
Abstract
Antibiotic resistance is an ever-growing problem faced by all major sectors of health care, including dentistry. Recurrent infections related to multidrug-resistant bacteria such as methicillin-resistant Staphylococcus aureus, carbapenem-resistant Enterobacteriaceae, and vancomycin-resistant enterococci (VRE) in hospitals are untreatable and question the effectiveness of notable drugs. Two major reasons for these recurrent infections are acquired antibiotic resistance genes and biofilm formation. None of the traditionally known effective techniques have been able to efficiently resolve these issues. Hence, development of a highly effective antibacterial practice has become inevitable. One example of a hard-to-eradicate pathogen in dentistry is Enterococcus faecalis, which is one of the most common threats observed in recurrent root canal treatment failures, of which the most problematic to treat are its biofilm-forming VRE strains. An effective response against such infections could be the use of bacteriophages (phages). Phage therapy was found to be highly effective against biofilm and multidrug-resistant bacteria and has other advantages like ease of isolation and possibilities for genetic manipulations. The potential of phage therapy in dentistry, in particular against E. faecalis biofilms in root canals, is almost unexplored. Here we review the efforts to develop phage therapy against biofilms. We also focus on the phages isolated against E. faecalis and discuss the possibility of using phages against E. faecalis biofilm in root canals.
Antibiotics, ‘the magic bullets’, have proved to be one of the most revolutionary discoveries of the twentieth century (Citation1, Citation2). However, their overuse and misuse in various cases, including viral and fungal infections, and patient failure to follow the prescribed course have led to a rise in antibiotic-resistant strains, the ‘post antibiotic era’ (Citation3). Consequently, many resistant pathogens like MRSA (methicillin-resistant Staphylococcus aureus), CRE (carbapenem-resistant Enterobacteriaceae), VRE (vancomycin-resistant enterococci) (Citation4, Citation5), multidrug-resistance Pseudomonas and Acinetobacter have developed into major threats. For instance, VRE exhibit resistance to vancomycin, which is considered ‘the last resort’ drug for Gram-positive bacteria, making their elimination almost impossible (Citation6, Citation7). The rate of acquired antibiotic resistance is also alarming. For example, Pseudomonas aeruginosa was shown to rapidly develop resistance against five relevant antibiotics upon exposure to stepwise increased concentrations (Citation8). Apart from being life threating, these antibiotic-resistant strains also lead to elevated health care costs (Citation9). Moreover, failure in surgeries and other medical procedures related to untreatable infections is expected to increase. Having said that, should we be alarmed that we are about to face an era similar to the one prior to the discovery of antibiotics, in which mortality will be caused by common infections?
Today, it is accepted that yet another reason for the failure of antibiotics is the formation of bacterial biofilms (Citation10). Biofilms are defined as dense aggregates of surface-adherent microorganisms that are embedded in a self-produced polymer matrix consisting of polysaccharide, protein, and extracellular DNA (Citation11, Citation12). Biofilms are characterized by the following factors: the environmental conditions and surfaces that favor their formation, the gene products that are required for their formation, the genes that are activated and required to maintain the biofilm, the architecture of the biofilm, and the types of extracellular products that are concentrated in the biofilm matrix (Citation13). According to the National Institutes of Health, biofilms account for more than 60% of the microbial infections in the body (Citation14). These infections can be caused either by a single microbial species or by a mixture of species (multispecies) (Citation15). Some of the common examples of biofilm infections are cystic fibrosis, native valve endocarditis, otitis media, periodontitis, and chronic prostatitis (Citation16).
A major problem of biofilms is their resistance against phagocytosis and their inherent tolerance to the host defense system, to antibiotic therapy, and to disinfectants like chlorine and alcohol, as well as heat (Citation17–Citation19). Factors like poor antibiotic penetration, nutrient limitation and slow bacterial growth, adaptive stress responses in bacteria, and formation of persister cells constitute a multilayered biofilm defense, which cannot easily be overcome (Citation20). The common techniques used for eradication of biofilms include mechanical disruption by physical means, such as tooth brushing and sonication (Citation21), but these are not 100% effective. The difficulties in destructing biofilms necessitate development of alternative ways to prevent and control biofilm-associated clinical infections.
In dentistry, bacterial biofilms are involved in almost all major diseases. Plaques which are actually multispecies biofilms growing on the teeth contain primary colonizers like streptococci on the acquired pellicle and are later colonized by Actinomyces, which may lead to caries (Citation21). Periodontitis is a classic example of a biofilm-mediated disease, which is refractory to antibiotic agents and the host defenses (Citation22). Lastly, the most common biological reason for root canal disease is endodontic biofilm (Citation23, Citation24), which is formed significantly by E. faecalis (Citation25, Citation26) commonly found in previously treated root canals along with other microorganisms.
Currently, endodontic treatment against E. faecalis and other root canal infections involves removing bacteria by biomechanical cleaning, root canal shaping, and disinfection followed by sealing and crown restoration. The purpose of root canal sealing is to provide a tight fluid seal from the coronal and apical part of the tooth. Ideally, endodontic treatment should achieve a sterile root canal system, but given the available materials and techniques, this is undoubtedly impossible (Citation26).
The case study of E. faecalis
E. faecalis is a commensal Gram-positive facultative anaerobic bacterium inhabiting the gastrointestinal tract of humans and various animals, but is also found in environments like soil and water (Citation27–Citation29). E. faecalis is one of the most frequently isolated species from hospital-associated infections; it causes endocarditis, bacteremia, urinary tract infections, meningitis, and other fatal forms of systemic and local infection in humans (Citation30).
The pathogenicity of E. faecalis can be attributed to the various virulence factors reported in clinical strains, including biofilm formation and the expression of surface adhesion components (Citation31). Additional virulence factors identified are hemolysin/bacteriocin, aggregation substance, gelatinase, enterococcal surface protein (Esp), endocarditis-associated antigen, or capsular polysaccharides (Citation32–Citation36). The ability of E. faecalis to adhere to medical devices such as ureteral stents and catheters and to develop biofilms on these devices is likely associated with its pathogenicity (Citation37).
Why is E. faecalis so hard to eradicate?
E. faecalis has become one of the most challenging bacteria to eradicate in the past few decades (Citation30, Citation38). As mentioned above, in root canals it is hidden from the immune system and antibiotics. Various antiseptic and antibiotic materials are used for intracanal bacterial eradication, which include calcium hydroxide or antibiotic pastes to improve bacterial control before root canal sealing. Yet, viable E. faecalis cells were found even after many days in root canals following endodontic treatment, regardless of the use of calcium hydroxide (Citation39–Citation41).
