Abstract
Atmospheric aerosol particles of biological origin are a very diverse group of biological materials and structures, including microorganisms, dispersal units, fragments and excretions of biological organisms. In recent years, the impact of biological aerosol particles on atmospheric processes has been studied with increasing intensity, and a wealth of new information and insights has been gained. This review outlines the current knowledge on major categories of primary biological aerosol particles (PBAP): bacteria and archaea, fungal spores and fragments, pollen, viruses, algae and cyanobacteria, biological crusts and lichens and others like plant or animal fragments and detritus. We give an overview of sampling methods and physical, chemical and biological techniques for PBAP analysis (cultivation, microscopy, DNA/RNA analysis, chemical tracers, optical and mass spectrometry, etc.). Moreover, we address and summarise the current understanding and open questions concerning the influence of PBAP on the atmosphere and climate, i.e. their optical properties and their ability to act as ice nuclei (IN) or cloud condensation nuclei (CCN). We suggest that the following research activities should be pursued in future studies of atmospheric biological aerosol particles: (1) develop efficient and reliable analytical techniques for the identification and quantification of PBAP; (2) apply advanced and standardised techniques to determine the abundance and diversity of PBAP and their seasonal variation at regional and global scales (atmospheric biogeography); (3) determine the emission rates, optical properties, IN and CCN activity of PBAP in field measurements and laboratory experiments; (4) use field and laboratory data to constrain numerical models of atmospheric transport, transformation and climate effects of PBAP.
Contents
Introduction2
History2
Definition and sources of primary biological aerosol particles3
Characteristic types of primary biological aerosol particles4
Bacteria and archaea4
Urban airborne bacteria5
Rural airborne bacteria9
Airborne bacteria at marine and coastal sites9
High altitude airborne bacteria10
Bacteria emission fluxes11
Archaea11
Fungal spores and fragments12
Pollen15
Viruses16
Algae and cyanobacteria18
Biological crusts and lichens18
Others19
Characteristic concentrations and emission estimates22
Techniques for PBAP collection and analysis22
PBAP sampling methods22
Traditional analysis methods23
Cultivation24
Light microscopy24
Fluorescence microscopy24
Modern analysis methods25
Molecular techniques25
Optical methods28
Non-optical methods30
Atmospheric relevance31
Atmospheric transport of biological particles31
PBAP as cloud condensation nuclei32
PBAP as ice nuclei32
Laboratory studies of ice nucleation active biological particles32
Observation of biological ice nuclei in the atmosphere33
Modeling of biological ice nuclei34
Optical properties39
Conclusions and outlook40
1. Introduction
1.1. History
The occurrence and relevance of airborne biological particles have been addressed since the beginnings of scientific investigations into atmospheric aerosols (Ehrenberg, Citation1847; Pasteur, Citation1861; Carnelly et al., Citation1887; De Bary, Citation1887). For example, by the late nineteenth century, Miquel (Citation1883) had already shown that airborne spore concentrations in France followed a seasonal cycle and were dependent on wind direction. He also showed that human mortality in Paris followed the bacteria concentration in the air. Since then, aerobiology (i.e. the study of airborne biological particles) has become well established as a multidisciplinary field of scientific research that interacts with a host of physical, biological and medical science disciplines (e.g. Gregory, Citation1973). The impact of aerobiology is especially notable in such diverse basic and applied sciences such as allergology, bioclimatology, palynology, biological pollution, biological warfare and terrorism, mycology, biodiversity studies, ecology, plant pathology, microbiology, indoor air quality, biological weathering, industrial aerobiology and cultural heritage.
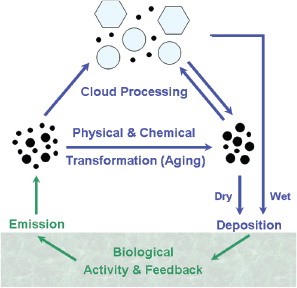
The potential relevance of biological particles for atmospheric processes has also been recognised for many years (e.g. Dingle, Citation1966; Schnell and Vali, 1972; Jaenicke and Matthias, Citation1988; Matthias-Maser and Jaenicke, Citation1995; Andreae and Crutzen, Citation1997; Jaenicke, Citation2005; Pöschl, Citation2005). outlines major processes in the cycling of primary biological aerosol particles (PBAP)Footnote1 between atmosphere and biosphere. After emission from the biosphere, PBAP undergo various physical and chemical aging processes in the atmosphere (coagulation, surface coating, reaction with photo-oxidants, etc.). Depending on their surface properties, PBAP can serve as nuclei for water droplets or ice crystals, leading to the formation of clouds and precipitation. Removal from the atmosphere proceeds via dry deposition (diffusion/sedimentation) or wet deposition with precipitation (nucleation/scavenging). After deposition, PBAP can interact with terrestrial or aquatic ecosystems and trigger biological activities leading to further PBAP emissions (growth and reproduction). However, biological particles in general have received less attention in atmospheric science than other types of aerosol particles such as sulfate, sea salt, mineral dust or volcanic ash (e.g. Junge, Citation1963; Hammond, Citation1971; Friedlander, Citation2000). This is primarily because the atmospheric impact of biological aerosols has been poorly understood, and because atmospherically relevant measurements have been costly and difficult to interpret. Also, global average number concentrations of biological particles have often been assumed to be insignificant compared to non-biological material and have thus not typically been considered for widespread measurements or included in global climate models. The Third Assessment Report (TAR) of the Intergovernmental Panel on Climate Change (IPCC) in 2001, for example, listed the global source strength of primary biological aerosol particles to be only 56 Tg/yr, in contrast to 3340 Tg/yr for sea salt and 2150 Tg/yr for mineral dust listed in the same report (Penner et al., Citation2001). Furthermore, the Fourth Assessment Report of the IPCC in 2007 stated that these estimates had not been refined, and primary biological particles were not mentioned in the contribution of Working Group I (Physical Science Basis) to the overall report (IPCC Citation2007b). PBAP concentrations have been estimated by other researchers (e.g. Matthias-Maser and Jaenicke, Citation1995) as comprising a much higher percentage of total atmospheric aerosol volume, however, and so important discrepancies exist.
Fig. 1. Cycling and effects of primary biological aerosol particles in the atmosphere and biosphere (adapted from Pöschl, Citation2005).
Interest in biological aerosol has been growing significantly in recent decades. This is highlighted by the fact that an Institute for Scientific Information (ISI) search for the term ‘biological aerosol’ (including quotation marks) results in less than a total of five publications until 1987, approximately one citation per year between 1987 and 1998, and then steadily increasing numbers with an average of ~9 citations per year between 1998 and 2011. Recently, several investigations have suggested that biological particles can have a substantial influence on clouds and precipitation and thus may influence the hydrological cycle and climate at least on regional scales (e.g. Andreae and Rosenfeld, Citation2008; Prenni et al., Citation2009; Pöschl et al., Citation2010). Various fields of medical research are also concerned with biological aerosols. Biological particles have been linked to many different adverse health effects spanning from infectious diseases to acute toxic effects, allergies, asthma and even cancer (Peccia et al., Citation2011). The negative effects that bioaerosols can play on the human respiratory system are particularly well documented (Verhoeff and Burge, Citation1997; Burge and Rogers, Citation2000; Douwes et al., Citation2003; Lee et al., Citation2005). Although medical research dealing with biological aerosols is indeed critical, this review article does not discuss medical applications of biological aerosols directly, providing instead a synthesis and overview of recent studies dealing with the observation and relevance of primary biological aerosol particles in an atmospheric context.
1.2. Definition and sources of primary biological aerosol particles
Aerosols are generally defined as colloidal systems of liquid or solid particles suspended in a gas (Hinds, Citation1999; Baron and Willeke, Citation2001; Fuzzi et al., Citation2006). Particle diameters are typically in the range of ~1 nm to around ~100 µm, where the lower limit is given by the size of small molecular clusters and the upper limit by high settling velocities comparable to the magnitude of atmospheric updraft velocities (~1 m s−1, Hinds, Citation1999; Seinfeld and Pandis, Citation2006). Primary atmospheric aerosol particles are emitted directly into the atmosphere from a source material, whereas secondary particles are formed in the atmosphere by condensation of gaseous precursors (Pöschl, Citation2005; Fuzzi et al., Citation2006).
The term ‘primary biological aerosol particles’ is defined to describe solid airborne particles derived from biological organisms, including microorganisms and fragments of biological materials such as plant debris and animal dander (IGAP, Citation1992)Footnote2. The definition for PBAP used within the text, as outlined in , can thus include all sorts of intact or fragmented biological cells, dispersal units or tissues.
Table 1. Characteristic types of primary biological aerosol particles (PBAP)
The term primary biological aerosol is more or less equivalent to the ‘soft’ term ‘bioaerosol’ (Reponen et al., Citation1995; Hinds, Citation1999). In contrast to the more rigorously defined ‘biological aerosol’, however, the term ‘bioaerosol’ is not very clearly defined and is frequently used with different meanings. In some cases, the term bioaerosol is used in a rather narrow sense, excluding biological secretion such as plant wax particles, for example (Gelencér, Citation2004). In other cases, it is used in a very broad sense, e.g. including any particle with biological activity/toxicity (Hirst, Citation1995), which would theoretically also comprise droplets of toxic chemicals such as sulphuric or nitric acid. Moreover, the term primary biological aerosol enables clear and easy distinction from biogenic secondary organic aerosols that are formed by atmospheric oxidation and gas-to-particle conversion of volatile organic compounds released from biological organisms (Hallquist et al., Citation2009; Jimenez et al., Citation2009). Therefore, we use the term biological aerosol within this review for the types of particles outlined in .
The size of PBAP can range from several nanometers (e.g. viruses, cell fragments) to a few hundred micrometres in aerodynamic diameter (e.g. pollen, plant debris; Cox and Wathes, Citation1995; Hinds, Citation1999; Jaenicke, Citation2005; Pöschl, Citation2005). Larger particles of biological material can also be lifted into the air, but due to high settling velocities they are rapidly deposited rather than being suspended over long times. Thus, they are usually not considered to be atmospheric aerosol particles, which is also the case for self-propelled organisms.
The biosphere, or the system in which all living things interact, dominates the Earth's surface, influencing the composition of land, water and air. Thus, PBAP can be released, both actively and passively, from every region of the globe. Key PBAP-producing systems include the following.
Plants release PBAP in the course of decay processes (see Section 2.7) as well as for reproduction, including pollen from higher plants and spores from ferns and mosses (see Section 2.3).
Microorganisms inhabit most plant, soil and rock surfaces (see Sections 2.1, 2.2, 2.5, 2.6). These microorganisms can be very numerous, contributing huge number concentrations per unit surface area (104 – 108 cells cm−2) in various natural environments (Morris and Kinkel, Citation2002; Lindow and Brandl, Citation2003; Yadav et al., Citation2004). Furthermore, the global leaf surface area is estimated to be roughly four times the terrestrial ground surface area (~6.4 108 km2 vs. ~1.5 108 km2) that provides a correspondingly large surface area for PBAP emission (Whittaker and Likens, Citation1973).
Primary biological aerosol particles originating from animals and humans include debris from skin or hair as well as, for example, excrements, brochosomes and eggs dispersed into the atmosphere by insects (see Section 2.7).
In areas of human activity, such as cities or agricultural managed areas, the numbers and composition of microorganisms such as bacteria or fungi are often increased and altered with respect to rural areas (see Sections 2.1.1, 2.2).
The cryosphere (e.g. Greenland, Antarctica, glaciers) is formed largely from precipitation, and this may be triggered by PBAP in some situations (Sands et al., Citation1982; Christner et al., Citation2008b; Pöschl et al., Citation2010). Bacteria have been discovered in ice cores from Antarctica at depths up to 3519 m (Raymond et al., Citation2008), giving possible evidence to the idea that these organisms have been introduced through precipitation. Thus, surface snow under windblown conditions could be a powerful source for PBAP via resuspension (Pomeroy and Jones, Citation1996).
Roughly, 70% of the globe is covered by oceans. They are full of living and decaying organisms, such as bacteria, archaea, fungi and algae, that are ejected from the ocean surface by bubble-bursting mechanisms, similar to the way other particles (e.g. sea salt) are emitted from such surfaces (Blanchard, Citation1983; O'Dowd et al., Citation2004).
In summary, the earth's biosphere provides many diverse and important sources of PBAP. We begin this review by discussing seven major categories of atmospherically relevant biological particles in detail. This is followed by a survey of available methods for PBAP detection and analysis, and overviews of the general atmospheric relevance of PBAP and their optical properties.
2. Characteristic types of primary biological aerosol particles
2.1. Bacteria and archaea
Due to their small size, bacteria have a relatively long atmospheric residence time (on the order of several days or more) compared to larger particles and can be transported over long distances (up to thousands of kilometres). Measurements show that mean concentrations in ambient air can be greater than 1×104 cells m−3 over land (Bauer et al., Citation2002a), whereas concentrations over the sea may be lower by a factor of ~100 – 1000 (Prospero et al., Citation2005; Griffin et al., Citation2006). Airborne bacteria may be suspended as individual cells but are more likely to be attached to other particles, such as soil or leaf fragments, or found as agglomerates of many bacterial cells (Bovallius et al., Citation1978; Lighthart, Citation1997). For this reason, whereas individual bacteria are typically on the order of ~1 µm or less in size, the median aerodynamic diameter of particles containing culturable bacteria at several continental sites has been reported to be ~4 µm, whereas at coastal sites it is ~2 µm (Shaffer and Lighthart, Citation1997; Tong and Lighthart, Citation1999; Wang et al., Citation2007).
The analysis of the diversity, composition and abundance of bacteria in air has experienced a recent growth in interest within the aerosol community (Morris et al., Citation2011). Understanding of the presence or properties of airborne bacteria is important because bacteria can influence atmospheric processes, function as human, plant and animal pathogens and be distributed over large physical scales from their natural or anthropogenic sources. In addition, the prediction of behavioural changes in bacterial colonisation of remote environments may be linked to climatic or anthropogenic changes that influence the atmosphere and thus could be a useful marker of changes in biodiversity.
Although in recent years, research on bacteria in the atmosphere has been constantly expanding, it remains difficult to establish a clear picture of the actual abundance and composition of bacteria in the air (Mancinelli and Shulls, Citation1978; Grinshpun and Clark, Citation2005; Maki et al., Citation2010). The presence of bacteria in air is strongly dependent on many factors such as seasonality, meteorological factors, anthropogenic influence, variability of bacterial sources and many other complicated variables. More importantly, the analysis of airborne bacteria still suffers from a lack of standardisation in air sampling and sample processing methods (Kuske, Citation2006; Peccia and Hernandez, Citation2006; Womack et al., Citation2010). Thus, differences in airborne concentration estimates, as well as in composition and abundance, could either be caused by biological variations or by differences in sampling or analysis strategies. Furthermore, many airborne bacteria studies were not designed to study a broad range of species but only to detect specific, and often pathogenic, species. Thus, current understanding of airborne bacteria concentrations and properties is undoubtedly influenced by such studies, and the literature should be understood with this in mind.
One area of expanding research is work towards the use of specific bacterial species for ‘source tracking’, conceptually similar to other atmospheric tracer methods. For example, the relative contribution of bacteria from various source environments can be determined, thus allowing measurements at an individual measurement site to predict the history of the air arriving at that location. Bowers et al. (Citation2010) made a contribution in this direction by identifying groups of bacteria in atmospheric samples, which are typically found primarily in the soil or on leaf-surface environments.
2.1.1. Urban airborne bacteria
The analysis and comparison of bacteria between urban and other environments has focused mostly on the comparison of bacterial concentration and not on diversity estimates. Historically, bacteria have typically been divided into Gram-positive and Gram-negative bacteria depending on their behaviour when their cell walls are treated by Gram staining. Culturing methods in general find more Gram-positive bacteria (e.g. Mancinelli and Shulls, Citation1978; Atlas and Bartha, Citation1997; Atlas and Bartha, Citation1997; Fuzzi et al., Citation1997; Kellogg and Griffin, Citation2006; Amato et al., Citation2007b; Fang et al., Citation2007; Fahlgren et al., Citation2010), whereas culture-independent techniques primarily find Gram-negative bacteria (e.g. Radosevich et al., Citation2002; Maron et al., Citation2005; Brodie et al., Citation2007; Després et al., Citation2007; Fierer et al., Citation2008; Fahlgren et al., Citation2010). Independent of the choice of detection method, several general trends can be observed from ambient measurements. A small number of existing studies suggest that both in urban and in natural areas, airborne bacterial communities are highly diverse, and variations in their species diversity are more complex than had previously been supposed (Bovallius et al., Citation1978; Jones and Cookson, Citation1983; Chihara and Someya, Citation1989). Bacterial diversity in rural areas is generally higher than at urban sites (Després et al., Citation2007). Still, the concentration of bacteria seems often to be higher in urban than in rural environments (Bovallius et al., Citation1978; Chihara and Someya, Citation1989; di Giorgio et al., 1996; Shaffer and Lighthart, Citation1997; Fang et al., Citation2007; Fahlgren et al., Citation2010), even when the samples are taken in the same geographic region. However, opposite trends have also been found: For example, Rosas et al. (Citation1993) described that bacterial concentrations in rural environments are higher than in urban sites. There is also evidence that the bacterial concentrations in the urban environment are influenced by human activities (Bovallius et al., Citation1978; Fang et al., Citation2007). Different cities do vary in their bacterial composition and abundance (Brodie et al., Citation2007); thus, there can be no single, typical description of urban bacterial composition.
A detailed description of the composition of bacteria found in urban air is restricted to only a few studies, as most culture-based studies classify cultured bacteria only as Gram positive or Gram negative, or as cocci or rods but do not provide taxonomic identification. Other studies dealing with the analysis of urban airborne bacteria give only the number of colony forming units (CFU) or other concentration estimates and do not try to identify the bacteria at all. Still, the general trend from available reports is that bacteria found in the air often belong to groups that are also common soil bacteria: on the taxonomic level of phyla, the Firmicutes (),Footnote3 Proteobacteria, Actinobacteria are often present, and on the level of superphyla the Cytophaga-Flavo-Bacteroidetes group is often being the most commonly observed. Additionally, in some studies bacteria from the phyla of Verrucomicrobia, Cyanobacteria, Acidobacteria, Planctomycetes and Chloroflexi have been detected. Within the airborne bacteria whose classes belong to Proteobacteria, Gamma- and Betaproteobacteria have been regularly identified, but Alpha-, Delta- and Epsilonproteobacteria have also been observed. Although Bacilli have been found often and in high numbers, drawing conclusions from these observations about their relative abundance is difficult due to high short-term variability and biases from the different detection methods.
Table 2. Taxonomic information on biological particles in air mentioned in the review
Concentrations of bacteria in cities exhibit especially high spatial variation because they are released from strong point sources, in contrast to the more spatially homogeneous release from, for example, an agricultural field. Areas with heavy vehicular traffic or sewage pollution have much higher concentrations of airborne bacteria, with a weaker or non-existent seasonal cycle, as compared to concentrations in more naturally influenced areas such as urban parks, forests or coastal sites (e.g. Miquel, 1883; Shaffer and Lighthart, Citation1997; Harrison et al., Citation2005; Fang et al., Citation2007).
The concentrations and composition of bacteria undergo daily, weekly and seasonal changes. It has often been found that numbers are greatest in summer and autumn (Bovallius et al., Citation1978; Jones and Cookson, Citation1983; di Giorgio et al., Citation1996; Tong and Lighthart, Citation1999; Fang et al., Citation2007; Kaarkainen et al., Citation2008), although exceptions exist; CitationFahlgren et al. (2010) found that bacteria counts at a coastal site were highest in winter, which they attributed to a strong winter marine sea spray source. Over a period of 1 d, bacteria have usually been observed to exhibit a peak airborne concentration in the morning and evening (Lighthart and Shaffer, Citation1995; Shaffer and Lighthart, Citation1997; Fang et al., Citation2007). It has been suggested by Maron et al. (Citation2006) that bacterial communities in cities show temporal variability, in which the daily and weekly variability is mainly influenced by anthropogenic sources, whereas seasonal variations are triggered by climate and atmospheric changes.
2.1.2. Rural airborne bacteria
Among the Gram-positive bacteria observed in rural air, Firmicutes and Actinobacteria are the prevalent groups, whereas Proteobacteria are the most prevalent Gram-negative bacteria (e.g. Lighthart, Citation1997; Maron et al., Citation2005; Després et al., Citation2007; Fang et al., Citation2007). Rural and urban sampling sites usually differ with regard to bacterial genetic diversity. Studies based on cultivation often find higher counts of CFUs at urban sites compared to rural sites in the same geographic regions (Bovallius et al., Citation1978; Jones and Cookson, Citation1983; Chihara and Someya, Citation1989; di Giorgio et al., 1996; Fang et al., Citation2007; Fahlgren et al., Citation2010). In contrast, some studies based on cultivation or deoxyribonucleic acid (DNA) analysis have detected higher diversity at rural than at urban sites (Rosas et al., Citation1993; Després et al., Citation2007). Some studies have shown strong correlations between bacteria concentrations at rural sites and meteorological conditions. For example, Lighthart et al. (Citation2009) found that six meteorological factors could account for 96% of the variance in culturable atmospheric bacteria concentrations measured in rural Oregon. Harrison et al. (Citation2005) measured boundary layer concentrations of total atmospheric bacteria at sites in England and found that they increased exponentially as a function of 24-h mean temperature and, except at the coastal site, decreased logarithmically with increasing wind speed, probably due to atmospheric dilution.
Shaffer and Lighthart (Citation1997) found that mean concentrations of culturable bacteria differed between four distinct land-use types chosen for study: urban, forest, rural and coastal. However, Bowers et al. (Citation2010) obtained an apparently contradictory result, finding concentrations to be 105 – 106 m−3 of air in each of three distinct land-use types (agricultural fields, suburban areas and forests). The difference could be related to the specific sites chosen, or could be methodological; whereas Shaffer and Lighthart (Citation1997) used cultivation to enumerate bacteria, Bowers et al. (Citation2010) used fluorescent microscopy to count DNA-containing particles in the size range 0.5 – 10 µm in diameter.
Despite the lack of difference in total bacteria numbers between the sites, Bowers et al. (Citation2010) found that concentrations of high-temperature ice nuclei, as determined by a droplet freezing assay, were on average two and eight times higher in the samples from agricultural areas than from the other two land-use types, which might indicate an agricultural, perhaps biological, source of ice nuclei, but the nature of the ice nuclei was not determined.
2.1.3. Airborne bacteria at marine and coastal sites
Although it has often been shown that airborne bacteria are dominated by bacterial groups that are prevalent in the soil, airborne bacteria, especially at marine and coastal sites, can also originate from marine sources. Bacteria are transmitted from water to the air by the bubble-bursting mechanism (e.g. Blanchard et al., Citation1981; Blanchard and Syzdek, Citation1982). Both, laboratory and field studies, have demonstrated that the concentration of bacteria in bubble bursting or sea spray aerosol greatly exceeds the concentration in the water from which the aerosol is produced (Blanchard and Syzdek, Citation1978; Blanchard et al., Citation1981; Blanchard and Syzdek, Citation1982; Blanchard, Citation1989; Marks et al., Citation2001; Aller et al., Citation2005). The mean atmospheric residence time of bacteria emitted from the oceans is expected to be shorter than for bacteria emitted from land surface due to their quicker removal by precipitation (Burrows et al., Citation2009b). In comparison with bacterial concentrations in urban and rural environments, CFU counts seem to be in general lower at coastal sites than at inland sites (Bovallius et al., Citation1978; Shaffer and Lighthart, Citation1997). Variation in bacterial concentrations over the course of a day can often be explained by considering onshore breezes that generally bring air with fewer bacteria than inland air (Lighthart, Citation1997).
Most of the studies dealing with the identification of airborne bacteria at marine and coastal sites have been conducted using culture-dependent techniques. However, it has also been shown in culture-independent analyses that bacteria at coastal and marine sites primarily stem from the phyla of the Proteobacteria, Firmicutes and Bacteroidetes. Within the Firmicutes, Bacillus seems to be prevalent (Shaffer and Lighthart, Citation1997), whereas within Proteobacteria mainly Alpha-, Beta- and Gammaproteobacteria were detected (Fahlgren et al., Citation2010; Urbano et al., Citation2011).
Although in general it has been reported that the concentration of bacteria is highest in summer and autumn, while lowest in winter (Vlodavets and Mats, Citation1958; Pady and Gregory, Citation1963; Borodulin et al., Citation2005), local exceptions can be found to this pattern. In a study analysing the airborne bacterial community at a sampling site near the Baltic Sea with mainly marine influenced air, Fahlgren et al., (Citation2010) detected higher CFU values during the winter compared with the summer. Because marine bacteria are ejected into the air along with sea spray aerosol particles, the source of marine bacteria to the atmosphere increases when winds become more powerful, generating more waves and surf, and thus sea spray. As a result, it is likely that marine bacteria are transferred to the atmosphere far more effectively by stronger winter winds (Nilsson et al., Citation2001; Nilsson et al., Citation2007; Fahlgren et al., Citation2010).
Examination of single particles collected in the air above biologically active ocean areas (Arctic and Southern) shows that bacteria are present, but their numbers are dwarfed by the large number of particles consisting of biogenic organic aggregates and colloids (Leck and Bigg, Citation2005a, Citation2005b; Bigg, Citation2007; Bigg and Leck, Citation2008).
Methodological issues may have confounded previous measurements of bacteria concentrations in marine air using culture methods. The culturability of bacteria in seawater is estimated to be between 0.001 and 0.1%, compared to 0.25% for freshwater and 0.3% for soil (Colwell, Citation2000). However, Fahlgren et al. (Citation2010) presented evidence that a majority of live airborne marine bacteria collected on the Swedish coast may be culturable on Zobell agar plates, which are based on Baltic seawater. Also, marine bacteria are smaller than land bacteria, with biovolumes often in the range 0.036 – 0.073 µm3 (Lee and Fuhrman, Citation1987), corresponding to equivalent spherical diameters of 0.20 – 0.26 µm. Consistent with the smaller size of marine bacteria, the count median diameter of particles associated with culturable bacteria has been found to be smaller at coastal sites – about 2 µm – compared to about 4 µm at continental sites (Shaffer and Lighthart, Citation1997; Tong and Lighthart, Citation2000; Wang et al., Citation2007).
2.1.4. High altitude airborne bacteria
The presence of bacteria at high altitudes in the atmosphere had already been detected as early as 1861 (Pasteur, 1861). Metabolically active microbes have been detected in air as high as 20–70 km in elevation (Imshenetsky et al., Citation1978; Griffin, Citation2004; Wainwright et al., Citation2004a; Bowers et al., Citation2009; Womack et al., Citation2010). Bacteria have been shown to survive long distance transport and also to be able to live and reproduce in airborne particles (Dimmick et al., Citation1979). Because bacteria metabolise within cloud droplets, some authors have proposed an impact on the chemistry of cloud droplets and air (Amato et al., Citation2005, 2007a, 2007c; Deguillaume et al., Citation2008; Vaitilingom et al., Citation2010). Vaitilingom et al. (Citation2010) showed that biodegradation is more likely to contribute to cloud chemistry at night than during the day because it must compete with photochemistry during the day. Bacteria collected in cloud water samples have been shown to metabolise and reproduce when those samples are incubated in the laboratory, even at supercooled conditions (Sattler et al., Citation2001; Amato et al., Citation2007a; Amato et al., Citation2007c). Sattler et al. (Citation2001) found that generation times varied between 3.6 and 19.5 d, comparable to those of phytoplankton in the ocean. The mean atmospheric residence time of a bacterial cell can be up to about 1 week (Burrows et al., Citation2009b), but the cell will spend only a small fraction of this time inside of a cloud droplet. Lelieveld and Heintzenberg (Citation1992) estimated that on average, tropospheric air spends about 5–6% of its time in clouds. It is, thus, unlikely that there is a significant primary production of bacteria within cloud droplets.
In addition to the effect of low temperature at high altitudes, it has been discussed repeatedly that air in general is a hostile environment for microorganisms, as they are exposed to UV light, have only small amounts of water available are subject to changing oxygen partial pressure, etc. (e.g. Womack et al., Citation2010 and references therein). Bacteria found at high altitudes probably either originate from high altitude habitats such as alpine sites, or they were transported to high altitudes with air currents. Rapid upwards transport of bacteria can be the result of storm activity over land and seas, volcanic activity, impact events and human activity such as weapons testing, aviation and spacecraft launches (Hall and Bruch, Citation1965; Bucker and Horneck, Citation1969; Simkin and Siebert, Citation1994; Kring, Citation2000; Griffin et al., Citation2002; Griffin, Citation2004). One evolutionary strategy for bacteria to survive in such hostile environments is the use of pigments to protect cells from harmful UV radiation. Thus, some studies on the diversity of bacteria in high altitude concentrate on pigmented bacteria (González-Toril et al., Citation2009).
The number of aerosol particles and PBAP decreases at high altitudes in general, and several studies could show that the diversity of airborne high-alpine bacteria is also reduced in comparison to urban and rural sites (Després et al., Citation2007; Bowers et al., Citation2009). These studies as well as others find primarily bacteria belonging to Proteo- and Actinobacteria as well as Bacteroidetes (González-Toril et al., Citation2009). Within the Proteobacteria, Gamma- and Betaproteobacteria are the prevalent classes detected. The presence of Bacillus species has also been reported from high altitude sites (Griffin, Citation2004; Maki et al., Citation2010). Between the different environmental types, bacteria phyla mainly differ in their relative proportions. Additional differences probably exist also on the species and family level.
2.1.5. Bacteria emission fluxes
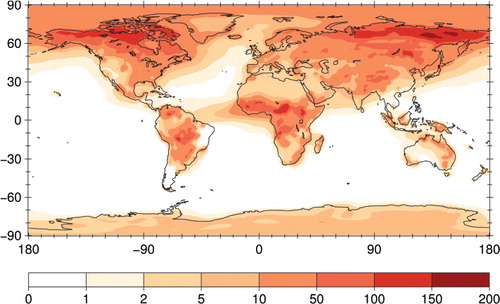
Only a few studies have attempted to directly measure the surface-atmosphere flux (net rate of emission and deposition) of bacteria. Flux measurements can be made using micrometeorological methods that rely on measuring the vertical gradient of bacteria concentrations in conjunction with gradients of air velocity, temperature or other parameters. These methods require fast and very short measurements for statistical significance and can be difficult to design, carry out and interpret. Existing estimates of the flux of culturable bacteria to the atmosphere range from 4.7 CFU m−2 s−1 for a high desert chaparral in Oregon, USA (Lighthart and Shaffer, Citation1994) to as much as 543 CFU m−2 s−1 for undisturbed croplands (Lindemann et al., Citation1982) and much stronger emissions for surfaces disturbed by human behaviour. For example, during harvesting of crops, emissions may be as high as 109 CFU m−2 s−1 (Lighthart, Citation1984). A first global model study estimates that average emissions from land of 250 m−2 s−1 (range: 140–380 m−2 s−1) would be required to reproduce observed mean concentrations of bacteria in the air (Burrows et al., Citation2009b; ), note that this estimate refers to total as opposed to culturable bacteria. Major obstacles to successful flux measurements include low number concentrations of airborne bacteria and the lack of automated methods for measuring concentrations because concentration measurements must be made continuously and at high time resolution.
