
Abstract
Our study makes use of a fortuitous oceanographic data set collected around the remote sub-Antarctic island of Bouvetøya by a conductivity–temperature–depth recorder (CTD) integrated with a satellite-relayed data logger deployed on an adult female southern elephant seal (Mirounga leonina) to describe the seasonal evolution of the western shelf waters. The instrumented seal remained in waters over the shelf for 259 days, collecting an average of 2.6 (±0.06) CTD profiles per day, providing hydrographic data encompassing the late austral summer and the entire winter. These data document the thermal stratification of the upper water layer due to summer surface heating of the previous year's Antarctic Surface Water, giving way to a cold subsurface layer at about 100 m as the austral winter progressed, with a concomitant increase in salinity of the upper layer. Upper Circumpolar Deep Water was detected at a depth of approximately 200 m along the western shelf of Bouvetøya throughout the year. These oceanographic data represent the only seasonal time series for this region and the second such animal–instrument oceanographic time series in the sub-Antarctic domain of the Southern Ocean.
To access the supplementary material for this article, please see supplementary files under Article Tools, online.
In the SO, large-scale physical oceanography is now reasonably well characterized (Whitworth & Nowlin Citation1987; Pakhomov & McQuaid Citation1996; Abbott et al. Citation2000). In the open ocean, the densest water masses, the CDW and Antarctic Bottom Water are responsible, amongst other things, for the injection of nutrients into the upper water layers by upwelling (Prézelin et al. Citation2000; Law et al. Citation2003). Generally, the lack of phytoplankton abundance in the open SO, despite the presence of adequate nutrients, is attributed to a lack of dissolved iron (Fe) in the euphotic zone (Blain et al. Citation2007). This has led to the general characterization of the SO as a high nutrient, low chlorophyll environment. However, phytoplankton blooms have been observed near shelf waters around SO islands using satellite imagery (Korb et al. Citation2004; Mongin et al. Citation2008), suggesting that these regions may be subjected to localized Fe enrichment (Blain et al. Citation2001). While remote sensing methods can provide information on the productivity of surface waters, this information is only available in cloud-free conditions, and detection of subsurface or deep chlorophyll maxima indicates that phytoplankton blooms do occur outside the detectable (vertical) range of satellite imagery (Ardyna et al. Citation2013).
An informative method for characterizing shelf-water productivity is sampling for phytoplankton and measuring physical properties of the water column using electronic devices to detect deep chlorophyll maxima and quantify the water properties in which they occur. Such empirical studies of the shelf waters around Kerguelen Island in the sub-Antarctic identified sufficient winter enrichment of the surface mixed layer with Fe to promote one of the largest and most predictable phytoplankton blooms in the SO (Blain et al. Citation2007). Unfortunately, similar studies of the shelf waters of other SO islands are limited, primarily because of logistic and financial considerations. A lack of empirical studies on shelf oceanography hinders our ability to mechanistically describe the relationships between predator demographic trends and biological and physical oceanography. Yet understanding these relationships is critical if we are to predict how populations of centrally foraging predators will respond to changes in ocean conditions and processes due to climate modification (Trathan et al. Citation2007).
Abbreviations in this article
AASW: Antarctic Surface Water
CDW: Circumpolar Deep Water
CTD: conductivity–temperature–depth recorder
CTD-SRDL: CTD integrated into a satellite relay data logger
MEOP: Marine Mammals Exploring the Oceans Pole to Pole research programme
NADW: North Atlantic Deep Water
PSU: practical salinity units
SO: Southern Ocean
T-S: temperature–salinity
UCDW: upper layer of Circumpolar Deep Water
WW: Winter Water
The sub-Antarctic island Bouvetøya (a) in the South Atlantic sector of the SO provides suitable terrestrial breeding habitat for large populations of central place foraging marine predators, including macaroni penguins (Eudyptes chrysolophus), chinstrap penguins (Pygoscelis antarctica) and the second largest population of Antarctic fur seals (Arctocephalus gazella) in the world (Hofmeyr et al. Citation2005). This makes the region of great interest in terms of the physical processes in the marine environment that support such a predator guild. All of these species prey on krill (Euphausia superba; Hofmeyr et al. Citation2010; Rombolá et al. Citation2010; Trathan et al. Citation2012) and, to a lesser extent, myctophid fishes (Myctophidae spp.; Cherel et al. Citation2010; Staniland et al. Citation2011). On account of the remoteness of Bouvetøya, only three recent studies of the oceanography near the island have been conducted (Krafft et al. Citation2010; González-Dávila et al. Citation2011; Klunder et al. Citation2011). Constraints on these studies meant they were unable to provide anything more than point estimates of oceanographic conditions during the austral summer.