In addition to root canals, E. faecalis is also hard to treat in the gut and other infection sites (Citation30). First, it has many strains that are antibiotic resistant (Citation42–Citation44). Second, the increased use of antibiotics in hospitals worldwide causes dysbiosis, changes in the gut microbiota that are leading to subsequent alterations in the local immune system (Citation45, Citation46). E. faecalis takes advantage of these alterations and takes over ‘the prized niche’ of the gastrointestinal tract, and this niche may be the primary source of organisms that cause enterococcal infections (Citation46–Citation48).
Persistence
Another reason for the difficulty to eradicate E. faecalis infections is its highly recalcitrant nature. This bacterium possesses exceptional surviving abilities and can persist in extreme conditions such as the gut (Citation49) and root canal system (Citation50) as a result of its ability to withstand an alkaline milieu and glucose starvation (Citation51, Citation52).
Antibiotic resistance
In addition to all that, E. faecalis strains are sometimes genetically resistant to antibiotics. According to the Centers for Disease Control and Prevention, VRE strains of E. faecalis are some of the most difficult to treat bacteria (www.cdc.gov/drugresistance/biggest_threats.html). Various studies conducted worldwide have demonstrated increasing rates of VRE-acquired cases; for example, more than 38% of such cases were revealed in Detroit Medical Center, Michigan, in 2009 and 11% of the cases were reported at the national level (Citation53). Vancomycin resistance has five well-recognized phenotypes: VanA, VanB, VanC, VanD, and VanE (Citation54–Citation56). Interestingly, two of these, VanA and VanB, are mediated by newly acquired gene clusters that provide resistant phenotypes primarily in E. faecalis and E. faecium (Citation57). Thus, when such resilient strains as E. faecalis also evolve to be antibiotic-resistant like VRE, it becomes almost impossible to control their infections. Nowadays, linezolid and daptomycin are the last resort drugs often used to treat infections caused by VRE E. faecalis (Citation58, Citation59). However, strains resistant even to these antibiotics have emerged (Citation60, Citation61). A recent study suggested that clinical samples not only had vancomycin-resistant E. faecalis but the isolates also showed resistance to tetracycline, linezolid, and ampicillin (Citation62).
E. faecalis in root canals
In dentistry E. faecalis is one of the main bacteria associated with chronic apical periodontitis in failed root canal treatments. Despite the fact that endodontic infections have a polymicrobial nature, the root canal environment may favor and support the survival of one species, which is commonly E. faecalis. Although E. faecalis is actually seldom present in primary endodontic infections, in cases of postendodontic treatment with apical periodontitis, failed cases are approximately nine times more likely to harbor E. faecalis than cases of primary infections (Citation63, Citation64). Furthermore, the prevalence of E. faecalis in periradicular disease including secondary endodontic infections was reported to be 33% (Citation65) and 24 to 77% in persistent infections (Citation50, Citation63) (Citation66) resulting in the development of lymphadenitis abscesses and cellulitis (Citation26, Citation46).
The way E. faecalis causes failure of root canal treatment is by entering via micro-leakage in faulty restorations, direct pulp exposure in cases of physical barrier breaks, and the gingival sulcus that reaches the pulp chamber through the periodontal membrane (Citation67). After penetrating the dentinal tubules, the root canal serves as a reservoir for bacteria that remain in the root canals protected from the immune system. These bacteria cause constant intracanal infections, endodontic diseases, and refractory or persistent periapical diseases (Citation52, Citation68–Citation70). They can also adhere to dentin collagen (main organic component of dentine), invade the dentinal tubules, and therefore withstand root canal debridement (Citation70).
In addition, E. faecalis contaminations were found to correlate with periodontitis where it was found to be prevalence in root canals of teeth with apical periodontitis requiring endodontic retreatment, or in saliva (Citation71).
The current infection control techniques in root canals fall short of the desired effectiveness against persistent infections. As antibiotics are useless, the endodontic treatment aims to eradicate bacteria from root canal and dentin tubules by mechanical removal of infected tissues and concomitant chemical treatment with antiseptic solution such as sodium hypochlorite and chlorhexidine (mechanochemical preparation). Despite these procedures, bacterial contamination, mainly E. faecalis, is histologically evident in dentine tubules (Citation72, Citation73). Furthermore, one of the disadvantages of root canal debridement is that it cannot prevent root canal late reinfection that may originate from the previously infected dentinal tubules. Ex vivo and clinical studies have shown that in spite of a temporary absence of bacteria following chemo-mechanical preparation, bacteria reappear following successive endodontic appointments. Antiseptic rinsing or antibacterial dressing reduces the bacterial counts; however, it does not completely eliminate the infecting bacteria (Citation74, Citation75). This suggests that intratubular bacteria may serve as a reservoir, out of reach of endodontic preparation.
Biofilms of E. faecalis in root canals
Biofilms, layers of bacteria growing together in a cooperative manner (Citation76, Citation77), are mechanically and physiologically more protected from antibiotics than planktonic cells (Citation78). This can be because most of the antimicrobial agents cannot penetrate into the deeply formed layers of bacteria in a biofilm. They can kill only the peripheral layers, and once the effect of the antibacterial agent has diminished, the surviving bacteria can form new layers of biofilm. The genetic basis of biofilm formation by E. faecalis is largely unknown. A recent study suggested that a specific enterococcus cell surface protein (Esp) is critical for biofilm formation by this organism (Citation79). Complete sterilization of an infected root canal is an important challenge in endodontic treatment, as because of the complexity of the root canal system, the traditional methods often cannot achieve sterilization (Citation80). Various protective measures of the E. faecalis biofilm increase its resistance to antibacterial treatment. This includes resistance to traditional antibacterial rinsing solutions such as chlorhexidine or sodium hypochlorite and the ability to adapt and grow in the presence of calcium hydroxide (Citation51, Citation81). Biofilm islands were reported to exist between the root canal filling and dentin walls despite root canal treatment (Citation82, Citation83). In general, it is found that resistance of biofilm to antibiotics may even increase up to 100–1,000-fold (Citation84). The physical removal of biofilm by endodontic instruments is only partially effective as biofilm may hide in areas unreachable by these instruments (Citation85). Moreover, any surviving biofilm may potentially recover, grow further, and spread apically, thus perpetuating the chronic apical periodontitis.
Phage therapy: is it the answer?
The increasing number of cases with infections related to antibiotic-resistant bacterial strains and biofilm formation, coupled with the failure of conventional measures to deal with them, necessitate the development and implementation of alternative methods. The use of bacteriophages against pathogenic bacteria, termed ‘phage therapy’, is one of the most promising methods being explored by scientists around the globe. A bacteriophage or phage is a virus that specifically targets and destroys disease-causing bacteria by invading bacterial cells, disrupting their metabolism, and causing lysis. Their lifecycle could also be lysogenic; however, for phage therapy, only lytic phages are used.
