Abstract
One challenge in current Arctic ecological research is to understand and predict how wildlife may respond to increased frequencies of “extreme” weather events. Heavy rain-on-snow (ROS) is one such extreme phenomenon associated with winter warming that is not well studied but has potentially profound ecosystem effects through changes in snow-pack properties and ice formation. Here, we document how ice-locked pastures following substantial amounts of ROS forced coastal Svalbard reindeer (Rangifer tarandus platyrhynchus) to use marine habitat in late winter 2010. A thick coat of ground ice covered 98% of the lowland ranges, almost completely blocking access to terrestrial forage. Accordingly, a population census revealed that 13% of the total population (n=26 of 206 individuals) and 21% of one sub-population were feeding on washed-up kelp and seaweed on the sea-ice foot. Calves were overrepresented among the individuals that applied this foraging strategy, which probably represents a last attempt to avoid starvation under particularly severe foraging conditions. The study adds to the impression that extreme weather events such as heavy ROS and associated icing can trigger large changes in the realized foraging niche of Arctic herbivores.
Current and future climate trends in most of the Arctic include rising temperatures, changing precipitation patterns and more frequent extreme weather events (Hassol Citation2004). The ecological effects of extreme weather phenomena are potentially large yet poorly explored (Post et al. Citation2009). For instance, heavy rain-on-snow (ROS) events, which are expected to become more frequent across the Arctic (Rennert et al. Citation2009), may strongly alter the snow-pack properties and even generate thick ice layers on the frozen ground when followed by sub-zero temperatures (Putkonen & Roe Citation2003; Kohler & Aanes Citation2004; Grenfell & Putkonen Citation2008; Rennert et al. Citation2009; Bartsch et al. Citation2010; Hansen et al. Citation2010a). There are indications that such ground-icing may have negative implications across trophic levels, including effects on plants (Robinson et al. Citation1998; Bjerke Citation2011), microbiota (Coulson et al. Citation2000) and small (Kausrud et al. Citation2008; Gilg et al. Citation2009) and large herbivores (e.g., Kohler & Aanes Citation2004; Hansen et al. Citation2010a; Stien et al. Citation2010; Hansen et al. Citation2011).
Despite its circumpolar relevance, our empirical knowledge of the ecological responses to heavy ROS events is limited to a few “early warning” systems, particularly the Ny-Ålesund area on the coast of Svalbard (Rennert et al. Citation2009). Here, heavy ROS and ground-icing occur rather regularly and have been shown to strongly reduce the population growth rates of the local meta-population of wild Svalbard reindeer (Rangifer tarandus platyrhynchus; Kohler & Aanes Citation2004; Hansen et al. Citation2011). Although these animals have a remarkable ability to locate ice-free microhabitat beneath the snow-pack (Hansen et al. Citation2010a), ground-icing can generate large changes in their behaviour, including exploratory movements across natural barriers (Stien et al. Citation2010) and range expansion to steep mountainous habitat (Hansen et al. Citation2010a). The present study documents how reindeer facing particularly icy conditions may further expand their niche to include washed-up kelp and seaweed (hereafter collectively referred to as kelp), a food source that is only occasionally included as a subsidy in the diets of large terrestrial herbivores (Carlton & Hodder Citation2003).
Materials and methods
Study area and species
The study area comprises the peninsula of Brøggerhalvøya and the coastal plains of Sarsøyra and Kaffiøyra, both located south of the peninsula, on the north-western coast of Spitsbergen, the largest island in the archipelago of Svalbard (see, e.g., Hansen et al. Citation2010b). Except for a small research settlement on Brøggerhalvøya (Ny-Ålesund), there is low human activity and no hunting. The climate is oceanic and mild for this latitude (79°N), with average temperature of −11.1°C and average total precipitation of 229 mm during November–April 1979–2010. The plant cover is scarce and dominated by mosses, lichens, dwarf willow (Salix polaris), purple saxifrage (Saxifraga oppositifolia) and graminoids. The vegetation rarely becomes taller than about 3–5 cm (except some graminoids) and is mainly confined to low altitudes.