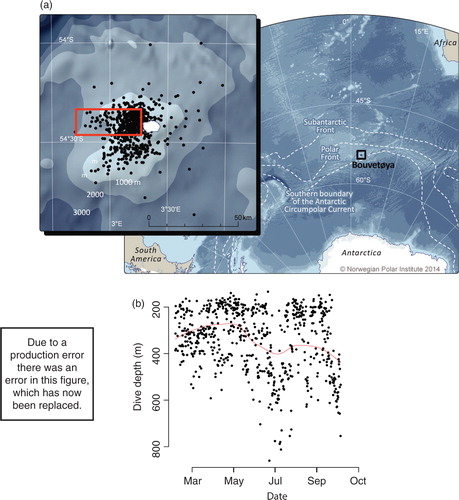
Fig. 1 (a) Filtered location estimates of CTD casts from a southern elephant seal instrumented on 20 January 2008 in relation to the sub-Antarctic island of Bouvetøya in the South Atlantic sector of the Southern Ocean. The red box highlights the section used to characterize the seasonal evolution of the water column in the areas typically used by resident krill–predator populations. Approximate locations of the sub-Antarctic Front, Polar Front and southern Boundary of the Antarctic Circumpolar Current are shown. (b) Diving during the resident period in the selected geographic box was consistent with a benthic foraging mode, with most dives being conducted within the 600-m isobath. The red line represents the fitted locally weighted scatter plot smoothed curve highlighting the temporal trend in dive behaviour.
In this study, an oceanographic “mooring” provided time-series data which afforded a unique opportunity to look at changes in the vertical structure of the shelf waters over time. This “mooring” was an adult female southern elephant seal (Mirounga leonina) instrumented with a CTD-SRDL. The seal remained in the immediate vicinity of Bouvetøya, providing a unique, continuous three-dimensional data set of the western shelf waters throughout the late austral summer until early October. This fortuitous data set allows us to characterize the composition of shelf waters and the seasonality of oceanographic processes around this rarely visited and consequently understudied island. We hypothesize that the physical oceanography on the shelf sets up conditions that can potentially support temporally and spatially predictable prey. Specifically, our study aims to (1) describe the three-dimensional seasonal evolution of the physical properties of the marine environment utilized by the predator guild during the austral summer and early autumn and (2) explore whether the UCDW, which is typically nutrient-rich, is present on the shelf over winter.
Materials and methods
This study was initiated at Nyrøysa, a beach on western Bouvetøya, in the Southern Ocean (54° 23’ S 3°47'E; a; Orsi et al. Citation1995). The oceanographic data reported herein are from a single adult female elephant seal tagged on 20 January 2008. The CTD-SRDL deployed on this seal was calibrated after assembly (Boehme et al. Citation2009) and CTD profiles collected were transmitted via the Argos satellite network. The CTD data collected were then post-processed (Boehme et al. Citation2009; Roquet et al. Citation2011), resulting in an estimated accuracy of ±0.03°C and 0.05 PSU. For broad-scale studies that seek to show relative changes over time, such as ours and that performed by Meredith et al. (Citation2011), these levels of accuracy and precision are quite reasonable, though readers must be aware that the data are not from traditional CTD platforms and interpret our findings accordingly.