Phages were first discovered by Fedrerick Twort in 1915. Felix d'Herelle developed the use of phages to treat various infectious diseases between 1917 and 1940. Along with George Eliava, he founded the George Eliava Institute in Tbilisi, Georgia, which uses phage therapy against bacterial infections even today. However, when Alexander Fleming discovered penicillin in 1928, its rapid success overtook the interest in phage therapy. The emergence of new resistant strains and the acceptance that biofilm formation is a major problem leading to treatment failure has recently rekindled the interest in phage therapy. Phage therapy offers various benefits over antibiotics (Citation86):
High specificity: The phages target specific pathogenic bacteria and pose no harm to the commensal microbiome of the body (Citation87).
Ease of isolation: Phages are bacteria-dependent and hence can be found wherever their target bacteria are present. It seems that each bacterium may have hundreds of phages as reflected, for example, in the collection of PhageDB, where 1,153 and 116 phages were isolated against Mycobacterium and Arthrobacter, respectively (PhageDB.org).
Possibility for clinical improvement: With the development of molecular biology and genetic engineering, the possibility of transforming phages increased in the past decades.
‘Single shot’: Phage multiplication occurs in correlation with the growth of the bacteria. Hence, theoretically only a single shot is required (Citation88).
No residual: Once the pathogenic bacteria have been completely eradicated from the host cell due to phage-induced lysis, because the phages can no longer grow without the bacterial host, it is expected that they will be shed from the cell without any harm to the cell (Citation89).
Biofilms destruction: Biofilm destruction by phages is much more efficient when compared with antibiotics () (Citation90). The phages can infect the bacteria present in the upper layer of the biofilm and replicate to form new phages, which can penetrate the inner layers and infect the remaining bacteria. The continuous replication of the phages and subsequent infection and killing of biofilm bacteria lead to their eradication. Moreover, when the target bacteria are killed, the phage particles are subsequently eliminated as they no longer have a host. As shown in , numerous experiments have been performed to date using single phages or phage cocktails or even a combination of phages and antibiotics against biofilms. To a large extent, all these trials successfully eliminated the targeted bacterial biofilms from the infection sites or reduced bacterial counts to noticeably lower numbers. Also, various reports of natural lytic phages with phage-borne polysaccharide depolymerases have shown that phage-induced lysis and extracellular polysaccharide degradation are used in combination in natural systems to reduce bacterial biofilms (Citation91–Citation95). These depolymerases appear to be carried on the surface of phages and degrade bacterial capsular polysaccharides to allow access to bacterial cell surfaces (Citation93). In addition, biofilm dispersion can be achieved by engineering a bacteriophage to express a biofilm-degrading enzyme during infection (Citation96). Thus, using bacteriophages as single phage or a combination of phages in cocktails could be a good approach for the treatment of biofilms in infectious bacterial diseases.
Fig. 1. Comparative analysis of the actions antibiotics and phages have on a mature biofilm. Antibiotics fail to penetrate the biofilm and only kill the bacteria superficially, and are thus unable to eradicate the biofilm. Phages, on the other hand, can infect bacterial cells on the outer layer of the biofilm, multiply, and in a chain reaction penetrate into the deeper layers, resulting in complete eradication of the biofilm in a single shot.
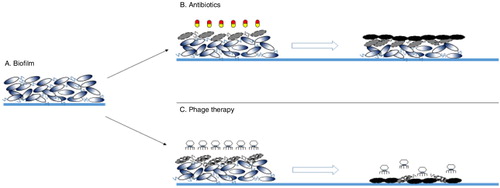
Table 1. Phage therapy trials on bacterial biofilms using different model systems
Phage therapy and human safety
Given the numerous advantages and the potential benefits of bacteriophages over the failing antibiotics, it is no wonder that scientists worldwide are delving into phage therapy. To the uninitiated, phage therapy not only holds great promise, but sets off alarm bells as well: is it safe for humans? This concern, although understandable, is virtually unfounded, as this method has been in use treating humans for decades. Indeed, it was first used over a century ago in France by Felix d'Herelle in 1919 to treat children suffering from severe dysentery (Citation110). Since then, many such trials have been conducted in France, Georgia, Poland, and many other places worldwide (Citation111, Citation112).
A recent trial was successfully performed in a patient suffering from an eye infection by Fadlallah et al., in 2015 at the Phage Therapy Center in Tbilisi, Georgia (Citation113). Many similar trials have achieved success without any harmful side effects (Citation114–Citation117). Phages were also found to have almost no harmful effects on the non-target microbiome (Citation118). Interestingly, phages exist everywhere in nature, and although up to date they have never been found to cause any harm or diseases in humans, there are certain temperate phages that might contain exotoxins that have harmful effects (Citation119–Citation121). However, pharmacological studies coupled with genetic tools, which are well-established nowadays, can help choose and purify the right target phage and remove unwanted virulence genes. Thus, in principal, phage therapy can be considered as a relatively safe technique (Citation122).
Phages against E. faecalis
For combatting VRE E. faecalis infections, phages have been isolated and tested for their efficacy by several researchers (). Most of these phages belong to the Myoviridae or the Siphoviridae families of tailed phages. In case of the phage IME-EF1, when administrated intraperitoneally in a murine sepsis model, one dose of IME-EF1 or its endolysin was found to reduce the bacterial blood count and protect the mice from a lethal challenge of E. faecalis, with a survival rate of 60 or 80% (Citation123). Similarly, in vivo therapeutic potential of ϕEF24C evaluated in a sepsis BALB/c mouse model proved to be effective at a low concentration with no host sensitivity and no change in mouse lethality following a single or repeated phage exposure (Citation124). In addition, the Q69 phage eradicated an E. faecalis strain mainly responsible for biogenic amines (BA) tyramine accumulation in food, which is considered as a toxicological hazard (Citation125).
Table 2. Phages isolated against E. faecalis, their accession numbers, and the family they belong to. This indicates the huge possibility of using these phages for phage therapy in the future.
Phage therapy against E. faecalis biofilms
Apart from planktonic bacteria eradication, a more challenging and relevant part of E. faecalis infections is eliminating its biofilms. So far, among the E. faecalis described phages (), only EFDG1 was tested on E. faecalis biofilms (Citation128). EFDG1, isolated from sewage water, was very efficient in nearly eliminating a 2-week-old E. faecalis biofilm of ~100 µM thickness. Evaluating the biofilm biomass showed a fivefold reduction within 7 days in the phage-treated samples compared with the untreated biofilms, which were stable and showed no reduction. Bacterial viable counts from the biofilm supported this notion by showing a five log reduction compared with the untreated biofilms. Scanning electron microscopy revealed the destruction of the treated biofilm which looks like clumps of distributed bacteria in comparison with the intact, untreated biofilm ().