Reindeer had been extinct in the area for almost a century due to past hunting when 15 wild animals were transferred to Brøggerhalvøya from Adventdalen, a valley to the south, in 1978 (Aanes et al. Citation2000). The population irrupted and then crashed from ca. 360 to ca. 80 individuals during the extremely icy winter of 1994 (Kohler & Aanes Citation2004), when about 40 individuals migrated to Sarsøyra and established a new population (N.A. Øritsland, pers. comm.). The population at Kaffiøyra, further south, was established in about 1996–97.
Svalbard reindeer are more or less solitary, and in contrast to most other Rangifer, they are highly sedentary and do not undertake long-distance migrations. Movement between the study ranges is restricted by glaciers, steep mountains and open sea or thin sea ice, but spatiotemporal variation in foraging conditions may induce migrations (Hansen et al. Citation2010b; Stien et al. Citation2010). Svalbard reindeer also differ from most other wild ungulates in that they are not subject to significant inter-specific competition or, in our study area, to insect harassment. Neither are they subject to predation: only a handful of killings by polar bear (Ursus maritimus) have been reported (Derocher et al. Citation2000; Sandal Citation2009). The population dynamics are shaped mainly by variation in climate and food availability (Reimers Citation1977 Citation1983; Aanes et al. Citation2000; Solberg et al. Citation2001; Aanes et al. Citation2002; Aanes et al. Citation2003; Kohler & Aanes Citation2004; Hansen et al. Citation2007; Tyler et al. Citation2008; Hansen et al. Citation2011), particularly during winter. Individual behaviour and range use are closely related to the distribution of food plants, both during summer (Hansen, Herfindal et al. Citation2009) and winter (Hansen, Aanes et al. Citation2009).
Data collection and analyses
As part of an annual monitoring programme, we surveyed the three sub-populations in late winter (1–8 April) 2010 from two snowmobiles crossing the landscape. Because of the open landscape and calm and stationary reindeer behaviour (see Hansen, Aanes et al. Citation2009), the animals can be approached at rather short distances (typically 100–200 m in winter), and census errors are assumed to be small. Each separate sub-population was surveyed during one day, except for Brøggerhalvøya (two days).
Following standard procedures (see Hansen et al. Citation2010a), we performed snow profile transects distributed in a 900× 1800-m grid system below 120 m a.s.l., providing a total of 57 snow profiles that reached the ground or, if present, ground-ice. Profiles were not completed when the snow-pack was >100 cm deep, which occurred in one instance. We performed ground-ice measurements in 20 sites on Brøggerhalvøya, 22 sites on Sarsøyra and 15 sites on Kaffiøyra. Altitudes above 120 m a.s.l. were not examined as they are generally not accessible by snowmobile. Note that the ground-ice thickness was usually only measured down to 14–15 cm depth for logistical reasons, but this did not influence our estimate of central tendency (i.e., the median, see below). Analyses were performed in R for Windows versions 2.12.0 (R Development Core Team Citation2010).
Results
Winter 2010 was the fourth mildest (−8.9°C) and the fourth wettest ever recorded (326 mm precipitation), with several heavy ROS events occurring during November–January (). Ground-ice (a) was present in 98% (n=56 of 57) of the snow profiles (median thickness 11 cm), and 89% of the profiles had ≥5 cm ground-ice. Under these severe foraging conditions, the population survey revealed that 13% of the total reindeer meta-population () was feeding on washed-up kelp on the sea-ice foot (b, c). The proportion of kelp feeders during this population “snap-shot” was 0% on Brøggerhalvøya, 12% on Kaffiøyra and 21% on Sarsøyra, which provided the best access to this food source. The demographic composition among kelp feeders () differed significantly from the non-kelp feeders (Fisher's exact test: P<0.01), and the few calves alive at this point in winter were overrepresented (Fisher's exact test: P<0.01). The adult (>1 yr) male segment (χ2=0.066, df=1, P=0.797), adult female segment (χ2=2.494, df=1, P=0.114) and unknown sex and age segment (Fisher's exact test: P=1) were neither overrepresented nor underrepresented among the kelp feeders.