To identify temporal trends in dive behaviour, a plot of the maximum depth of CTD profiles over time was fitted with a locally weighted scatter plot smoothed curve. We characterized the dive behaviour of the animal during the tracking period by plotting maximum dive depth against date. Error correction of Argos location data followed an iterative two-step process. Firstly, raw locations were pre-processed to remove extreme outliers by removing those with unclassified error estimates (LC-Z) and, secondly, a swim speed filter of 2 ms−1 was applied (McConnell et al. Citation1992). Subsequently, locations were estimated using a Kalman filter under a state-space framework in the R package “crawl” (Johnson et al. Citation2008). Applying this spatial error correction and filtering process, we estimated a location for each CTD profile that was likely accurate to within ±5 km of the true location (Kuhn et al. Citation2009; Lowther, Lydersen, Fedak et al. Citation2015).
We calculated seawater potential density anomalies (σ, kg m−3) at 1-m intervals for each CTD profile (McDougall et al. Citation2009). Monthly cross-sectional oceanographic profiles were generated for temperature (°C), salinity (PSU) and seawater potential density anomaly along a section running west from Nyrøysa out to a distance of 35 km, representing the approximate distance to the 2000-m isobath (a). Given the coarse nature of depth sampling provided by compressing T-S profiles, we define the surface mixed layer depth as a finite change in density from surface values (Nilsen & Falck Citation2006). To avoid aliasing issues with the diurnal heating–cooling cycle and to acknowledge the effect of surface turbulence, we use a conservative value of 0.125 kg m−3 and define the surface σ as the value calculated for the bottom of the first depth bin (4 m) to determine the surface mixed layer depth at the location of each CTD profile (Brainerd & Gregg Citation1995; Rudnick & Ferrari Citation1999; Huyer et al. Citation2005). To determine the surface mixed layer depth (m), monthly means were calculated and interpolated across the study area. A T-S plot of the hydrographic data was constructed using a reference pressure of 0 dbar to characterize the temporal evolution of the water column throughout the residency period of the elephant seal.
Results
The animal remained close to Bouvetøya from the time it entered the water post-tagging on 20 January until 5 October, during which time it commenced a directed south-east movement to approximately 57°S. Thus, for the purpose of this study, the seal's CTD-SRDL provided oceanographic data for 259 days, resulting in 675 CTD profiles collected at a rate of approximately 2.61±0.06 per day (a). Within a defined geographical section of interest (a), 424 CTD profiles (b) were collected during the 259-day deployment, resulting in an average 1.64 (±0.2) profiles per day. The mean depth of profiles was 337 m (±5 m), with a maximum depth of 860 m; of all oceanographic profiles, 31% were from water depths greater than 400 m. Most dives during the animal's sedentary period were conducted inside the 600-m isobath along the western shelf of Bouvetøya ().
There was a clear seasonal evolution of the water column along the western shelf of Bouvetøya (see Supplementary Fig. S1). During February and March, the upper 200 m of water were stratified, with a cold layer of water (0.25°C–0.75°C) between 80 and 200 m overlaid by surface waters reaching up to 1.5°C. The cold layer represents the remnants of the cold WW of the AASW from the previous year (Supplementary Fig. S1). The pronounced upper-layer thermocline persisted until the end of April, but as the austral winter progressed, rapid surface cooling led to the formation of a strong thermocline between 150 and 200 m, representing the new WW. Salinity also followed a seasonal progression with two pronounced haloclines separating the two upper summer layers (salinities approximately 33.8 and 34.4 PSU, respectively) and the more saline UCDW (ca. 34.8 PSU). These haloclines persisted throughout the rest of the study period, although an increase in salinity of the surface waters to ca. 34.2 PSU was apparent during the autumn and into winter. Concurrent with an increase in salinity through this period, the halocline also appeared to become deeper, reflecting the thickening of the upper layer of cold water during the winter.
A T-S plot showed the presence of a cold layer—WW AASW 2007—in mid-summer on top of the warmer UCDW of which North Atlantic Deep Water is a part (). By early autumn (April and May), T-S values covered a broader range, changing to near straight line T-S curves crossing a reduced number of isopycnals. During mid-winter (September and October), there was a cold (ca. −1.5°C) AASW layer above a denser layer of UCDW ( and Supplementary Fig. S1). The depth of the mixed layer varied spatially and temporally, ranging between 60 and 160 m to the west of the island, occurring shallower than 60 m across large regions between February and April (Supplementary Fig. S2).