Fig. 2. Column density of bacterial tracer (106 m−2), simulated from estimated emissions for a set of ten ecosystems estimates (Burrows et al., Citation2009b).
2.1.6. Archaea
All known life on earth can be categorised into one of three broad categories: archaea, bacteria and eukarya. As with bacteria, the DNA of archaea is not contained within a cell nucleus (prokaryotes). Little is known about archaea in the atmosphere. They had long been thought to occur only in very restricted, extreme environments, but by now they have been found in a wide variety of habitats (Schleper et al., Citation 2005). They are one of the most diverse and widespread forms of life on Earth and as major players in the biogeochemical cycles of nitrogen and carbon they are involved in the production of methane, the assimilation of amino acids and the oxidation of ammonium (Schleper et al., Citation2005). Archaea in the atmosphere have thus far been difficult to detect and characterise. Many studies have reported the detection of archaea from samples collected from air above compost piles and biosolids (e.g. Baertsch et al., Citation2007; Moletta et al., Citation2007; Thummes et al., Citation2007), but these are specialised environments rich in biological material. Three aerosol studies (Després et al., Citation2007; Fierer et al., Citation2008; Bowers et al., Citation2009) utilised polymerase chain reaction (PCR) primers capable of amplifying archaeal DNA, but were unable to amplify and detect genetic material from archaea. This may be due to limitations of the applied PCR primers and amplification conditions as described in Section 3.3.1.3. Radosevich et al. (Citation2002) is the only published report to successfully detect DNA sequences of three crenarchaeal clones from ambient air but no further information is provided.
2.2. Fungal spores and fragments
Fungal spores and fragments are understood to be one of the most common classes of airborne PBAP in a number of environments (Womiloju et al., Citation2003; Elbert et al., Citation2007; Bauer et al., Citation2008b; Crawford et al., Citation2009). Fungi are comprised of vegetative mycelia that consist of a large number of branched hyphae and grow in virtually every ecosystem on Earth and are also capable of efficient aerosolisation (Adhikari et al., Citation2009). Spores are often released by active processes such as osmotic pressure ‘cannons’ and surface-tension catapults (e.g. Buller's Drop; Ingold, Citation1971; Lacey, Citation1996; Ingold, Citation1999; Pringle et al., Citation2005). Spores can be released as a part of the sexual and/or asexual morph (stage) of the lifecycle of a fungus, and many species are able to produce spores from both stages. Fungi that actively release spores during the sexual morph, or telemorph, can typically produce one of three different kinds of spores: ascospores are released from an ascus (a long tube that typically holds eight spores), basidiospores from a basidium (small pedestal on fruiting bodies) or teliospores. The asexual stage, anamorph, of some fungi produce conidia, also known as conidiospores, that are produced by hyphal portions called conidiophores. Many studies thus refer to spores as a singular class, referring to both sexual and asexual morphs together. Spores are also often subdivided for practical reasons into hyaline (colorless) and dematiaceous (colored) spores (e.g. Pady and Gregory, Citation1963; Adams et al., Citation1968). Spores most commonly observed to dominate ambient concentrations of airborne fungal material have been from the species: Cladosporium, Alternaria, Penicillium, Aspergillus, as well as Epicoccum and a variety of yeasts, smuts and rusts (plant pathogens) and other basidiomycetes (e.g. Madelin, Citation1994). Detailed reviews on fungal spores in the atmosphere are available elsewhere (e.g. Madelin, Citation1994; Elbert et al., Citation2007) and as an example presents an example of fungal spore micrographs with and without coating by secondary organic aerosol. Some plants such as ferns and mosses also disperse spores that are typically larger and less numerous than fungal spores (Graham et al., Citation2003; Elbert et al., Citation2007).
Fig. 3. Fungal spores with and without coating by secondary organic aerosol (dark gray envelope in left panel). Electron micrographs of aerosol filter samples from pristine tropical rainforest air in the Amazon (CitationPöschl et al., 2010), (Reproduced with permission from AAAS).

In recent PBAP analyses, molecular genetic methods have been used that analyse the genetic substance, DNA, of biological material. As discussed by Després et al. (Citation2007), fungal DNA detected in aerosol filter samples is most likely to originate from spores that are known to resist environmental stress and survive atmospheric transport (Madelin, Citation1994; Griffin, Citation2004; Griffin and Kellogg, Citation2004). But, fungal DNA may also be derived from other fungal material such as hyphae and tissue fragments. Although it is generally assumed that spores comprise the majority of airborne fungal material, this remains largely unsubstantiated and has important implications for interpretation of both, laboratory or field measurements. Hyphal fragments have been observed in ambient air in a number of studies (Gorny et al., Citation2002; Green et al., Citation2006). Sinha and Kramer (Citation1971) suggested that airborne hyphae are most commonly unbranched conidiophores that can be 1–100 µm in length but that are more commonly 5–40 µm. Pady and Gregory (Citation1963) summarise the observed concentrations to be within the range 100–103 m−3 and observed that a large fraction of ambient hyphae were viable and able to germinate resulting in fungal colony growth. As most fungal species in the biosphere are still unknown, the detection and characterisation of fungi in atmospheric aerosol samples by DNA analysis can help to elucidate the global spread and diversity of fungi. As a by-product of the active emission process, fungal spores can be coated with specific sugar (e.g. arabitol, mannitol) and sterol (e.g. ergosterol) compounds that have thus been utilised as chemical tracers for ambient fungal spore concentrations (Lau et al., Citation2006; Bauer et al., 2007). Spores can also be coated with hydrophobin compounds that may affect both, their ice-nucleating ability (Iannone et al., Citation2011) and the immune response they cause after human inhalation (Aimanianda et al., Citation2009).
Depending on biological species, age and ambient conditions, the diameter of fungal spores can vary (~1–50 µm); most frequently it is in the range of 2–10 µm (Elbert et al., Citation2007; Wang et al., Citation2008; Fröhlich-Nowoisky et al., Citation2009; Huffman et al., Citation2010). Furthermore, fungal spores are often observed to aggregate into long chains of spores that greatly affect their aerodynamic diameter and have implications for both, atmospheric lifetime and deposition into human tissues (Lacey, Citation1991; Reponen et al., Citation2001).
The number and mass concentrations of fungal spores are typically observed to be ~104 m−3 and ~1 µg m−3, respectively, in continental boundary layer air ( and ). They account for up to ~10% of organic carbon (OC) and ~5% of PM10 at urban and suburban locations (Bauer et al., Citation2002a Citation2000b, 2008a). In pristine tropical rainforest air, fungal spores account for up to ~45% of coarse particulate matter. Thus, the properties and effects of fungal spores may be particularly important in tropical regions where both, physico-chemical processes in the atmosphere and biological activity at the Earth's surface are particularly intense (Graham et al., Citation2003; Gilbert, Citation2005; Elbert et al., Citation2007; CitationPöschl et al., 2010; Zhang et al., Citation2010), but chemical tracers typical of fungal spores have also been reported in aerosols from semi-arid and arid sites (Graham et al., Citation2004).
Table 3. Global emission estimates for different types of PBAP and size ranges of air particulate matter (PMx, x = upper limit of particle diameter; TSP = total suspended particulates)
Table 4. Characteristic magnitudes of the number and mass concentrations of PBAP in air over vegetated regions
Most airborne fungi belong to the divisions of Ascomycota and Basidiomycota (Fröhlich-Nowoisky et al., Citation2009). Most Ascomycota and Basidiomycota actively eject their spores with liquid jets or droplets (osmotic pressure and surface tension effects), whereas others rely on dry spore detachment by wind or other external forces. Dry-discharged spore concentrations tend to be enhanced during warm, dry weather conditions, whereas actively wet discharged spores tend to be enhanced during humid conditions such as those at night and in the early morning hours (Graham et al., Citation2003; Elbert et al., Citation2007). Emission and dispersal of fungal spores can thus be selectively correlated with various meteorological parameters and usually have specific behaviours, depending on the species involved (Fitt et al., Citation1989; Pasanen et al., Citation1991; Calderon et al., Citation1995; Katial et al., Citation1997; Sabariego et al., Citation2000; Troutt and Levetin, Citation2001; Burch and Leventin, Citation2002; Jones and Harrison, Citation2004; Grinn-Gofron and Mika, Citation2008; Oliviera et al., Citation2009).
From spore counts and molecular tracers, Elbert et al. (Citation2007) derived a global emission rate of ~50 Tg a−1 for fungal spores, corresponding to average mass and number emission fluxes of ~23 ng m−2 s−1 and 200 m−2 s−1, respectively, over land. These values are in fair agreement with the global average model estimates of Heald and Spracklen (Citation2009): ~28 Tg a−1 or ~6 ng m−2 s−1 over land, and 189 Tg a−1 calculated by Jacobson and Streets (Citation2009) for the year 2000. For Europe, Winiwater et al. (2009) derived a value of ~0.6 ng m−2 s−1, and recently Sesartic and Dallafior (Citation2011) estimated average mass and number flux values of ~17 ng m−2 s−1 and 513 m−2 s−1 over land. Overall, the studies indicate that the global average emission rates of fungal spores are uncertain by about one order of magnitude, which appears comparable to many aerosol sources and less than for other types of PBAP (). More field data are required to constrain better the actual emission flux of fungal spores on regional and global scales.
Studies based on DNA obtained directly from atmospheric aerosol samples offer new possibilities to identify the origin of fungal matter, independent of viability, cultivability and fragmentation (e.g. Boreson et al., Citation2004; Peccia and Hernandez, Citation2006; Fierer et al., Citation2008; Bowers et al., Citation2009; Fröhlich-Nowoisky et al., Citation2009). DNA-based techniques can amplify target regions of the DNA extracted directly from atmospheric aerosol samples. Amplification of the internal transcribed spacer (ITS) regions between the 18S and 28S ribosomal ribonucleic acid (rRNA) genes provides good target regions to identify fungi to genus and often to species level (O'Brien et al., Citation2005; Fröhlich-Nowoisky et al., Citation2009). DNA sequencing of this region has proven to be an efficient tool for the detection of rare and hard-to-cultivate fungi (e.g. Blumeria graminis) as well as highly abundant and easy-to-cultivate fungi (e.g. Cladosporium sp.) in aerosol samples (Fröhlich-Nowoisky et al., Citation2009) and other habitats (e.g. Hunt et al., Citation2004; O'Brien et al., Citation2005).
With regard to species richness, Fröhlich-Nowoisky et al. (Citation2009) detected 64% Basidiomycota and 34% Ascomycota in a semi-urban environment in central Europe, whereas Bowers et al. (Citation2009) found only 4% Basidiomycota but 82–92% Ascomycota at a mountain site in North America. On the class level within the Ascomycota, Dothideomycetes and Eurotiomycetes seem to be the prevalent groups (Bowers et al., Citation2009; Fröhlich-Nowoisky et al., Citation2009). These findings may be influenced not only by regional differences but also by measurement issues, and the actual biogeographic distribution of airborne fungal species is a subject of ongoing studies (Womack et al., Citation2010; Fröhlich-Nowoisky et al., Citation2011). As in bacteria, seasonal variation exists and differs between sampling sites also for fungi. During hot summer seasons, a decrease in the airborne microflora was reported for some fungi (Mullius et al., Citation1984; Filipello-Marchision et al., Citation1992; Fröhlich-Nowoisky et al., Citation2011).
The detection and apparent frequency of occurrence of different species can be affected by technical factors such as extraction efficiency of genomic DNA, varying rRNA gene copy number in the species, primer matching and performance, amplification efficiency of the target region, and cloning success. To our knowledge, from the few studies that have reported DNA analyses of fungi in atmospheric aerosol samples, some had neither found the expected high abundance of, e.g. Cladosporium sp. nor a high species richness (in particular Basidiomycota), which may well be due to limitations of the applied PCR primers (e.g. Després et al., Citation2007; Fierer et al., Citation2008; Bowers et al., Citation2009), although over 1500 fungal DNA sequences from 5 urban air samples were measured by Fierer et al. (Citation2008) and several dozens of filter samples of urban, rural and high-alpine air were analysed by Després et al. (Citation2007). Thus, careful selection and combination of multiple PCR primer pairs and other materials for the extraction and amplification of DNA obtained from aerosol samples are key elements for achieving high coverage of species richness (>300 species), as discussed in Fröhlich-Nowoisky et al. (Citation2009). The high number of fungal species that were detected only once indicates that a higher number of samples and clones would have to be investigated for a complete coverage of species diversity (Fröhlich-Nowoisky et al., Citation2009). In any case, the detected and reported numbers and frequencies of occurrence of species, families and classes can only be taken as a lower limit for the actual diversity and frequencies of occurrence.
2.3. Pollen
Among PBAP types, pollen grains can be among the largest in physical size and represent the reproductive units of plants that contain the male gamete. Pollen grains vary in size between 10 and 100 µm, have various shapes and have a hard shell that protects the sperm cells during the transportation processes. Pollen grains occur as biological aerosols not only as complete units but also as fragmented pieces. Pollen can rupture when the humidity is high, and these fragments have been shown to be in the range from 30 nm to 5 µm (Taylor et al., Citation2002, Citation2004; Miguel et al., Citation2006). Pollen of anemophilous plants use wind as their dispersal vector and have a typical diameter of 17–58 µm (Stanley and Linskins, Citation1974; Kuparinen, Citation2006; Nathan et al., Citation2008; Pope, Citation2010). They are usually dispersed in large amounts and over wide ranges because of their floating ability (Straka, Citation1975). The pollen is often ejected in clumps that stick to their neighbouring vegetation and are blown away after drying (Jones and Harrison, Citation2004).
The ability of pollen to disperse into the atmosphere depends on several parameters. On the one hand, dispersal depends on resuspension as described in detail by Jones and Harrison (Citation2004). On the other hand, pollen dispersal depends on meteorological factors. For example, bonding of pollen to surfaces is affected by temperature and moisture and thus by temperature and humidity of the air (Jones and Harrison, Citation2004). It has also been shown that pollen counts in, for example, the cypress family show significant positive correlations with daily minimum, mean and maximum temperatures and negative correlations with precipitation (Lo and Levetin, Citation2007). After temperature and humidity, wind and rain are typically the most important parameters for pollen dispersal, and it has been shown that pollen emission is reduced in the presence of rain or when the wind speed was low (Ogden et al., Citation1969). In general, the pollen concentration decreases with height, and for birch it could be shown that at 2000 m just 40% of the ground concentration were present (Rempe, Citation1937). This general phenomenon can be modified by inversion layers (Linskens and Jorge, Citation1986). The horizontal distance over which pollen can be carried with the wind can depend on prevailing temperatures, and it has been suggested that an increase in air temperature may induce atmospheric instability and thus promote pollen dispersal (Kuparinen et al., Citation2009). However, pollen has been observed to be lifted into the upper layers of the atmosphere by convection (Monin and Obukhov, Citation1954; Tackenberg, Citation2003; Taylor and Jonsson, Citation2004; Wright et al., Citation2008), and this ability makes pollen grains possibly relevant as ice nuclei in many environments.
The residence time of pollen in the atmosphere depends on their settling velocities that depend on morphology (shape), density and size of the pollen and vary widely among pollen types (Digiovanni et al., Citation1995; Diehl et al., Citation2001). The time pollen stay aloft in the atmosphere also influences the horizontal distance they can travel. Long distance dispersal (LDD) is not only interesting from the atmospheric point of view but also shapes many fundamental processes in plant ecology and evolution (Ellstrand, Citation1992; Kawecki and Ebert, Citation2004; Neilson et al., Citation2005; Nathan et al., Citation2008), e.g. the ability of plants to spread into new areas (IPCC Citation2007a). Dust is well known to frequently travel long distances, even across oceans (Kellogg and Griffin, Citation2006; Ben-Ami et al., Citation2010). Biological particles can also be transported over similar distances and have been observed to accompany dust plumes thousands of miles from their assumed sources (Shinn et al., Citation2000; Kellogg and Griffin, Citation2006; Polymenakou et al., Citation2008; Hallar et al., Citation2011). The LDD of wind-dispersed pollen is typically promoted by turbulent vertical fluctuations in wind and by coherency in vertical eddy motion that uplifts seeds well above the vegetation canopy (Nathan et al., Citation2002; Tackenberg, Citation2003; Soons et al., Citation2004). For a wide range of plant types, a positive relationship has been observed between mean air temperature and the frequency of LDD by wind in a boreal forest. Thus, an increase in local air temperature increases pollen dispersal distances (Kuparinen et al., Citation2009).
Another characteristic of pollen is their seasonality, as the presence of pollen in the atmosphere follows a clear seasonal cycle in response to the flowering seasons of the plant sources (Tormo et al., Citation2010). On a local scale, the pollination season for each plant is predictable and only shifts slightly as a function of meteorological parameters. The amount of pollen disposed by individual plants can vary greatly from 1 yr to another. Pollination during a given season always has a date, at which the pollen begins to disperse but at lower numbers, followed by the main pollination season when most of the pollen is dispersed, and conclude by a date when the plant stops its pollen-producing phase. However, usage of this nomenclature varies greatly within the literature and can be confusing (Jato et al., Citation2006). In addition, sedimented pollen might be resuspended again from dry surfaces. Pollen grains occasionally show a diurnal cycle with concentrations rising 1–2 h after dawn, peaking a few hours later and decreasing through the afternoon (Jones and Harrison, Citation2004). The phenomenon of diurnal variability is also common among other biological aerosol types such as bacteria and fungal spores (Jones and Harrison, Citation2004; Huffman et al., Citation2010).
Pollen grains are large PBA particles that typically lead to short atmospheric residence times. However, they can be up-drafted to high altitudes and have large residence times, and thus pollen can reach concentrations comparable to ice nuclei in some circumstances (Scheppegrell, Citation1924; Pruppacher and Klett, Citation1997; Diehl et al., Citation2001). Detailed information on the behaviour of pollen in ice nucleation is given in section 4.2.
Global changes, such as increasing atmospheric CO2 concentrations, increasing temperature, changes in the amount, distribution, and intensity of precipitation events, increases in the intensity and frequency of certain extreme weather events, changes in land use, and urbanisation, will likely have an impact on the production, distribution and dispersion of pollen (IPCC Citation2007b; Reid and Gamble, Citation2009). Climatic changes may lead to shifted or even elongated pollinations seasons. As pollen is known to cause allergies in humans, these shifts may also lead to changes in human exposure and changes in the prevalence and severity of symptoms in individuals with allergic diseases (Reid and Gamble, Citation2009). Alterations in the timing of aeroallergen production in response to weather variables have been clearly demonstrated for certain tree species, but less for grass, weed and mould (Katial et al., Citation1997; Emberlin et al., Citation2002; Clot, Citation2003).
Because pollen allergies can cause such medical problems, various pollen-monitoring programmes, networks and databases have been developed recently to provide data on pollen observation, share methods, encourage collaborations and thus create foundation for intensive pollen research. Examples include the Pollen Monitoring Program in Europe (Giesecke et al., Citation2010) and the Pollen Biology Research Coordination Network in the United States. The Global Pollen Database combines pollen information from Africa, the Americas and northern Asia made available by regional networks such as the Indo Pacific Database, the Latin America Pollen Database and the North American Pollen Database. These international collaborations are helping to advance knowledge of global pollen distribution.
In addition to climate-related changes in the atmospheric abundance of pollen and fungal spores, the allergenic potential of PBAP can also be enhanced by interactions with air pollutants (Taylor et al., Citation2002; Franze et al., Citation2003; Taylor and Jonsson, Citation2004; Franze et al., Citation2005; Pöschl, Citation2005; Reid and Gamble, Citation2009). For example, the reaction with ozone and nitrogen oxides leads to the formation of reactive oxygen intermediates and nitrated proteins that can influence the interaction of PBAP with the immune system and trigger or exacerbate allergic diseases (Gruijthuijsen et al., Citation2006; Shiraiwa et al., Citation2011; Zhang et al., Citation2011).
2.4. Viruses
Viruses are among the smallest of common PBAP classes, with physical diameter as low as 20 nm (Dongsheng, Citation2006). However, viruses are not commonly airborne as individuals and are more likely attached to other suspended particles (e.g. Yang et al., Citation2011).
Many diseases present in humans, animals, birds, fish, insects and plants are caused by viruses found in aerosols (one of possible routes of infection transmission), confirmed by numerous laboratory studies (Akers, Citation1969; Akers, Citation1973; Verreault et al., Citation2008). However, publications revealing the presence of viruses as PBAP are not numerous (Gloster et al., Citation1982; Christensen et al., Citation1990; Grant et al., Citation1994; Chen et al., Citation2008b). This perceived lack of research might be connected to the fact that before the development of molecular biological methods (e.g. PCR) to detect genetic material of microorganisms (Alvarez et al., Citation1995; Peccia and Hernandez, Citation2006), only viable viruses could be found in air samples. There are no universal test systems for virus detection such as nutrient media for bacteria or fungi. Instead, sensitive cell cultures, embryonated chicken eggs or susceptible laboratory animals are required (Zhdanov and Gaudamovich, Citation1982a; Zhdanov and Gaudamovich, Citation1982b), and specific test systems allow only the detection of viruses replicating in them. No other viable viruses, which can also be present in aerosol samples, will be detected.
Another possible reason for the scarcity of publications containing information on viable viruses in atmospheric aerosol is inactivation of viruses in the atmosphere under the influence of different environmental factors (changes in temperature, relative humidity, solar radiation, etc.). Unlike bacteria, fungi and algae, viruses have no repair systems, and therefore, their inactivation rates are usually higher than those of living microorganisms.
According to Posada et al. (Citation2010), the inactivation rate of viruses in an aerosol can be described by:
Here C is the concentration of viable viruses at the time t, C 0 is the initial concentration of viable viruses and k is the rate coefficient of inactivation. Even for the most stable viruses, k is typically of the order of 0.01 min−1, corresponding to an effective half-life of about one hour (Donaldson, Citation1973; McDevitt et al., Citation2008). For most other viruses, the inactivation rate in aerosol is considerably higher (Harper, Citation1961; Miller et al., Citation1963; de Jong, Citation1965; Miller and Artenstein, Citation1967; Songer, Citation1967; Akers, Citation1969; Benbough, Citation1971; Barlow, Citation1972; Donaldson, Citation1972; Akers, Citation1973; Donaldson and Ferris, Citation1974). Viruses are almost completely inactivated in aerosols in the span of 1 d under such conditions.
Experimental data on the survival of viruses show that, on the whole, the inactivation rate becomes higher for all viruses with increasing temperature (Zhdanov and Gaudamovich, Citation1982a; Zhdanov and Gaudamovich, Citation1982b; Weber and Stilianakis, Citation2008). Also, the number of surviving viral particles in aerosol decreases with increased radiation dose that the virus aerosol is exposed to (Jensen, Citation1964; Zhdanov and Gaudamovich, Citation1982a; Zhdanov and Gaudamovich, Citation1982b; Sagripanti and Lytle, Citation2007; McDevitt et al., Citation2008). Relative humidity influences the survival of viruses in aerosol differently. For example, the survival rate of influenza virus in aerosol is highest at high relative humidity, whereas that of foot-and-mouth disease virus is highest at intermediate relative humidity (50–60%) (Akers, Citation1969; Donaldson, Citation1972; Schaffer et al., Citation1976; Weber and Stilianakis, Citation2008).
The use of molecular biological methods for detecting genetic material of viruses also involves certain difficulties. As genetic material of different viruses is presented by very different variants: DNA or RNA (ribonucleic acid) molecules, single-stranded genomes (with + or – strand), fragmented genomes, etc. (Zhdanov and Gaudamovich, Citation1982a), no universal PCR primer for detection of all viruses has been developed so far. Consequently, such methods determine only the presence of expected viruses in samples. For example, Chen et al. (Citation2008c) presented the results of search for different subtypes of influenza-A virus in the atmosphere of Taiwan. Other viruses in atmospheric air samples were not even sought.
Virus-containing aerosols are formed in spray from water surfaces (Baylor et al., Citation1977a; Baylor et al., Citation1977b; Baylor and Baylor, Citation1980), from aerosolised virus-destroyed tissues of plants, insects, animals and birds; they are also shed by sick animals, birds and humans (Zhdanov and Gaudamovich, Citation1982b; Jones and Harrison, Citation2004). One should also note the intentional use of aerosols of insect viruses for plant protection (Morris, Citation1980; Maiorov et al., Citation1985; Jinn et al., Citation2009). The transfer of virus-containing aerosols in the atmosphere has been described for local scales, e.g. near sludge wastewater treatment plants and stock farms (Strauch and Ballarini, Citation1994; Carducci et al., Citation1995; Sigari et al., Citation2006; Langley and Morrow, Citation2010) as well as for regional scales, e.g. transfer of foot-and-mouth disease virus across the English Channel (Gloster et al., Citation1982) and transfer of avian influenza virus from continental China to Taiwan (Chen et al., Citation2008b). The hypothesis of transcontinental transfer of influenza A virus aerosol was demonstrated by Hammond et al. (Citation1989). Consequently, virus-containing aerosols can spread worldwide.
Relatively few studies have attempted to comprehensively estimate the concentration of different viruses in ambient air and evaluate their source strength. One example of such a study was reported by Safatov et al. (Citation2010) who collected 30 samples of atmospheric air (10–15 m3 each) onto fibrous filters during different seasons in Southwestern Siberia. Samples were analysed using the PCR method for the presence of viruses known to cause respiratory diseases, although no virus genetic material was found. It should be noted, however, that Southwestern Siberia is not endemic to airborne viral infections largely because it is located far from sources of viruses such as influenza (Chen et al., Citation2008c) and habitats of migrant birds transmitting influenza viruses (Liu et al., Citation2005). Electron microscopy during the same study detected bacteria, fungal spores and plant fragments (Safatov et al., Citation2010), but virus-like particles are impossible to confidently detect without genetic methods such as PCR, as discussed above.
As noted by Chen et al. (Citation2008b), the total concentration of different subtypes of influenza A viruses can reach 800 m−3 copies of genetic material of these viruses in the ambient atmosphere of Taiwan and up to 3·104 m−3 in outdoor pet markets in Taiwan. It should be noted that Taiwan, like the whole of South-East Asia, is an endemic area for this virus, which explains the higher influenza virus concentrations in this region. For other viruses originating from local sources, such as excreta of infected animals and humans, sewage treatment plants, use in agriculture, etc. (Fannin et al., Citation1985; Carducci et al., Citation1995; Sigari et al., Citation2006), the total numbers of viruses in aerosol are not large, and their contribution to the total mass of aerosol is negligible. As for more powerful sources, such as soil, vegetation and water surfaces, data on virus aerosol in the air in natural conditions are available only for the latter (Baylor et al., Citation1977a, 1977b; Baylor and Baylor, Citation1980). Virus-like particles can also be present in the atmosphere and water, such as those described by Leck and Bigg (Citation2005a) using electron microscopy. The concentration of virus-containing particles in the air is low, however (less than 100 cm−3 – total concentration of aerosol particles with diameter larger than 90 nm in marine atmosphere; data from Bigg et al. (Citation1995)), and by these methods it is impossible to distinguish viruses from particles on which they are bounded.
Virus aerosols collected from the exhaled air of infected animals have been the subject of various research studies. For example, for pigs infected with classical swine fever virus, the concentration of viral particles may reach 104 m−3 of air (Weesendorp et al., Citation2008). Other infections of animals and birds (such as foot-and-mouth disease, Newcastle disease, etc.) are characterised by virus aerosol exhalation of the same order of magnitude (Downie et al., Citation1965; Donaldson et al., Citation1983; Donaldson and Alexandersen, Citation2002; Li et al., Citation2009). Taking into account that the number of infected animals is typically not large, the total number of exhaled viruses in aerosol is similarly small.
To evaluate the total mass of viruses in 1 m3 of the atmosphere, let us consider an upper limit to the ambient concentration of viral particles to be 3·104 m−3. Assuming a per virus mass of 2·10−17 kg, one of the heaviest known viruses (vaccina virus, Zhdanov and Gaudamovich, Citation1982b) yields an estimated virus mass concentration of 6·10−4 µg m−3. This estimate is approximately four orders of magnitude smaller than the total concentration of biogenic substance in the atmosphere (Jaenicke, Citation2005; ) and thus, even if the concentration of viruses in the atmosphere was underestimated by an order of magnitude, the average mass concentration of viruses in atmospheric air would be very small compared to other biogenic aerosol mass.
2.5. Algae and cyanobacteria
The occurrence of algae in fresh and sea water is well known. However, algae living outside of the aqueous environment rarely attract attention. These algae are termed terrestrial, aeroterrestrial, aerophytic or subaerial, are able to reside on almost all substrates, natural or artificial and can become airborne, constituting the aero-(phyto) plankton. Chlorophycean and xanthophycean species are common worldwide (Printz, Citation1921; Laundon, Citation1985; Dubovik, Citation2002; Reisser, Citation2002; Sharma et al., Citation2007; Neustupa and Skaloud, Citation2010) and exist free living as well as lichenised (Bubrick et al., Citation1984). Due to the size of the algae and their spores, many smaller than 10 µm in diameter (Printz, Citation1921; Dubovik, Citation2002; Burchardt and Dankowska, Citation2003; Neustupa and Skaloud, Citation2010), they can be easily dispersed in the atmosphere. In a recent review article, the mechanisms involved in the aerosolisation of algae were presented, and Sharma et al. (2007) stated that ‘airborne algae are the least-studied organisms in both aerobiological and phycological studies’. This seems to be valid for aerosol studies, too. Quantitative measurements of algae in ambient air are very rare and it was reported (Reisser, Citation2002) that algal cells are present at a concentration of 300–500 cells m−3 of air on a dry and sunny summer day ().
Cyanobacteria belong taxonomically to bacteria, although they have long been considered as algae, as they have the ability to obtain their energy through photosynthesis. Due to their colour, they have been called blue-green algae, but they still belong – as they lack a cell nucleus – to the bacteria domain. As cyanobacteria can be found in almost every environment, have habitats across all latitudes, are widespread in oceans and freshwater, terrestrial ecosystems and bare rock and soils as well as in extreme habitats such as hot springs, they have been considered to be one of the most successful groups of microorganisms. They fulfil vital ecological functions in the world's oceans as important contributors to global carbon and nitrogen budgets (Stewart and Falconer, Citation2008). Cyanobacteria can not only occur as planktonic cells but also create biofilms in marine, freshwater and terrestrial environments. A few are involved in symbiosis with lichen, plants and other organisms and provide energy for their host. Although well studied in the marine environment, not much is known about their presence in the atmospheric environment.