Fig. 1 The weather in Ny-Ålesund during winter 2010. Bars represent daily total precipitation (measured between 6:00 h day t − 1 and 6:00 h day t), solid line indicates fluctuations in daily average temperature (00:00–24:00 h day t). Arrows indicate when major precipitation events (i.e., >10 mm daily precipitation measured in day t) occurred at above-zero temperatures (measured in day t − 1). These occurred on 11–12 November (total 43 mm), 11 December (17 mm), and 17–19 January (total 55 mm). Source: the Norwegian Meteorological Institute.
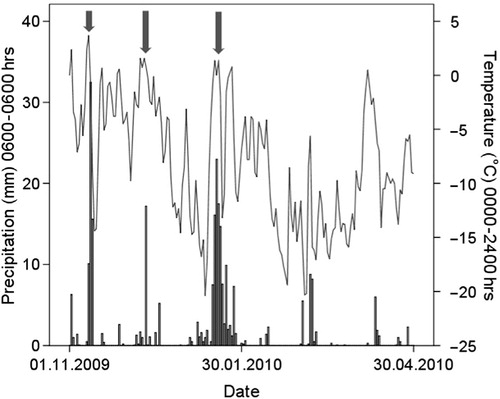
Fig. 2 (a) Ridge habitat coated with ground-ice on the coastal plain of Sarsøyra during April 2010. (b) Reindeer feeding on kelp on the beaches of Sarsøyra, with blocks of sea-ice in the background. (c) Reindeer feeding craters with kelp fragments at Sarsøyra. Photographer: B.B. Hansen (Norwegian Polar Institute).

Table 1 Distribution of kelp-feeders versus non-kelp feeders across populations and demographic groups in a Svalbard reindeer meta-population during a “snap-shot” (population survey) in late winter 2010. Total estimated population size was 206 animals.
Discussion
The present study has documented how parts of a coastal Svalbard reindeer meta-population used kelp as food during a winter with extremely poor foraging conditions due to heavy ROS and extensive ground-icing. To our knowledge, such use of non-terrestrial food has neither been demonstrated nor quantified previously in Svalbard reindeer. Likewise, non-anecdotal reports of other large terrestrial herbivores feeding on marine algae are few (Carlton & Hodder Citation2003) and largely limited to introduced reindeer (R. tarandus) on South Georgia (Leader-Williams et al. Citation1981) and red deer (Cervus elaphus) on the Isle of Rum (Conradt Citation2000), which occasionally fed on washed-up seaweed at low tide in winter.
Why should the reindeer use marine algae as food? Carlton & Hodder (2003) suggested that terrestrial mammals may utilize the intertidal zone as a seasonal food subsidy during resource-restricted periods of the year. Although quantitative data on kelp feeding are not available from other years, we have only observed this foraging strategy during winters when the accessibility of terrestrial forage has been particularly limited due to icing (R. Aanes, pers. obs.). The nutritional value of kelp and seaweed for terrestrial herbivores is largely unknown, but the in vitro digestibility of seaweed in the Isle of Rum system was comparable to terrestrial forage (Conradt Citation2000), and high mineral contents may possibly also have a significant nutritional value. On the other hand, roughly one fourth of the kelp feeders in our study apparently had diarrhoea (R. Aanes, pers. obs.). This was observed only occasionally among animals utilizing terrestrial foraging sites and could, therefore, be associated with, e.g., high salt intake from marine algae. The overrepresentation of calves among the kelp feeders further indicates that kelp is not a high-quality food source, as this demographic group has the poorest ability to compete for (or dig their way down to) the few ice-free terrestrial feeding sites.