Fig. 2. Scanning electron microscopy images of untreated and treated biofilms with the phage EFDG1. The image on the left shows a well-developed biofilm, while the right image is one with EFDG1 phage treatment. Both the biofilms are 2 weeks old.

The activity of EFDG1 was further tested in post-treated root canal infections (Citation128) using an ex vivo two-chamber bacterial leakage model of human teeth (Citation139). Measurements of bacterial leakage from the root apex showed that the obturated root canals subjected to EFDG1 irrigation resulted in dramatic reduction of eight logs in bacterial leakage compared with the conventional sample. Confocal microscopy images of horizontal root sections demonstrated that live bacteria were evident in the dentinal tubules of the control group, whereas dead bacteria were seen in the phage-treated teeth (Citation128).
These results indicate that phage therapy might be a worthy additive solution in combatting E. faecalis biofilms in root canals where all other anti-infective and aseptic technique strategies, including the current use of increased apical preparation sizes, and inclusion of chlorhexidine in combination with sodium hypochlorite, fail (Citation66, Citation140) (Citation141).
Future perspectives
A lot is known about the importance of phages in nature (Citation142, Citation143); however, the phage–bacteria interaction in the oral microbiome still needs to be explored. Moreover, using phages to remove specific bacteria from the microbiome will allow us to study the role of their host in the microbiome and identify keystone pathogens in various infections. Thus, the use of phages will be beneficial both in gaining knowledge about oral pathogens and in removing them. Understanding the oral microbiome with the help of phages can potentially lead to the development of ‘microbiome engineering’ to prevent infections. Using ‘good’ bacteria as a probiotic (Citation118) and phages against the pathogens might be a new avenue yet to be explored in oral health. However, the inadequate number of phages which can specifically target oral bacteria raise the need for the isolation and characterization of more phages against oral pathogens, for example, the ones responsible for root canal infections.
In conclusion, considering all the available positive outcomes from the usage of phages against not only E. faecalis but also other bacteria in biofilms, phage therapy appears to be a tool against infectious biofilms. In the future, phages such as EFDG1 and other phages of E. faecalis like phiEF24C, IME-EF1, and EFLK1 can be used either as cocktails or as combinations with antibiotics to combat VRE E. faecalis in dental biofilms. In root canal treatments, although alternative antibacterial irrigants (such as chlorhexidine and sodium hypochlorite) were shown to be effective, they still do not prevent recurrent E. faecalis infections. Consequently, combinations of anti E. faecalis phages and antibacterial agents can benefit the host by reducing the chances of recurrent infections.
Conflict of interest and funding
There is no conflict of interest in the present study for any of the authors.
Acknowledgements
The authors thank the Rosetrees trust for kindly funding this work and the Ariane de Rothschild Women Doctorial program for funding LK. They also thank Elisheva Dorfman for editing the manuscript.
Related Research Data
References
- Gradmann C. Re-inventing infectious disease: antibiotic resistance and drug development at the Bayer Company 1945–80. Med Hist. 2016; 60: 155–80.
- Gradmann C. Magic bullets and moving targets: antibiotic resistance and experimental chemotherapy, 1900–1940. Dynamis. 2011; 31: 305–21.
- Price NL, Goyette-Desjardins G, Nothaft H, Valguarnera E, Szymanski CM, Segura M, etal. Glycoengineered outer membrane vesicles: a novel platform for bacterial vaccines. Sci Rep. 2016; 6: 24931.
- Drusano GL, Louie A, MacGowan A, Hope W. Suppression of emergence of resistance in pathogenic bacteria: keeping our powder dry. Part 1. Antimicrob Agents Chemother. 2016; 60: 1183–93.
- Drusano GL, Hope W, MacGowan A, Louie A. Suppression of emergence of resistance in pathogenic bacteria: keeping our powder dry, Part 2. Antimicrob Agents Chemother. 2016; 60: 1194–201.
- O'Driscoll T, Crank CW. Vancomycin-resistant enterococcal infections: epidemiology, clinical manifestations, and optimal management. Infect Drug Resist. 2015; 8: 217–30.
- Yarlagadda V, Sarkar P, Samaddar S, Haldar J. A vancomycin derivative with a Pyrophosphate-Binding Group: a strategy to combat vancomycin-resistant bacteria. Angew Chem Int Ed Engl. 2016; 55: 7836–40.
- Feng Y, Jonker MJ, Moustakas I, Brul S, Ter Kuile BH. Dynamics of mutations during development of resistance by Pseudomonas aeruginosa against five antibiotics. Antimicrob Agents Chemother. 2016; 60: 4229–36.
- Durai R, Ng PC, Hoque H. Methicillin-resistant Staphylococcus aureus: an update. Aorn J. 2010; 91: 599–606. quiz 7–9.
- Emami S, Nikokar I, Ghasemi Y, Ebrahimpour M, Sedigh Ebrahim-Saraie H, Araghian A, etal. Antibiotic resistance pattern and distribution of pslA Gene among biofilm producing Pseudomonas aeruginosa isolated from waste water of a burn center. Jundishapur J Microbiol. 2015; 8: e23669.
- Hoiby N, Ciofu O, Johansen HK, Song ZJ, Moser C, Jensen PO, etal. The clinical impact of bacterial biofilms. Int J Oral Sci. 2011; 3: 55–65.
- Bhinu VS. Insight into biofilm-associated microbial life. J Mol Microbiol Biotechnol. 2005; 10: 15–21.
- Karatan E, Watnick P. Signals, regulatory networks, and materials that build and break bacterial biofilms. Microbiol Mol Biol Rev. 2009; 73: 310–47.
- Lewis K. Riddle of biofilm resistance. Antimicrob Agents Chemother. 2001; 45: 999–1007.
- Romling U, Balsalobre C. Biofilm infections, their resilience to therapy and innovative treatment strategies. J Intern Med. 2012; 272: 541–61.
- Donlan RM. Biofilms: microbial life on surfaces. Emerg Infect Dis. 2002; 8: 881–90.
- Hoiby N, Bjarnsholt T, Givskov M, Molin S, Ciofu O. Antibiotic resistance of bacterial biofilms. Int J Antimicrob Agents. 2010; 35: 322–32.
- Donlan RM, Costerton JW. Biofilms: survival mechanisms of clinically relevant microorganisms. Clin Microbiol Rev. 2002; 15: 167–93.
- Bradley CR, Fraise AP. Heat and chemical resistance of enterococci. J Hosp Infect. 1996; 34: 191–6.
- Stewart PS. Mechanisms of antibiotic resistance in bacterial biofilms. Int J Med Microbiol. 2002; 292: 107–13.