Airborne cyanobacteria were found in Varanasi City, India, where they were more abundant than green algae and diatoms; Phormidium fragile as well as Nostoc muscorum were recorded throughout the 2 yr of sampling (Sharma and Rai, Citation2008). Similar observations were made in Cairo, Egypt. The species Chroococcus limenticus, Lyngbya lagerheimii and Schizothrix purpurascens appeared in all seasons (El-Gamal, Citation2008). Genitsari et al. (Citation2011) recently published an overview of taxa of airborne algae and cyanobacteria found in aerobiological studies.
2.6. Biological crusts and lichens
Arid and semi-arid regions of the globe exhibit biological crusts (also called rock varnish, cryptobiotic, microbiotic or biological soil crust) consisting of bacteria, fungi, algae, lichens and bryophytes in variable proportions (Belnap et al., Citation2003). Rock varnishes are natural, thin (5 µm – l mm), brown, black or grey (lead colour) coatings on rock surfaces (Krumbein and Jens, Citation1981). They consist largely of iron and manganese oxides with some quartz, clays and carbonates admixed. They may contain considerable amounts of organic material that sometimes may be responsible for the gel-like or lacquer-like appearance. The grey or black films may sometimes consist predominantly of dry cyanobacteria. Rocks outside of deserts often are also covered by a varnish (Douglas, Citation1987). Although these rock crusts are not suspended into the air by active mechanisms, they can be eroded from their host surface and broken into fragments producing small dust particles (Grini et al., Citation2002; Büdel et al., Citation2004). Parts of biological soil crusts can also be ablated from desert surfaces, and all these particles may contribute to the content of organic material in dust transported over long distances during storms (Grini et al., Citation2002; Büdel et al., Citation2004; Prospero et al., Citation2005; Hua et al., Citation2007).
As lichens are a symbiotic form of life between a wide range of fungi, algae or cyanobacteria (Hawksworth et al., Citation1995; Nash III, Citation1996) and distribute separately or together via the air, they also can, in certain circumstances, constitute a subgroup of PBAP. As biological aerosols, they may play a role in ice nuclei activity as well as in health effects. Lichen, for example, can cause human allergenic reactions (Richardson, Citation1975; Fahselt, Citation1994; Ingolfsdottir, Citation2002).
Although lichens are regarded as an individual taxonomic group – lichen taxonomy is based on the taxonomy of the fungal partner, the mycobiont (Tehler, Citation1996) – they also must be considered separately from their host because lichens are different in behaviour and life form from their isolated partners (Nash III, 1996; Tehler, Citation1996). Nearly 20% of all known fungi species are lichenised (Lutzoni et al., Citation2001) and more than 98% of the lichenised fungi belong to Ascomycota and a few to Basidiomycota (Hawksworth et al., Citation1995; Tehler, Citation1996). Most of the lichenised fungi form symbioses with Chlorophyta, whereas only about 10% lichenise with cyanobacteria and 3% with both (Tschermal-Woess, Citation1988; Lewis and McCourt, Citation2004).
In general, lichens can be found in three major life forms that all can contribute to biological aerosol particles: a crust-like biofilm (crustose), a leaf-like (foliose) and a branched tree-like biofilm (fruticose) as described in Hawksworth et al. (Citation1995) and Büdel and Scheidegger (Citation1996). Especially for PBAP formation, the presence of lichens in extreme habitats is interesting. They can deal with extreme light, dryness or temperature that are less favourable for higher plants. They are also tolerant to extreme desiccation and UV light exposure due to their cortical pigments (Nybakken et al., Citation2004; Gauslaa, Citation2005; Vráblíkova et al., Citation2006). Their ability to prevent the formation or to scavenge free radicals also increases their chance of surviving during their transport in air. Lichens face considerable problems in colonising new sites and maintaining existing populations. Mycobionts can distribute separately via Asco- or Basidiospores, or asexual by singular lichen structures, that contain both, mycobiont and phycobiont, such as insidia (protuberances from the thallus of corticated algae and medullary tissue), soredia (several algal cells encased by a hyphae) or hormocrusts (fragments if filamentous cyanophyte Nostoc spp with hypha penetrating the thallus fragments (Marshall, Citation1996; Oksanen, Citation2006)).
Although lichen species have been detected in the atmosphere, estimates of lichens biomass and consequently estimates of the number of lichen-derived aerosol particles are difficult. Lichen species are adapted to almost every temperate to extreme environment on Earth, such as arctic tundra, hot deserts, rocky coasts, or toxic slag heaps, but the spread of lichen population within each environment type is different. They are abundant on plant leaves or branches in rain forests or temperate woodlands, as well as on bare rock including walls, and on exposed soil surfaces. Henderson-Begg et al. (Citation2009) suggested that canopy lichen biomass in temperate forest is similar to leaf biomass. Lichen detection directly in air has been pursued by Marshall (Citation1996) in an aerobiological monitoring programme, where he found lichen soredia to be the most abundant airborne propagule with a size range of 30–100 µm on Signy Island in Maritime Antarctica.
Lichens are not only adapted to survive hot and dry environmental conditions but are also often frost tolerant (Kershaw, Citation1985). In ice nucleation studies pursued by Kieft (Citation1988), nearly all lichens tested showed ice nucleation activity at temperatures above −8 °C or even above −5 °C (see also ). In theory, the IN could be the bacterium or the fungal partner, but the failure to culture IN bacteria from lichen and also the high density of IN in active lichen carrying only few numbers of bacteria suggests a non-bacterial source of lichen IN. Studies by Kieft and Ahmadjian (1989) show that in their tests, always the mycobiont was responsible for IN activity. IN activity in lichen might enhance the uptake of atmospheric moisture by enhancing the condensation or causing deposition of ice from water vapour. Lichen-associated IN might even contribute to atmospheric IN (Kieft, Citation1988).
2.7. Others
Apart from the PBAP defined above, there exist a variety of other primary aerosol particles from the biosphere. Plant fragments, in particular, are one of the largest mass fractions of atmospheric PBAP and comprise a wide spectrum of decaying matter. However, drawing a consistent definition between PBAP and other organic matter can be difficult because plant materials can eventually be broken down into humic-like substances (HULIS, Fuzzi et al., Citation2006; Graber and Rudich, Citation2006) by oxidative modification and degradation of biopolymers, and are therefore, important components of soil. Cellulose, a homopolymer of D-glucose (Pöhlker et al., Citation2011), is the most frequently occurring biopolymer in the terrestrial environment (Butler and Bailey, Citation1973; Sánchez-Ochoa et al., Citation2007), although other biopolymers and structural components (e.g. lignin, chitin) may also be atmospherically relevant (Pöhlker et al., Citation2011). Cellulose is a major component of plant tissue and pollen and can be produced by some bacteria (Cannon and Anderson, Citation1991) but is not present in insects and animalian tissue (Winiwarter et al., Citation2009). Atmospheric concentrations of cellulose thus exhibit a strongly seasonal cycle and have been shown to have mass concentrations at least in the order of ~0.5 µg m−3 (Puxbaum and Tenze-Kunit, Citation2003). Presence of cellulose in atmospheric aerosols has also been frequently used as a proxy for total concentrations of plant debris (e.g. Sánchez-Ochoa et al., Citation2007; Winiwarter et al., Citation2009). Due to large particles of biological material being lifted into the atmosphere (e.g. air-dispersed seeds), the actual concentration of cellulose and related materials in total air particulate matter may be substantially higher under windy conditions.
Emissions of liquid and solid secretions from organisms are also found in the atmosphere. Brochosomes and epicuticular wax (Wittmaack, Citation2005; Wittmaack et al., Citation2005) are expelled from leafhoppers and do not contain cells. Brochosomes serve as a highly water-repellent body coating of leafhoppers. They usually become airborne not as individuals but in the form of large clusters containing up to 10 000 individuals or even more, but then usually separate into individual brochosomes or small clusters. They are extremely widespread and can be found in aerosol samples from all continental regions (Bigg, Citation2003). Exopolymer secretions (EPS) of microalgae and bacteria in water become airborne with the bubble- bursting mechanism (Leck and Bigg, Citation2005a, Citation2005b). EPS might deteriorate the ability of sea spray particles acting as Cloud condensation nuclei (CCN, see studies in Section 4.1).
Other sources of PBAP are fur fibres, dandruffs and skin fragments of animals (including humans) that are shed in large amounts per day and can also be airborne (Cox and Clark, Citation1973). Insect fragments have also been observed in a number of studies and thus may represent an important class of physically large PBAP in some areas (e.g. Wittmaack, Citation2005). Whole macroscopic organisms, such as spiders lofted to air by silk ‘ballooning’ lines, have been observed to travel extremely long distances (>several 100 km) and to high altitudes (stratosphere) (e.g. Thomas et al., 2003). These organisms may contribute to PBAP dispersal efficiently through body or silk fragments.
2.8. Characteristic concentrations and emission estimates
Biological particles cover an extremely broad range of sizes and are morphological very diverse. Thus, measurements of ambient PBAP concentrations are extremely challenging and reports from literature have been very few. PBAP have unique properties, such as large particle size, cell viability concerns and physical changes associated with environmental variables (see Section 3.1). In addition, biological material suspended in the atmosphere undergoes drastic changes as a function of manifold biological variables such as species and ecosystem health, seasonal and episodic cycles and meteorology. As a result, average concentrations for PBAP are extremely difficult to estimate without rigorously taking all of these variables into account. However, PBAP concentrations have been estimated to be up to 25% of total aerosol mass on a global basis (Jaenicke, Citation2005), and so more detailed measurements are desperately needed.
As a first overview of global importance, present a survey of global emission estimates and characteristic magnitudes for the number and mass concentration of major PBAP classes, respectively. Such synthesis provides means for discussion and motivation of future measurements.
3. Techniques for PBAP collection and analysis
3.1. PBAP sampling methods
The sampling and collection of atmospheric aerosols requires a sound understanding of the physical principles that govern interactions with suspended particles (<100 µm). As such, particle size is by far the most important characteristic for choosing a sampling procedure for both PBAP and all other classes of airborne particles (Nicholson, Citation1995). Losses of particles within inlets, inlet lines and instruments can cause huge biases in both quantitative and qualitative understanding, and therefore sampling procedures must be designed properly (Zimmermann et al., Citation1987). While small particles (<0.1 µm) are most susceptible to Brownian diffusion and electrostatic forces, large particles (>5 µm) are more influenced by losses due to gravitational settling and inertial surface impaction. In addition, losses of particles to surfaces due to air turbulence can affect particles of all sizes. These issues are dealt with in detail elsewhere (e.g. Hinds, Citation1999; Baron and Willeke, Citation2001).
However, the sampling of biological aerosols often requires special handling procedures as compared to the sampling of inert particles and these are introduced briefly here. Although sampling issues due to particle size effects are not unique to biological aerosol sampling, PBAP size and mass are often hard to predict based on complicated relationships between the biological tissue and environmental variables such as temperature and relative humidity (e.g. Madelin and Johnson, Citation1992; Reponen et al., Citation1996). In addition, many biological particles (e.g. chain agglomerates of fungal spores) can have very elongated morphologies that can create large differences between physical and aerodynamic diameter measurements (Lacey, Citation1991; Reponen et al., Citation2001). Collection of living, or viable, material requires care in order not only to sample microorganisms appropriately but also to keep them alive until the desired analysis or cultivation can be performed. Certain species of bacteria and fungal spores are more easily cultivated than others, and in many cases this relates to the ability of sampled particles to survive harsh conditions related to sample collection (i.e. sun exposure, extended period of high airflow). Bacteria, in particular, are susceptible to damage or death when impacted too forcefully onto collection substrates (e.g. Stewart et al., Citation1995). Samples also must often be immediately frozen after collection to preserve DNA for later analysis. This can complicate sampling of PBAP in remote environments and add both uncertainty and bias to measurements. Small biological particles (e.g. viruses, bacteria) may also be attached to larger, non-biological particles such as mineral dust and thus be sized much larger than the microorganism by itself. This fact must be kept in mind when analysing size-resolved PBAP measurements. Due to the additional complexities in determining the biological nature of a collected set of particles compared to that of non-biological particles, PBAP measurements should be carefully scrutinised and in many circumstances taken as lower limit values.
As a result of the issues specific to biological aerosol sampling, a number of sampling methods have been developed primarily for PBAP collection. Again, a review of these topics can be found elsewhere (e.g. Henningson and Ahlberg, Citation1994; Madelin, Citation1994; Crook and Sherwood-Higham, Citation1997; Levetin, Citation2004; Xu et al., Citation2011) and so only a very brief overview of key classes of PBAP samplers will be given here. Devices that collect aerosol based on inertial forces have been particularly well utilised for PBAP measurement. The most common single-stage impactor used for microbiological sampling has been the relatively simple slit-to-agar sampler (Henningson and Ahlberg, Citation1994), first developed by Bourdillon et al. (Citation1941) and capable of providing CFU counts with high time resolution of approximately 1 h. Many different cascade impactors (multiple stages) have also been developed and extensively used for PBAP sampling (Mitchel, Citation1995). Probably, the most widely used of these has been the Andersen sampler (Andersen, Citation1958). This device has been frequently used as a reference method for sampling of culturable microorganisms, providing six or eight stages of particle sizing but due to physical constraints, the inner jets on each plate have a different sampling efficiency than the outer jets. An alternative in which this phenomenon is considered is the Berner-Impactor (Hillamo et al., Citation1991). The Marple 8-stage impactor is a personal cascade impactor design that provides size-resolved PBAP information directly from the breathing zone of an individual who wears it (Rubow et al., Citation1987). The Burkhardt spore trap has been an important sampler for collecting large PBAP such as fungal spores and pollen (Hirst, Citation1952). In it, air is brought in through a small slit in a housing designed to face the prevailing wind, and material collected continuously for up to a week can later be analysed via microscopy. Impingers collect airborne particles into liquid media and offer the possibilities of extended sampling times and increased collection efficiency with respect to many agar samplers (Crook, Citation1995a). Greenburg and Smith (Citation1922) developed the first liquid impinger, and many types have since been designed for PBAP sampling (e.g. Henderson, Citation1952.). The AGI-30 has been particularly important and has been used as a reference sampler for the collection of viable organisms (Brachman et al., Citation1964; Henningson and Ahlberg, Citation1994). Other inertial samplers that have been commonly applied to PBAP collection include rotary arm samplers and both single- and multiple-stage centrifugal cyclones (e.g. IPCC Citation2007b; Crook, Citation1995a).
Non-inertial sampling methods such as passive settling and filtration have also both been widely used for PBAP collection. Many types of filter substrates have been used, including fibrous, flat (e.g. Nucleopore™) and membrane (Crook, Citation1995b). Each have their own advantages and disadvantages and these should be investigated thoroughly by the individual investigator. Electrostatic (Mainelis et al., Citation1999 Citation2002; Tan et al., Citation2011) and thermal precipitators (Kethley et al., Citation1952; Orr et al., Citation1956) have been developed in an attempt to collect viable aerosols with less damaging force. Recently, methods for online detection of biological aerosols have been developed using a variety of optical and additional techniques. These will be discussed in detail in Section 3.3.
3.2. Traditional analysis methods
Before the invention of molecular and other physicochemical methods, the diversity, identity and concentration of airborne microorganisms were primarily studied via microscopic analysis and cultivation methods on both selective and non-selective media. These methods have long and rich histories within aerobiological studies and still remain useful tools for PBAP analysis. Thus, the terms provided here (‘traditional’ and ‘modern’) are historical and somewhat arbitrary, but helpful for rough categorisation. A detailed discussion of these methods is beyond the scope of this text and can be found elsewhere (e.g. Cox and Wathes, Citation1995).
3.2.1. Cultivation
Cultivation methods are only capable of collecting and detecting certain viable bacteria, fungi and algae, whereas they are incapable of detecting all other biological aerosol particles from these and other organism groups (e.g. dead bacteria and fungi, tissue fragments such as cell walls or cytoplasmic material). Although not capable of directly infecting a host, non-viable PBAP classes are still extremely important because they can provoke deleterious health effects (Gorny et al., Citation2002; Green et al., Citation2006) and may still be relevant for cloud formation.
As mentioned, studies investigating only the culturable fraction of PBAP have historically comprised a large portion of reported measurements. However, studies investigating cell viability suggest that the vast majority of environmental microbiota is non-culturable, even when viable (Staley and Konopka, Citation1985; Roszak and Colwell, Citation1987; Amann et al., Citation1995; Colwell, Citation2000; Rappé and Giovannoni, Citation2003; Wainwright et al., Citation2004b). Thus, studies based on culturing alone usually drastically underestimate microbial diversity (Fierer et al., Citation2008) and concentration. Only ~17% of known fungal species can be grown in culture (Bridge and Spooner, Citation2001) and for bacteria, the fraction is typically less than 10%, with an observed range of 0.01%–75%, and average values estimated at ~1% (Heidelberg et al., Citation1997; Lighthart, Citation2000; Chi and Li, Citation2007). In addition, the culturability of bacteria decreases rapidly following aerosolisation, (Heidelberg et al., Citation1997) and bacteria can be easily damaged when impacted via traditional sampling devices (Stewart et al., Citation1995). Within the culturing processes, additional biases may occur. For example, the sampling efficiency and culturability of viable bacteria depend strongly not only on the bacterial strain but also on experimental and environmental factors, including the growth medium used (ZoBell and Mathews, Citation1936; Kelly and Pady, Citation1954; Shahamat et al., Citation1997; Griffin et al., Citation2006), the choice of impaction versus filtration as a collection method (Stewart et al., Citation1995), the incubation temperature and length of incubation time (Amato et al., Citation2007c; Wang et al., Citation2007; Wang et al., Citation2008), the air sampler volume (Griffin et al., Citation2006), relative humidity (Wang et al., Citation2001), time of day (Tong and Lighthart, Citation1999) and even the collection season (Amato et al., Citation2007c). Finally, single particles containing bacteria may represent a colony consisting of many cells, leading to a further underestimation of their abundance (Tong and Lighthart, Citation2000).
Despite these limitations, cultivation methods have the advantage that they are far less expensive than molecular techniques and can give an indication of the number of viable cells in the air, for species that respond well to the culturing method used. Cultivation is particularly useful for targeting individual species or groups of microorganisms and for creating culture collections. Culture studies can also characterise certain bacterial strains by various biochemical methods and give qualitative information about relative changes in the concentration over the course of days or season.
3.2.2. Light microscopy
In addition to cultivation, microscopic techniques have been extremely important to the history and development of PBAP analysis and continue to be invaluable tools. Light microscopy of various types has been applied to the characterisation of collected PBAP (Spurny, Citation1994; Cox and Wathes, Citation1995 and references therein). For pollen, it still remains the most common analysis method. Simple optical microscopy was the first technique to be applied to PBAP analysis in the seventeenth century (e.g. Miquel, Citation1883) and continues to be utilised extensively. But, it has to be taken into consideration that particles <2 µm are only visible as dots under an optical microscope and thus cannot be analysed in detail according to their size and shape. Aerosol samples collected by sedimentation and impaction devices can be directly visualised and counted by light microscopy. If the sampling device used is volumetric, the concentration of particles in the air can be quantified. However, the identification of biological particles via direct counting is both very tedious and somewhat subjective, as the particles must be counted by eye. Various traditional stains can be used in combination with light microscopy, including methylene blue for simple examination, and Gram staining and other differential staining techniques that aid classification into groups of species. Protein staining gives an estimate of all (total) PBAP (Matthias-Maser and Jaenicke, Citation1995). For the analysis of the diversity and composition of bacteria and fungi, microscopic analyses are not usually reliable, as small non-descript spores and hyphae or fragments of fungal tissues cannot be classified (Pitt and Hocking, Citation1997). Some fungi or bacteria may remain morphologically undistinguishable or may only be identifiable to a class or family level.
3.2.3. Fluorescence microscopy
Fluorescence microscopy has been applied both to look at the autofluorescence of PBAP (Pöhlker et al., Citation2011), and especially with the use of fluorescent dye labelling (Karlsson and Malmberg, Citation1989; Hernandez et al., Citation1999). The most common traditional method for determining the total count of environmental microorganism is direct fluorescence microscopy, either by taking advantage of the autofluorescence of certain biological compounds or by using samples that have been treated with a fluorescent dye, most commonly 4.6-diamidino-2-phenylindole (DAPI) or acridine orange (Francisco et al., Citation1973; Hobbie et al., Citation1977; Kepner and Pratt, Citation1994; Matthias-Maser and Jaenicke, Citation1995; Harrison et al., Citation2005). Acridine orange binds to both DNA and RNA, fluorescing green when bound to DNA and red when bound to RNA and some mucins. DAPI fluoresces blue when bound to DNA and yellow when unbound or bound to a non-DNA material. Traditionally, particles have been classified and counted by a human investigator, taking into account the size and morphology of stained particles. Epifluorescence microscopy permits counting of the total number of unlysed cells containing DNA; a number that includes both viable and non-viable microorganisms. It is not always possible to unambiguously distinguish different types of biological particles, such as bacterial and fungal spores, via fluorescence microscopy.
However, the colour of autofluorescence combined with the application of fluorescent stains can be used to distinguish some broad groups of microorganisms. Fluorescence microscopy is both tedious and labour intensive, although the use of fluorescence aids the identification of biological particles compared to light microscopy. Recently, more automatic techniques have been attempted, including computer analysis of microscopic images (Carrera et al., Citation2005) and fluorescence spectroscopy (Reyes et al., Citation1999; Courvoisier et al., Citation2008). Other modern fluorescence-based techniques will be described in the next section.
3.3. Modern analysis methods
3.3.1. Molecular techniques
3.3.1.1. Chemical tracers
Chemical tracers, such as the sugar alcohols mannitol and arabitol, can be used to not only assess the abundance of PBAP, especially fungal spores in air particulate matter, but also characterise other aerosol types (Hensel and Petzhold, Citation1995; Graham et al., Citation2003; Graham et al., Citation2004; Lau et al., Citation2006; Elbert et al., Citation2007; Yttri et al., Citation2007; Bauer et al., Citation2008a; Engling et al., Citation2009; IInuma et al., 2009; Burshtein et al., Citation2011). A number of other chemical tracers have been used as proxies for various types of biological aerosol particles, such as endotoxins, mycotoxins, glucan, ergosterol, extracellular polymeric substances, carbohydrates, proteins, peptides, sugars and adenosine triphospate (ATP). These can be analysed by a wide range of instrumental and bioanalytical techniques such as chromatography coupled to mass spectrometry, spectrophotometry, fluorescence spectroscopy, immunoassays, dye assays, etc. (Griffith and DeCosemo, Citation1994; Reponen et al., Citation1995; Franze et al., Citation2005; Pöschl, Citation2005; Demirev and Fenselau, Citation2008).
Chemical tracer analysis has the advantage of providing quantitative information, but it does usually not provide information about the identity and biodiversity of PBAP on the species level.
3.3.1.2. Nucleic acid sampling and extraction
For the study of biological aerosols molecular techniques, e.g. those based on molecular genetic analyses, have some advantages compared to traditional methods. The results can enable the identification and quantification of culturable as well as uncultivable microorganisms, of viable and dead cells, and of plant and animal fragments (Després et al., Citation2007).
To analyse biological aerosols with molecular genetic tools, biological aerosols need to be collected and the nucleic acids extracted. Biological aerosols are either collected in culturing media (liquid or plates) or water (Boreson et al., Citation2004), special biological aerosol particle collectors (e.g. the Wetted-wall Cyclone sampler (Biotrace) in Maron et al. (Citation2006)) or on appropriate air filter samples. Several different filter types have already successfully been used for the DNA analysis but not been compared quantitatively to each other (e.g. glass fibre filters (Després et al., Citation2007; Fröhlich-Nowoisky et al., Citation2009); quartz fibre filters and polypropylene filtres (Després et al., Citation2007); cellulose nitrate filtres (Després et al., Citation2007; Bowers et al., Citation2010); Celanex polyethylene terephthalate filtres (Brodie et al., Citation2007); borosilicate filtres (DeSantis et al., Citation2005) etc.).
The basis for most molecular analyses techniques is the successful extraction of DNA. Every cell carries this molecule: eukaryotes carry DNA in the form of chromosomes within the cell nucleus, whereas prokaryotes and archaea lack a nucleus and carry DNA in genophores directly within their cell body. During the DNA extraction processes, proteins are denatured and separated together with lipids, pigments, cell wall fragments and organelle structures.
DNA extraction protocols vary according to the tissue for which they are used. Plant cells, with strong cell walls, present different extraction challenges than animal tissue cells. Endospores of bacterial and fungal spores have very strong cell walls that need to be broken down before DNA extraction. Thus, some compromise is always required in selecting a DNA extraction method for ambient air filter samples that contain a mixture of various organisms and tissue types. Either the DNA is extracted using a generic method that does not exclude any tissue type but is also not ideally matched to any specific type such that some organisms and tissue types are undersampled, or the DNA is extracted using a method specific to a particular organism or tissue type, and other types are undersampled. Thus, for ambient samples that include a mixture of many types of biological material, PBAP that contain thick membranes or cell walls could elude extraction of DNA by a generic method, leading to an underestimation of such species. Usually, commercial kits can be used for the extraction (e.g. soil DNA extraction kit (Després et al., Citation2007; Fröhlich-Nowoisky et al., Citation2009)); Fast Prep 120 agitator together with Spin column of MolBio Laboratories (DeSantis et al., Citation2005); alternatively, some investigators develop their own extraction methods (e.g. Boreson et al., Citation2004; Maron et al., Citation2005).
3.3.1.3. Amplification of genomic DNA
The DNA extract of an air filter sample still contains the genomic DNA of all sorts of organisms; thus, it is a mixture of DNA from living and dead material of fungi, bacteria, archaea, plants, animals or viruses. To be able to identify single genera or species, it is necessary to enrich the DNA of the organism of interest relative to other genomes. The PCR efficiently amplifies characteristic regions of the DNA of a species, or a group of species, for detailed analyses. The method is based on thermal cycling in which the DNA is heated, thereby separating the double strands. Short DNA sequences, called primers, attach to matching sequences on the single stranded DNA that mark the borders of the region of interest on the genome. In the final step of PCR, the DNA polymerase enzymatically assembles a new matching DNA strand in the target region using single nucleotides. PCR, followed by genetic sequencing of the amplified DNA, can be used to identify organisms, to understand phylogenetic relationships or to study specific genes. Thus, it is an essential technique when studying the identity, diversity and composition of biological aerosols.
Within every genomic DNA, there are areas that are highly conserved between organisms such as housekeeping genes, whereas other areas vary enormously. The primers used in PCR should ideally attach to highly conserved areas, whereas the sequence between the primers should be diverse and thus uniquely present in the targeted organisms or group of organisms. Areas such as ribosomal RNA genes or the ITS-regions with a huge variability are often used in taxonomic identification studies, as they enable to differentiate between genera or even species and thus can be used as a fingerprint of a species. For bacteria, e.g. often the 16S rRNA genes are amplified (Moffett et al., Citation2000).
While amplification followed by taxonomic analysis allows the identification of PBAP species within aerosol samples, it gives no information about the quantity of particular species in the air. Quantitative PCR allows the calculation of the number of DNA template molecules in the DNA extract. Thus, when studying, e.g. a single copy gene within a single bacterial species, it is possible to estimate the number of individuals of this particular species that was present in the air filter sample. This could be an especially valuable tool for studying pathogenic biological aerosol particles.
Although molecular techniques are thought to detect biological aerosols unambiguously, they may not amplify all airborne bacteria in a sample (Peccia and Hernandez, Citation2006; Fierer et al., Citation2008). Theoretically, the DNA polymerase can amplify a single DNA copy and thus detect even organisms that are present in minor quantities. But, the sensitivity of the PCR depends on several factors. One major issue is the choice of the primer pair used for the amplification process. Different primer pairs have different specificity and sensitivity (Alvarez et al., Citation1995; Polz and Cavanaugh, Citation1998). Thus, while one primer pair might start the amplification from just a single DNA strand, a less sensitive primer pair might need 100 molecules to get started. It is also possible that a primer pair is more sensitive to some species than to others within a targeted group. Although a primer pair might have been designed to perfectly match the target organism, it might also co-amplify other organisms. This is especially important for biological aerosol DNA extracts where many competing DNA molecules may be present, possibly even in higher concentrations than the organism of interest. Literature research can help to find suitable specific primers, but often the specificity of primers is overstated in the literature (e.g. Fröhlich-Nowoisky et al., Citation2009).
Another important point that must be considered is the possible presence of PCR inhibitors. Substances such as humic acids inhibit the DNA amplification process either by hindering the attachment of polymerase to the primers to initiate amplification, or by binding to the DNA and thereby preventing primers or enzymes from attaching. Different PCR primers as well as DNA polymerases vary in their ability to overcome inhibitory factors. In addition, filter materials on which biological aerosol particles are collected can inhibit PCR (Després et al., Citation2007).
Finally, although DNA is a stable molecule, which under cool, dark and dry conditions can sometimes be preserved for several thousand years (Pääbo et al., Citation2004), DNA starts to degrade, breaks into smaller pieces and is chemically modified as soon as an organism dies (Pääbo, Citation1989; Lindahl, Citation1993; Höss et al., Citation1996; Smith et al., Citation2001). UV light, ozone and melting-freezing processes speed up these degradation processes, and thus a long residence time of biological material in air leads to deterioration of DNA and loss of genetic information. Pollen grains, as well as many spores of fungi and bacteria, are encased by thick cell walls to protect their DNA from DNA-destroying environmental processes.
3.3.1.4. Restriction fragment length polymorphism techniques
A standard molecular genetic technique is the fragmentation of DNA by a restriction enzyme that cuts the DNA wherever a specific target sequence occurs. The resulting restriction fragments are separated according to their lengths by gel electrophoresis. It can be applied following colony PCR (confirmation of plasmid insertion) to select as many different clones as possible for the sequencing reaction (Fröhlich-Nowoisky et al., Citation2009). For a broad characterisation of the community structure and diversity of a PBAP sample, the terminal restriction fragment length polymorphism (T-RFLP) technique can be used to get a rough estimate of the diversity and relative abundances (Després et al., Citation2007; Georgakopoulos et al., Citation2009). The first step of a T-RFLP is a PCR amplification in which one primer is fluorescently labelled. The PCR products are then digested with a restriction enzyme that cuts each DNA strand at the target sequence, resulting in shorter, labelled fragments. Because the position of the target sequence on the genome varies among bacterial strains, the length of the labelled fragments also varies. The fluorescently labelled end fragments are separated by electrophoresis, and the strand lengths and fluorescence intensities are calculated. The fluorescence intensity of strands of a given length indicates the frequency of the corresponding bacterial strains in the original sample. The genetic diversity indicated by a T-RFLP profile (as measured by the number of different strand lengths) is highly dependent on the choice of the restriction enzyme and the part of the gene used to generate the terminal fragments. If the PCR products are simultaneously cloned and sequenced, the size of the terminal fragment that each will produce can be calculated, and so each sequence can be attributed to a T-RFLP peak (Després et al., Citation2007; Georgakopoulos et al., Citation2009).