Our results confirm that heavy ROS and icing can block access to winter forage (Hansen et al. Citation2010a) and thereby generate important changes in the realized foraging niche of Arctic herbivores. In the Ny-Ålesund area, icing has been shown to induce sudden range displacements (Stien et al. Citation2010) and force parts of the population to seek steep mountainous habitat (Hansen et al. Citation2010a) or alternative marine food sources (this study). These refuges are unlikely to provide resources sufficient to maintain the population in the long-term, and icing has been shown to strongly suppress the reindeer population growth rates (Kohler & Aanes Citation2004; Hansen et al. Citation2011). This was supported when, presumably because of the difficult winter conditions in 2009/10, the population underwent an 18% reduction in size from the summer of 2009 to the summer of 2010, despite initially very low animal densities (R. Aanes, unpubl. data). Accordingly, whether the reindeer can dig their way out of the expected future increase in heavy ROS and icing (Rennert et al. Citation2009) is highly uncertain (Hansen et al. Citation2011). The behavioural plasticity demonstrated in the rapidly warming Ny-Ålesund area may nonetheless serve as a bellwether of changes in range use and foraging niche expansion in Arctic herbivores facing an increase in extreme weather (such as heavy ROS) or climate change in general. Furthermore, the present study adds to the impression that warmer and wetter winters may reduce forage accessibility and have overall negative implications for herbivore populations across the Arctic (Forchhammer & Boertmann Citation1993; Aanes et al. Citation2000 Citation2003; Solberg et al. Citation2001; Miller & Gunn Citation2003; Kohler & Aanes Citation2004; Tews et al. Citation2007; Hansen et al. Citation2010a; Stien et al. Citation2010; Hansen et al. Citation2011).
Acknowledgements
The study was financed by the Norwegian Research Council (NORKLIMA grants to RA and BBH and an Arctic Field Grant to BBH), the Norwegian Polar Institute, and the Centre for Conservation Biology (co-funded through the Norwegian University of Science and Technology). A. Stien and V. Veiberg gave valuable comments on the manuscript.
References
- Aanes R. Sæther B.-E. Øritsland N.A. Fluctuations of an introduced population of Svalbard reindeer: the effects of density dependence and climatic variation. Ecography. 2000; 23: 437–443.
- Aanes R. Sæther B.-E. Smith F.M. Cooper E.J. Wookey P.A. Øritsland N.A. The Arctic Oscillation predicts effects of climate change in two trophic levels in a High-Arctic ecosystem. Ecology Letters. 2002; 5: 445–453.
- Aanes R. Sæther B.-E. Solberg E.J. Aanes S. Strand O. Øritsland N.A. Synchrony in Svalbard reindeer population dynamics. Canadian Journal of Zoology. 2003; 81: 103–110.
- Bartsch A. Kumpula T. Forbes B.C. Stammler F. Detection of snow surface thawing and refreezing in the Eurasian Arctic with QuikSCAT: implications for reindeer herding. Ecological Applications. 2010; 20: 2346–2358.
- Bjerke J.W. Winter climate change: ice encapsulation at mild subfreezing temperatures kills freeze-tolerant lichens. Environmental and Experimental Botany. 2011; 72: 404–408.
- Carlton J.T. Hodder J. Maritime mammals: terrestrial mammals as consumers in marine intertidal communities. Marine Ecology Progress Series. 2003; 256: 271–286.
- Conradt L. Use of a seaweed habitat by red deer (Cervus elaphus L.). Journal of Zoology. 2000; 250: 541–549.
- Coulson S.J. Leinaas H.P. Ims R.A. Soevik G. Experimental manipulation of the winter surface ice layer: the effects on a High Arctic soil microarthropod community. Ecography. 2000; 23: 299–306.
- Derocher A.E. Wiig O. Bangjord G. Predation of Svalbard reindeer by polar bears. Polar Biology. 2000; 23: 675–678.
- Forchhammer M.C. Boertmann D. The muskoxen Ovibos moschatus in north and northeast Greenland: population trends and the influence of abiotic parameters on population dynamics. Ecography. 1993; 16: 299–308.
- Gilg O. Sittler B. Hanski I. Climate change and cyclic predator–prey population dynamics in the High Arctic. Global Change Biology. 2009; 15: 2634–2652.
- Grenfell T.C. Putkonen J. A method for the detection of the severe rain-on-snow event on Banks Island, October 2003, using passive microwave remote sensing. Water Resources Research. 2008; 44: 1–9.
- Hansen B.B. Aanes R. Herfindal I. Kohler J. Sæther B.-E. Climate, icing, and wild arctic reindeer: past relationships and future prospects. Ecology. 2011; 92: 1917–1923.
- Hansen B.B. Aanes R. Herfindal I. Sæther B.-E. Henriksen S. Winter habitat-space use in a large Arctic herbivore facing contrasting forage abundance. Polar Biology. 2009; 32: 971–984.
- Hansen B.B. Aanes R. Sæther B.-E. Feeding-crater selection by High-Arctic reindeer facing ice-blocked pastures. Canadian Journal of Zoology. 2010a; 88: 170–177.