Fig. 2 Potential T-S plots for the water column around the sub-Antarctic island of Bouvetøya, colour coded by month. Vertical dashed lines represent isopycnals and the horizontal dashed line approximates the freezing point of sea water at different salinities. Distinct water masses identified by potential temperature and salinity values are identified as WW, Antarctic Surface Water from the previous year (AASW 2007) and NADW.
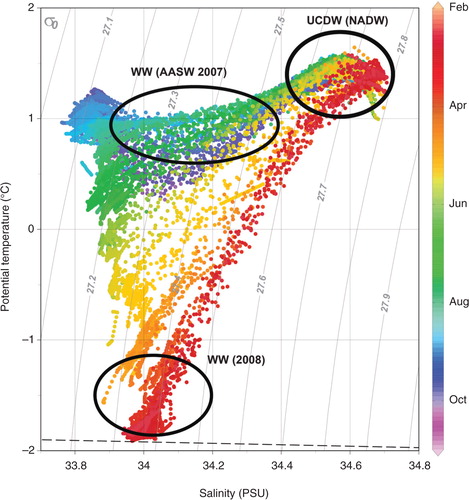
By the time the elephant seal departed the selected intensive study area, a stable, homogeneous layer of cold water (−1°C to −2°C) occupied the top 100 m, representing the fully developed WW, which lay above the upper layer of the CDW (Supplementary Fig. S1).
Discussion
In this study, the tagging of a deep-diving elephant seal that subsequently remained resident in the shelf waters around the remote sub-Antarctic island of Bouvetøya provided a unique data set that allowed the first description of the seasonal evolution of the water column in this region. Our study represents one of the two animal-borne instrument deployments that have collected a time series of ocean data comprehensive enough to quantify such changes, with the pattern of T-S plots in our study from 2008 tracking closely those describing the upper water column around the South Orkney Islands in 2007 (Meredith et al. Citation2011).
Two concurrent studies, independent of ours, detected elevated levels of dissolved nutrients, including Fe (0.5–1.0 nM), and heterogeneously distributed regions of high krill abundance throughout the upper water column and on the shelf to the west and south of Bouvetøya during the summer (Krafft et al. Citation2010; Klunder et al. Citation2011). The latter study documented that swarms of krill with high proportions of adults were found in waters with salinities and temperatures ranging from 33.96 (±0.05) to 34.19 (±0.09) PSU and 1.06 (±0.54)°C to 0.2 (±0.49)°C, respectively, with silicate levels five times the magnitude of those recorded when krill were absent. The shelf waters around Bouvetøya support a rich predator guild including chinstrap and macaroni penguins and Antarctic fur seals, colonies of which breed at Nyrøysa, amongst many other locations on the island. These predators feed on krill, which can aggregate in swarms that occupy the upper mixed layer during the night, descending to depths between100 and 250 m during daylight (Zhou & Dorland Citation2004). Penguins and Antarctic fur seals do not dive as deeply as elephant seals, so the consistent proximity of shallower regions of the mixed layer to the breeding beach plays an important role in their foraging ecology by providing access to krill (Lowther, Lydersen & Kovacs Citation2015). These resources are particularly crucial during the late breeding season for both fur seals and penguins when young require a lot of food that parents must bring back to the colony (Blanchet et al. Citation2013; Lowther et al. Citation2014).
In our study, we identified above ca. 200 m remnants of the previous year's cooler water AASW during mid-to-late summer, in agreement with the findings of a boat-based transect across the region conducted at the same time as our elephant seal was collecting ocean profiles in this area (González-Dávila et al. Citation2011). It is worth noting that AASW, derived from CDW upwelled on the continental shelf of Antarctica and advected north by Ekman transport, is rich in silicate (silicic acid), which is an essential nutrient for diatom productivity (Hutchins & Bruland Citation1998). Salinity and temperature profiles of the subsurface, remnant AASW we describe are closely matched with those described by Krafft et al. (Citation2010) as containing high levels of silicic acid, suggesting conditions in this western shelf water mass were favourable for krill feeding between February and March 2008.