3.3.1.5. Sequencing methods
To determine the identity of the genomes obtained from atmospheric aerosol samples, the PCR products are often cloned and sequenced (e.g. Boreson et al., Citation2004; Maron et al., Citation2005; Després et al., Citation2007; Fierer et al., Citation2008; Bowers et al., Citation2009; Fröhlich-Nowoisky et al., Citation2009; Georgakopoulos et al., Citation2009). Species can often be identified by comparing the obtained sequences with those that are already available in online databases, e.g. that of the National Center for Biotechnology Information (NCBI). The BLAST searchFootnote4 is the easiest way to search for matching sequences. If a new organism (e.g. new bacterial strain) is found, it is possible to identify the most closely matching sequences already available in the database. For bacteria, the generally accepted levels of discrimination are 97–99% similarity for species and 95–97% for genera; for fungi, clustering sequences with 97% or greater sequence similarity are accepted (O'Brien et al., Citation2005; Bowers et al., Citation2009; Fröhlich-Nowoisky et al., Citation2009; Georgakopoulos et al., Citation2009). The traditional sequencing technique is using a chain termination (Sanger and Coulson, Citation1975; Sanger, Citation1981). The sequencing process is essentially another PCR but only one primer instead of two is used. The reaction mix contains not only the standard DNA bases (deoxynucleotide-triphosphate, dNTP) but also chain terminator dideoxynucleotide-triphosphates (ddNTPs) that are labelled with fluorescent dyes that emit light at different wavelengths (dye-terminator sequencing). The incorporation of a ddNTP results in a chain termination, and the fluorescence peaks are detected after capillary electrophoresis resulting in the DNA sequence.
The high demand for low-cost sequencing in recent years has led to the development of high-throughput sequencing technologies. In these technologies, the sequencing process is parallelised for several samples, and thus in a short time thousands or millions of sequences are produced. One high-throughput technique, 454 pyrosequencing, has been applied successfully several times in biological aerosol research (Bowers et al., Citation2009, Citation2011). The DNA is amplified within an oil droplet by the so-called emulsion PCR. Each oil droplet contains a single DNA template attached to a single primer-coated bead that then forms a clonal colony. The sequencing machine contains many picolitre-volume reactions areas, each containing a single bead and sequencing enzyme. Pyrosequencing uses luciferase to generate light for detection of the individual nucleotides added to the nascent DNA, and the combined data are used to generate sequence readouts. For studies in which the taxonomic identification of biological aerosols is anticipated, the amplified sequences must have a minimum length to enable differentiation at the genus or species level. Often, this minimum length is around 600–1000 bp (base pairs). In traditional sequencing, this length can be reached easily, whereas in high throughput sequencing the length of the different sequences is usually only around 300–400 bp. Thus, with current high-throughput techniques, it is often not possible to identify biological aerosol particles on a species or genus level.
3.3.1.6. Hybrid and chip technology
Microarrays can be used for the characterization of PBAP (Wilson et al., Citation2002; Brodie et al., Citation2007; Georgakopoulos et al., Citation2009). Species or group specific probes (e.g. bacterial 16S rRNA genes of DNA; Loy and Bodrossy, Citation2006) are fixed on glass slides. DNA from atmospheric samples is fluorescently labelled and can hybridise with the DNA on the microarray if complementary sequences exist in the microarray. The sequence is determined from the position of the fluorescence on the chip. This technology was recently used on atmospheric aerosol samples for bacterial 16S rRNA genes (Wilson et al., Citation2002; Brodie et al., Citation2007). Both studies compared the cloning and sequencing method with microarray technology that was found to be more sensitive in detecting bacterial taxa.
3.3.2. Optical methods
Most of the above-mentioned methods for detecting PBAP are limited to offline analyses that can be time consuming, costly and which often provide measurements that suffer from poor time resolution (hours/days). However, much effort has been invested in the last decades, largely by military research facilities interested in quick detection of bio-warfare agents, to be able to detect biological aerosols in real time and with high time resolution. However, a comprehensive review of all modern methods for PBAP detection is well beyond the scope of this text and the following section is intended to present an overview of the most important classes of techniques. It focuses on field-based techniques, although some techniques that are primarily laboratory based are also discussed, as they may have important relevance to ambient detection.
3.3.2.1. On-line autofluorescence methods
All biological materials contain fluorophores that may be helpful for identification. Among most existing instruments that utilise biological auto-fluorescence for online determination of biological aerosol particle concentration, the choices of wavelength for the excitation source can generally be classified into one of two regions. Sources that provide light in the region of approximately 350–370 nm enable detection of reduced pyridine nucleotides (e.g. NAD(P)H) and riboflavin that are biological molecules linked to cellular metabolism (Harrison and Chance, Citation1970; Eng et al., Citation1989; Kell et al., Citation1991; Li et al., Citation1991; Iwami et al., Citation2001). Detection of auto-fluorescence under these conditions may indicate the presence of viable biological material in the aerosol particles, although other biological molecules (e.g. chlorophyll, cellulose) can also auto-fluoresce under many environmental conditions. The second region of fluorescence excitation commonly utilised by biological aerosol instrumentation is approximately 260–280 nm and highlights amino acids, such as tryptophan, tyrosine and phenylalanine that are present in all proteins (e.g. Teale and Weber, Citation1957; Pöhlker et al., Citation2011).
Thus, it can be broadly stated that detection of fluorescence by this class of instruments allows determination of fluorescent biological aerosol particles (FBAP). However, different instrument designs measure and report different fractions of the PBAP and comparisons across instruments should be conducted with this in mind. Among the uncertainties involved is the possibility that material of non-biological origin will also fluoresce within a given particle, causing a positive artifact. However, non-biological particles producing such artefacts are unlikely to contribute significantly to the coarse (>1 µm) fraction of ambient particulate mass (Huffman et al., Citation2010) and may also exhibit much weaker fluorescence than biological particles. For example, results of a study performed in remote Amazonia indicated that coarse FBAP closely approximated PBAP concentrations and size distributions (Pöschl et al., Citation2010). It is also known that not all biological microorganisms or fractions will fluoresce under the experimental conditions of such instruments, and opaque or absorbing PBAP are likely to fluoresce only very weak at best (Pöhlker et al., Citation2011). The weak fluorescence of some PBAP suggests that fluorescence measurements underestimate the total biological material present. Further investigation will be required to achieve full understanding of the response of fluorescence-based biological aerosol particle detectors to all types of biogenic aerosol particles, and to quantify potential interferences by fluorescence of non-biological particles. However, our present understanding is that FBAP can generally be regarded as an approximate lower limit for the actual abundance of PBAP (Huffman et al., Citation2010).
The Ultraviolet Aerodynamic Particle SizerFootnote5 (UV-APS) was the first commercially available, fluorescence-based instrument for real-time analysis of biological aerosols (Hairston et al., Citation1997; Brosseau et al., Citation2000). The UV-APS measures the aerodynamic diameter and side scatter parameter (analogous to the optical diameter) of incoming particles by measuring their time-of-flight between two lasers (633 nm) and then detects fluorescence in the wavelength range of 420–575 nm after excitation by a pulsed ultraviolet laser (Nd:YAG, 355 nm). An example of UV-APS measurement data is given in . This shows time series and size distributions of FBAPs detected with a UV-APS at a site in central Europe (Mainz, Germany, October 2006). The figure highlights the ability of the UV-APS to measure FBAP size distributions of discrete particle events with much higher time and size resolution than is achievable by most traditional sampling and detection methods (Huffman et al., Citation2010).
Fig. 4. Characteristic time series and number size distribution of fluorescent biological aerosol particles (FBAPs) measured with an ultraviolet aerodynamic particle sizer (UV-APS) in central Europe (Mainz, Germany, October 2006). The peaks at ~1.5, ~3 and ~13 µm can be attributed to bacteria, fungal spores, and pollen. N F,c is the number concentration of FBAPs, and N T,c is the number concentration of total aerosol particles with aerodynamic diameters D a>1 µm; dN F/dlogD a is the number size distribution function of FBAPs (CitationHuffman et al., 2010).
The Wide Issue Bioaerosol Spectrometer (WIBS) is also available in limited commercial production and not only provides conceptually similar information to the UV-APS but also provides a rough estimate of particle sphericity (Kaye et al., Citation2000 Citation2005). Incoming particles are first optically sized by measurement of scattered laser light. UV pulses at ~280 and ~370 nm from a xenon flash lamp excite each particle, and multiple bands of fluorescent emission in the range of 310–400 nm (for 280 nm excitation) and 420–650 nm (for 280 and 370 nm excitation) are recorded.
A number of other instruments have been developed for military research use and have not been commercialised or widely used for measurements published within peer-reviewed literature. However, these technical and scientific developments have been significant contributions to the scientific community (e.g. Hill et al., Citation1995; Cheng et al., Citation1999; Reyes et al., Citation1999; Seaver et al., Citation1999; Kopczynski et al., Citation2005; Cabredo et al., Citation2007; Campbell et al., Citation2007; Manninen et al., Citation2008; Pan et al., Citation2009; Sivaprakasam et al., Citation2009).
Although the development of such instruments has left a detailed trail in publically accessible military reports and peer-reviewed literature, ambient measurements by such techniques have been less frequently documented or de-classified. An early generation UV-APS (Ho, Citation2002) was used for short periods to measure background FBAP concentrations at several military locations within Canada and Sweden (Ho and Spence, Citation1998; Ho et al., Citation2004), and a different LIF spectrometer design was utilised for detection of FBAP in ambient air in several US locations (Pinnick et al., Citation2004; Pan et al., Citation2007; Pan et al., Citation2009). Recently, fluorescence measurements of biological aerosols have also been reported in more detail for tropical rainforest air (Prenni et al., Citation2009; Gabey et al., Citation2010; Pöschl et al., Citation2010) as well as in two urban European locations (Huffman et al., Citation2010; Gabey et al., Citation2011).
3.3.2.2. Flow cytometry
Flow cytometry has also long been an important tool in the investigation of ambient PBAP. Among the papers that report successful applications of flow cytometry to real-time investigation of biological aerosol are those by Sincock et al. (Citation1999), Chen and Li (Citation2005) and Chen and Li (Citation2007). Fluorescent in situ hybridization (FISH) flow cytometry has been used by a number of groups to characterise airborne bacteria and aerosolised byproducts (e.g. Lange et al., Citation1997). Using a flow, cytometric system (Ho and Fisher, Citation1993) investigated Bacillus subtilis bacterial spores, and Prigione et al. (Citation2004) developed a flow cytometer that could be selectively applied to the investigation of airborne fungi.
3.3.2.3. LIght Detection And Ranging (LIDAR) and remote sensing
The LIght Detection And Ranging (LIDAR) technique has been utilised to quickly and remotely monitor the presence of PBAP over a larger spatial range. A LIDAR system (Evans et al., Citation1994) was operated to determine its sensitivity to aerosolised Bacillus subtilis spores, and subsequent efforts have investigated LIDAR performance for PBAP detection in more detail (e.g. Simard et al., Citation2004; Glennon et al., Citation2009). The detection of fluorescent ambient aerosol was reported for the first time using a LIDAR system (Immler et al., Citation2005), attributing the signals to a combination of PAH-containing particles and/or PBAP. Furthermore, it was shown (Sassen, Citation2008) that pollen can generate strong laser depolarisation in LIDAR backscatter during Alaskan springtime measurements and suggested that pollen plumes may be mistaken for upper cirrus clouds and therefore introduce important errors into identifying aerosols in the atmosphere. Atmospheric optical phenomena caused by pollen also have been reported,Footnote6 and will be discussed briefly in Section 5.
3.3.2.4. Fluorescent and Raman spectroscopy
Fluorescent properties of collected biological aerosols have been widely used for assessment of concentrations and properties of ambient PBAP. Fisar et al. (Citation1990) showed that fluorescence detected from dyed biological aerosol particles could be effectively scaled to more traditionally measured CFUs. Raman spectroscopy has been used for investigation and characterisation of individual pollen grains in a number of different manners by various groups (Laucks et al., Citation2000; Boyain-Goitia et al., Citation2003; Kano and Hamaguchi, Citation2006; Schulte et al., Citation2008). A particularly interesting technique utilised fluorescence measured from particles impacted on a surface as a pre-selector, before Raman spectroscopy enabled bacterial identification in more detail (Rosch et al., Citation2006). Surface-enhanced Raman spectroscopy has also been employed to sensitively detect and characterise pollen and bacteria after injection into a silver suspension (Sengupta et al., Citation2005, 2006, Citation2007). Raman microscopy has been utilised by Ivleva et al. (Citation2005) for the chemical investigation and discrimination of ambient PBAP. They used a combination of Raman microscopy with multivariate analyses to characterise sampled pollen with the goal of differentiating between allergenic species.
3.3.2.5. Additional optical methods
While the classes of optical techniques mentioned have been most commonly utilised for biological aerosol analysis, a review of the literature highlights the virtually endless list of techniques that have been and could be applied. Among notable additional efforts include the use of X-ray fluorescence (Pepponi et al., Citation2004) and total internal reflection fluorescence microscopy (TIRFM) (Axelrod, Citation2008) for the investigation of biological aerosol properties. Infrared vibrational spectroscopy has also been widely used for pollen identification (Pappas et al., Citation2003; Gottardini et al., Citation2007; Dell'Anna et al., Citation2009; Zimmermann, Citation2010). Scanning electron microscopy (SEM) has been particularly useful at the investigation of PBAP, allowing a close look at the morphology and surface of particles (Karlsson and Malmberg, Citation1989; Wittmaack et al., Citation2005; Coz et al., Citation2010; Pöschl et al., Citation2010; Gilardoni et al., Citation2011).
A recent study has applied Scanning Transmission X-ray Microscopy with Near-Edge X-ray Absorption Fine Structure (STXM-NEXAFS) spectroscopy to the study of PBAP (Pöhlker, personal communication). This technique allows the structural examination of particles with a resolution down to about 30 nm, combined with spatially resolved chemical characterisation. It is suitable for the determination of the major elements, carbon, nitrogen and oxygen, as well as some minor elements, e.g. iron and chlorine. In addition to the measurement of elemental abundances, it can also yield information on the bonding state of the various elements. Using STXM-NEXAFS, Pöhlker (personal communication) could show that the coarse fraction of the Amazonian aerosol consisted predominantly of intact PBAP.
3.3.3. Non-optical methods
Several reviews of modern techniques of biological aerosol detection and analysis have been published (Spurny, Citation1994; Ho, Citation2002; Douwes et al., Citation2003; Lim et al., Citation2005; Kuske, Citation2006) and so, again, only highlights of important classes of non-optical modern techniques will be discussed here.
3.3.3.1. Mass spectrometry
Advances in mass spectrometry (MS) have been utilised in many areas of physical and biological science in the last decades to provide detailed information about chemical composition. The analysis of biological aerosols has been no different, and many different MS techniques have been employed for PBAP characterisation. Matrix-Assisted Laser Desorption Ionization Time-Of-Flight (MALDI-TOF) has been successfully applied in several PBAP studies (Kim et al., Citation2005; van Wuijckhuijse et al., Citation2005; Kleefsman et al., Citation2008; Russell, Citation2009). Parker et al. (Citation2000) developed an instrument that utilised ion-trap mass spectrometry for detection and analysis of pollen, bacteria and other aerosol types. Stowers et al. (Citation2006) demonstrated the utility of laser-induced fluorescence as a pre-selector before more detailed MS identification. Lawrence Berkeley National Lab has also utilised a similar concept for its biological aerosol mass spectrometry (BAMS) instrument (e.g. Fergenson et al., Citation2004) and has deployed the instrument in a number of field locations. Other real-time MS instruments designed primarily for other aerosol-related detection purposes have also been applied to PBAP characterisation. For example, an Aerodyne Aerosol Mass Spectrometer (AMS) was deployed in Amazonia, Brazil and was able to detect molecular ion markers in submicron aerosol consistent with the presence of biological aerosols (Chen et al., Citation2009; Pöschl et al., Citation2010; Schneider et al., Citation2011). Pratt et al. (Citation2009) utilised an Aerosol Time-of-Flight Mass Spectrometer (ATOFMS) to detect the presence of biological aerosols as ice nuclei during some flights of an aircraft campaign in the Western United States. While not technically a mass spectrometer, both Dworszanski (Citation1997) and Snyder et al. (Citation2001) report development of automated instruments that utilise pyrolysis-gas chromatography in front of an ion mobility spectrometer for rapid detection of biological aerosol particles in the field.
3.3.3.2. Breakdown spectroscopy
Many groups have also utilised different types of breakdown spectroscopy (BS) to identify elemental composition as means of biological aerosol detection and analysis, although these have not been as widely applied to ambient measurements. Laser-induced breakdown spectroscopy (LIBS) has easily been the most common form of BS method and has been employed to characterise pollen (Boyain-Goitia et al., Citation2003), and fungal spores (Hybl et al., Citation2003), bacteria (Morel et al., Citation2003) and a variety of biological aerosol particle types (Samuels et al., Citation2003). Other forms of elemental analysis such as spark-induced breakdown spectroscopy (SIBS), particle-induced X-ray emission (PIXE) and various forms of combustion analysis have also been utilised for laboratory study of biological aerosol (Sarantaridis and Caruana, Citation2010; Schmidt and Bauer, Citation2010).
3.3.3.3. Miscellaneous non-optical-methods
As has been the case for recently developed optical methods of biological aerosol analysis, a large variety of non-optical techniques of chemical and physical analysis have been employed. Bioluminescence and chemiluminescence have been extensively explored for potential analytical benefits, and much of this effort has focused on the detection of ATP within collected aerosol (e.g. Lee et al., Citation2008; Seshadri et al., Citation2009; Yoon et al., Citation2010). Electrochemical, immunochemical and immune biological methods have also been investigated for ambient PBAP characterisation (Rishpon et al., Citation1992; Spurny, Citation1994; Sarantaridis and Caruana, Citation2010; Schmidt and Bauer, Citation2010). It should be noted here that complementary instruments such as biological aerosol particle concentrators have also helped enable analyses by a variety of instruments (e.g. Pan et al., Citation2004).
4. Atmospheric relevance
4.1. Atmospheric transport of biological particles
Particles in the atmosphere, both biological and non-biological, are transported primarily together with air currents, as well as vertically downwards by gravitational sedimentation and inside of airborne water droplets and ice crystals. Particles are removed from the air either by sedimentation and deposition onto the ground and plant or other surfaces, or through washout by precipitation. Depending on their size and aerodynamic properties, the average residence time of various biological particles in the atmosphere can range from less than a day to a few weeks. We do not attempt a complete review of atmospheric transport processes and their particular impact on biological aerosols here, but only give a brief overview of some key research areas. Existing reviews cover aspects of this topic in more detail (Gregory, Citation1973; Niklas, Citation1985; Aylor, Citation1986; Nagarajan and Singh, Citation1990; Brown and Hovmøller, Citation2002).
Dry deposition (sedimentation and interception/impaction onto surfaces) is the most important removal mechanism for particles tens of microns in diameter or larger. Dry deposition rates are characterised primarily by the particle's aerodynamic diameter (diameter of a spherical particle of unit density with the same terminal velocity in air as the particle in question (e.g. Hinds, Citation1999)). Because biological particles often have complex structure (rough surfaces, internal pores and asymmetric shape), their physical and aerodynamic sizes can differ significantly. In particular, certain fungal spores, pollen and seeds are adapted to be aerodynamically buoyant, to promote long-range airborne transport. Observations show that the aerodynamic diameter may be either larger or smaller than the physical diameter, with considerable variation between species (Gregory, Citation1973; , p. 17; Madelin and Johnson, Citation1992; Reponen et al., Citation1998; Reponen et al., Citation2001), and microorganisms may sometimes be present in the atmosphere in clumps or attached to other particles (Lacey, Citation1991; Tong and Lighthart, Citation2000).
For particles ~0.1–10 µm in diameter, washout by falling precipitation is the most efficient removal mechanism. A single rain event (even a slight drizzle) can efficiently remove a large percentage of particles from the air in many circumstances (McDonald, Citation1962).
In spite of limited differences related to buoyancy and IN activity, biological particles can be expected, to a good first approximation, to be transported in the atmosphere similarly to mineral dust and are treated similarly to dust in most model studies of atmospheric dispersion. Desert dust particles of a similar size to bacteria-carrying particles are well known to be transported over long distances, particularly during intermittent dust storms that are visible to satellites. Biological particles, undistinguishable to satellites, can be transported over long distance in a similar fashion, as is borne out by case studies showing marked changes in the concentration and composition of airborne microorganisms during some dust transport events (e.g. Bovallius et al., Citation1978; Prospero et al., Citation2005; Jeon et al., Citation2011); global atmospheric model simulations produce similar results (Burrows et al., Citation2009b; Wilkinson et al., 2012).
A wide variety of approaches have been used for modelling the transport of biological particles on different spatial scales: Gaussian plume models (distances up to ~100 m; Ganio et al., Citation1995; Skelsey et al., Citation2008), Lagrangian stochastic models (Jarosz et al., Citation2004), regional-scale models (Helbig et al., Citation2004; Sofiev et al., Citation2006) and global climate and transport models (Burrows et al., Citation2009b; Wilkinson et al., 2012). Most studies of the airborne dispersal of biological particles have focused on the spread of specific human, animal and plant diseases on the local or regional scale (Donaldson et al., 1982; Gloster et al., Citation1982; Aylor et al., Citation2003; Isard et al., Citation2005). Others have addressed the potential for dispersal and cross-fertilization of GM organisms (Aylor et al., Citation2003; Jarosz et al. Citation2004) and seed dispersal (Nathan et al., Citation2002).
4.2. PBAP as cloud condensation nuclei
Aerosol particles form the nucleus for the condensation of cloud droplets, and their number and properties influence cloud microphysical properties. How active particles are as cloud condensation nuclei (CCN) depends on their size and hygroscopicity (Petters and Kreidenweis, Citation2007). Primary biological aerosol particles are generally assumed to be efficient CCN, provided that their surfaces are wettable (Andreae and Rosenfeld, Citation2008; Ariya et al., Citation2009). It has been suggested that the largest PBAP (e.g. pollens grains) may act as the so-called ‘giant CCN’, i.e. they may form cloud droplets at lower supersaturations than most other aerosol particles and quickly grow to large droplet sizes, thereby facilitating rain formation (Dingle, Citation1966; Möhler et al., Citation2007; Pope, Citation2010).
lists measurements of hygroscopic growth of bacteria, fungal spores, pollen and algal exudate below water saturation and shows critical supersaturations that have been determined for some bacterial species and algal exudate (exopolymer secretions, EPS). Below the saturation point of water, hygroscopicity is quantified by the diameter increase relative to the dry diameter (‘diameter growth factor’) or by the mass of the water taken up at a certain relative humidity (RH) compared to the dry mass. The diameter growth factors (GF) observed for bacteria and fungal spores are usually modest (GF ~1.05 to 1.3 at RH = 98%) compared to those of inorganic salt particles such as sodium chloride (GF ~1.65 at RH = 98%) (Lee et al., Citation2002). Pollen grains usually show no or only little increase in geometric size at increasing relative humidities, but can take up substantial amounts of water, e.g. up to three times their dry weight at RH = 95% (Diehl et al., Citation2001). Critical supersaturations for bacteria have been reported over a wide range between 0.07 and more than 2% (Franc and DeMott, Citation1998; Bauer et al., Citation2003). Algal exudate, which is mixed into sea spray particles, reduces the hygroscopicity and CCN activity relative to artificial seawater devoid of exudate (Wex et al., Citation2010; Fuentes et al., Citation2011).
Table 5. Compilation of laboratory measurements of the hygroscopic properties of biological particles (n.a.=data not available)
4.3. PBAP as ice nuclei
Cloud water droplets do not freeze directly at 0°C but can remain in a supercooled liquid state down to temperatures of approximately −38°C. At higher temperatures, aerosol particles are required as ice nuclei to initiate ice formation via heterogeneous freezing, i.e. the formation of ice germs by the aid of crystal-like structure elements or other the so-called active sites on the particle surface. If an ice nucleus (IN) is contained inside a liquid droplet when initiating freezing, the process is termed ‘immersion freezing’. ‘Condensation freezing’ is a special case of immersion freezing, in which the CCN of a cloud droplet acts as an IN during the condensational growth phase. If a supercooled droplet collides with an aerosol particle and freezes as a result of the collision, one speaks of ‘contact nucleation’. On dry particles (‘deposition nucleation’), ice can also form directly from the vapour phase (Pruppacher and Klett, Citation1997). Freezing of a relatively small number of cloud droplets can trigger glaciation, i.e. turning a whole cloud or a region within a cloud into pure ice. Glaciation is driven by the different values of the saturation vapour pressure over supercooled liquid water and over ice, which leads to depositional growth of the ice crystals at the expense of evaporating droplets once the relative humidity falls below water saturation (Wegener-Bergeron-Findeisen process, Findeisen, Citation1938). This process is connected to the formation of large crystals that tend to fall out as precipitation. Furthermore, ice-multiplication processes can occur, in which additional ice crystals are produced from existing ice crystals, for example due to the formation of ice splinters during riming of ice particles (Hallett and Mossop, Citation1974).
In numerous studies, it has been established that mineral dust particles are relatively efficient ice nuclei. At lower temperatures, soot particles also can nucleate ice. Interestingly, the most active IN (those nucleating ice at the highest subzero temperatures) discovered so far are of biological origin. Overviews of biological ice nucleation measurements and discussions of their possible implications are given in previous reviews (Lee et al., Citation1995; Szyrmer and Zawadzki, Citation1997; Möhler et al., Citation2007; Delort et al., Citation2010; DeMott and Prenni, Citation2010). Here, we summarise the observational basis of biological ice nucleation seen in laboratory experiments and, as far as available, in the atmosphere, including both, historical and recent measurements. With the help of modelling studies, the observations are put into the context of typical atmospheric concentrations and atmospherically relevant processes.
4.3.1. Laboratory studies of ice nucleation active biological particles
Since the first ice-nucleating biological particles were discovered, numerous microorganisms have been screened for ice nucleation activity. A selection of results is summarised in . Listed are the highest temperatures at which ice nucleation was observed, which are often cited as key findings of the experiments. Equally important is the fraction of ice-nucleating particles to total particles because often the apparent freezing onset depends on the particle concentration (Yankofsky et al., Citation1981). For many biological species, only a small fraction of the total particles (i.e. in the case of bacteria: cells) actually nucleates ice, even at relatively low temperatures. Nevertheless, the entire species or strain is referred to as ‘ice nucleation active’ (INA). The IN number fractions at different temperatures, as far as they are available from the experiments in , are displayed in .
Fig. 5. Ice nucleating number fraction f IN at the observed IN onset and maximum activity temperatures from the experiments listed in Table 6. For comparison, f IN data for immersion freezing on mineral dust (natural soil samples, median diameters of 0.2–1 µm) are included (M. Niemand, personal communication).
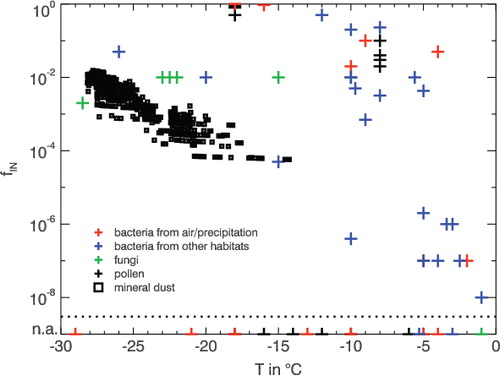
Table 6. Compilation of laboratory measurements of the IN properties of biological particles (n.a.=data not available)
Many studies listed in used biological particles sampled not from the atmosphere, but from plants, lichens, soils, ocean water and insect guts. Some of them were cultivated under favourable conditions being tested for IN activity. It can thus be questioned whether the investigated samples are representative of biological aerosols. Most studies employ the so-called droplet freezing assay with rather large volumes (several µl) of particle suspensions (thus testing for immersion freezing). Only a few experiments employ atmospherically relevant droplet sizes (e.g. Möhler et al., Citation2008) or test for other nucleation modes (e.g. Levin and Yankofsky, Citation1983).
Among bacterial ice nucleators, Pseudomonas syringae is the most common. Other INA species include Pseudomonas fluorescens and Erwinia herbicola. In general, not all strains within a species are INA, and also within the INA strains, the fraction of ice-nucleating cells varies significantly (Hirano and Upper, Citation1995). Active number fractions between 10−8 and close to 1 are reported, and the onset of ice nucleation can be seen at temperatures between −2 and −10°C ( and ). Species with intermediate IN activity begin ice nucleation only at lower temperatures (e.g. Mortazavi et al., Citation2008).
A number of fungi (both free living and lichen fungi) were found to nucleate ice at temperatures comparable to INA bacteria (), some even at −1°C. Fungal spores, which are more likely to become airborne than other parts of the fungal thallus, have been observed to nucleate ice at lower temperatures (−10 to − 28.5°C, Jayaweera and Flanagan, Citation1982; Iannone et al., Citation2011). Lichen photobionts (algae or cyanobacteria) are in general less efficient IN than the corresponding lichen fungi (Kieft and Ahmadjian, Citation1989).
Pollen of various plant species have been shown by a number of studies (Diehl et al., Citation2001, Citation2002; von Blohn et al., 2005) to nucleate ice starting at temperatures around −10°C (). Very high active fractions (up to 1.0) can be reached at −18°C (Diehl et al., Citation2001). Other biological materials, such as algae, leaf litter and plankton initiate freezing at temperatures comparable to bacteria and fungi. IN numbers are usually given in relation to the mass of the bulk material, which is of only limited relevance for atmospheric applications, where particle surface area and number fractions matter.
Also shown in are ice nucleating number fractions for mineral dust particles of atmospherically relevant sizes (median diameters of 0.2–1 µm) in the immersion freezing mode (Niemand, personal communication). The observed ice nucleating fractions for dust range from about 10−4 to 10−2 at temperatures between −15 and −28°C. Larger dust particles (not shown) exhibit larger ice-nucleating number fractions because ice nucleation is proportional to the particle surface area. CitationConen et al. (2011) showed that soil with high organic content exhibits higher IN activity than montmorillonite and postulate that ice active proteins in the soil from fragments of decaying biological material are likely the cause. The atmospheric importance of different INA materials depends not only on their ice-nucleating fractions that are shown in but also on their number concentrations at cloud altitudes and the prevailing environmental conditions (e.g., temperature and humidity).
4.3.2. Observation of biological ice nuclei in the atmosphere
The plant pathogenic bacteria Pseudomonas are frequent in the biosphere, and Pseudomonas bacteria are also frequently found as airborne particles or in cloud droplets (e.g. Fuzzi et al., Citation1997; Amato et al., Citation2005, 2007a, 2007c). However, in these studies, the ice nucleation activity of the isolated Pseudomonas strains was not determined. Little is known about the atmospheric abundance of INA lichen and fungal spores. Pollen, for which ice nucleation activity seems to be a common property of many species, can reach peak concentrations on the order of several per litre during the pollination season (Vogel et al., Citation2008).