- Hansen B.B. Aanes R. Sæther B.-E. Partial seasonal migration in High-Arctic Svalbard reindeer (Rangifer tarandus platyrhynchus). Canadian Journal of Zoology. 2010b; 88: 1202–1209.
- Hansen B.B. Henriksen S. Aanes R. Sæther B.-E. Ungulate impact on vegetation in a two-level trophic system. Polar Biology. 2007; 30: 549–558.
- Hansen B.B. Herfindal I. Aanes R. Sæther B.-E. Henriksen S. Functional response in habitat selection and the tradeoffs between foraging niche components in a large herbivore. Oikos. 2009; 118: 859–872.
- Hassol S.J. Impacts of a warming Arctic. Arctic climate impact assessment. Cambridge University Press. Cambridge, 2004
- Kausrud K.L. Mysterud A. Steen H. Vik J.O. Østbye E. Cazelles B. Framstad E. Eikeset A.M. Mysterud I. Solhøy T. Stenseth N.C. Linking climate change to lemming cycles. Nature. 2008; 456: 93–97.
- Kohler J. Aanes R. Effect of winter snow and ground-icing on a Svalbard reindeer population: results of a simple snowpack model. Arctic Antarctic and Alpine Research. 2004; 36: 333–341.
- Leader-Williams N. Scott T.A. Pratt R.M. Forage selection by introduced reindeer on South Georgia, and its consequences for the flora. Journal of Applied Ecology. 1981; 18: 83–106.
- Miller F.L. Gunn A. Catastrophic die-off of Peary caribou on the western queen Elizabeth Islands, Canadian High Arctic. Arctic. 2003; 56: 381–390.
- Post E. Forchhammer M.C. Bret-Harte M.S. Callaghan T.V. Christensen T.R. Elberling B. Fox A.D. Gilg O. Hik D.S. Høye T.T. Ims R.A. Jeppesen E. Klein D.R. Madsen J. McGuire A.D. Rysgaard S. Schindler D.E. Stirling I. Tamstorf M.P. Tyler N.J.C. van der Wal R. Welker J. Wookey P.A. Schmidt N.M. Aastrup P. Ecological dynamics across the Arctic associated with recent climate change. Science. 2009; 325: 1355–1358.
- Putkonen J. Roe G. Rain-on-snow events impact soil temperatures and affect ungulate survival. Geophysical Research Letters. 2003; 30: 4.
- R Development Core Team. 2010. R: a language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
- Reimers E. Population-dynamics in two subpopulations of reindeer in Svalbard. Arctic and Alpine Research. 1977; 9: 369–381.
- Reimers E. Mortality in Svalbard reindeer. Holarctic Ecology. 1983; 6: 141–149.
- Rennert K.J. Roe G. Putkonen J. Bitz C.M. Soil thermal and ecological impacts of rain on snow events in the circumpolar Arctic. Journal of Climate. 2009; 22: 2302–2315.
- Robinson C.H. Wookey P.A. Lee J.A. Callaghan T.V. Press M.C. Plant community responses to simulated environmental change at a High Arctic polar semi-desert. Ecology. 1998; 79: 856–866.
- Sandal T. Blant rein og bjørn. (Among reindeer and bears.). Villreinen. 2009; 24: 68–69.
- Solberg E.J. Jordhøy P. Strand O. Aanes R. Loison A. Sæther B.-E. Linnell J.D.C. Effects of density-dependence and climate on the dynamics of a Svalbard reindeer population. Ecography. 2001; 24: 441–451.
- Stien A. Loe L.E. Mysterud A. Severinsen T. Kohler J. Langvatn R. Icing events trigger range displacement in a High-Arctic ungulate. Ecology. 2010; 91: 915–920.
- Tews J. Ferguson M.A.D. Fahrig L. Potential net effects of climate change on High Arctic Peary caribou: lessons from a spatially explicit simulation model. Ecological Modelling. 2007; 207: 85–98.
- Tyler N.J.C. Forchhammer M.C. Øritsland N.A. Nonlinear effects of climate and density in the dynamics of a fluctuating population of reindeer. Ecology. 2008; 89: 1675–1686.