- Aparna MS, Yadav S. Biofilms: microbes and disease. Braz J Infect Dis. 2008; 12: 526–30.
- Schaudinn C, Gorur A, Keller D, Sedghizadeh PP, Costerton JW. Periodontitis: an archetypical biofilm disease. J Am Dent Assoc. 2009; 140: 978–86.
- Ricucci D, Siqueira JF Jr. Biofilms and apical periodontitis: study of prevalence and association with clinical and histopathologic findings. J Endod. 2010; 36: 1277–88.
- Nair PN. On the causes of persistent apical periodontitis: a review. Int Endod J. 2006; 39: 249–81.
- Mohammadi Z, Soltani MK, Shalavi S. An update on the management of endodontic biofilms using root canal irrigants and medicaments. Iran Endod J. 2014; 9: 89–97.
- Pericas JM, Zboromyrska Y, Cervera C, Castaneda X, Almela M, Garcia-de-la-Maria C, etal. Enterococcal endocarditis revisited. Future Microbiol. 2015; 10: 1215–40.
- Lebreton F, Willems RJL, Gilmore MS, Gilmore MS, Clewell DB, Ike Y, Shankar N. Enterococcus diversity, origins in nature, and gut colonization. Enterococci: from commensals to leading causes of Drug resistant infection. 2014; Boston: MA: Massachusetts Eye and Ear Infirmary. , PMID: 24649513.
- Palmer KL, van Schaik W, Willems RJL, Gilmore MS, Gilmore MS, Clewell DB, Ike Y, Shankar N. Enterococcal genomics. Enterococci: from commensals to leading causes of drug resistant infection. 2014; Boston: MA: Massachusetts Eye and Ear Infirmary. , PMID: 24649511.
- Ramsey M, Hartke A, Huycke M, Gilmore MS, Clewell DB, Ike Y, Shankar N. The physiology and metabolism of Enterococci. Enterococci: from commensals to leading causes of rug resistant infection . 2014; Boston: MA: Massachusetts Eye and Ear Infirmary. , PMID: 24649507.
- Agudelo Higuita NI, Huycke MM. Gilmore MS, Clewell DB, Ike Y, Shankar N. Enterococcal disease, epidemiology, and implications for treatment. Enterococci: from commensals to leading causes of rug resistant infection . 2014; MA: Massachusetts Eye and Ear Infirmary: Boston. , PMID: 24649504.
- Brito CS, Queiroz LL, Campos PA, Batistao DW, Silva Hde A, de Agostini GG, etal. The nares as a CA-MRSA reservoir in the healthy elderly. Rev Soc Bras Med Trop. 2015; 48: 614–16.
- Coque TM, Patterson JE, Steckelberg JM, Murray BE. Incidence of hemolysin, gelatinase, and aggregation substance among enterococci isolated from patients with endocarditis and other infections and from feces of hospitalized and community-based persons. J Infect Dis. 1995; 171: 1223–9.
- Lowe AM, Lambert PA, Smith AW. Cloning of an Enterococcus faecalis endocarditis antigen: homology with adhesins from some oral streptococci. Infect Immun. 1995; 63: 703–6.
- Eaton TJ, Gasson MJ. Molecular screening of Enterococcus virulence determinants and potential for genetic exchange between food and medical isolates. Appl Environ Microbiol. 2001; 67: 1628–35.
- Baeten JM, McClelland RS, Overbaugh J, Richardson BA, Emery S, Lavreys L, etal. Vitamin A supplementation and human immunodeficiency virus type 1 shedding in women: results of a randomized clinical trial. J Infect Dis. 2002; 185: 1187–91.
- Theilacker C, Krueger WA, Kropec A, Huebner J. Rationale for the development of immunotherapy regimens against enterococcal infections. Vaccine. 2004; 22(Suppl 1): S31–8.
- Mohamed JA, Huang DB. Biofilm formation by enterococci. J Med Microbiol. 2007; 56: 1581–8.
- Guzman Prieto AM, van Schaik W, Rogers MR, Coque TM, Baquero F, Corander J, etal. Global emergence and dissemination of enterococci as nosocomial pathogens: attack of the clones?. Front Microbiol. 2016; 7: 788.
- Sjogren U, Figdor D, Spangberg L, Sundqvist G. The antimicrobial effect of calcium hydroxide as a short-term intracanal dressing. Int Endod J. 1991; 24: 119–25.
- Vivacqua-Gomes N, Gurgel-Filho ED, Gomes BP, Ferraz CC, Zaia AA, Souza-Filho FJ. Recovery of Enterococcus faecalis after single- or multiple-visit root canal treatments carried out in infected teeth ex vivo . Int Endod J. 2005; 38: 697–704.
- Bystrom A, Sundqvist G. Bacteriologic evaluation of the efficacy of mechanical root canal instrumentation in endodontic therapy. Scand J Dent Res. 1981; 89: 321–8.
- Sahm DF, Kissinger J, Gilmore MS, Murray PR, Mulder R, Solliday J, etal. In vitro susceptibility studies of vancomycin-resistant Enterococcus faecalis . Antimicrob Agents Chemother. 1989; 33: 1588–91.
- Huycke MM, Sahm DF, Gilmore MS. Multiple-drug resistant enterococci: the nature of the problem and an agenda for the future. Emerg Infect Dis. 1998; 4: 239–49.
- Miller WR, Murray BE, Rice LB, Arias CA. Vancomycin-resistant enterococci: therapeutic challenges in the 21st century. Infect Dis Clin North Am. 2016; 30: 415–39.
- Belkaid Y, Hand TW. Role of the microbiota in immunity and inflammation. Cell. 2014; 157: 121–41.
- Arias CA, Murray BE. The rise of the Enterococcus: beyond vancomycin resistance. Nat Rev Microbiol. 2012; 10: 266–78.
- Brandl K, Plitas G, Mihu CN, Ubeda C, Jia T, Fleisher M, etal. Vancomycin-resistant enterococci exploit antibiotic-induced innate immune deficits. Nature. 2008; 455: 804–7.
- Park SY, Shin YP, Kim CH, Park HJ, Seong YS, Kim BS, etal. Immune evasion of Enterococcus faecalis by an extracellular gelatinase that cleaves C3 and iC3b. J Immunol. 2008; 181: 6328–36.
- Fisher K, Phillips C. The ecology, epidemiology and virulence of Enterococcus . Microbiology. 2009; 155: 1749–57.
- Sedgley C, Buck G, Appelbe O. Prevalence of Enterococcus faecalis at multiple oral sites in endodontic patients using culture and PCR. J Endod. 2006; 32: 104–9.
- Liu H, Wei X, Ling J, Wang W, Huang X. Biofilm formation capability of Enterococcus faecalis cells in starvation phase and its susceptibility to sodium hypochlorite. J Endod. 2010; 36: 630–5.