In the meridional plane, upwelling and subsequent Ekman transport moves CDW northwards from the Antarctic continent where it meets the western shelf topography of Bouvetøya (Johnson & Bryden Citation1989). We detected the presence of UCDW on the western shelf of Bouvetøya throughout the study period, which was clearly identified by the presence of relatively warm, saline water below 200 m. Notably, our results depict the presence of nutrient-rich North Atlantic Deep Water within the UCDW which persists until early winter before being replaced exclusively by cooler UCDW.
Unlike ship-borne oceanographic studies, we are unable to influence the “design” of the sampling regime conducted by instrumented diving predators. The spatial variability in sampling effort is therefore likely biased towards areas where an individual can find food, with “unprofitable” areas being under-represented in our data set. However, given the sparse coverage of Argo floats in the area covered by our study, it is unlikely that a similar fine-scale four-dimensional characterization of the shelf hydrography of Bouvetøya could be made using only satellite-derived measurements. The opportunistic collection of time-series oceanographic data from a deep-diving marine predator has allowed us to characterize the seasonal evolution of the physical environment of a sub-Antarctic island that hosts large numbers of breeding central place foragers. Furthermore, these data have proven invaluable in garnering a clearer understanding of the links between physical oceanography and ecological processes (Lowther, Lydersen & Kovacs Citation2015), which would have been impossible using remote, satellite-derived measurements of near-surface ocean characteristics.
Supplementary Material
Download PDF (336.4 KB)Acknowledgements
This study is part of the International Polar Year MEOP (Norway) programme. Fieldwork was funded and carried out as part of the Norwegian Antarctic Research Expedition to Bouvetøya. The marine mammal data were collected and made freely available by the International MEOP Consortium and the national programmes that contribute to it (www.meop.net). We are greatly indebted to Martin Biuw who led the International Polar Year expedition and also to Greg Hofmeyr, Nico deBruyn, Petrus Kritzinger and Aline Arriola for their efforts during the 2007/08 fieldwork. We also acknowledge the comments from one anonymous reviewer and Fabien Roquet in refining this manuscript.
Notes
To access the supplementary material for this article, please see supplementary files under Article Tools, online.
References
- Abbott M.R., Richman J.G., Letelier R.M., Bartlett J.S. The spring bloom in the Antarctic Polar Frontal Zone as observed from a mesoscale array of bio-optical sensors. Deep-Sea Research Part II. 2000; 47: 3285–3314.
- Ardyna M., Babin M., Gosselin M., Devred E., Bélanger S., Matsuoka A., Tremblay J.-É. Parameterization of vertical chlorophyll a in the Arctic Ocean: impact of the subsurface chlorophyll maximum on regional, seasonal and annual primary production estimates. Biogeosciences Discussions. 2013; 10: 1345–1399.
- Blain S., Quéguiner B., Armand L., Belviso S., Bombled B., Bopp L., Bowie A., Brunet C., Brussaard C., Carlotti F. Effect of natural iron fertilization on carbon sequestration in the Southern Ocean. Nature. 2007; 446: 1070–1074.
- Blain S., Tréguer P., Belviso S., Bucciarelli E., Denis M., Desabre S., Fiala M., Jézéquel V.M., Le Fèvre J., Mayzaud P. A biogeochemical study of the island mass effect in the context of the iron hypothesis: Kerguelen Islands, Southern Ocean. Deep-Sea Research Part I. 2001; 48: 163–187.
- Blanchet M.-A., Biuw M., Hofmeyr G.J.G., de Bruyn P.J.N., Lydersen C., Kovacs K.M. At-sea behaviour of three krill predators breeding at Bouvetøya—Antarctic fur seals, macaroni penguins and chinstrap penguins. Marine Ecology Progress Series. 2013; 477: 285–302.