Direct observations of the involvement of biological particles in cloud ice and precipitation formation are difficult to obtain. The presence (or absence) of INA biological particles in the air, cloud condensate or precipitation can give some indications on their role in the atmosphere. Such observations are discussed in the following section.
Kumai (Citation1961) found indirect evidence of the possible involvement of biological particles in atmospheric ice nucleation by microscopically identifying bacteria at the centre of 3 out of 307 examined snow crystals. In the pioneering studies by Maki and Willoughby (Citation1978) and Sands et al. (Citation1982), ice-nucleating bacteria were isolated from rain and snow. Some were even sampled at altitudes up to 2500 m above ground (Sands et al., Citation1982). Jayaweera and Flanagan (1982) and Lindemann et al. (1982) were the first to determine the ice nucleation activity of biological particles sampled directly from the atmosphere. In the Arctic, two types of bacteria (an unidentified strain and a Pseudomonas strain) and five fungal spores were observed with moderate to high ice nucleation activity (Jayaweera and Flanagan, Citation1982). Lindemann et al. (Citation1982) found significant concentrations of INA bacteria at 10°C above different plant canopies and bare soil and identified them as Pseudomonas syringae and Erwinia herbicola. The INA bacteria accounted for 0–4% of the total bacteria. Constantinidou et al. (Citation1990) isolated Pseudomonas syringae from rainwater and aerosol samples over a soy bean field and tested them as INA at −5°C. Recently, Morris et al. (Citation2008) studied Pseudomonas syringae strains isolated from rain, snow, alpine streams, lakes and wild plants and found that all the strains isolated from snow showed ice nucleation activity at −2 to −5°C, while this property was rare among the other strains. However, other samples of bacteria isolated from precipitation exhibited only moderate ice nucleation activity (Ahern et al., Citation2007; Mortazavi et al., Citation2008).
Christner et al. (Citation2008a Citationb) demonstrated that snow and rain samples from around the world contain ice nuclei active at temperatures above −10°C. A majority of these ice nuclei was found to be inactivated through heat treatment (at 95°C) and/or lysozyme digestion, indicating a probable bacterial and/or proteinaceous origin. The observed concentrations of warm-temperature ice nuclei ranged between below 10 and several hundred per litre of snow/rain water, which is much lower than the estimated numbers of hydrometers per litre precipitation (Diehl and Wurzler, Citation2010; Hoose et al., Citation2010a). Ice nuclei sampled from the urban atmosphere (Henderson-Begg et al., Citation2009), which were also active at temperatures above −10°C, were not deactivated by lysozyme treatment and heating to 60 °C (but by heating to 90 °C). This was interpreted as an indication that these ice nuclei were not bacterial, but rather from lichen or fungi. Also, ice nuclei concentrations at 10°C in air and snow at a high-elevation mountain site did not correlate with the relative abundances of well-known INA bacteria species such as Pseudomonas syringae and Erwinia herbicola (Bowers et al., Citation2009). It was suggested that ice nucleation at this site was due to other bacterial species, varying expression of the bacterial ice-nucleation capacities, or other biological particles.
It can be expected that biological ice nuclei are particularly abundant in regions with active vegetation. Prenni et al. (Citation2009) investigated aerosol particles sampled above the canopy in the Amazon forest and inferred that the majority of the ice nuclei at −25°C and warmer were of biological origin (but absolute concentrations of ice nuclei at these temperatures were low, on the order of 1–2 l−1). Ice nuclei at colder temperatures were in large part contributed by mineral dust. At cirrus cloud altitude, Pratt et al. (Citation2009) observed a case of co-occurrence of biological material with mineral dust after long-range transport: From mass-spectroscopic measurements of ice crystal residues in wave clouds, 50% mineral dust particles and 33% biological particles were identified. But, only in one out of many research flights was a measurable fraction of biological material observed.
In summary, the presently available observations do not give a consistent picture. The strongest indications of ice-nucleation activity in clouds are available for bacterial or fungal ice nucleators. Quantitative measurements are unfortunately rare and cover only anecdotal events. In particular, more studies quantifying the atmospheric concentrations of INA biological particles in comparison to other ice-nucleating agents, such as mineral dust and soot, would be desirable.
4.3.3. Modelling of biological ice nuclei
Numerical simulations of the effects biological ice nuclei in clouds began with Levin et al. (Citation1987), who studied the efficiency of cloud seeding with INA bacteria for precipitation enhancement in a 1.5-dimensional cloud model. Later, Diehl and Wurzler (Citation2004); Diehl et al. (2006) parameterised freezing of droplets containing or colliding with unknown amounts of different biological ice nucleators for a parcel model with detailed microphysics. In a follow-up study (Diehl and Wurzler, Citation2010), the freezing rates were scaled with typical fractions of droplets containing bacteria, mineral dust and soot particles, respectively, and it was shown that typical bacteria concentrations yielded much lower ice crystal concentrations than typical mineral dust concentrations. Ariya et al. (Citation2009), in studies with a 1.5-dimensional model of a convective cloud, and employing INA biological aerosol concentrations of less than 1 l−1, pointed out that even such low IN concentrations can trigger cloud glaciation through ice-multiplication processes. The comprehensive empirical ice nucleation parameterisation by Phillips et al. (Citation2008) includes biological material as part of the ‘insoluble organic’ particles. In a cloud-system resolving model, cloud micro- and macrophysical properties, including precipitation, were sensitive to variations of the ‘insoluble organic’ aerosol by a factor of 100 (Phillips et al., Citation2009). The first global model studies of the impact of biological particles by Hoose et al. (Citation2010a), Hoose et al. (Citation2010b) used simple emission parameterisations for bacteria, fungal spores and pollens to simulate their concentration in the atmosphere. The freezing rates were derived from classical nucleation theory and laboratory data (Chen et al., Citation2008a). The average contribution of biological particles to heterogeneous ice nucleation in mixed-phase clouds (integrated over all temperature regions in the troposphere) was calculated to be only 0.00001%, with an uppermost estimate of 0.6%. Instead, atmospheric ice nucleation was found to be dominated by mineral dust, which is also in agreement with recent observations (CitationChoi et al., 2010; DeMott and Prenni, Citation2010; CitationDeMott et al., 2010; CitationKlein et al., 2010). The contribution of biological particles to ice nucleation in relatively warm cloud layers (above −10°C) might be higher but was not quantified in these studies.
In summary, the effect of biological particles as ice nuclei on clouds and precipitation is strongly dependent on the atmospheric concentrations of INA species. Most likely, these concentrations exhibit strong temporal and spatial variations, and any effect on clouds is therefore likely seasonal and local in nature. However, more quantitative observations of INA biological particles in the atmosphere are necessary to allow better estimates of their effects on clouds.
4.4. Optical properties
The absorption and scattering of radiation by aerosol particles are important physical properties that influence regional and global radiation budgets. Better understanding of the effect that natural aerosols have on the atmosphere is necessary to constrain effects that anthropogenic influence may have on global climate. Because PBAP can be a major fraction of aerosol number and surface area in certain locations, it is possible that they may also affect climate forcing both directly (by absorbing or scattering radiation) and indirectly (through cloud processes). Certain fungal spores and other PBAP classes can be highly coloured and absorbing, which may increase their direct influence on the surrounding atmosphere (e.g. Adams et al., Citation1968; Troutt and Levetin, Citation2001). However, there have been very few studies estimating the direct effect of PBAP on climate, in part because geographically or temporally comprehensive PBAP measurements are not yet available. A theoretical description of the interaction between electromagnetic radiation and PBAP is difficult because the Mie theory is only valid for spheres, and thus many biological aerosol particles cannot be well described (Bohren and Huffman, Citation1983).
Some work has been done to directly measure certain optical properties of PBAP that could be relevant to the investigation of atmospheric aerosol. Spankuch et al. (Citation2000) showed that down-welling infrared flux was significantly increased when pine pollen concentrations were higher, suggesting that emissions of certain pollens may cause localised atmospheric warming events. Several groups have shown that pollen can cause visible coronae around the sun and moon and could therefore influence local solar radiation properties (Parviainen et al., Citation1994; Trankle and Mielke, Citation1994; Mims, Citation1998; Schneider and Vollmer, Citation2005).
Most work involving the optical properties of PBAP has used detailed physical measurements in the laboratory and, as such, has only limited obvious application to the atmosphere at large. Surbek et al. (Citation2009) characterised the elastic light scattering of a number of pollen species. Several groups have made detailed measurements of the polarisation and scattering properties specific to bacteria and other biological aerosol types (Bickel et al., Citation1976 and references therein; Bohren and Huffman, Citation1983; Vandemerwe et al., Citation1989). Gurton et al. (Citation2001) measured infrared extinction by bacterial spores. Harding and Johnson (Citation1984) measured quasi-elastic light scattering, and Gittins et al. (Citation1999) measured infrared absorption of Bacillus subtilis bacteria. Yabushita and Wada (Citation1985) performed IR and UV absorption measurements of E. coli and yeast organisms.
In view of the much higher abundance of mineral dust discussed by CitationHoose et al. (2010b), the influence of biological material on aerosol optical properties may be relatively small on global scales. However, on regional scales PBAP may sometimes have a substantial influence on the total scattering and absorption of light by aerosols. Using chemical tracers and multivariate statistical analysis, Guyon et al. (Citation2004) showed that up to 66% of aerosol mass and 47% of the light absorption in air over the Amazon Rainforest were attributable to biogenic particles during the wet season. During the wet-to-dry transition period, biogenic particles still accounted for up to 35% of light absorption, even though there was already a substantial amount of biomass burning aerosol present. These results are consistent with many earlier measurements (e.g. Artaxo et al., Citation1988 Citation1998), which showed that biogenic aerosol makes up a large fraction of the Amazonian aerosol mass burden, especially in the coarse fraction. In the absence of definitive identification, earlier studies assumed that it consisted mostly of PBAP (Andreae and Crutzen, Citation1997). This was confirmed with less uncertainty using multiple techniques by Pöschl et al. (Citation2010), who showed that PBAP accounted for ~67% of the aerosol mass (or volume) of all particle sizes sampled above the rainforest in a remote section of Amazonia, Brazil during a time period of very low influence from airborne mineral dust or anthropogenic pollution. In addition, biological organisms or material may often be attached to mineral dust particles and might also influence the optical properties of these combined particles. Thus, PBAP need to be considered when modelling local and regional optical properties above and downwind of biologically active regions. Additional direct atmospheric measurements of light absorption and scattering by biogenic particles will likely be necessary to help provide suitable model inputs and such measurements are strongly recommended.
5. Conclusions and outlook
Scientific investigations of biological aerosol particles in the atmosphere have a long history going back to the nineteenth century. Since then, the topic has attracted attention in various different research areas. In the last decade, there has been a surge in interest within the atmospheric community based partly on the development of new measurement techniques and on studies indicating that PBAP may play an important role as ice nuclei influencing the formation of clouds and precipitation.
Depending on the character of the biological particle of interest, as well as on the scope of the scientific questions asked, manifold methods are available to study PBAP. Thus, we have included an overview of current methods for sampling and analysis of biological aerosols including traditional and modern techniques.
Further studies of biological aerosols will lead to a better understanding of their role in climate and atmospheric processes and will also help to improve understanding of their impacts on humans. Recently developed and emerging techniques for sampling and analysing airborne biological particles have bolstered efforts to understand the properties of ambient PBAP. However, for better comparison among different datasets, sampling and analysis techniques have to be standardised. Efficient and reliable analytical techniques must be developed for the identification and quantification of PBAP as well as for the determination of the abundance and diversity of PBAP and their seasonal variation on regional and global scales (atmospheric biogeography).
One of the main influences of PBAP on climate and atmosphere is through the capability of certain PBAP to function as excellent ice nuclei. Future research should thus concentrate on the determination of actual emission rates and optical properties of PBAP and to link results from laboratory experiments concerning the IN ability of PBAP to atmospheric measurements.
Global modelling is one of the rising methods to track and understand the worldwide global distribution of PBAP. Atmospheric models can estimate the atmospheric effects of IN active biological aerosol particles on clouds and help identify targets for experimental research. However, the models need to be constrained and confirmed by experimental data. Future research should use field and laboratory data to better constrain numerical models of PBAP sources, their transformation in the atmosphere and effects on climate. An important task is the delivery of larger and more comprehensive datasets from sampling sites worldwide and their incorporation into atmospheric models, to ground model efforts in a solid empirical foundation.
Acknowledgements
We would like to thank M. Hummel, I. Mueller-Germann, C. Pöhlker and H. Paulsen for support and the members of the bioaerosol community for stimulating discussions. This research was funded in part by the Max Planck Society, the German Research Foundation (DE1161/2-1) and the LEC Geocycles. C.H. acknowledges support by the President's Initiative and Networking Fund of the Helmholtz Association.
Notes
1A list of abbreviations can be found in the appendix.
2Originally defined by Griffith, W. D.; Jaenicke, R.; Levin, Z.; Matthias-Maser, S; Rantio-Lehtimaki, A.; Schnell, R. C.; and Sinha, M. P.
3Table 2 provides an overview of all taxonomic terms used in the review
4Provided by NCBI http://blast.ncbi.nlm.nih.gov/Blast.cgi, 28 March 2011.
5UV-APS; TSI Inc., Model 3314, St. Paul, MN
6 http://www.atoptics.co.uk/droplets/pollen1.htm, 28 March 2011
Related Research Data
References
- Adams K. F, Hyde H. A, Williams D. A. Woodlands as a source of allergens. Allergy. 1968; 23: 265–281.
- Adhikari, A, Jung, J, Reponen, T, Lewis, J. S, DeGrasse, E. C. and co-authors. 2009. Aerosolization of fungi, (1- > 3)-beta-D Glucan, and Endotoxin from materials collected in New Orleans homes after hurricanes Katrina and Rita. General Meeting of the American Society of Microbiology. Environ. Res. 109, 215–224.
- Ahern H. E, Walsh K. A, Hill T. C. J, Moffett B. F. Fluorescent pseudomonads isolated from Hebridean cloud and rain water produce biosurfactants but do not cause ice nucleation. Biogeosciences. 2007; 4: 115–124.
- Aimanianda, V, Bayry, J, Bozza, S, Kniemeyer, O, Perruccio, K. and co-authors. 2009. Surface hydrophobin prevents immune recognition of airborne fungal spores. Nature. 460: 1117–1121.
- Akers, T. G. 1969. Survival of airborne virus, phage and other minute microbes. In: Introduction to Experimental Aerobiology. (R. L.Dimmick and A. B.Akers. Wiley Interscience, New York. 296–339.
- Akers, T. G. 1973. Some aspects of airborne inactivation of viruses. In: Airborne Transmission and Airborne Infection. VIth International Symposium on Aerobiology Technical University at Ensched, Netherlands. (J. F. P.Hers and J. C.Winkler). John Wiley & Sons: New York, 73–81.
- Aller J, Kuznetsova M, Jahns C, Kemp P. The sea surface microlayer as a source of viral and bacterial enrichment in marine aerosols. J. Aerosol Sci. 2005; 30: 801–812.
- Alvarez A. J, Buttner M. P, Stetzenbach L. D. PCR for bioaerosols monitoring: sensitivity and environmental interference. Appl. Environ. Microbiol. 1995; 61: 3639–3644.
- Amann R. I, Ludwig W, Schleifer K. H. Phylogenetic identification and in-situ detection of individual microbial-cells without cultivation. Microbiol. Rev. 1995; 59: 143–169.
- Amato, P, Demeer, F, Melaouhi, A, Fontanella, S, Martin-Biesse, A. and co-authors. 2007a. A fate for organic acids, formaldehyde and methanol in cloud water: their biotransformation by microorganisms. Atmospheric Chemistry and Physics. 7: 4159–4169.
- Amato, P, Hennebelle, R, Magand, O, Sancelme, M, Delort, A. M. and co-authors. 2007b. Bacterial characterization of the snow cover at Spitzberg, Svalbard. FEMS Microbiol. Ecol. 59: 255–264.
- Amato, P, Menager, M, Sancelme, M, Laj, P, Mailhot, G. and co-authors. 2005. Microbial population in cloud water at the Puy de Dôme: implications for the chemistry of clouds. Atmos. Environ. 39: 4143–4153.
- Amato, P, Parazols, M, Sancelme, M, Laj, P, Mailhot, G. and co-authors. 2007c. Microorganisms isolated from the water phase of tropospheric clouds at the Puy de Dôme: major groups and growth abilities at low temperatures. FEMS Microbiol. Ecol. 59: 255–264.
- Andersen A. A. New sampler for the collection, sizing and enumeration of viable airborne particles. J. Bacteriol. 1958; 76: 471–484.
- Andreae M. O, Crutzen P. J. Atmospheric aerosols: biogeochemical sources and role in atmospheric chemistry. Science. 1997; 276: 1052–1056.
- Andreae M. O, Rosenfeld D. Aerosol-cloud-precipitation interactions. Part 1. The nature and sources of cloud-active aerosols. Earth Sci. Rev. 2008; 89: 13–41.
- Ariya, P. A, Sun, J, Eltouny, N. A, Hudson, E. D, Hayes, C. T. and co-authors. 2009. Physical and chemical characterization of bioaerosols – implications for nucleation processes. Int. Rev. Phys. Chem. 28: 1–32.
- Artaxo, P, Fernandes, E. T, Martins, J. V, Yamasoe, M. A, Hobbs, P. V. and co-authors. 1998. Large-scale aerosol source apportionment in Amazonia. J. Geophys. Res. 103: 31837–31847.
- Artaxo P, Storms H, Bruynseels F, van Grieken R, Maenhaut W. Composition and sources of aerosols from the Amazon basin. J. Geophys. Res. 1988; 93: 1605–1615.
- Atlas R. M, Bartha R. Microbial Ecology: Fundamentals and Applications. Benjamin/Cummings Publishing Co: San Francisco CA, 1997
- Axelrod D. Total internal reflection fluorescence microscopy. Biophysical Tools for Biologists, Vol 2: In Vivo Techniques. 2008; 89: 169–221.
- Aylor D. A framework for examining inter-regional aerial transport of fungal spores. Agric. Forest Meteorol. 1986; 38: 263–288.
- Aylor D. E. Spread of plant disease on a continental scale: role of aerial dispersal of pathogens. Ecology. 2003; 84: 1989–1997.
- Baertsch C, Paez-Rubio T. P.-R, Viau E, Peccia J. Source tracking aerosols released from land-applied Class B biosolids during high-wind events. Appl. Environ. Microbiol. 2007; 73: 4522–4531.
- Barlow D. F. The effects of various protecting agents on the inactivation of foot-and-mouth disease virus in aerosols and during freeze-drying. J. Gen. Virol. 1972; 17: 281–288.
- Baron P. A, Willeke K. Aerosol Measurement. Principles, Techniques, and Applications. Wiley Interscience: New York, 2001
- Bauer, H, Claeys, M, Vermeylen, R, Schueller, E, Weinke, G. and co-authors. 2008a. Arabitol and mannitol as tracers for the quantification of airborne fungal spores. Atmos. Environ. 42: 588–593.
- Bauer, H, Kasper-Giebl, A, Loflund, M, Giebl, H, Hitzenberger, R. and co-authors. 2002a. The contribution of bacteria and fungal spores to the organic carbon content of cloud water, precipitation and aerosols. Atmos. Res. 64: 109–119.
- Bauer, H, Kasper-Giebl, A, Zibuschka, F, Hitzenberger, R, Kraus, G. F. and co-authors. 2002b. Determination of the carbon content of airborne fungal spores. Anal. Chem. 74: 91–95.
- Bauer H, Giebl R, Hitzenberger R, Kasper-Giebl A. Airborne bacteria as cloud condensation nuclei. J. Geophys. Res. 2003; 108(D21): 4658.
- Bauer, H, Schueller, E, Weinke, G, Berger, A, Hitzenberger, R. and co-authors. 2008b. Significant contributionss of fungal spores to the organic carbon and to the aerosol mass balance of the urban atmospheric aerosol. Atmos. Environ. 42: 588.
- Baylor E. R, Baylor M. B, Blanchard D. C, Syzdek L. D, Appel C. Water-to-air transfer of virus. Science. 1977a; 198: 575–580.
- Baylor E. R, Peters V, Baylor M. B. Water-to-air transfer of virus. Science. 1977b; 197: 763–764.
- Baylor E. R, Baylor M. B. Surf-to-wind transfer of viruses. Ann. N. Y. Acad. Sci. 1980; 353: 201–208.
- Belnap, J, Büdel, B and Lange, O. L. 2003. Biological soil crusts: characteristics and distribution. In: Biological Soil Crusts: Structure, Function, and Management. (J.Belnap and O. L.Lange). Springer-Verlag:: BerlinHeidelberg, 3–29.
- Ben-Ami, Y, Koren, I, Rudich, Y, Artaxo, P, Martin, S. T. and co-authors. 2010. Transport of Saharan dust from the Bodélé Depression to the Amazon Basin: a case study. Atmos. Chem. Phys. 10: 7533–7544.
- Benbough J. E. Some factors affecting the survival of airborne viruses. J. Gen. Virol. 1971; 10: 209–220.
- Bickel W. S, Davidson J. F, Huffman D. R, Kilkson R. Application of polarization effects in light-scattering – new biophysical tool. Proc. Natl. Acad. Sci. USA. 1976; 73: 486–490.
- Bigg E. K. Brochosomes – a tracer for near-surface air. Atmos. Res. 2003; 66: 141–144.
- Bigg E. K. Sources, nature and influence on climate of marine airborne particles. Environ. Chem. 2007; 4: 155–161.
- Bigg E. K, Gras J. L, Mossop D. J. C. Wind-produced submicron particles in the marine atmosphere. Atmos. Res. 1995; 36: 55–68.
- Bigg, E. K and Leck, C. 2008. The composition of fragments of bubbles bursting at the ocean surface. J. Geophys. Res. 113: 10.3402/tellusb.v64i0.15598.
- Blanchard, D. C. 1983. The production, distribution, and bacterial enrichment of the sea-salt aerosol. In: Air-Sea Exchange of Gases and Particles. (P. S.Liss and W. G. N.Slinn), 407–454.
- Blanchard D. C. The ejection of drops from the sea and their entrichment with bacteria and other materials – a review. Estuaries. 1989; 12: 127–137.
- Blanchard, D. C and Syzdek, L. D. 1978. Problems in bubble and jet drop researches. Limnol. Oceanogr. 2: 3, 389–400.
- Blanchard D. C, Syzdek L. D. Water-to-air transfer and enrichment of bacteria in drops from bursting bubbles. Appl. Environ. Microbiol. 1982; 43: 1001–1005.
- Blanchard D. C, Syzdek L. D, Weber M. E. Bubble scavenging of bacteria in fresh-water quickly produces bacterial enrichment in airborne jet drops. Limnol. Oceanogr. 1981; 26: 961–964.
- Bohren C. F, Huffman D. R. Biological Particles. Absorption and Scattering of Light by Small Particles. New York, NY: John Wiley & Sons, 1983; 425.
- Boreson J, Dillner A. M, Peccia J. Correlating bioaerosol load with PM2.5 and PM10cf concentrations: a comparison between natural desert and urban-fringe aerosols. Atmos. Environ. 2004; 38: 6029–6041.
- Borodulin, A. I, Safatov, A. S, Shabanov, A. N, Yarygin, A. A, Khutorova, O. G. and co-authors. 2005. Physical characteristics of concentration fields of tropospheric bioaerosols in the South of Western Siberia. J. Aerosol Sci. 36: 785–800.
- Bourdillon R. B, Lidwell O. M, Thomas J. C. A slit sampler for collecting and counting air-borne bacteria. J. Hyg. 1941; 41: 197–224.
- Bovallius A, Bucht B, Roffey R, Anas P. Long-range air transmission of bacteria. Appl. Environ. Microbiol. 1978; 35: 1231–1232.
- Bowers, R. M, Lauber, C. L, Wiedinmyer, C, Hamady, M, Hallar, A. G. and co-authors. 2009. Characterization of airborne microbial communities at a high-elevation site and their potential to act as atmospheric ice nuclei. Appl. Environ. Microbiol. 75: 5121–5130.
- Bowers R. M, McLetchie S, Knight R, Fierer N. Spatial variability in airborne bacterial communities across land-use types and their relationship to the bacterial communities of potential source environments. ISME J. 2010; 5: 1–12.
- Bowers R. M, Sullivan A. P, Costello E. K, Collett J. L, Knight R, Fierer N. Sources of bacteria in outdoor air across cities in the Midwestern United States. Appl. Environ. Microbiol. 2011; 77: 6350–6356.
- Boyain-Goitia A. R, Beddows D. C. S, Griffiths B. C, Telle H. H. Single-pollen analysis by laser-induced breakdown spectroscopy and Raman microscopy. Appl. Opt. 2003; 42: 6119–6132.
- Brachman, P. S, Gabelli, V. J, Kethley, T. W, Ehrlich, R, Maltman, J. R. and co-authors. 1964. Standard sampler for assay of airborne microorganisms. Science. 144: 1295.
- Bridge P, Spooner B. Soil fungi: diversity and detection. Plant Soil. 2001; 232: 147–154.
- Brodie, E. L, DeSantis, T. Z, Parker, J. P. M, Zubietta, I. X, Piceno, Y. M. and co-authors. 2007. Urban aerosols harbor diverse and dynamic bacterial populations. Proc. Natl. Acad. Sci. USA. 104: 299–304.
- Brosseau, L. M, Vesley, D, Rice, N, Goodell, K, Nellis, M. and co-authors. 2000. Differences in detected fluorescence among several bacterial species measured with a direct-reading particle sizer and fluorescence detector. Aerosol Sci. Technol. 32: 545–558.
- Brown J. K. M, Hovmøller M. S. Aerial dispersal of pathogens on the global and continental scales and its impact on plant disease. Science. 2002; 297: 537–541.
- Bubrick P, Galun M, Frensdorff A. Observations on free-living Trebouxia De Puymaly and Pseudotrebouxia Archibald, and evidence that both symbionts from Xanthoria parientina (L.) Th. Fr. can be found free-living in nature. New Phytologist. 1984; 97: 455–462.
- Bucker H, Horneck G. Discussion of a possible contamination of space with terrestrial life. Life Sci. Space. Res. 1969; 7: 21–27.
- Büdel, B and Scheidegger, C. 1996. Thallus morphology and anatomy. In: Lichen biology. (T. H.Nash III). ich finde: Cambridge University Press, Cambridge, 37–64.
- Büdel, B, Weber, B, Kühl, M, Pfanz, H, Sültemeyer, D. and co-authors. 2004. Reshaping of sandstone surfaces by cryptoendolithic cyanobacteria: bioalkalization causes chemical weathering in arid landscapes. Geobiology. 2: 261–268.
- Burch M, Leventin E. Effects of meteorological conditions on spore plumes. Int. J. Biometeorol. 2002; 46: 107–117.
- Burchardt L, Dankowska E. Development of algae on artificial substrates used for plant cultivation. Biol. Lett. 2003; 40: 53–59.
- Burge H. A, Rogers C. A. Outdoor allergens. Environ. Health Perspect. 2000; 108: 653–659.
- Burrows, S. M, Butler, T, Jockel, P, Tost, H, Kerkweg, A. and co-authors. 2009b. Bacteria in the global atmosphere – Part 2: Modeling of emissions and transport between different ecosystems. Atmos. Chem. Phys. 9: 9281–9297.
- Burrows S. M, Elbert W, Lawrence M. G, Poschl U. Bacteria in the global atmosphere – Part 1: Review and synthesis of literature data for different ecosystems. Atmos. Chem. Phys. 2009a; 9: 9263–9280.
- Burshtein N, Lang-Yona N, Rudich Y. Ergosterol, arabitol and mannitol as tracers for biogenic aerosols in the eastern Mediterranean. Atmos. Chem. Phys. 2011; 11: 829–839.
- Butler G. W, Bailey R. W. Chemistry and Biochemistry of Herbage. New York: Academic Press, 1973
- Cabredo S, Parra A, Anzano J. Bacteria spectra obtained by laser induced fluorescence. J. Fluoresc. 2007; 17: 171–180.
- Calderon C, Lacey J, McCartney H. A, Rosas I. Seasonal and diurnal variation of airborne basidiomycete spore concentration in Mexico City. Grana. 1995; 34: 260–268.
- Campbell S. D, Jeys T. H, Le Eapen X. Bioaerosol optical sensor model development and initial validation. Proceed. SPIE. 2007; 6538: 65380P65381–65380P65389.
- Cannon R. E, Anderson S. M. Biogenesis of bacterial cellulose. Crit. Rev. Microbiol. 1991; 17: 345–447.
- Carducci A, Arrighi S, Ruschi A. Detection of coliphages and enteroviruses in sewage and aerosol from an activated sludge wastewater treatment plant. Lett. Appl. Microbiol. 1995; 21: 207–209.
- Carnelly T, Haldane J. S, Anderson A. M. The carbon acid, organic matter, and micro-organisms in air, more especially of dwellings and schools. Philos. Transact. R. Soc. Lond. B. 1887; 178: 61–111.
- Carrera M, Kesavan J, Zandomeni R, Sagripanti J. L. Method to determine the number of bacterial spores within aerosol particles. Aerosol Sci. Technol. 2005; 39: 960–965.
- Chen, Q, Farmer, D. K, Schneider, J, Zorn, S. R, Heald, C. L. and co-authors. 2009. Mass spectral characterization of submicron biogenic organic particles in the Amazon Basin. Geophys. Res. Lett. 36: L20806.
- Chen J. P, Hazra A, Levin Z. Parameterizing ice nucleation rates using contact angle and activation energy derived from laboratory data. Atmos. Chem. Phys. 2008a; 8: 7431–7449.
- Chen P. S, Li C. S. Sampling performance for bioaerosols by flow cytometry with fluorochrome. Aerosol Sci. Technol. 2005; 39: 231–237.
- Chen P. S, Li C. S. Real-time monitoring for bioaerosols – flow cytometry. Analyst. 2007; 132: 14–16.
- Chen, P. S, Lin, Q. K, Tsai, F. T and Kuo, W. C. 2008b. Airborne influenza and avian influenza viruses from long term transportation and its health effect. In: European Aerosol Conference 2008. Thessaloniki, Greece..
- Chen, P. S, Lin, Q. K, Tsai, F. T and Kuo, W. C. 2008c. Detection of influenza and avian influenza virus. In: European Aerosol Conference 2008. Thessaloniki, Greece.
- Cheng, Y. S, Barr, E. B, Fan, B. J, Hargis, P. J, Rader, D. J. and co-authors. 1999. Detection of bioaerosols using multiwavelength UV fluorescence spectroscopy. Aerosol Sci. Technol. 30: 186–201.
- Chernoff, D. I and Bertram, A. K. 2010. Effects of sulfate coatings on the ice nucleation properties of a biological ice nucleus and several types of minerals. J. Geophys. Res. Atmos. 115: 10.3402/tellusb.v64i0.15598.