- Saleh IM, Ruyter IE, Haapasalo M, Orstavik D. Survival of Enterococcus faecalis in infected dentinal tubules after root canal filling with different root canal sealers in vitro . Int Endod J. 2004; 37: 193–8.
- Hayakawa K, Marchaim D, Palla M, Gudur UM, Pulluru H, Bathina P, etal. Epidemiology of vancomycin-resistant Enterococcus faecalis: a case-case-control study. Antimicrob Agents Chemother. 2013; 57: 49–55.
- Arthur M, Courvalin P. Genetics and mechanisms of glycopeptide resistance in enterococci. Antimicrob Agents Chemother. 1993; 37: 1563–71.
- Fines M, Perichon B, Reynolds P, Sahm DF, Courvalin P. VanE, a new type of acquired glycopeptide resistance in Enterococcus faecalis BM4405. Antimicrob Agents Chemother. 1999; 43: 2161–4.
- Perichon B, Reynolds P, Courvalin P. VanD-type glycopeptide-resistant Enterococcus faecium BM4339. Antimicrob Agents Chemother. 1997; 41: 2016–18.
- Palmer SM, Rybak MJ. Vancomycin-resistant enterococci. Pharmacotherapy. 1996; 16: 819–29.
- Britt NS, Potter EM, Patel N, Steed ME. Comparison of the effectiveness and safety of linezolid and daptomycin in vancomycin-resistant enterococcal bloodstream infection: a national cohort study of veterans affairs patients. Clin Infect Dis. 2015; 61: 871–8.
- Rincon S, Panesso D, Diaz L, Carvajal LP, Reyes J, Munita JM, etal. Resistance to ‘last resort’ antibiotics in Gram-positive cocci: the post-vancomycin era. Biomedica. 2014; 34(Suppl 1): 191–208.
- Johnson AP, Tysall L, Stockdale MV, Woodford N, Kaufmann ME, Warner M, etal. Emerging linezolid-resistant Enterococcus faecalis and Enterococcus faecium isolated from two Austrian patients in the same intensive care unit. Eur J Clin Microbiol Infect Dis. 2002; 21: 751–4.
- Munoz-Price LS, Lolans K, Quinn JP. Emergence of resistance to daptomycin during treatment of vancomycin-resistant Enterococcus faecalis infection. Clin Infect Dis. 2005; 41: 565–6.
- Talebi M, Asghari Moghadam N, Mamooii Z, Enayati M, Saifi M, Pourshafie MR. Antibiotic resistance and biofilm formation of Enterococcus faecalis in patient and environmental samples. Jundishapur J Microbiol. 2015; 8: e23349.
- Rocas IN, Siqueira JF Jr., Santos KR. Association of Enterococcus faecalis with different forms of periradicular diseases. J Endod. 2004; 30: 315–20.
- Zhang C, Du J, Peng Z. Correlation between Enterococcus faecalis and persistent intraradicular infection compared with primary intraradicular infection: a systematic review. J Endod. 2015; 41: 1207–13.
- Tennert C, Fuhrmann M, Wittmer A, Karygianni L, Altenburger MJ, Pelz K, etal. New bacterial composition in primary and persistent/secondary endodontic infections with respect to clinical and radiographic findings. J Endod. 2014; 40: 670–7.
- Stuart CH, Schwartz SA, Beeson TJ, Owatz CB. Enterococcus faecalis: its role in root canal treatment failure and current concepts in retreatment. J Endod. 2006; 32: 93–8.
- Narayanan LL, Vaishnavi C. Endodontic microbiology. J Conserv Dent. 2010; 13: 233–9.
- Al-Nazhan S, Al-Sulaiman A, Al-Rasheed F, Alnajjar F, Al-Abdulwahab B, Al-Badah A. Microorganism penetration in dentinal tubules of instrumented and retreated root canal walls. In vitro SEM study. Restor Dent Endod. 2014; 39: 258–64.
- Souto R, Colombo AP. Prevalence of Enterococcus faecalis in subgingival biofilm and saliva of subjects with chronic periodontal infection. Arch Oral Biol. 2008; 53: 155–60.
- Love RM. Enterococcus faecalis – a mechanism for its role in endodontic failure. Int Endod J. 2001; 34: 399–405.
- Wang QQ, Zhang CF, Chu CH, Zhu XF. Prevalence of Enterococcus faecalis in saliva and filled root canals of teeth associated with apical periodontitis. Int J Oral Sci. 2012; 4: 19–23.
- Berutti E, Marini R, Angeretti A. Penetration ability of different irrigants into dentinal tubules. J Endod. 1997; 23: 725–7.
- Zou L, Shen Y, Li W, Haapasalo M. Penetration of sodium hypochlorite into dentin. J Endod. 2010; 36: 793–6.
- Chavez De Paz LE, Dahlen G, Molander A, Moller A, Bergenholtz G. Bacteria recovered from teeth with apical periodontitis after antimicrobial endodontic treatment. Int Endod J. 2003; 36: 500–8.
- Distel JW, Hatton JF, Gillespie MJ. Biofilm formation in medicated root canals. J Endod. 2002; 28: 689–93.
- Kolenbrander PE. Oral microbial communities: biofilms, interactions, and genetic systems. Annu Rev Microbiol. 2000; 54: 413–37.
- Davey ME, O'Toole GA. Microbial biofilms: from ecology to molecular genetics. Microbiol Mol Biol Rev. 2000; 64: 847–67.
- de la Fuente-Nunez C, Reffuveille F, Fernandez L, Hancock RE. Bacterial biofilm development as a multicellular adaptation: antibiotic resistance and new therapeutic strategies. Curr Opin Microbiol. 2013; 16: 580–9.
- Toledo-Arana A, Valle J, Solano C, Arrizubieta MJ, Cucarella C, Lamata M, etal. The enterococcal surface protein, Esp, is involved in Enterococcus faecalis biofilm formation. Appl Environ Microbiol. 2001; 67: 4538–45.
- Pan J, Sun K, Liang Y, Sun P, Yang X, Wang J, etal. Cold plasma therapy of a tooth root canal infected with Enterococcus faecalis biofilms in vitro . J Endod. 2013; 39: 105–10.
- Shen Y, Stojicic S, Haapasalo M. Antimicrobial efficacy of chlorhexidine against bacteria in biofilms at different stages of development. J Endod. 2011; 37: 657–61.
- Ricucci D, Siqueira JF Jr., Bate AL, Pitt Ford TR. Histologic investigation of root canal-treated teeth with apical periodontitis: a retrospective study from twenty-four patients. J Endod. 2009; 35: 493–502.
