-related genes are required for colony pigmentation of Porphyromonas gingivalis. Microbiology. 2009; 155: 1282–93.
- Davey ME, Duncan MJ. Enhanced biofilm formation and loss of capsule synthesis: deletion of a putative glycosyltransferase in Porphyromonas gingivalis. J Bacteriol. 2006; 188: 5510–23.
- Sjoberg BM, Torrents E. Shift in ribonucleotide reductase gene expression in Pseudomonas aeruginosa during infection. Infect Immun. 2011; 79: 2663–9.
- Liang H, Duan J, Sibley CD, Surette MG, Duan K. Identification of mutants with altered phenazine production in Pseudomonas aeruginosa. J Med Microbiol. 2011; 60: 22–34.
- Kim SH, Lee BY, Lau GW, Cho YH. IscR modulates catalase A (KatA) activity, peroxide resistance and full virulence of Pseudomonas aeruginosa PA14. J Microbiol Biotechnol. 2009; 19: 1520–6.
- Lee JS, Heo YJ, Lee JK, Cho YH. KatA, the major catalase, is critical for osmoprotection and virulence in Pseudomonas aeruginosa PA14. Infect Immun. 2005; 73: 4399–403.
- Shin DH, Choi YS, Cho YH. Unusual properties of catalase A (KatA) of Pseudomonas aeruginosa PA14 are associated with its biofilm peroxide resistance. J Bacteriol. 2008; 190: 2663–70.
- Lau GW, Britigan BE, Hassett DJ. Pseudomonas aeruginosa OxyR is required for full virulence in rodent and insect models of infection and for resistance to human neutrophils. Infect Immun. 2005; 73: 2550–3.
- Erickson DL, Lines JL, Pesci EC, Venturi V, Storey DG. Pseudomonas aeruginosa relA contributes to virulence in Drosophila melanogaster. Infect Immun. 2004; 72: 5638–45.
- Vogt SL, Green C, Stevens KM, Day B, Erickson DL, Woods DE, et al.. The stringent response is essential for Pseudomonas aeruginosa virulence in the rat lung agar bead and Drosophila melanogaster feeding models of infection. Infect Immun. 2011; 79: 4094–104.
- D'Argenio DA, Gallagher LA, Berg CA, Manoil C. Drosophila as a model host for Pseudomonas aeruginosa infection. J Bacteriol. 2001; 183: 1466–71.
- Yang L, Chen L, Shen L, Surette M, Duan K. Inactivation of MuxABC-OpmB transporter system in Pseudomonas aeruginosa leads to increased ampicillin and carbenicillin resistance and decreased virulence. J Microbiol. 2011; 49: 107–14.
- Potvin E, Lehoux DE, Kukavica-Ibrulj I, Richard KL, Sanschagrin F, Lau GW, et al.. In vivo functional genomics of Pseudomonas aeruginosa for high-throughput screening of new virulence factors and antibacterial targets. Environ Microbiol. 2003; 5: 1294–308.
- Kurz CL, Chauvet S, Andres E, Aurouze M, Vallet I, Michel GP, et al.. Virulence factors of the human opportunistic pathogen Serratia marcescens identified by in vivo screening. EMBO J. 2003; 22: 1451–60.
- Pili-Floury S, Leulier F, Takahashi K, Saigo K, Samain E, Ueda R, et al.. In vivo RNA interference analysis reveals an unexpected role for GNBP1 in the defense against Gram-positive bacterial infection in Drosophila adults. J Biol Chem. 2004; 279: 12848–53.
- Ulvila J, Vanha-Aho LM, Kleino A, Vaha-Makila M, Vuoksio M, Eskelinen S, et al.. Cofilin regulator 14-3-3{zeta} is an evolutionarily conserved protein required for phagocytosis and microbial resistance. J Leukoc Biol. 2011; 89: 649–59.