- Chi M. C, Li C. S. Fluorochrome in monitoring atmospheric bioaerosols and correlations with meteorological factors and air pollutants. Aerosol Sci. Technol. 2007; 41: 672–678.
- Chihara S, Someya T. Dynamic aspects of airborne bacterial flora over an experimental area in suburb and distribution of resistant strains to antibacterial agents among airborne staphylococci. Nippon-Eiseigaku-Zasshi. 1989; 44: 756–762.
- Choi Y. S, Lindzen R. S, Ho C. H, Kim J. Space observations of cold-cloud phase change. Proc. Natl. Acad. Sci. USA. 2010; 107: 11211–11216.
- Christensen, L. S, Mousing, J, Mortensen, S, Soerensen, K. J, Strandbygaard, B. S. and co-authors. 1990. Evidence of long distance airborne transmission of Aujesky's disease (pseudorabies) virus. Vet. Rec. 127: 471–474.
- Christner, B. C, Cai, R, Morris, C. E, McCarter, K. S, Foreman, C. M. and co-authors. 2008a. Geographic, seasonal, and precipitation chemistry influence on the abundance and activity of biological ice nucleators in rain and snow. Proc. Natl. Acad. Sci. USA. 105: 18854–18859.
- Christner B. C, Morris C. E, Foreman C. M, Cai R. M, Sands D. C. Ubiquity of biological ice nucleators in snowfall. Science. 2008b; 319: 1214–1214.
- Clot B. Trends in airborne pollen: an overview of 21 years of data in Neuchâtel (Switzerland). Aerobiologia. 2003; 19: 227–234.
- Colwell R. Viable but nonculturable bacteria: a survival strategy. J. Infect. Chemother. 2000; 6: 121–125.
- Conen F, Morris C. E, Leifeld J, Yakutin M. V, Alewell C. Biological residues define the ice nucleation properties of soil dust. Atmos. Chem. Phys. 2011; 11: 9643–9648.
- Constantinidou H. A, Hirano S. S, Baker L. S, Upper C. D. Atmospheric dispersal of ice nucleation-active bacteria – the role of rain. Phytopathology. 1990; 80: 934–937.
- Courvoisier, F, Bonacina, L, Boutou, V, Guyon, L, Bonnet, C. and co-authors. 2008. Identification of biological microparticles using ultrafast depletion spectroscopy. Faraday Discuss. 137: 37–49.
- Cox C. S, Wathes C. M. Bioaerosols Handbook. Boca Raton, FL: CRC Press, 1995
- Cox R. N, Clark R. P. Natural-convection flow about the human body. Revue Generale de Thermique. 1973; 12: 11–19.
- Coz, E, Artíñano, B, Clark, L. M, Hernandez, M, Robinson, A. L. and co-authors. 2010. Characterization of fine primary biogenic organic aerosols in an urban area in the northeastern United States. Atmos. Environ. 44: 3952–3962.
- Crawford, C, Reponen, T, Lee, T, Iossifova, Y, Levin, L. and co-authors. 2009. Temporal and spatial variation of indoor and outdoor airborne fungal spores, pollen, and (1- > 3)-beta-d-glucan. Aerobiologia. 25: 147–158.
- Crook, B. 1995a. Inertial samplers: biological perspectives. In: Bioaerosols Handbook. (C. S.Cox and C. M.Wathes). CRC-Press: Boca RatonFL, 247–268.
- Crook, B. 1995b. Non-inertial Samplers. In: Bioaerosols Handbook. (C. S.Cox and C. M.Wathes). Boca RatonFL, 269–284.
- Crook B, Sherwood-Higham J. L. Sampling and assay of bioaerosols in the work environment. J. Aerosol Sci. 1997; 28: 417–426.
- De Bary A. Comparative Morphology and Biology of the Fungi, Mycetozoa and Bacteria. Oxford: Clarendon Press, 1887
- de Jong J. G. The survival of measles virus in air, in relation to the epidemiology of measles. Archiv für die Gesamte Virusforschung. 1965; 16: 97–102.
- Deguillaume, L, Leriche, M, Amato, P, Ariya, P. A, Delort, A. M. and co-authors. 2008. Microbiology and atmospheric processes: chemical interactions of primary biological aerosols. Biogeosciences. 5: 1073–1084.
- Dell'Anna, R, Lazzeri, P, Frisanco, M, Monti, F, Campeggi, F. M. and co-authors. 2009. Pollen discrimination and classification by Fourier transform infrared (FT-IR) microspectroscopy and machine learning. Anal. Bioanal. Chem. 394: 1443–1452.
- Delort, A. M, Vaitilingom, M, Amato, P, Sancelme, M, Parazols, M. and co-authors. 2010. A short overview of the microbial population in clouds: potential roles in atmospheric chemistry and nucleation processes. Atmos. Res. 98: 249–260.
- Demirev P. A, Fenselau C. Mass spectrometry for rapid characterization of microorganisms. Annu. Rev. Anal. Chem. 2008; 1: 71–93.
- DeMott P. J, Prenni A. J. New directions: need for defining the numbers and sources of biological aerosols acting as ice nuclei. Atmos. Environ. 2010; 44: 1944–1945.
- DeMott, P. J, Prenni, A. J, Liu, X, Kreidenweis, S. M, Petters, M. D. and co-authors. 2010. Predicting global atmospheric ice nuclei distributions and their impacts on climate. Proc. Natl. Acad. Sci. USA. 107: 11217–11222.
- DeSantis T. Z, Stone C. E, Murray S. R, Moberg J. P, Andersen G. L. Rapid quantification and taxonomic classification of environmental DNA from both procaryotic and eucaryotic originsusing a microarray. FEMS Microbiol. Lett. 2005; 245: 271–278.
- Després, V. R, Nowoisky, J. F, Klose, M, Conrad, R, Andreae, M. O. and co-authors. 2007. Characterization of primary biogenic aerosol particles in urban, rural, and high-alpine air by DNA sequence and restriction fragment analysis of ribosomal RNA genes. Biogeosciences. 4: 1127–1141.
- di Giorgio, C, Krempff, A, Guiraus, H, Binder, P, Tiret, C. and co-authors. 1996. Atmospheric pollution by airborne microorganisms in the city of Marseilles. Atmos. Environ. 30: 155–160.
- Diehl K, Matthias-Maser S, Jaenicke R, Mitra S. K. The ice nucleating ability of pollen: part II. Laboratory studies in immersion and contact freezing modes. Atmos. Res. 2002; 61: 125–133.
- Diehl K, Quick C, Matthias-Maser S, Mitra S. K, Jaenicke R. The ice nucleating ability of pollen – Part I: laboratory studies in deposition and condensation freezing modes. Atmos. Res. 2001; 58: 75–87.
- Diehl, K, Simmel, M and Wurzler, S. 2006. Numerical sensitivity studies on the impact of aerosol properties and drop freezing modes on the glaciation, microphysics, and dynamics of clouds. J. Geophys. Res. Atmos. 111: 10.3402/tellusb.v64i0.15598.
- Diehl K, Wurzler S. Heterogeneous drop freezing in the immersion mode: model calculations considering soluble and insoluble particles in the drops. J. Atmos. Sci. 2004; 61: 2063–2072.
- Diehl K, Wurzler S. Air parcel model simulations of a convective cloud: bacteria acting as immersion ice nuclei. Atmos. Environ. 2010; 44: 4622–4628.
- Digiovanni F, Kevan P. G, Nasr M. E. The variability in settling velocities of some pollen and spores. Grana. 1995; 34: 39–44.
- Dimmick R. L, Wolochow M. A, Chatigny A. A. Evidence for more than one diversion of bacteria with airborne particles. Appl. Environ. Microbiol. 1979; 38: 642–643.
- Dingle A. N. Pollens as condensation nuclei. Journal de Recherches Atmospheriques. 1966; 2: 231–237.
- Donaldson A. I. The influence of relative humidity on the aerosol stability of different strains of foot-and-mouth disease virus suspended in saliva. J. Gen. Virol. 1972; 15: 25–33.
- Donaldson A. I. The influence of relative humidity on the stability of foot-and-mouth disease virus in aerosols from milk and faecal slurry. Res. Vet. Sci. 1973; 15: 96–101.
- Donaldson A. I, Alexandersen S. Predicting the spread of foot-and-mouth disease by airborne virus. Revue Scientifique et Technique. 2002; 21: 569–575.
- Donaldson A. I, Ferris N. P. Airborne stability of swine vesicular disease virus. Vet. Rec. 1974; 95: 19–23.
- Donaldson, A. I, Gloster, J, Harvey, L. D and Deans, D.H. 1982. Use of prediction models to forecast and analyse airborne spread during the foot-and-mouth disease outbreaks in Brittany, Jersey and the Isle of Wight in 1981. Vet. Rec. 110: 53–57. 10.3402/tellusb.v64i0.15598.
- Donaldson A. I, Wardley R. C, Martin S, Ferris N. P. Experimental Aujeszky's disease in pigs: excretion, survival and transmission of the virus. Discov. Med. 1983; 113: 490–494.
- Dongsheng D. From the smallest virus to the biggest gene: marching towards gene therapy for duchenne muscular dystrophy. Process. Land. 2006; 6: 103–106.
- Douglas G. R. Manganese-rich coatings from Iceland. Earth Surf. Ann. Occup. Hyg. 1987; 12: 301–310.
- Douwes J, Thorne P, Pearce N, Heederik D. Bioaerosol health effects and exposure assessment: progress and prospects. Bull. WHO. 2003; 47: 187–200.
- Downie, A. W, Meiklejohn, M, Vincent, L. S, Rao, A. R, Sundara Babu, B. V. and co-authors. 1965. The recovery of smallpox virus from patients and their environment in a smallpox hospital. Int. J. Algae. 33: 615–622.
- Dubovik I. E. Migrations of aerophytic algae and their colonization on different substrata. J. Allergy. 2002; 4: 48–55.
- Durham O. C. The volumetric incidence of atmospheric allergens: I. Specific gravitiy of pollen grains. Field Anal. Chem. Technol. 1943; 14: 455–461.
- Dworzanski, J. P, McClennen, W. H, Cole, P. A, Thornton, S. N, Meuzelaar, H. L. C. and co-authors. 1997. Field-portable, automated pyrolysis-GC/IMS system for rapid biomarker detection in aerosols: a feasibility study. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin, Physikalische-mathematische Klasse. 1: 295–305.
- Ehrenberg, C. G. 1847. Passatstaub und Blutregen. Ein großes organisches unsichtbares Leben in der Atmosphäre. Pak. J. Biol. Sci. 269–460.
- El-Gamal A. D. Aerophytic Cyanophyceae (cyanobacteria) from some Cairo districts, Egypt. Atmos. Chem. Phys. 2008; 11: 1293–1302.
- Elbert W, Taylor P. E, Andreae M. O, Poschl U. Contribution of fungi to primary biogenic aerosols in the atmosphere: wet and dry discharged spores, carbohydrates, and inorganic ions. Oikos. 2007; 7: 4569–4588.
- Ellstrand N. C. Gene flow by pollen: implications for plant conservation genetics. Int. J. Biometeorol. 1992; 63: 77–86.
- Emberlin, J, Detandt, M, Gehrig, R, Jaeger, S, Nolard, N. and co-authors. 2002. Responses in the start of Betula (birch) pollen seasons to recent changes in spring temperatures across Europe. Biophys. J. 46: 159–170.
- Eng J, Lynch R. M, Balaban R. S. Nicotinamide adenine dinucleotide fluorescence spectroscopy and imaging of isolated cardiac myocytes. Aerosol Sci. Technol. 1989; 55: 621–630.
- Engling, G, Lee, J. J, Tsai, Y. W, Lung, S. C. C, Chou, C. C. K. and co-authors. 2009. Size resolved anhydrosugar composition in smoke aerosol from controlled field burning of rice straw. J. Aerosol Sci. 43: 662–672.
- Evans B. T. N, Yee E, Roy G, Ho J. Remote detection and mapping of bioaerosols. Appl. Environ. Microbiol. 1994; 25: 1549–1566.
- Fahlgren C, Hagstrom A, Nilsson D, Zweifel U. L. Annual variations in the diversity, viability, and origin of airborne bacteria. Aerobiologia. 2010; 76: 3015–3025.
- Fahlgren C, Bratbak G, Sandaa R.-A, Thyrhaug R, Zweifel U. L. Diversity of airborne bacteria in samples collected using different devices for aerosol collection. Symbiosis. 2010; 27: 107–120.
- Fahselt D. Secondary biochemistry of lichens. Microb. Ecol. 1994; 16: 117–165.
- Fang Z. G, Ouyang Z. Y, Zheng H, Wang X. K, Hu L. F. Culturable airborne bacteria in outdoor environments in Beijing, China. Appl. Environ. Microbiol. 2007; 54: 487–496.
- Fannin K. F, Vana S. C, Jakubowski W. Effect of an activated sludge wastewater treatment plant on ambient air densities of aerosols containing bacteria and viruses. Anal. Chem. 1985; 49: 1191–1196.
- Fergenson, D. P, Pitesky, M. E, Tobias, H. J, Steele, P. T, Czerwieniec, G. A. and co-authors. 2004. Reagentless detection and classification of individual bioaerosol particles in seconds. Appl. Environ. Microbiol. 76: 373–378.
- Fierer, N, Liu, Z. Z, Rodriguez-Hernandez, M, Knight, R, Henn, M. and co-authors. 2008. Short-term temporal variability in airborne bacterial and fungal populations. Mycoses. 74: 200–207.
- Filipello-Marchision V, Cassinelli C, Tulloi V, Piscozzi A. Outdoor airborne dermatophytes and related fungi: a survey in Turin. Meteorologische Zeitschrift. 1992; 35: 251–257.
- Findeisen W. Kolloidmeteorologischen Vorgänge bei der Niederschlagsbildung. Int. J. Biometeorol. 1938; 55: 121–133.
- Fisar Z, Hysek J, Binek B. Quantification of Airborne Microorganisms and Investigation of their Interactions with Nonliving Particles. 1990; 34: 189–193.
- Fitt B. D. L, McCartney H. A, Rosas I. The role of rain in dispersal of pathogen inoculum. Ann. Rev. Phytopathol. 1989; 27: 241–270.
- Franc G. D, DeMott P. J. Cloud activation characteristics of airborne Erwinia carotovora cells. J. Appl. Meteorol. 1998; 37: 1293–1300.
- Francisco D. E, Mah R. A, Rabin A. C. Acridine orange-epifluorescence technique for counting bacteria in natural waters. Transact. Am. Microsc. Soc. 1973; 92: 416–421.
- Franze T, Weller M. G, Niessner R, Pöschl U. Enzyme immunoassays for the investigation of protein nitration by air pollutants. Analyst. 2003; 128: 824–831.
- Franze T, Weller M. G, Niessner R, Poeschl U. Protein nitration by polluted air. Environ. Sci. Technol. 2005; 39: 1673–1678.
- Friedlander, S. K. 2000. Smoke, dust, and haze. Fundamentals of aerosol dynamics. Oxford University Press, Oxford, New York.
- Fröhlich-Nowoisky J, Pickersgill D. A, Després V. R, Pöschl U. High diversity of fungi in air particulate matter. Proc. Natl. Acad. Sci. USA. 2009; 106: 12814–12819.
- Fröhlich-Nowoisky J, Burrows S. M, Xie Z, Engling G, Solomon P. A, Fraser M. P, Mayol-Bracero O. L, Artaxo P, Begerow D, Conrad R, Andreae M. O, Després V. R, Pöschl U. Biogeography in the air: fungal diversity over land and oceans. Biogeosci. Discuss. 2011; 8: 7071–7096.
- Fuentes E, Coe H, Green D, McFiggans G. On the impacts of phytoplankton-derived organic matter on the properties of the primary marine aerosol – part 2: composition, hygroscopicity and cloud condensation activity. Atmos. Chem. Phys. 2011; 11: 2585–2602.
- Fuzzi, S, Andreae, M. O, Huebert, B. J, Kulmala, M, Bond, T. C. and co-authors. 2006. Critical assessment of the current state of scientific knowledge, terminology, and research needs concerning the role organic aerosols in the atmosphere, climate, and global change. Atmos. Chem. Phys. 6: 2017–2038.
- Fuzzi S, Mandrioli P, Perfetto A. Fog droplets – an atmospheric source of secondary biological aerosol particles. Atmos. Environ. 1997; 31: 287–290.
- Gabey A, Stanley W. R, Gallagher M, Kaye P. H. The fluorescence properties of aerosol larger than 0.8 (m in urban and tropical raisnforest locations. Atmos. Chem. Phys. 2011; 11: 5491–5504.
- Gabey, A. M, Gallagher, M. W, Whitehead, J, Dorsey, J. R, Kaye, P. H. and co-authors. 2010. Measurements and comparison of primary biological aerosol above and below a tropical forest canopy using a dual channel fluorescence spectrometer. Atmos. Chem. Phys. 10: 4453–4466.
- Ganio L. M, Mohr A. J, Lighthart B. A comparison between computer modeled bioaerosol dispersion and a bioaerosol field spray event. Aerobiologia. 1995; 11: 183–188.
- Gauslaa Y. Lichen palatability depends on investment in herbivore defence. Oecologia. 2005; 143: 94–105.
- Gelencér A. Carbonaceous Aerosols. the Netherlands: Springer Verlag Dordrecht, 2004
- Genitsaris S, Kormas K. A, Moustaka-Gouni M. Airborne algae and cyanobacteria: occurrence and related health effects. Front. Biosci. 2011; 3: 772–787.
- Georgakopoulos, D. G, Després, V, Fröhlich-Nowoisky, J, Psenner, R, Ariya, P. A. and co-authors. 2009. Microbiology and atmospheric processes: biological, physical and chemical characterization of aerosol particles. Biogeosciences. 6: 721–737.
- Giesecke T, Fontna S. L, van der Knaap W. O, Pardoe H. S, Pidek I. A. From early pollen trapping experiments to the Pollen Monitoring Programme. Veg. Hist. Archaeobot. 2010; 19: 247–258.
- Gilardoni, S, Vignati, E, Marmer, E, Cavalli, F, Belis, C. and co-authors. 2011. Sources of carbonaceous aerosol in the Amazon basin. Atmos. Chem. Phys. 11: 2747–2764.
- Gilbert G. S. Nocturnal fungi: airborne spores in the canopy and understory of a tropical rain forest. Biotropica. 2005; 37: 462–464.
- Gittins, C. M, Piper, L. G, Rawlins, W. T, Marinelli, W. J, Jensen, J. O. and co-authors. 1999. Passive and active standoff infrared detection of bio-aerosols. Field Anal. Chem. Technol. 3: 274–282.
- Glennon, J. J, Nichols, T, Gatt, P, Baynard, T, Marquardt, J. H. and co-authors. 2009. System performance and modeling of a bioaerosol detection lidar sensor utilizing polarization diversity. Proceed. SPIE. 7323: 0T1–0T12.
- Gloster J, Sellers R. F, Donaldson A. I. Long distance transport of foot-and-mouth disease virus over the sea. Vet. Rec. 1982; 10: 47–52.
- González-Toril, E, Amils, R, Delmas, R. J, Petit, J.-R, Komárek, J. and co-authors. 2009. Bacterial diversity of autotrophic enriched cultures from remote, glacial Antarctic, Alpine and Andean aerosol, snow and soil samples. Biogeosciences. 6: 33–44.
- Gorny, R. L, Reponen, T, Willeke, K, Schmechel, D, Robine, E. and co-authors. 2002. Fungal fragments as indoor air biocontaminants. Appl. Environ. Microbiol. 68: 3522–3531.
- Gottardini E, Rossi S, Cristofolini F, Benedetti L. Use of Fourier transform infrared (FT-IR) spectroscopy as a tool for pollen identification. Aerobiologia. 2007; 23: 211–219.
- Graber E. R, Rudich Y. Atmospheric HULIS: how humic-like are they? A comprehensive and critical review. Atmos. Chem. Phys. 2006; 6: 729–753.
- Graham, B, Falkovich, A. H, Rudich, Y, Maenhaut, W, Guyon, P. and co-authors. 2004. Local and regional contributions to the atmospheric aerosol over Tel Aviv, Israel: a case study using elemental, ionic and organic tracers. Atmos. Environ. 38: 1593–1604.
- Graham, B, Guyon, P, Maenhaut, W, Taylor, P. E, Ebert, M. and co-authors. 2003. Composition and diurnal variability of the natural Amazonian aerosol. J. Geophys. Res. 108 (D24), 4765.
- Grant R. H, Scheidt A. B, Rueff L. R. Aerosol transmission of viable virus affecting swine: explanations of an epizootic of pseudorabies. Int. J. Biometeorol. 1994; 38: 33–39.
- Green, B. J, Tovey, E. R, Sercombe, J. K, Blachere, F. M, Beezhold, D. H. and co-authors. 2006. Airborne fungal fragments and allergenicity. Med. Mycol. 44: S245–S255.
- Greenberg, L and Smith, G. W. 1922. A new instrument for sampling aerial dust. U.S. Bureau of Mines, 1–3.
- Gregory, P. H. 1973. The Microbiology of the Atmosphere. 2nd ed. Leonard Hill Books, Plant Series Monographs, Plymouth..
- Griffin D. W. Terrestrial microorganisms at an altitude of 20,000 m in Earth's atmosphere. Aerobiologia. 2004; 20: 135–140.
- Griffin D. W, Kellogg C. A. Dust storms and their impact on ocean and human health: dust in Earth's atmosphere. Ecohealth. 2004; 1: 284–295.
- Griffin D. W, Kellogg C. A, Garrison V. H, Shinn E. A. The global transport of dust. Am. Scientist. 2002; 90: 228–235.
- Griffin D. W, Westphal D. L, Gray M. A. Airborne microorganisms in the African desert dust corridor over the mid-Atlantic ridge, Ocean Drilling Program, Leg 209. Aerobiologia. 2006; 22: 211–226.
- Griffith W. D, DeCosemo G. A. L. The assessment of bioaerosols: a critical review. J. Aerosol Sci. 1994; 25: 1425–1458.
- Grini A, Zender C. S, Colarco P. R. Saltation sandblasting behavior during mineral dust aerosol production. Geophys. Res. Lett. 2002; 29: 1868–1871.
- Grinn-Gofron A, Mika A. Selected airborne allergenic fungal spores and meteorological factors in Szczecin, Poland, 2004–2006. Aerobiologia. 2008; 24: 89–97.
- Grinshpun S. A, Clark J. M. Measurement and characterization of bioaerosols. J. Aerosol Sci. 2005; 36: 553–555.
- Gross D. C, Cody Y. S, Proebsting E. L, Radamaker G. K, Spotts R. A. Distribution, population-dynamics, and characteristics of ice nucleation-active bacteria in deciduous fruit tree orchards. Appl. Environ. Microbiol. 1983; 46: 1370–1379.
- Gruijthuijsen Y. K, Grieshuber I, Stöcklinger A, Tischler U, Fehrenbach T, Weller M. G, Vogel L, Vieths S, Pöschl U, Duschl A. Nitration enhances the allergenic potential of proteins. Int. Arch. Allergy Immunol. 2006; 141: 265–275.
- Gurton K. P, Ligon D, Kvavilashvili R. Measured infrared spectral extinction for aerosolized Bacillus subtilis var. niger endospores from 3 to 13 mu m. Appl. Opt. 2001; 40: 4443–4448.
- Guyon, P, Graham, B, Roberts, G. C, Mayol-Bracero, O. L, Maenhaut, W. and co-authors. 2004. Sources of optically active aerosol particals over the Amazon forest. Atmos. Environ. 38: 1039–1051.
- Hairston P. P, Ho J, Quant F. R. Design of an instrument for real-time detection of bioaerosols using simultaneous measurement of particle aerodynamic size and intrinsic fluorescence. J. Aerosol Sci. 1997; 28: 471–482.
- Hall L. B, Bruch C. W. Procedures necessary for the prevention of planetary contamination. Life Sci. Space. Res. 1965; 3: 48–62.
- Hallar, A. G, Chirokova, G, McCubbin, I, Painter, T. H, Wiedinmyer, C. and co-authors. 2011. Atmospheric bioaerosols transported via dust storms in the western United States. Geophys. Res. Lett. 38: L17801. 10.3402/tellusb.v64i0.15598.
- Hallett J, Mossop S. C. Production of secondary ice particles during the riming process. Nature. 1974; 249: 26–28.
- Hallquist, M, Wenger, J. C, Baltensperger, U, Rudich, Y, Simpson, D. and co-authors. 2009. The formation, properties and impact of secondary organic aerosols: current and emerging issues. Atmos. Chem. Phys. 9: 5155–5236.
- Hammond, A. L. 1971. Inadvertent Climate Modification. Report of the Study of Man's Impact on Climate (SMIC). MIT-Press, Cambridge, Massachusetts.
- Hammond G. W, Raddatz R. L, Gelskey D. E. Impact of atmospheric dispersion and transport of viral aerosols on the epidemiology of influenza. Rev. Infect. Dis. 1989; 11: 494–497.
- Harding S. E, Johnson P. Quasi-elastic light-scattering studies on dormant and germinating bacillus-subtilis spores. Biochem. J. 1984; 220: 117–123.
- Harper G. J. Airborne microorganisms: survival test with four viruses. J. Hyg. (Camb). 1961; 59: 479–486.
- Harrington J. B, Metzger K. Ragweed pollen density. Am. J. Bot. 1963; 50: 532–539.
- Harrison D. E, Chance B. Fluorimetric technique for monitoring changes in level of reduced nicotinamide nucleotides in continuous cultures of microorganisms. Appl. Microbiol. 1970; 19: 446–450.
- Harrison, R. M, Jones, A. M, Biggins, P. D. E, Pomeroy, N, Cox, C. S. and co-authors. 2005. Climate factors influencing bacterial count in background air samples. Int. J. Biometeorol. 49: 167–178.
- Hawksworth, D. L, Kirk, P. M, Sutton, B. C and D. N.Pegler. 1995. Ainsworth & Bisby's Dictionary of the Fungi. CAB International, Wallingford.
- Heald C. L, Spracklen D. V. Atmospheric budget of primary biological aerosol particles from fungal spores. Geophys. Res. Lett. 2009; 36: L09806.
- Heidelberg, J. F, Shahamat, M, Levin, M, Rahman, I, Stelma, G. and co-authors. 1997. Effect of aerosolization on culturability and viability of gram-negative bacteria. Appl. Environ. Microbiol. 63: 3585–3588.
- Helbig N, Vogel B, Vogel H, Fiedler F. Numerical modelling of pollen dispersion on the regional scale. Aerobiologia. 2004; 20: 3–19.
- Henderson D. W. An apparatus for the study of airborne infection. J. Hyg. 1952; 50: 53–68.
- Henderson-Begg S. K, Hill T, Thyrhaug R, Khan M, Moffett B. F. Terrestrial and airborne non-bacterial ice nuclei. Atmos. Sci. Lett. 2009; 10: 215–219.
- Henningson E. W, Ahlberg M. S. Evaluation of microbiological aerosol samplers – a review. J. Aerosol Sci. 1994; 25: 1459–1492.
- Hensel, A and Petzhold, K. 1995. Biological and biochemical analysis of bacteria and viruses. In: Bioaerosols Handbook. (C. S.Cox and C. M.Wathes). CRS-Press: BOCA RatonFL.
- Hernandez M, Miller S. L, Landfear D. W, Macher J. M. A combined fluorochrome method for quantitation of metabolically active and inactive airborne bacteria. Aerosol Sci. Technol. 1999; 30: 145–160.
- Hill, S. C, Pinnick, R. G, Nachman, P, Chen, G, Chang, R. K. and co-authors. 1995. Aerosol-fluorescence spectrum analyzer: real-time measurement of emission spectra of airborne biological particles. Appl. Opt. 34: 7149–7155.
- Hillamo R. E, Kauppinen E. I. On the performance of the Berner low pressure impactor. Aerosol Sci. Technol. 1991; 14: 7149–7155.
- Hinds W. C. Aerosol Technology. New York: Wiley, 1999
- Hirano S. S, Baker L. S, Upper C. D. Ice nucleation temperature of individual leaves in relation to population sizes of ice nucleation active bacteria and frost injury. Plant Physiol. 1985; 77: 259–265.
- Hirano, S. S and Upper, C. D. 1995. Ecology of ice nucleation-active bacteria. In: Biological Ice Nucleation and Its Applications. (R. E.Lee, G. J.Warren and L. V.Gusta). The American Phytopathological Society: St PaulMN, 41–61.
- Hirst J. M. An automatic volumetric spore trap. Ann. Appl. Biol. 1952; 39: 257–265.
- Hirst, J. M. 1995. Introduction, Retrospect and Prospect. In: Bioaerosol Handbook. (S.Cox and C. M.Wathes). CRC-Press: Boca RatonFL, 5–14.
- Ho J. Future of biological aerosol detection. Analytica Chimica Acta. 2002; 457: 125–148.
- Ho, J and Fisher, G. 1993. Detection of BW agents: flow cytometry measurement of Bacillus Subtilis (BS) spore fluorescence. Defence Research Establishment Suffield. Suffield, Medicine Hat: AlbertaCanada, 1–34.
- Ho J, Spence M. Background Aerosol Characteristics Measured With A Fluorescence Aerodynamic Particle Sizer: Sensitivity of FLAPS Performance. Defense Research Establishment Suffield Memorandum. 1998; 1504: 1–20.
- Ho, J, Tjarnhage, T, Burke, J and Stadnyk, L. 2004. Background aerosol sampling: optical characteristics of live particles associated with thundershowers in Umea, Sweden in September 2003. In: Proceedings of 8th International Symposium on Protection Against Chemical and Biological Warfare Agents (including Bioterrorism), Gothenburg, Sweden, 2–6. June, 2004.
- Hobbie J. E, Daley R. J, Jasper S. Use of nuclepore filters for counting bacteria by fluorescence microscopy. Appl. Environ. Microbiol. 1977; 33: 1225–1228.
- Hoose, C, Kristjansson, J. E and Burrows, S. M. 2010a. How important is biological ice nucleation in clouds on a global scale?. Environ. Res. Lett. 5: 024009. 10.3402/tellusb.v64i0.15598.
- Hoose C, Kristjansson J. E, Chen J. P, Hazra A. A classical-theory-based parameterization of heterogeneous ice nucleation by mineral dust, soot, and biological particles in a global climate model. J. Atmos. Sci. 2010b; 67: 2483–2503.
- Höss M, Jaruga P, Zastawny T, Dizdaroglu M, Pääbo S. DNA damage and DNA sequence retrieval from ancient tissues. Nuc. Acid Res. 1996; 24: 1304–1307.
- Hua N.-P, Kobayashi F, Iwasaka Y, Shi G.-Y, Naganuma T. Detailed identification of desert-originated bacteria carried by Asian dust storms to Japan. Aerobiologia. 2007; 23: 291–298.