- Vieira AR, Siqueira JF Jr., Ricucci D, Lopes WS. Dentinal tubule infection as the cause of recurrent disease and late endodontic treatment failure: a case report. J Endod. 2012; 38: 250–4.
- Ceri H, Olson ME, Stremick C, Read RR, Morck D, Buret A. The Calgary Biofilm Device: new technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J Clin Microbiol. 1999; 37: 1771–6.
- Lin J, Shen Y, Haapasalo M. A comparative study of biofilm removal with hand, rotary nickel-titanium, and self-adjusting file instrumentation using a novel in vitro biofilm model. J Endod. 2013; 39: 658–63.
- Loc-Carrillo C, Abedon ST. Pros and cons of phage therapy. Bacteriophage. 2011; 1: 111–14.
- Ly-Chatain MH. The factors affecting effectiveness of treatment in phages therapy. Front Microbiol. 2014; 5: 51.
- Capparelli R, Nocerino N, Iannaccone M, Ercolini D, Parlato M, Chiara M, etal. Bacteriophage therapy of Salmonella enterica: a fresh appraisal of bacteriophage therapy. J Infect Dis. 2010; 201: 52–61.
- Clokie MR, Millard AD, Letarov AV, Heaphy S. Phages in nature. Bacteriophage. 2011; 1: 31–45.
- Stambolis C. [On the nature and significance of nodular renal blastoma (author's transl)]. Zentralbl Allg Pathol. 1979; 123: 3–8.
- Cornelissen A, Ceyssens PJ, T'Syen J, Van Praet H, Noben JP, Shaburova OV, etal. The T7-related Pseudomonas putida phage phi15 displays virion-associated biofilm degradation properties. PLoS One. 2011; 6: e18597.
- Hughes KA, Sutherland IW, Clark J, Jones MV. Bacteriophage and associated polysaccharide depolymerases – novel tools for study of bacterial biofilms. J Appl Microbiol. 1998; 85: 583–90.
- Hughes KA, Sutherland IW, Jones MV. Biofilm susceptibility to bacteriophage attack: the role of phage-borne polysaccharide depolymerase. Microbiology. 1998; 144: 3039–47.
- Vivaldi EA. [Informatics in the School of Medicine of the University of Chile. II. The school network and access to data bases]. Rev Med Chil. 1990; 118: 1387–92.
- Gutierrez D, Vandenheuvel D, Martinez B, Rodriguez A, Lavigne R, Garcia P. Two phages, phiIPLA-RODI and phiIPLA-C1C, Lyse Mono- and Dual-Species Staphylococcal Biofilms. Appl Environ Microbiol. 2015; 81: 3336–48.
- Lu TK, Collins JJ. Dispersing biofilms with engineered enzymatic bacteriophage. Proc Natl Acad Sci USA. 2007; 104: 11197–202.
- Fu W, Forster T, Mayer O, Curtin JJ, Lehman SM, Donlan RM. Bacteriophage cocktail for the prevention of biofilm formation by Pseudomonas aeruginosa on catheters in an in vitro model system. Antimicrob Agents Chemother. 2010; 54: 397–404.
- Alemayehu D, Casey PG, McAuliffe O, Guinane CM, Martin JG, Shanahan F, etal. Bacteriophages phiMR299-2 and phiNH-4 can eliminate Pseudomonas aeruginosa in the murine lung and on cystic fibrosis lung airway cells. MBio. 2012; 3: e00029–12.
- Basu S, Agarwal M, Kumar Bhartiya S, Nath G, Kumar Shukla V. An in vivo wound model utilizing bacteriophage therapy of Pseudomonas aeruginosa biofilms. Ostomy Wound Manage. 2015; 61: 16–23.
- Nzakizwanayo J, Hanin A, Alves DR, McCutcheon B, Dedi C, Salvage J, etal. Bacteriophage can prevent encrustation and blockage of urinary catheters by Proteus mirabilis . Antimicrob Agents Chemother. 2015; 60: 1530–6.
- Curtin JJ, Donlan RM. Using bacteriophages to reduce formation of catheter-associated biofilms by Staphylococcus epidermidis . Antimicrob Agents Chemother. 2006; 50: 1268–75.
- Seth AK, Geringer MR, Nguyen KT, Agnew SP, Dumanian Z, Galiano RD, etal. Bacteriophage therapy for Staphylococcus aureus biofilm-infected wounds: a new approach to chronic wound care. Plast Reconstr Surg. 2013; 131: 225–34.
- Sillankorva S, Oliveira D, Moura A, Henriques M, Faustino A, Nicolau A, etal. Efficacy of a broad host range lytic bacteriophage against Escherichia coli adhered to urothelium. Curr Microbiol. 2011; 62: 1128–32.
- Ahiwale S, Tamboli N, Thorat K, Kulkarni R, Ackermann H, Kapadnis B. In vitro management of hospital Pseudomonas aeruginosa biofilm using indigenous T7-like lytic phage. Curr Microbiol. 2011; 62: 335–40.
- Sillankorva S, Oliveira R, Vieira MJ, Azeredo J. Real-time quantification of Pseudomonas fluorescens cell removal from glass surfaces due to bacteriophage varphiS1 application. J Appl Microbiol. 2008; 105: 196–202.
- Yilmaz C, Colak M, Yilmaz BC, Ersoz G, Kutateladze M, Gozlugol M. Bacteriophage therapy in implant-related infections: an experimental study. J Bone Joint Surg Am. 2013; 95: 117–25.
- Rahman M, Kim S, Kim SM, Seol SY, Kim J. Characterization of induced Staphylococcus aureus bacteriophage SAP-26 and its anti-biofilm activity with rifampicin. Biofouling. 2011; 27: 1087–93.
- Phee A, Bondy-Denomy J, Kishen A, Basrani B. Azarpazhooh A, Maxwell K. Efficacy of bacteriophage treatment on Pseudomonas aeruginosa biofilms. J Endod. 2013; 39: 364–9.
- Paisano AF, Spira B, Cai S, Bombana AC. In vitro antimicrobial effect of bacteriophages on human dentin infected with Enterococcus faecalis ATCC 29212. Oral Microbiol Immunol. 2004; 19: 327–30.
- Summers WC. Felix d'Herelle and the origins of molecular biology. 1999; New Haven, CT: Yale University Press.
- Kutter E, De Vos D, Gvasalia G, Alavidze Z, Gogokhia L, Kuhl S, etal. Phage therapy in clinical practice: treatment of human infections. Curr Pharm Biotechnol. 2010; 11: 69–86.
- Kutter EM, Kuhl SJ, Abedon ST. Re-establishing a place for phage therapy in Western medicine. Future Microbiol. 2015; 10: 685–8.