- Boehme L., Lovell P., Biuw M., Roquet F., Nicholson J., Thorpe S.E., Meredith M.P., Fedak M. Technical note: animal-borne CTD-satellite relay data loggers for real-time oceanographic data collection. Ocean Science. 2009; 5: 685–695.
- Brainerd K.E., Gregg M.C. Surface mixed and mixing layer depths. Deep-Sea Research Part I. 1995; 42: 1521–1543.
- Cherel Y., Fontaine C., Richard P., Labat J.-P. Isotopic niches and trophic levels of myctophid fishes and their predators in the Southern Ocean. Limnology and Oceanography. 2010; 55: 324–332.
- González-Dávila M., Santana-Casiano J.M., Fine R.A., Happell J., Delille B., Speich S. Carbonate system in the water masses of the southeast Atlantic sector of the Southern Ocean during February and March 2008. Biogeosciences. 2011; 8: 1401–1413.
- Hofmeyr G.J.G., Bester M.N., Kirkman S.P., Lydersen C., Kovacs K.M. Intraspecific differences in the diet of Antarctic fur seals at Nyrøysa, Bouvetøya. Polar Biology. 2010; 33: 1171–1178.
- Hofmeyr G.J.G., Krafft B.A., Kirkman S.P., Bester M.N., Lydersen C., Kovacs K.M. Population changes of Antarctic fur seals at Nyrøysa, Bouvetøya. Polar Biology. 2005; 28: 725–731.
- Hutchins D.A., Bruland K.W. Iron-limited diatom growth and Si: N uptake ratios in a coastal upwelling regime. Nature. 1998; 393: 561–564.
- Huyer A., Fleischbein J.H., Keister J., Kosro P.M., Perlin N., Smith R.L., Wheeler P.A. Two coastal upwelling domains in the northern California Current system. Journal of Marine Research. 2005; 63: 901–929.
- Johnson D.S., London J.M., Lea M.A., Durban J.W. Continuous-time correlated random walk model for animal telemetry data. Ecology. 2008; 89: 1208–1215.
- Johnson G.C., Bryden H.L. On the size of the Antarctic Circumpolar Current. Deep Sea Research Part A. 1989; 36: 39–53.
- Klunder M.B., Laan P., Middag R., De Baar H.J.W., Van Ooijen J.C. Dissolved iron in the Southern Ocean (Atlantic sector). Deep-Sea Research Part II. 2011; 58: 2678–2694.
- Korb R.E., Whitehouse M.J., Ward P. SeaWiFS in the southern ocean: spatial and temporal variability in phytoplankton biomass around South Georgia. Deep-Sea Research Part II. 2004; 51: 199–116.
- Krafft B.A., Melle W., Knutsen T., Bagøien E., Broms C., Ellertsen B., Siegel V. Distribution and demography of Antarctic krill in the southeast Atlantic sector of the Southern Ocean during the austral summer 2008. Polar Biology. 2010; 33: 957–968.
- Kuhn C.E., Johnson D.S., Ream R.R., Gelatt T.S. Advances in the tracking of marine species: using GPS locations to evaluate satellite track data and a continuous-time movement model. Marine Ecology Progress Series. 2009; 393: 97–109.
- Law C.S., Abraham E.R., Watson A.J., Liddicoat M.I. Vertical eddy diffusion and nutrient supply to the surface mixed layer of the Antarctic Circumpolar Current. Journal of Geophysical Research—Oceans. 2003; 108: article no. 3272, doi: http://dx.doi.org/10.1029/2002JC001604.
- Lowther A.D., Lydersen C., Biuw M., de Bruyn P.J.N., Hofmeyr G.J.G., Kovacs K.M. Post-breeding at-sea movements of three central-place foragers in relation to submesoscale fronts in the Southern Ocean around Bouvetøya. Antarctic Science. 2014; 6: 533–544.