- Huffman J. A, Treutlein B, Pöschl U. Fluorescent biological aerosol particle concentrations and size distributions measured with an Ultraviolet Aerodynamic Particle Sizer (UV-APS) in Central Europe. Atmos. Chem. Phys. 2010; 10: 3215–3233.
- Hunt J, Boddy L, Randerson P. F, Rogers H. J. An evaluation of 18S rDNA approaches for the study of fungal diversity in grassland soils. Microb. Ecol. 2004; 47: 385–395.
- Hybl J. D, Lithgow G. A, Buckley S. G. Laser-induced breakdown spectroscopy detection and classification of biological aerosols. Appl. Spectroscopy. 2003; 57: 1207–1215.
- Iannone R, Chernoff D. I, Pringle A, Martin S. T, Bertram A. K. The ice nucleation ability of one of the most abundant types of fungal spores found in the atmosphere. Atmos. Chem. Phys. 2011; 11: 1191–1201.
- IGAP. 1992. The International Global Aerosol Program. Deepak Publishing, Hampton, VA.
- IInuma, Y, Engling, G, Puxbaum, H and Herrmann, H. 2009. A highly resolved anion-exchange chromatographic method for determination of saccaridic tracers for biomass combustion and primary bio-particles in atmospheric aerosol. Atmos. Environ. 43: 1367–1371.
- Immler F, Engelbart D, Schrems O. Fluorescence from atmospheric aerosol detected by a lidar indicates biogenic particles in the lowermost stratosphere. Atmos. Chem. Phys. 2005; 5: 345–355.
- Imshenetsky A. A, Lysenko S. V, Kazakov G. A. Upper boundary of the biosphere. Appl. Environ. Microbiol. 1978; 35: 1–5.
- Ingold C. T. Fungal Spores: Their Liberation and Dispersal. Clarendon Press: Oxford, 1971
- Ingold C. T. Active liberation of reproductive units in terrestrial fungi. Mycologist. 1999; 13: 113–116.
- Ingolfsdottir K. Usnic acid. Phytochemistry. 2002; 61: 729–736.
- IPCC., 2007a. Climate Change 2007: Impacts, Adaptation and Vulnerability. In:Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. (M. L.Parry, O. F.Canziani, J. P.Palutikof, P. J.van der Linden and C.E.Hanson). Cambridge University Press: CambridgeUK, 976. pp.
- IPCC., 2007b. Climate Change 2007: The Physical Science Basis. In:Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. (S.Solomon, D.Qin, M.Manning, Z.Chen, M.Marquis, K.B.Averyt, M.Tignor and H.L.Miller). Cambridge University Press: CambridgeUnited Kingdom and New York, NY, USA, 996. pp.
- Isard S. A, Gage S. H, Comtois P, Russo J. M. Principles of the atmospheric pathway for invasive species applied to soybean rust. Oral Microbiol. Immunol. 2005; 55: 851.
- Ivleva N, Niessner R, Panne U. Characterization and discrimination of pollen by Raman microscopy. J. Geophys. Res. Atmos. 2005; 381: 261–267.
- Iwami, Y, Takahashi-Abbe, S, Takahashi, N, Yamada, T, Kano, N. and co-authors. 2001. The time-course of acid excretion, levels of fluorescence dependent on cellular nicotinamide adenine nucleotide and glycolytic intermediates of Streptococcus mutans cells exposed and not exposed to air in the presence of glucose and sorbitol. Science. 16: 34–39.
- Jacobson M. Z, Streets D. G. Influence of future anthropogenic emissions on climate, natural emissions, and air quality. 2009; 114: D08118.
- Jaenicke R. Abundance of cellular material and proteins in the atmosphere. Atmos. Environ. 2005; 308: 73.
- Jaenicke, R. andMatthias, S. 1988. The primary biogenic fraction of the atmospheric aerosol. In: Aerosols and Climate. (P. V.Hobbs, and M. P.McCormick). Deepak Publishing: HamptonVA, 31–38.
- Jarosz N, Loubet B, Huber L. Modelling airborne concentration and deposition rate of maize pollen. Geophys. Res. Lett. 2004; 38: 5555–5566.
- Jato, V, Rodriguez-Rajo, F. J, Alcázar, P, De Nuntiis, P, Galán, C. and co-authors. 2006. May the definition of pollen season influence aerobiological results?. Appl. Microbiol. 22: 13–25.
- Jayaweera K, Flanagan P. Investigations on biogenic ice nuclei in the arctic atmosphere. Atmos. Environ. 1982; 9: 94–97, doi: 10.1029/GL009i001p00094.
- Jensen M. M. Inactivation of airborne viruses by ultraviolet irradiation. Science. 1964; 12: 418–420.
- Jeon, E. M, Kim, H. J, Jung, K, Kim, J. H, Kim, M. Y. and co-authors. 2011. Impact of Asian dust events on airborne bacterial community assessed by molecular analyses. Biotechnol. Prog. 45: 4313–4321.
- Jimenez, J. L, Canagaratna, M. R, Donahue, N. M, Prevot, A. S, Zhang, Q. and co-authors. 2009. Evolution of organic aerosols in the atmosphere. Aerosol Sci. Technol. 326: 1525–1529.
- Jinn T. R, Kao S. S, Tseng Y. C, Chen Y. J, Wu T. Y. Aerosol infectivity of a Baculovirus to Trichoplusia ni larvae: an alternative larval inoculation strategy for recombinant protein production. Appl. Environ. Microbiol. 2009; 25: 384–389.
- Johnson D. L, Pearce T. A, Esmen N. A. The effect of phosphate buffer on aerosol size distribution of nebulized Bacillus subtilis and Pseudomonas fluorescens bacteria. Sci. Tot. Environ. 1999; 30: 202–210.
- Jones B, Cookson J. Natural atmospheric microbial conditions in a typical suburban area. 1983; 45: 919–934.
- Jones A. M, Harrison R. M. The effect of meteorological factors on atmospheric bioaerosols concentrations – a review. Biogeosciences. 2004; 326: 151–180.
- Junge, C. E. 1963. Air Chemistry and Radioactivity. Academic Press, New York and London. 382.
- Junge K, Swanson B. D. High-resolution ice nucleation spectra of sea-ice bacteria: implications for cloud formation and life in frozen environments. Chem. Lett. 2008; 5: 865–873.
- Kaarkainen, P, Meklin, T, Rintala, H, Hyvärinen, A, Kärkäinen, P. and co-authors. 2008. Seasonal variation in airborne microbial concentrations and diversity in landfill, urban, and rural sites. Scand. J. Work Environ. Health. 36: 556–563.
- Kano H, Hamaguchi H. O. Vibrational imaging of a single pollen grain by ultrabroadband multiplex coherent anti-stokes Raman scattering microspectroscopy. Int. J. Biometeorol. 2006; 35: 1124–1125.
- Karlsson K, Malmberg P. Characterization of exposure to molds and actinomycetes in agricultural dusts by scanning electron microscopy, fluorescence microscopy and the culture method. Ecol. Lett. 1989; 15: 353–359.
- Katial R. K, Zhang Y, Jones R. H, Dyer P. D. Atmospheric mold spore counts in relation to meteorlogical parameters. Appl. Opt. 1997; 41: 17–22.
- Kawecki T. J, Ebert D. Conceptual issues in local adaptation. Opt. Exp. 2004; 7: 1224–1241.
- Kaye P. H, Barton J. E, Hirst E, Clark J. M. Simultaneous light scattering and intrinsic fluorescence measurement for the classification of airborne particles. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2000; 39: 3738–3745.
- Kaye, P. H, Stanley, W. R, Hirst, E, Foot, E. V, Baxter, K. L. and co-authors. 2005. Single particle multichannel bio-aerosol fluorescence sensor. Trend. Ecol. Evol. 13: 3583–3593.
- Kell D. B, Ryder H. M, Kaprelyants A. S, Westerhoff H. V. Quantifying heterogeneity – flow cytometry of bacterial cultures. Canad. J. Bot. –Revue Canadienne De Botanique. 1991; 60: 145–158.
- Kellogg C. A, Griffin D. W. Aerobiology and the global transport of desert dust. Microbiol. Rev. 2006; 21: 638–644.
- Kelly C. D, Pady S. M. Microbiological studies of air masses over Montreal during 1950 and 1951. 1954; 32: 591–600.
- Kepner R. L, Pratt J. R. Use of fluorochromes for direct enumeration of total bacteria in environmental-samples – past and present. Science. 1994; 58: 603–615.
- Kershaw K. A. Physiological Ecology of Lichens. Cambridge University Press, Cambridge. 1985
- Kethley T. W, Gordon M. T, Orr C. A thermal precipitator for aerobacteriology. Lichenologist. 1952; 116: 368–369.
- Kieft T. L. Ice nucleation activity in lichens. Rapid Commun. Mass Spectrom. 1988; 54: 1678–1681.
- Kieft T. L, Ahmadjian V. Biological ice nucleation activity in lichen mycobionts and photobionts. Kona Powder Particle J. 1989; 21: 355–362.
- Kim J. K, Jackson S. N, Murray K. K. Matrix-assisted laser desorption/ionization mass spectrometry of collected bioaerosol particles. Atmos. Chem. Phys. 2005; 19: 1725–1729.
- Kleefsman W. A, Stowers M. A, Verheijen P. J. T, Marijnissen J. C. M. Single particle mass spectrometry – bioaerosol analysis by MALDI MS. Proceed. SPIE. 2008; 26: 205–214.
- Klein, H, Nickovic, S, Haunold, W, Bundke, U, Nillius, B. and co-authors. 2010. Saharan dust and ice nuclei over Central Europe. GSA Today. 10: 10211–10221.
- Kopczynski, K, Kwasny, M, Mierczyk, Z and Zawadzki, Z. 2005. Laser induced fluorescence system for detection of biological agents: European project FABIOLA. Oecologia. 1–12.
- Kring D. A. Impact events and their effect on the origin, evolution, and distribution of life. J. Meteorol. 2000; 10: 1.
- Krumbein W. E, Jens K. Biogenic rock varnishes of the negev desert (Israel) an ecological study of iron and manganese transformation by cyanobacteria and fungi. Trend. Plant Sci. 1981; 50: 25–38.
- Kumai M. Snow crystals and the identification of the nuclei in the United-States of America. Proceed. R. Soc. Lond. B. 1961; 18: 139–150.
- Kuparinen A. Mechanistic models for wind dispersal. Curr. Opin. Biotechnol. 2006; 11: 296–301.
- Kuparinen A, Katul G, Nathan R, Schurr F. M. Increases in air temperature can promote wind-driven dispersal and spread of plants. Grana. 2009; B276: 3081–3087.
- Kuske C. R. Current and emerging technologies for the study of bacteria in the outdoor air. Mycol. Res. 2006; 17: 291–296.
- Lacey J. Aggregation of spores and its effect on aerodynamic behaviour. J. Agromed. 1991; 30: 437–445.
- Lacey J. Spore dispersal – its role in ecology and disease: the British contribution to fungal aerobiology. Appl. Environ. Microbiol. 1996; 100: 641–660.
- Langley R. L, Morrow W. E. M. Livestock handling – minimizing worker injuries. Atmos. Environ. 2010; 15: 226–235.
- Lange J. L, Thorne P. S, Lynch N. Application of flow cytometry and fluorescent in situ hybridization for assessment of exposures to airborne bacteria. J. Aerosol Sci. 1997; 63: 1557–1563.
- Lau A. P. S, Lee A. K. Y, Chan C. K, Fang M. Ergosterol as a biomarker for the quantification of the fungal biomass in atmospheric aerosols. Taxon. 2006; 40: 249–259.
- Laucks M. L, Roll G, Schweiger G, Davis E. J. Physical and chemical (Raman) characterization of bioaerosols – pollen. Biogenic particles in the surface microlayer and overlaying atmosphere in the central Arctic Ocean during summer Tellus . 2000; 31: 307–319.
- Laundon J. R. Desmococcus olivaceus – the name of the common subaerial green alga. Geophys. Res. Lett. 1985; 34: 671–672.
- Leck C, Bigg E. K. 2005a; 57B: 305–316.
- Leck C, Bigg E. K. Source and evolution of the marine aerosol – a new perspective. Indoor Air. 2005b; 32: L19803.
- Lee S, Fuhrman J. A. Relationships Between Biovolume and Biomass of Naturally Derived Marine Bacterioplankton. J. Aerosol Sci. 1987; 53: 1298–1303.
- Lee T, Grinshpun S. A, Martuzevicius D, Adhikari A, Crawford C. M, Luo J, Reponen T. Relationship between indoor and outdoor bioaerosols collected with a button inhalable aerosol sampler in urban homes. Sens. Actuators B Chem. 2005; 16: 37–47.
- Lee B. U, Kim S. H, Kim S. S. Hygroscopic growth of E-coli and B-subtilis bioaerosols. 2002; 33: 1721–1723.
- Lee S. J, Park J. S, Im H. T, Jung H. I. A microfluidic ATP-bioluminescence sensor for the detection of airborne microbes. Science. 2008; 132: 443–448.
- Lee, R. E, Warren, G. J and Gusta, L. V. 1995. Biological Ice Nucleation and its Applications. The American Phytopathological Society, St Paul, MN.
- Lelieveld J, Heintzenberg J. Sulfate cooling effect on climate through in-cloud oxidation of anthropogenic SO2. J. Climate Appl. Meteorol. 1992; 258: 117–120.
- Levetin E. Methods for aeroallergen sampling. J. Climate Appl. Meteorol. 2004; 4: 376–383.
- Levin Z, Yankofsky S. A. Contact versus immersion freezing of freely suspended droplets by bacterial ice nuclei. Am. J. Bot. 1983; 22: 1964–1966.
- Levin Z, Yankofsky S. A, Pardes D, Magal N. Possible application of bacterial condensation freezing to artificial rainfall enhancement. Biotechnol. Prog. 1987; 26: 1188–1197.
- Lewis L. A, McCourt R. M. Green algae and the origin of the land plants. Vet. Microbiol. 2004; 91: 1535–1556.
- Li J. K, Asali E. C, Humphrey A. E. Monitoring cell concentration and activity by multiple excitation flourometry. Appl. Environ. Microbiol. 1991; 7: 21–27.
- Li, X, Chai, T, Wang, Z, Song, C, Cao, H. and co-authors. 2009. Occurrence and transmission of Newcastle disease virus aerosol originating from infected chickens under experimental conditions. FEMS Microbiol. Ecol. 36: 226–232.
- Lighthart B. Microbial aerosols – estimated contribution of combine harvesting to an airshed. Aerobiologia. 1984; 47: 430–432.
- Lighthart B. The ecology of bacteria in the alfresco atmosphere. Atmos. Environ. 1997; 23: 263–274.
- Lighthart B. Mini-review of the concentration variations found in the alfresco atmospheric bacterial populations. Appl. Environ. Microbiol. 2000; 16: 7–16.
- Lighthart B, Shaffer B. T. Bacterial flux from Chaparral into the atmosphere in midsummer at a high desert location. Aerobiologia. 1994; 28: 1267–1274.
- Lighthart B, Shaffer B. T. Airborne bacteria in the atmospheric surface layer: temporal distribution above grass seed field. Clin. Microbiol. Rev. 1995; 61: 1492–1496.
- Lighthart B, Shaffer B. T, Frisch A. S, Paterno D. Atmospheric culturable bacteria associated with meteorological conditions at a summer-time site in the mid-Willamette Valley, Oregon. Nature. 2009; 25: 285–295.
- Lim D. V, Simpson J. M, Kearns E. A, Kramer M. F. Current and developing technologies for monitoring agents of bioterrorism and biowarfare. Appl. Environ. Microbiol. 2005; 18: 583–607.
- Lindahl T. Instability and decay of the primary structure of DNA. Appl. Environ. Microbiol. 1993; 362: 709–715.
- Lindemann J, Constantinidou H. A, Barchet W. R, Upper C. D. Plants as sources of airborne bacteria, including ice nucleation-active bacteria. Mol. Plant-Microbe Interact. 1982; 44: 1059–1063.
- Lindow S. E, Brandl M. T. Microbiology of the phyllosphere. Allergologie. 2003; 69: 1875–1883.
- Lindow S. E, Lahue E, Govindarajan A. G, Panopoulos N. J, Gies D. Localization of ice nucleation activity and the Icec Gene-Product in Pseudomonas-Syringae and Escherichia-Coli. Science. 1989; 2: 262–272.
- Linskens H. F, Jorge W. Pollenstransport in größen Höhen. Beobachtungen während der Fahrt mit einem Gasballon. J. Allergy Clin. Immunol. 1986; 9: 55–58.
- Liu, J, Xiao, H, Lei, F, Zhu, Q, Qin, K. and co-authors. 2005. Highly pathogenic H5N1 influenza virus infection in migratory birds. Clinica Chimica Acta. 309: 1206.
- Lo, E and Levetin, E. 2007. Influence of meteorological conditions on early spring pollen in the Tulsa atmosphere from 1987–2006. J. Phycol. 119.
- Loy A, Bodrossy L. Highly parallel microbial diagnostics using oligonucleotide microarrays. Nature. 2006; 363: 106–119.
- Lundheim R. Ice nucleation in seaweeds in relation to vertical zonation. J. Aerosol Sci. 1997; 33: 739–742.
- Lutzoni F, Pagel M, Reeb V. Major fungal lineages derived from lichen symbiotic ancestors. J. Appl. Bacteriol. 2001; 411: 937–940.
- Madelin T. M. Fungal aerosols: a review. Glob. Biogeochem. Cycle. 1994; 25: 1405–1412.
- Madelin T. M, Johnson H. E. Fungal and actinomycete spore aerosols mearsured at different humidities with an aerodynamic particle sizer. J. Aerosol Sci. 1992; 72: 400–409.
- Mahowald, N, Jickells, T. D, Baker, A. R, Artaxo, P, Benitez-Nelson, C. R. and co-authors. 2008. Global distribution of atmospheric phosphorus sources, concentrations and deposition rates, and anthropogenic impacts. Aerosol Sci. Technol. 22: 10.3402/tellusb.v64i0.15598.
- Mainelis, G, Adhikari, A, Willeke, K, Lee, S. A, Reponen, T. and co-authors. 2002. Collection of airborne microorganisms by a new electrostatic precipitator. Doclady Acad. Nauk Tadjic. SSR. 33: 1417–1432.
- Mainelis, G, Grinshpun, S. A, Willeke, K, Reponen, T, Ulevicius, V. and co-authors. 1999. Collection of airborne microorganisms by electrostatic precipitation. Appl. Microbiol. 30: 127–144.
- Maiorov, V. I, Scherbinin, A. V, Bogachev, S. S, Gileva, I. P, Muzychenko, M. V. and co-authors. 1985. Effectiveness of insecticides based on nuclear polyhedrosis virus of ballworm. Sci. Tot. Environ. 28: 733–736.
- Maki, L. R, Galyan, E. L, Changchi.Mm and Caldwell, D. R. 1974. Ice nucleation induced by Pseudomonas-Syringae. J. Appl. Meteorol. 28: 456–459.
- Maki, T, Susuki, S, Kobayashi, F, Kakikawa, M, Tobo, Y. and co-authors. 2010. Phylogenetic analysis of atmospheric halotolerant bacterial communities at high altitude in an Asian dust (KOSA) arrival region, Suzu City. Appl. Environ. Microbiol. 408: 4556–4562.
- Maki L. R, Willoughby K. J. Bacteria as biogenic sources of freezing nuclei. Appl. Opt. 1978; 17: 1049.
- Mancinelli R. L, Shulls W. A. Airborne bacteria in an urban environment. J. Aerosol Sci. 1978; 35: 1095–1101.
- Manninen, A, Putkiranta, M, Rostedt, A, Saarela, J, Laurila, T. and co-authors. 2008. Instrumentation for measuring fluorescence cross sections from airborne microsized particles. Atmos. Environ. 47: 110–115.
- Marks R, Kruczalak K, Jankowska K, Michalska M. Bacteria and fungi in air over the Gulf of Gdansk and Baltic sea. Atmos. Environ. 2001; 32: 237–250.
- Maron, P.-A, Lejon, D. P. H, Carvalho, E, Bizet, K, Lemanceau, P. and co-authors. 2005. Assessing genetic structure and diversity of airborne bacterial communities by DNA Fingerprinting and 16S rDNA clone library. New Phyologists. 39: 3687–3695.
- Maron, P.-A, Mougel, C, Lejon, D. P. H, Carvalho, E, Bizet, K. and co-authors. 2006. Temporal variability of airborne bacterial community structure in an urban area. Atmos. Res. 40: 8074–8080.
- Marshall W. Aerial dispersal of lichen soredia in the maritime Antarctic. Environ. PLoS ONE. 1996; 134: 523–530.
- Matthias-Maser S, Jaenicke R. Size distribution of primary biological aerosol particles with radii > 0.2 (m in an urban/rural influenced region. Science. 1995; 39: 279–286.
- McDevitt, J. J, Milton, D. K, Rudnick, S. N and First, M. W. 2008. Inactivation of poxviruses by upper-Room UVC light in a simulated hospital room. Aerosol Sci. Technol. 3: 10.3402/tellusb.v64i0.15598. e3186.
- McDonald J. E. Collection and washout of airborne pollens and spores by raindrops. Proceed. Soc. Exp. Biol. Med. 1962; 135: 435–437.
- Miguel A. G, Taylor P. E, House J, Glovsky M. M, Flagan R. C. Meteorological influences on respirable fragment release from Chinese elm pollen. Am. J. Hyg. 2006; 40: 690–696.
- Miller W. S, Artenstein M. S. Aerosol stability of three acute respiratory disease viruses. Appl. Opt. 1967; 125: 222–227.
- Miller W. S, Demchak P, Rosenberger C. R, Dominik J. W, Bradshaw J. L. Stability and infectivity of airborne Yellow fever and Rift Valley fever viruses. 1963; 77: 114–121.
- Mims F. M. Solar corona caused by juniper pollen in Texas. 1998; 37: 1486–1488.
- Miquel, P. 1883. Les Organismes Vivants de l'atmosphere. Gauthier-Villars, Paris.
- Mitchel, J. P. 1995. Particel size analyzer: practical procedures and laboratory techniques. In: Bioaerosols Handbook. (C. S.Cox, and C. M.Wathes). CRC-Press: Boca RatonFL, 177–246.
- Moffett B. F, Walsh K. A, Harris J. A, Hill T. C. J. erschienen in Journal. Biogeosciences. 2000; 6: 129–131.
- Möhler O, DeMott P. J, Vali G, Levin Z. Microbiology and atmospheric processes: the role of biological particles in cloud physics. Sci. Tot. Environ. 2007; 4: 1059–1071.
- Möhler, O, Georgakopoulos, D. G, Morris, C. E, Benz, S, Ebert, V. and co-authors. 2008. Heterogeneous ice nucleation activity of bacteria: new laboratory experiments at simulated cloud conditions. Trudy Geofizicheskogo Instituta Akademiya Nauk SSSR. 5: 1425–1435.
- Moletta M, Delgenes J.-P, Godon J.-J. Differences in the aerosolization behavior of microorganisms as revealed through their transport by biogas. Appl. Opt. 2007; 379: 75–88.
- Monin A. S, Obukhov A. M. Basic laws of turbulent mixing in the ground layer of the atmosphere. Can. Entomol. 1954; 151: 163–187.
- Morel S, Leone N, Adam P, Amouroux J. Detection of bacteria by time-resolved laser-induced breakdown spectroscopy. 2003; 42: 6184–6191.
- Morris O. N. Entomopathogenic viruses: strategies for use in forest insect pest management. Isme Journal. 1980; 112: 573–584.
- Morris, C. E and Kinkel, L. L. 2002. Fifty years of phyllosphere microbiology: significant contributions to research in related fields. In: Phyllosphere Microbiology. (S. E.Lindow, E. I.Hecht-Poinar and V. J.Elliot). APS Press: MN, 365–375.
- Morris, C. E, Sands, D. C, Vinatzer, B. A, Glaux, C, Guilbaud, C. and co-authors. 2008. The life history of the plant pathogen pseudomonas syringae is linked to the water cycle. Environ. Chem. 2: 321–334.
- Morris C. E, Sands D. C, Bardin M, Jaenicke R, Vogel B, Leyronas C, Ariya P. A, Psenner R. Microbiology and atmospheric processes: research challenges concerning the impact of airborne micro-organisms on the atmosphere and climate. 2011; 8: 17–25.
- Mortazavi R, Hayes C. T, Ariya P. A. Ice nucleation activity of bacteria isolated from snow compared with organic and inorganic substrates. Clin. Allergy. 2008; 5: 373–381.
- Mücke, W and Lemmen, C. 2008. Bioaerosole und Gesundheit, ecomed Medizin Verlagsgruppe Hüthig Jehle Rehm GmbH, 1–240.
- Mullius J, Hutcheson P, Slain R. Aspergillus spore concentration on outside air. Cardiff and St.Louis compared. 1984; 14: 351–364.
- Nagarajan S, Singh D. V. Long-distance dispersion of rust pathogens. Nature. 1990; 28: 139–53.
- Nash III, T. H. 1996. Lichen Biology. (T. H.Nash III). Cambridge University Press, Cambridge, New York, 1–7.
- Nathan, R, Katul, G, Horn, H. S, Thomas, S. M, Oren, R. and co-authors. 2002. Mechanisms of long-distance dispersal of seeds by wind. Bioscience. 418: 409–413.
- Nathan, R, Schurr, G. M, Spiegel, O, Steinitz, O, Trakhtenbroth, A. and co-authors. 2008. Mechanisms of long-distance seed dispersal. Plant Ecol. Evol. 23: 638–647.
- Neilson, R. P, Pitelka, L. F, Solomon, A. M, Nathan, R, Midgley, G. F. and co-authors. 2005. Forecasting regional to global plant migration in response to climatic change. 55: 749–759.
- Neustupa J, Skaloud P. Diversity of subaerial algae and cyanobacteria growing on bark and wood in the lowland tropical forests of Singapore. Bot. Rev. 2010; 143: 51–62.
- Nicholson, K. W. 1995. Physical aspects if bioaerosol sampling and deposition. In: Bioaerosols Handbook. (Cox, C. S. and C. M.Wathes), CRC-Press, Boca Raton: FloridaUSA, 27–47.
- Niklas K. J. The aerodynamics of wind pollination. J. Geophys. Res. 1985; 51: 328–386.
- Nilsson, E. D, Martensson, E. M, Van Ekeren, J. S, Leeuw, G. d, Moerman, M. and co-authors. 2007. Primary marine aerosol emission: size resolved eddy covariance measurements with estimates of the sea salt and organic carbon fractions. Oecologia. 7: 13345–13400.
- Nilsson, E. F, Rannik, U, Swietlicki, E, Leck, C, Aalto, P. P. and co-authors. 2001. Turbulent aerosol fluxes over the Arctic Ocean 2. Winddriven sources from the seas. Appl. Environ. Microbiol. 106: 32139–32154.
- Nybakken L, Solhaug K. A, Bilger W, Gauslaa Y. The lichens Xanthoriaelegans and Cetraria islandica maintain a high protection against UV-B radiation in Arctic habitats. Nature. 2004; 140: 211–216.
- O'Brien H. E, Parrent J. L, Jackson J. A, Moncalvo J. M, Vilgalys R. Fungal community analysis by large-scale sequencing of environmental samples. Cryobiology. 2005; 71: 5544–5550.
- O'Dowd, C. D, Facchini, M. C, Cavalli, F, Ceburnis, D, Mircea, M. and co-authors. 2004. Biogenically driven organic contribution to marine aerosol. Am. J. Bot. 431: 676–680.
- Obata H, Muryoi N, Kawahara H, Yamade K, Nishikawa J. Identification of a novel ice-nucleating bacterium of Antarctic origin and its ice nucleation properties. Appl. Microbiol. Biotechnol. 1999; 38: 131–139.
- Ogden E. C, Hayes J. V, Raynor G. S. Diurnal patterns of pollen emission in Ambrosia, Phleum, Zea, and Ricinus. Int. J. Biometeorol. 1969; 56: 16–21.
- Oksanen I. Ecological and biotechnological aspects of lichens. Appl. Microbiol. 2006; 73: 723–734.
- Oliviera M, Ribeiro H, Delgado J, Abreu I. The effects of meteorological factors on airborne fungal spore concentration in two areas differing in urbanisation level. Proc. Natl. Acad. Sci. USA. 2009; 53: 61–73.
- Orr C, Gordon M. T, Kordecki M. C. Thermal precipitation for sampling air-borne microorganisms. Annu. Rev. Genet. 1956; 4: 116–118.
- Pääbo S. Ancient DNA extraction, characterization, molecular cloning, and enzymatic amplification. Transact. Br. Mycol. Soc. 1989; 86: 1939–1943.
- Pääbo, S, Poinar, H, Serre, D, Jaenicke-Després, V, Hebler, J. and co-authors. 2004. Genetic analysis from ancient DNA. Aerosol Sci. Technol. 38: 645–679.
- Pady S. M, Gregory P. H. Numbers and viability of airborne hyphal fragments in England. Environ. Sci. Technol. 1963; 46: 609–613.
- Pan, Y. L, Boutou, V, Bottiger, J. R, Zhang, S. S, Wolf, J. P. and co-authors. 2004. A puff of air sorts bioaerosols for pathogen identification. J. Geophys. Res. Atmos. 38: 598–602.
- Pan Y. L, Pinnick R. G, Hill S. C, Chang R. K. Particle-Fluorescence Spectrometer for Real-Time Single-Particle measurements of atmospheric organic carbon and biological aerosol. Appl. Spectroscopy. 2009; 43: 429–434.
- Pan Y. L, Pinnick R. G, Hill S. C, Rosen J. M, Chang R. K. Single-particle laser-induced-fluorescence spectra of biological and other organic-carbon aerosols in the atmosphere: measurements at New Haven, Connecticut, and Las Cruces, New Mexico. Field Anal. Chem. Technol. 2007; 112: D24S19.
- Pappas C. S, Tarantilis P. A, Harizanis P. C, Polissiou M. G. New method for pollen identification by FT-IR spectroscopy. Appl. Opt. 2003; 57: 23–27.
- Parker, E. P, Trahan, M. W, Wagner, J. S, Rosenthal, S. E, Whitten, W. B.and co-authors. 2000. Detection and classification of individual airborne microparticles using laser ablation mass spectroscopy and multivariate analysis. Atmos. Environ. A Gen. Topics. 4: 31–42.
- Parviainen P, Bohren C. F, Makela V. Vertical elliptic coronas caused by pollen. Annales des Science Naturelles (partie zoologique). 1994; 33: 4548–4551.
- Pasanen A. L, Pasanen P, Jantunen M. J, Kalliokoski P. Significance of air humidity and air velocity for fungal spore release into the air. Atmos. Environ. 1991; 25: 459–462.
- Pasteur L. Mémoire sur les corpuscles organises qui existent dans l'atmosphere. Examen de la doctrine des générations spontanées. Atmos. Environ. 1861; 16: 5–98.