- Fadlallah A, Chelala E, Legeais JM. Corneal infection therapy with topical bacteriophage administration. Open Ophthalmol J. 2015; 9: 167–8.
- Brussow H. What is needed for phage therapy to become a reality in Western medicine?. Virology. 2012; 434: 138–42.
- Bruttin A, Brussow H. Human volunteers receiving Escherichia coli phage T4 orally: a safety test of phage therapy. Antimicrob Agents Chemother. 2005; 49: 2874–8.
- Rose T, Verbeken G, Vos DD, Merabishvili M, Vaneechoutte M, Lavigne R, etal. Experimental phage therapy of burn wound infection: difficult first steps. Int J Burns Trauma. 2014; 4: 66–73.
- Sarker SA, Sultana S, Reuteler G, Moine D, Descombes P, Charton F, etal. Oral phage therapy of acute bacterial iarrhea with two coliphage preparations: a andomized trial in children from Bangladesh. EBioMedicine. 2016; 4: 124–37.
- Abedon ST, Kuhl SJ, Blasdel BG, Kutter EM. Phage treatment of human infections. Bacteriophage. 2011; 66–85.
- Casas V, Maloy S. Role of bacteriophage-encoded exotoxins in the evolution of bacterial pathogens. Future Microbiol. 2011; 6: 1461–73.
- Abedon ST, Lejeune JT. Why bacteriophage encode exotoxins and other virulence factors. Evol Bioinform Online. 2005; 1: 97–110.
- Casas V, Magbanua J, Sobrepena G, Kelley ST, Maloy SR. Reservoir of bacterial exotoxin genes in the environment. Int J Microbiol. 2010; 2010: 754368.
- Abedon ST, Thomas-Abedon C. Phage therapy pharmacology. Curr Pharm Biotechnol. 2010; 11: 28–47.
- Zhang W, Mi Z, Yin X, Fan H, An X, Zhang Z, etal. Characterization of Enterococcus faecalis phage IME-EF1 and its endolysin. PLoS One. 2013; 8: e80435.
- Uchiyama J, Rashel M, Takemura I, Wakiguchi H, Matsuzaki S. In silico and in vivo evaluation of bacteriophage phiEF24C, a candidate for treatment of Enterococcus faecalis infections. Appl Environ Microbiol. 2008; 74: 4149–63.
- Ladero V, Gomez-Sordo C, Sanchez-Llana E, Del Rio B, Redruello B, Fernandez M, etal. Q69 (an Enterococcus faecalis-Infecting Bacteriophage) as a biocontrol agent for reducing tyramine in dairy products. Front Microbiol. 2016; 7: 445.
- Uchiyama J, Rashel M, Maeda Y, Takemura I, Sugihara S, Akechi K, etal. Isolation and characterization of a novel Enterococcus faecalis bacteriophage phiEF24C as a therapeutic candidate. FEMS Microbiol Lett. 2008; 278: 200–6.
- Horiuchi T, Sakka M, Hayashi A, Shimada T, Kimura T, Sakka K. Complete genome sequence of bacteriophage BC-611 specifically infecting Enterococcus faecalis strain NP-10011. J Virol. 2012; 86: 9538–9.
- Khalifa L, Brosh Y, Gelman D, Coppenhagen-Glazer S, Beyth S, Poradosu-Cohen R, etal. Targeting Enterococcus faecalis biofilms with phage therapy. Appl Environ Microbiol. 2015; 81: 2696–705.
- Khalifa L, Coppenhagen-Glazer S, Shlezinger M, Kott-Gutkowski M, Adini O, Beyth N, etal. Complete genome sequence of Enterococcus bacteriophage EFLK1. Genome Announc. 2015; 3: e01308–15.
- Parasion S, Kwiatek M, Mizak L, Gryko R, Bartoszcze M, Kocik J. Isolation and characterization of a novel bacteriophage phi4D lytic against Enterococcus faecalis strains. Curr Microbiol. 2012; 65: 284–9.
- Li X, Ding P, Han C, Fan H, Wang Y, Mi Z, etal. Genome analysis of Enterococcus faecalis bacteriophage IME-EF3 harboring a putative metallo-beta-lactamase gene. Virus Genes. 2014; 49: 145–51.
- Fard RM, Barton MD, Arthur JL, Heuzenroeder MW. Whole-genome sequencing and gene mapping of a newly isolated lytic enterococcal bacteriophage EFRM31. Arch Virol. 2010; 155: 1887–91.
- Yasmin A, Kenny JG, Shankar J, Darby AC, Hall N, Edwards C, etal. Comparative genomics and transduction potential of Enterococcus faecalis temperate bacteriophages. J Bacteriol. 2010; 192: 1122–30.
- Yoon BH, Chang HI. Genomic annotation for the temperate phage EFC-1, isolated from Enterococcus faecalis KBL101. Arch Virol. 2015; 160: 601–4.
- Stevens RH, Ektefaie MR, Fouts DE. The annotated complete DNA sequence of Enterococcus faecalis bacteriophage phiEf11 and its comparison with all available phage and predicted prophage genomes. FEMS Microbiol Lett. 2011; 317: 9–26.
- Rigvava S, Tchgkonia I, Jgenti D, Dvalidze T, Carpino J, Goderdzishvili M. Comparative analysis of the biological and physical properties of Enterococcus faecalis bacteriophage vB_EfaS_GEC-EfS_3 and Streptococcus mitis bacteriophage vB_SmM_GEC-SmitisM_2. Can J Microbiol. 2013; 59: 18–21.
- Young-Woo K, Hyo-Thl C. Isolation and molecular characterization of ΦFC1,a new temperate phage from Enterococcus faecalis . Mol Cell. 1994; 4: 155–8.
- Nigutova K, Styriak I, Javorsky P, Pristas P. Partial characterization of Enterococcus faecalis bacteriophage F4. Folia Microbiol (Praha). 2008; 53: 234–6.
- Wolanek GA, Loushine RJ, Weller RN, Kimbrough WF, Volkmann KR. In vitro bacterial penetration of endodontically treated teeth coronally sealed with a dentin bonding agent. J Endod. 2001; 27: 354–7.
- Chavez de Paz LE. Redefining the persistent infection in root canals: possible role of biofilm communities. J Endod. 2007; 33: 652–62.
- Saber Sel D, El-Hady SA. Development of an intracanal mature Enterococcus faecalis biofilm and its susceptibility to some antimicrobial intracanal medications; an in vitro study. Eur J Dent. 2012; 6: 43–50.
- Sime-Ngando T. Environmental bacteriophages: viruses of microbes in aquatic ecosystems. Front Microbiol. 2014; 5: 355.
- Shapiro OH, Kushmaro A. Bacteriophage ecology in environmental biotechnology processes. Curr Opin Biotechnol. 2011; 22: 449–55.