- Lowther A.D., Lydersen C., Fedak M.A., Lovell P., Kovacs K.M. The Argos-CLS Kalman Filter: error structures and state-space modelling relative to Fastloc GPS data. PLoS One. 2015; 10: e0124754, doi: http://dx.doi.org/10.1371/journal.pone.0124754.
- Lowther A.D., Lydersen C., Kovacs K.M. A sum greater than its parts: merging multi-predator tracking studies to increase ecological understanding. Ecosphere. 2015; 6: article no. 251, doi: http://dx.doi.org/10.1890/ES15-00293.1.
- McConnell B.J., Chambers C., Nicholas K.S., Fedak M.A. Satellite tracking of grey seals (Halichoerus grypus). Journal of Zoology. 1992; 226: 271–282.
- McDougall T.J., Feistel R., Millero F.J., Jackett D.R., Wright D.G., King B.A., Marion G.M., Chen C., Spitzer P., Seitz S. The international thermodynamic equation of seawater—2010: calculation and use of thermodynamic properties. Manuals and guides 56. 2009; Paris: Intergovernmental Oceanographic Commission.
- Meredith M.P., Nicholls K.W., Renfrew I.A., Boehme L., Biuw M., Fedak M. Seasonal evolution of the upper-ocean adjacent to the South Orkney Islands, Southern Ocean: results from a “lazy biological mooring.”. Deep-Sea Research Part II. 2011; 58: 1569–1579.
- Mongin M., Molina E., Trull T.W. Seasonality and scale of the Kerguelen plateau phytoplankton bloom: a remote sensing and modeling analysis of the influence of natural iron fertilization in the Southern Ocean. Deep-Sea Research Part II. 2008; 55: 880–892.
- Nilsen J., Falck E. Variations of mixed layer properties in the Norwegian Sea for the period 1948–1999. Progress in Oceanography. 2006; 70: 58–90.
- Orsi A.H., Whitworth T., Nowlin W.D. On the meridional extent and fronts of the Antarctic Circumpolar Current. Deep-Sea Research Part I. 1995; 42: 641–673.
- Pakhomov E.A., McQuaid C.D. Distribution of surface plankton and seabirds across the Southern Ocean. Polar Biology. 1996; 16: 271–286.
- Prézelin B.B., Hofmann E.E., Mengelt C., Klinck J.M. The linkage between Upper Circumpolar Deep Water (UCDW) and phytoplankton assemblages on the west Antarctic Peninsula continental shelf. Journal of Marine Research. 2000; 58: 165–202.
- Rombolá E.F., Marschoff E., Coria N. Inter-annual variability in chinstrap penguin diet at South Shetland and South Orkneys islands. Polar Biology. 2010; 33: 799–806.
- Roquet F., Charrassin J.-B., Marchand S., Boehme L., Fedak M., Reverdin G., Guinet C. Delayed-mode calibration of hydrographic data obtained from animal-borne satellite relay data loggers. Journal of Atmospheric and Oceanic Technology. 2011; 28: 787–801.
- Rudnick D.L., Ferrari R. Compensation of horizontal temperature and salinity gradients in the ocean mixed layer. Science. 1999; 283: 526–529.
- Staniland I.J., Morton A., Robinson S.L., Malone D., Forcada J. Foraging behaviour in two Antarctic fur seal colonies with differing population recoveries. Marine Ecology Progress Series. 2011; 434: 183–196.
- Trathan P.N., Forcada J., Murphy E.J. Environmental forcing and Southern Ocean marine predator populations: effects of climate change and variability. Philosophical Transactions of the Royal Society B. 2007; 362: 2351–2365.
- Trathan P.N., Ratcliffe N., Masden E.A. Ecological drivers of change at South Georgia: the krill surplus, or climate variability. Ecography. 2012; 35: 983–993.
- Whitworth T., Nowlin W.D. Water masses and currents of the Southern Ocean at the Greenwich Meridian. Journal of Geophysical Research—Oceans. 1987; 92: 6462–6476.
- Zhou M., Dorland R.D. Aggregation and vertical migration behavior of Euphausia superba . Deep-Sea Research Part II. 2004; 51: 2119–2137.