- Peccia J, Hernandez M. Incorporating polymerase chain reaction-based identification, population characterization, and quantification of microorganisms into aerosol science: a review. 2006; 40: 3941–3961.
- Peccia J, Hospodsky D, Bibby K. New directions: a revolution in DNA sequencing now allows meaningful integration of biology with aerosol science. 2011; 45: 1896–1897.
- Penner, J. E. 1995. Carbonaceous aerosols influencing atmospheric radiation: black and organic Carbon. In: Aerosol Forcing of Climate. (R. J.Charlson and J.Heintzenberg). John Wiley and Sons: Chichester, 91–108.
- Penner, J. E, Andreae, M. O, Annegarn, H, Barrie, L, Feichter, J. and co-authors. 2001. Aerosols, their direct and indirect effects. In: The Scientific Basis. Contribution of Working Group I To The Third Assessment Report Of The Intergovernmental Panel On Climate Change. (J. T.Houghton, Y.Ding, D. J.Griggs, M.Noguer, P. J.van der Linden, X.Dai, K.Maskell and C. A.Johnson) Cambridge University Press, Cambridge, UK and New York, NY, USA, 289–348.
- Pepponi, G, Lazzeri, P, Coghe, N, Bersani, M, Gottardini, E. and co-authors. 2004. Total reflection X-ray fluorescence analysis of pollen as an indicator for atmospheric pollution. J. Bacteriol. 59: 1205–1209.
- Petters M. D, Kreidenweis S. M. A single parameter representation of hygroscopic growth and cloud condensation nucleus activity. Biogeosciences. 2007; 7: 1961–1971.
- Phelps P, Giddings T. H, Prochoda M, Fall R. Release of cell-free ice nuclei by Erwinia-Herbicola. J. Atmos. Sci. 1986; 167: 496–502.
- Phillips, V. T. J, Andronache, C, Christner, B, Morris, C. E, Sands, D. C. and co-authors. 2009. Potential impacts from biological aerosols on ensembles of continental clouds simulated numerically. Environ. 6: 987–1014.
- Phillips V. T. J, DeMott P. J, Andronache C. An empirical parameterization of heterogeneous ice nucleation for multiple chemical species of aerosol. 2008; 65: 2757–2783.
- Pinnick R. G, Hill S. C, Pan Y. L, Chang R. K. Fluorescence spectra of atmospheric aerosol at Adelphi, Maryland, USA: measurement and classification of single particles containing organic carbon. Atmos. Atmos. Measure. Techniq. Discuss. 2004; 38: 1657–1672.
- Pitt J. I, Hocking A. D. Fungi and food spoilage. Springer. 1997
- Pöhlker C, Huffman J. A, Pöschl U. Autofluorescence of atmospheric bioaerosols – fluorescent biomolecules and potential interferences. Microbiol. 2011; 4: 5857–5933.
- Polymenakou P. A, Mandalakis M, Stephanou E. G, Tselepides A. Particle size distribution of airborne microorganisms and pathogens during an intense African dust event in the eastern Meidterranean. 2008; 116: 292–296.
- Polz M. F, Cavanaugh C. M. Bias in template-to-product ratios in multitemplate PCR. Appl. Environ. FEMS Microbiol. Ecol. 1998; 64: 3724–3730.
- Pomeroy, J. W and Jones, H. G. 1996. Wind-blown snow: sublimation, transport and changes to polar snow. In: Chemical Exchange between the Atmosphere and Polar Snow. (E. W.Wolff and R. C.Bales). Springer-Verlag, Berlin. 678.
- Ponder, M. A, Gilmour, S. J, Bergholz, P. W, Mindock, C. A, Hollingsworth, R. and co-authors. 2005. Characterization of potential stress responses in ancient Siberian permafrost psychroactive bacteria. J. Virol. Methods. 53: 103–115.
- Pope F. D. Pollen grains are efficient cloud condensation nuclei. Angewandte Chemie-International Edition . 2010; 5: 1–6.
- Posada J. A, Redrow J, Celik I. A mathematical model for predicting the viability of airborne viruses. Science. 2010; 164: 88–95.
- Pöschl U. Atmospheric aerosols: composition, transformation, climate and health effects. Appl. Environ. Microbiol. 2005; 44: 7520–7540.
- Pöschl, U, Martin, S. T, Sinha, B, Chen, Q, Gunthe, S. S. and co-authors. 2010. Rainforest aerosols as biogenic nuclei of clouds and precipitation in the Amazon. Nat. Geosci. 329: 1513–1516.
- Pouleur S, Richard C, Martin J. G, Antoun H. Ice nucleation activity in fusarium-acuminatum and fusarium-avenaceum. Nat. Geosci. 1992; 58: 2960–2964.
- Pratt, K. A, DeMott, P. J, French, J. R, Wang, Z, Westphal, D. L. and co-authors. 2009. In situ detection of biological particles in cloud ice-crystals. Appl. Environ. Microbiol. 2: 397–400.
- Prenni, A. J, Petters, M. D, Kreidenweis, S. M, Heald, C. L, Martin, S. T. and co-authors. 2009. Relative roles of biogenic emissions and Saharan dust as ice nuclei in the Amazon basin. Mycologia. 2: 401–404.
- Prigione V, Lingua G, Marchisio V. F. Development and use of flow cytomefry for detection of airborne fungi. Det Kongelige Norske Videnskabers Selskabs Skrifter. 2004; 70: 1360–1365.
- Pringle A, Patek S. N, Fischer M, Stolze J, Money N. P. The captured launch of a ballistospore. Aerobiologia. 2005; 97: 866–871.
- Printz H. Subaerial algae from South Africa. 1921; 120: 1–54.
- Prospero J. M, Blades E, Mathison G, Naidu R. Interhemispheric transport of viable fungi and bacteria from Africa to the Caribbean with soil dust. Atmos. Environ. 2005; 21: 1–19.
- Pruppacher H. R, Klett J. D. Microphysics of Clouds and Precipitation. Kluwer Academic Publisher: Dordrecht, 1997
- Puxbaum H, Tenze-Kunit M. Size distribution and season variation of atmospheric cellulose. Annu. Rev. Microbiol. 2003; 37: 3693–3699.
- Radosevich J. L, Wilson W. J, Shinn J. H, DeSantis T. Z, Andersen G. L. Development of a high-volume aerosole collection system fort he identification of airborne microorganisms. Extremophiles. 2002; 34: 162–167.
- Rappé M. S, Giovannoni S. J. The uncultured microbial majority. Eco. Health. 2003; 57: 369–394.
- Raymond J. A, Christner B. C, Schuster S. C. A bacterial ice-binding protein from the Vostok ice core. 2008; 12: 713–717.
- Reid C. E, Gamble J. L. Aeroallergens, allergic disease, and climate change: impacts and adaptation. Planta. 2009; 6: 458–470.
- Reisser, W. 2002. Algae living on trees. In: Symbiosis: Mechanisms and Model Systems. (J.Seckbach). Kluwer Academic Publishers, The Netherlands. 387–395.
- Rempe H. Untersuchungen über die Verbreitung des Blütenstaubes durch die Luftströmungen. Grana. 1937; 27: 93–147.
- Reponen T. A, Gazenko S. V, Grinshpun S. A, Willeke K, Cole E. C. Characteristics of airborne actinomycete spores. 1998; 64: 3807–3812.
- Reponen T, Grinshpun S. A, Conwell K. L, Wiest J, Anderson M. Aerodynamic versus physical size of spores: measurement and implication for respiratory deposition. Atmos. Environ. 2001; 40: 119–125.
- Reponen, T, Willeke, K, Grinshpun, S and Nevalainen, A. 1995. Biological particle sampling. In: Bioaerosol Handbook. (C. S.Cox, and C. M.Wathes). CRC-Press: Boca RatonFL, 751–778.
- Reponen T, Willeke K, Ulevicius V, Reponen A, Grinshpun S. A. Effect of relative humidity on the aerodynamic diameter and respiratory deposition of fungal spores. Phytoprotection. 1996; 30: 3967–3974.
- Reyes, F. L, Jeys, T. H, Newbury, N. R, Primmerman, C. A, Rowe, G. S. and co-authors. 1999. Bio-aerosol fluorescence sensor. 3: 240–248.
- Richard C, Martin J. G, Pouleur S. Ice nucleation activity identified in some phytopathogenic Fusarium species. ACS Symp. Ser. 1996; 77: 83–92.
- Richardson, D. H. S. 1975. The vanishing lichens: their history, biology and importance.
- Rishpon J, Gezundhajt Y, Soussan L, Rosenmargalit I, Hadas E. Immunoelectrodes for the Detection of Bacteria. Anal. Chem. 1992; 511: 59–72.
- Rosas I, Calderon C, Ulloa M, Lacey J. Abundance of airborne Penicillium CFU in relation to urbanization in Mexico City. Microbiol. Rev. 1993; 59: 2648–2652.
- Rosch, P, Harz, M, Peschke, K. D, Ronneberger, O, Burkhardt, H. and co-authors. 2006. On-line monitoring and identification of bioaerosols. Am. Indust. Hyg. Assoc. J. 78: 2163–2170.
- Roszak D. B, Colwell R. R. Survival strategies of bacteria in the natural-environment. Mass Spectrom. Rev. 1987; 51: 365–379.
- Rubow K. L, Marple V. A, Olin J, McCawley M. A. A personal cascade impactor: design, evaluation and calibration. Int. J. Biometeorol. 1987; 48: 532–538.
- Russell S. C. Microorganism characterization by single particle mass spectrometry. 2009; 28: 376–387.
- Sabariego S, Díaz de la Guardia C, Alba F. The effect of meteorological factors on the daily variation of airborne fungal spores in Granada (southern Spain). Photochem. Photobiol. 2000; 44: 1–5.
- Safatov, A. S, Buryak, G. A, Andreeva, I. S, Olkin, S. E, Reznikova, I. K. and co-authors. 2010. Atmospheric bioaerosols. In: Aerosols Science and Technology. (I.Agranovski). Wiley – VCH: Weinheim, 407–454.
- Sagripanti J. L, Lytle C. D. Inactivation of influenza virus by solar radiation. J. Geophys. Res. 2007; 83: 1278–1282.
- Samuels A. C, DeLucia F. C, McNesby K. L, Miziolek A. W. Laser-induced breakdown spectroscopy of bacterial spores, molds, pollens, and protein: initial studies of discrimination potential. Idöjárás. 2003; 42: 6205–6209.
- Sánchez-Ochoa, A, Kasper-Giebl, A, Puxbaum, H, Gelencserm, A, Legrand, M. and co-authors. 2007. Concentration of atmospheric cellulose: a proxy for plant debris across a west-east transect over Europe. Biosci. Rep. 112: 10.3402/tellusb.v64i0.15598.
- Sands D. C, Langhans V. E, Scharen A. L, de Smet G. The association between bacteria and rain and possible resultant meteorological implications. J. Mol. Biol. 1982; 86: 148–152.
- Sanger F. Determination of nucleotide sequences in DNA. Anal. Chem. 1981; 1: 3–18.
- Sanger F, Coulson A. R. A rapid method for determining sequences in DNA by primed synthesis with DNA polymerase. 1975; 94: 441–446.
- Sarantaridis D, Caruana D. J. Potentiometric detection of model bioaerosol particles. Geophys. Res. Lett. 2010; 82: 7660–7667.
- Sassen, K. 2008. Boreal tree pollen sensed by polarization lidar: depolarizing biogenic chaff. Geophys. Res. Lett. L18810.
- Sattler B, Puxbaum H, Psenner R. Bacterial growth in supercooled cloud droplets. Med. J. Rec. 2001; 28: 239–242.
- Schaffer F. L, Soergel M. E, Straube D. C. Survival of airborne influenza virus: effect of propagation host, relative humidity, and composition of spray fluids. Nat. Rev. Microbiol. 1976; 51: 263–273.
- Scheppegrell W. Airplane tests of hayfever pollen density in the upper air. Appl. Opt. 1924; 119: 185–189.
- Schleper C, Jurgens G, Jonuscheit M. Genomic studies of uncultivated archaea. 2005; 3: 479–488.
- Schmidt M. S, Bauer A. J. R. Preliminary correlations of feature strength in spark-induced breakdown spectroscopy of bioaerosols with concentrations measured in laboratory analyses. Appl. Opt. 2010; 49: C101–C109.
- Schneider, J, Freutel, F, Zorn, S. R, Chen, Q, Farmer, D. K. and co-authors. 2011. Mass-spectrometric identification of primary biological particle markers: indication for low abundance of primary biological material in the pristine submicron aerosol of Amazonia. Atmos. Chem. Phys. Discuss. 11: 19143–19178.
- Schneider W. B, Vollmer M. Experimental simulations of pollen coronas. Tellus. 2005; 44: 5746–5753.
- Schnell R. C. Ice nuclei produced by laboratory cultured marine phytoplankton. Nature. 1975; 2: 500–502.
- Schnell R. C, Tan-Schnell S. N. Kenyan tea-Litter – a source of ice nuclei. Tellus. 1982; 34: 92–95.
- Schnell R. C, Vali G. Atmospheric ice nuclei from decomposing vegetation. J. Atmos. Sci. 1972; 236: 163–165.
- Schnell R. C, Vali G. Freezing nuclei in marine waters. Anal. Chem. 1975; 27: 321–323.
- Schnell R. C, Vali G. Biogenic ice nuclei: part I, Terrestrial and marine sources. Aerosol Sci. Technol. 1976; 33: 1554–1564.
- Schulte F, Lingott J, Panne U, Kneipp J. Chemical characterization and classification of pollen. 2008; 80: 9551–9556.
- Seaver M, Eversole J. D, Hardgrove J. J, Cary W. K, Roselle D. C. Size and fluorescence measurements for field detection of biological aerosols. J. Colloid Interface Sci. 1999; 30: 174–185.
- Seinfeld J. H, Pandis S. N. Atmospheric chemistry and physics. From air pollution to climate change. John Wiley & Sons Inc: New Jersey, 2006
- Sengupta A, Brar N, Davis E. J. Bioaerosol detection and characterization by surface-enhanced Raman spectroscopy. Anal. Bioanal. Chem. 2007; 309: 36–43.
- Sengupta A, Laucks M. L, Davis E. J. Surface-enhanced Raman spectroscopy of bacteria and pollen. Biogeosciences. 2005; 59: 1016–1023.
- Sengupta A, Mujacic M, Davis E. J. Detection of bacteria by surface-enhanced Raman spectroscopy. J. Aerosol Sci. 2006; 386: 1379–1386.
- Sesartic A, Dallafior T. N. Global fungal spore emissions, review and synthesis of literature data. Microb. Ecol. 2011; 8: 1181–1192.
- Seshadri S, Han T, Krumins V, Fennell D. E, Mainelis G. Application of ATP bioluminescence method to characterize performance of bioaerosol sampling devices. Aerobiologia. 2009; 40: 113–121.
- Shaffer B. T, Lighthart B. Survey of culturable airborne bacteria at four diverse locations in Oregon: urban, rural, forest, and coastal. Ecotoxicol. Environ. Saf. 1997; 34: 167–177.
- Shahamat, M, Levin, M, Rahman, I, Grim, C, Heidelberg, J. and co-authors. 1997. Evaluation of media for recovery of aerosolized bacteria. J. Phycol. 13: 219–226.
- Sharma N. K, Rai A. K. Allergenicity of airborne cyanobacteria Phormidium fragile and Nostoc muscorum. Geophys. Res. Lett. 2008; 69: 158–162.
- Sharma N. K, Rai A. K, Singh S, Brown R. M. Airborne algae: their present status and relevance. Nat. Chem. 2007; 43: 615–627.
- Shinn, E. A, Smith, G. W, Prospero, J. M, Betzer, P, Hayes, M. L. and co-authors. 2000. African dust and the demise of Caribbean coral reefs. J. Preven. Med. Hyg. 27: 3029–3032.
- Shiraiwa M, Sosedova Y, Rouvière A, Yang H, Zhang Y, Abbatt J. P. D, Ammann M, Pöschl U. The role of long-lived reactive oxygen intermediates in the reaction of ozone with aerosol particles. IEEE Transact. Geosci. Rem. Sens. 2011; 3: 291–295.
- Sigari, G, Panatto, D, Lai, P, Stefani, L, Giuntini, A. and co-authors. 2006. Virological investigation on aerosol from waste depuration plants. 47: 4–7.
- Simard, J. R, Roy, G, Mathieu, P, Larochelle, V, McFee, J. and co-authors. 2004. Standoff sensing of bioaerosols using intensified range-gated spectral analysis of laser-induced fluorescence. Field Anal. Chem. Technol. 42: 865–874.
- Simkin, T and Siebert, L. 1994. Volcanoes of the World. Geoscience Press, TusconAZ.
- Sincock S. A, Kulaga H, Cain M, Anderson P, Stopa P. J. Applications of flow cytometry for the detection and characterization of biological aerosols. Appl. Opt. 1999; 3: 291–306.
- Sinha R. J, Kramer C. L. Identifying hyphal fragments in the atmosphere. Trans. Kansas Acad. Agric. Forest Meteorol. 1971; 74: 48–51.
- Sivaprakasam, V, Pletcher, T, Tucker, J. E, Huston, A. L, McGinn, J. and co-authors. 2009. Classification and selective collection of individual aerosol particles using laser-induced fluorescence. Nature. 48: B126–B136.
- Skelsey P, Holtslag A, Vanderwerf W. Development and validation of a quasi-Gaussian plume model for the transport of botanical spores. Journal de Recherches Atmospheriques. 2008; 148: 1383–1394.
- Smith, C. I, Chamberlain, A. T, Riley, M. S, Cooper, A, Stringer, C. B. and co-authors. 2001. Neanderthal DNA: not just old but old and cold?. Field Anal. Chem. Technol. 410: 771–772.
- Snider J. R, Layton R. G, Caple G, Chapman D. Bacteria as condensation nuclei. Int. J. Biometeorol. 1985; 19: 139–145.
- Snyder A. P, Tripathi A, Maswadeh W. M, Ho J, Spence M. Field detection and identification of a bioaerosol suite by pyrolysis-gas chromatography-ion mobility spectrometry. 2001; 5: 190–204.
- Sofiev M, Silijamo P, Ranta H, Rantio-Lehtimäki A. Towards numerical forecasting of long-range air transport of birch pollen: theoretical considerations and a feasibility study. J. Allergy. 2006; 50: 392–402.
- Solomon W. R, Stohrer A. W, Gilliam J. A. The “fly-shield” rotobar: a simplified impaction sampler with motion-regulated shielding. Appl. Microbiol. 1968; 41: 290–296.
- Songer J. R. Influence of relative humidity on the survival of some airborne viruses. Ecology. 1967; 15: 35–42.
- Soons M. B, Heil G. W, Nathan R, Katul G. G. Determinants of long-distance seed dispersal by wind in grasslands. J. Geophys. Res. Atmos. 2004; 85: 3056–3068.
- Spankuch D, Dohler W, Guldner J. Effect of coarse biogenic aerosol on downwelling infrared flux at the surface. J. Aerosol Sci. 2000; 105: 17341–17350.
- Spurny K. R. On the chemical detection of bioaerosols. Annu. Rev. Microbiol. 1994; 25: 1533–1547.
- Staley, J and Konopka, A. 1985. Measurement of insitu activities of nonphotosynthetic microorganisms in aquatic and terrestrial habitats. 39: 321–346.
- Stanley R, Linskins H. Pollen: Biology, Chemistry and Management. Springer-Verlag: Berlin, 1974
- Stewart, I and Falconer, I. R. 2008. Cyanobacteria and cyanobacterial toxins. In: Oceans and human health: risks and remedies from the seas. (P. J.Walsh, S. L.Smith and L. E.Fleming). Academic Press, Burlington, MA. 271–296.
- Stewart, S. L, Grinshpun, S. A, Willeke, K, Terzieva, S, Ulevicius, V. and co-authors. 1995. Effect of impact stress on microbial recovery on an agar surface. Appl. Opt. 61: 1232–1239.
- Stowers M. A, van Wuijckhuijse A. L, Marijnissen J. C. M, Kientz C. E, Ciach T. Fluorescence preselection of bioaerosol for single-particle mass spectrometry. 2006; 45: 8531–8536.
- Straka H. Pollen- und Sporenkunde. Fischer Verlag: Stuttgart, 1975
- Strauch D, Ballarini G. Hygienic aspects of the production and agricultural use of animal wastes. Aerosol Sci. Technol. 1994; 41B: 176–228.
- Surbek M, Esen C, Schweiger G. Elastic light scattering on single pollen: scattering in a large space angle. Bulletin of the Amer. Meteor. Soc. 2009; 43: 679–684.
- Szyrmer W, Zawadzki I. Biogenic and anthropogenic sources of ice-forming nuclei: a review. Ecol. Monogr. 1997; 78: 209–228.
- Tackenberg O. Modelling long-distance dispersal if plant diaspores by wind. Aerosol Sci. Technol. 2003; 73: 173–189.
- Tan M, Shen F, Yao M, Zhu T. Development of automated electrostatic sampler (AES) for bioaerosol detection. Clin. Exp. Allergy. 2011; 45: 1154–1160.
- Taylor P. E, Flagan R, Miguel A. G, Valenta R, Glovsky M. M. Birch pollen rupture and the release of aerosols of respirable allergens. J. Allergy Clin. Immunol. 2004; 34: 1591–1596.
- Taylor P. E, Flagan R. C, Valenta R, Glovsky M. M. Release of allergens as respirable aerosols: a link between grass pollen and asthma. Curr. Allergy Asthma Rep. 2002; 109: 51–56.
- Taylor P. E, Jonsson H. Thunderstorm asthma. Biochem. J. 2004; 4: 409–413.
- Teale F. W. J, Weber G. Ultraviolet fluorescence of the aromatic amino acids. 1957; 65: 476–482.
- Tehler, A. 1996. Systematics, phylogeny and classification. In: Lichen Biology. (T. H.Nash). Cambridge University Press, Cambridge, 217–239.
- Thummes K, Schäfer J, Kämpfer P, Jäckel U. Thermophilic methanogenic Archaea in compost material: occurrence, persistence and possible mechanisms for their distribution to other environments. Int. J. Biometeorol. 2007; 30: 634–643.
- Tormo, R, Silva, I, Gonzalo, A, Moreno, A, Pérez, R. and Fernández, S. 2010. Phenological records as complement to aerobiological data. Aerosol Sci. Technol. 55, 51–65.
- Tong Y, Lighthart B. Diurnal distribution of total and culturable atmospheric bacteria at a rural site. Aerosol Sci. Technol. 1999; 30: 246–254.
- Tong Y, Lighthart B. The annual bacterial particle concentration and size distribution in the ambient atmosphere in a rural area of the Willamette Valley, Oregon. Opt. 2000; 32: 393–493.
- Trankle E, Mielke B. Simulation and analysis of pollen coronas. Appl. Int. J. Biometeorol. 1994; 33: 4552–4562.
- Troutt C, Levetin E. Correlation of spring spore concentrations and meteorological conditions in Tulsa, Oklahoma. 2001; 45: 64–74.
- Tschermal-Woess, E. 1988. The algal partner. In: CRC Handbook of Lichenology. vol. I. (M.Galun). CRC Press, Boca Raton, Florida, 39–92.
- Tsumuki H, Konno H, Maeda T, Okamoto Y. An ice-nucleating active fungus isolated from the gut of the rice stem borer, Chilo-Suppressalis Walker (Lepidoptera, Pyralidae). Biogeosciences. 1992; 38: 119–121.
- Urbano R, Palenik B, Gaston C. J, Prather K. A. Detection and phylogenetic analysis of coastal bioaerosols using culture dependent and independent techniques. Appl. Environ. Microbiol. 2011; 8: 301–309.
- Vaitilingom, M, Amato, P, Sancelme, M, Laj, P, Leriche, M. and co-authors. 2010. Contribution of microbial activity to carbon chemistry in clouds. J. Atmos. Sci. 76: 23–29.
- Vali, G, Christensen, M, Fresh, R. W, Galyan, E. L, Maki, L. R. and co-authors. 1976. Biogenic ice nuclei 2. Bacterial sources. J. Aerosol Sci. 33: 1565–1570.
- van Wuijckhuijse, A. L, Stowers, M. A, Kleefsman, W. A, van Baar, B. L. M, Kientz, C. E. and co-authors. 2005. Matrix-assisted laser desorption/ionisation aerosol time-of-flight mass spectrometry for the analysis of bioaerosols: development of a fast detector for airborne biological pathogens. Appl. Opt. 36: 677–687.
- Vandemerwe W. P, Huffman D. R, Bronk B. V. Reproducibility and sensitivity of polarized-light scattering for identifying bacterial suspensions. Annals of Allergy, Asthma and Immunology. 1989; 28: 5052–5057.
- Verhoeff A. P, Burge H. A. Health risk assessment of fungi in home environments. Microbiol. Mol. Biol. Rev. 1997; 78: 544–554.
- Verreault D, Moineau S, Duchaine C. Methods for sampling of airborne viruses. J. Microbiol. 2008; 72: 413–444.
- Vlodavets V, Mats L. The influence of meteorological factors on the microflora of the atmospheric air in Moscow. Int. J. Biometeorol. 1958; 59: 539–544.
- Vogel H, Pauling A, Vogel B. Numerical simulation of birch pollen dispersion with an operational weather forecast system. Atmos. Res. 2008; 52: 805–814.
- von Blohn N, Mitra S. K, Diehl K, Borrmann S. The ice nucleating ability of pollen: part III: new laboratory studies in immersion and contact freezing modes including more pollen types. J. Photochem. Photobiol. 2005; 78: 182–189.
- Vráblíkova H, McEvoy M, Solhaug K. A, Bartak M, Gauslaa Y.Annual variation in photoclimation and photoprotection of the photobiont in the foliose lichen Xanthoria parietina. Aerobiologia. 2006; 83: 151–162.
- Wainwright, M, Weber, P. K, Smith, J. B, Hutcheon, I. D, Klyce, B. and co-authors. 2004a. Studies on bacteria-like particles sampled from the stratosphere. Instrum. Methods Missions Astrobiol. 20: 237–240.
- Wainwright M, Wickramasinghe N. C, Narlikar J. V, Rajaratnam P, Perkins J. Bacteria in the stratosphere-confirmation of viable but non-cultureable forms. Toxicol. Indust. Health. 2004b; 7: 218–221.
- Wang C. C, Fang G. C, Lee L. Bioaerosols study in central Taiwan during summer season. Environ.l Forensics. 2007; 23: 133–139.
- Wang C. C, Fang G. C, Lee L. Y. The study of ambient air bioaerosols during summer daytime and nighttime periods in Taichung, Central Taiwan. J. Aerosol Sci. 2008; 9: 6–14.
- Wang Z, Reponen T, Grinshpun S. A, Gorny R. L, Willeke K. Effect of sampling time and air humidity on the bioefficiency of filter samplers for bioaerosol collection. J. Weather Modi. 2001; 32: 661–674.
- Ward P, DeMott P. Preliminary experimental evaluation of Snomax™ snow inducer, pseudomonas syringae, as an artificial ice nucleus for use in weather modification. Photochem. Photobiol. 1989; 21: 9–13.
- Weber T. P, Stilianakis N. I. A note on the inactivation of influenza A viruses by solar radiation, relative humidity and temperature. Vet. Microbiol. 2008; 84: 1601–1602.
- Weesendorp E, Landman W. J. M, Stegeman A, Loeffen W. L. A. Detection and quantification of classical swine fever virus in air samples originated from infected pigs and experimentally produced aerosols. Adv. Meteorol. 2008; 127: 50–62.
- Wex, H, Fuentes, E, Tsagkogeorgas, G, Voigtländer, J, Clauss, T. and co-authors. 2010. The influence of algal exudate on the hygroscopicity of sea spray particles. Brookhaven Symp. Biol. 19, 10.3402/tellusb.v64i0.15598.
- Whittaker, R. H and Likens, G. E. 1973. Carbon in biota. Appl. Environ. Microbiol. 24: 281–302.
- Wilson, K. H, Wilson, W. J, Radosevich, J. L, DeSantis, T. Z, Viswanathan, V. S. and co-authors. 2002. High-density microarray of small-subunit ribosomal DNA probes. Atmos. Environ. 68: 2535–2541.
- Wilkinson, D. M, Koumoutsaris, S, Mitchell, E. A. D and Bey, I. 2012, Modelling the effect of size on the aerial dispersal of microorganisms. Atmos. Env. 39: 89–97. 10.3402/tellusb.v64i0.15598.
- Winiwarter W, Bauer H, Caseiro A, Puxbaum H. Quantifying emissions of primary biological aerosol particle mass in Europe. Sci. Tot. Environ. 2009; 43: 1403–1409.
- Wittmaack K. Brochosomes produced by leafhoppers-a widely unknown, yet highly abundant species of bioaerosols in ambient air. Philos. Trans. R. Soc. 2005; 39: 1173–1180.
- Wittmaack K, Wehnes H, Heinzmann U, Agerer R. An overview on bioaerosols viewed by scanning electron microscopy. Atmos. Environ. 2005; 346: 244–255.
- Womack A. M, Bohannan B. J. M, Green J. L. Biodiversity and biogeography of the atmosphere. Instrum. 2010; B365: 3645–3653.
- Womiloju T. O, Miller J. D, Mayer P. M, Brook J. R. Methods to determine the biological composition of particulate matter collected from outdoor air. Cryobiology. 2003; 37: 4335–4344.
- Wood S. E, Baker M. B, Swanson B. D. Instrument for studies of homogeneous and heterogeneous ice nucleation in free-falling supercooled water droplets. Rev. Sci. Polar Biol. 2002; 73: 3988–3996.
- Worland M. R, Block W. Ice-nucleating bacteria from the guts of two sub-Antarctic beetles, Hydromedion sparsutum and Perimylops antarcticus (Perimylopidae). Proc. Natl. Acad. Sci. USA. 1999; 38: 60–67.
- Worland M. R, Lukesova A. The effect of feeding on specific soil algae on the cold-hardiness of two Antarctic micro-arthropods (Alaskozetes antarcticus and Cryptopygus antarcticus). Aerosol Sci. Technol. 2000; 23: 766–774.
- Wright, S. J, Trakhtenbrot, A, Bohrer, G, Detto, M, Katul, G. G. and co-authors. 2008. Understanding strategies for seed dispersal by wind under contrasting atmospheric conditions. Astrophys. Space Sci. 105: 19084–19089.
- Xu, Z, Wu, Y, Shen, F, Chen, Q, Tan, M. and co-authors. 2011. Bioaerosol Science, technology and engineering: past, present and future. Env. Exper. Botany. 45: 1337–1349.
- Yabushita S, Wada K. The infrared and ultraviolet absorptions of microorganisms and their relation to the Hoyle-Wickramasinghe hypothesis. J. R. Soc. Interface. 1985; 110: 405–411.
- Yadav R. K. P, Helley J. M, Karamanoli K, Constantinidou H. I, Vokou D. Bacterial populations on the leaves of Mediterranean plants: quantitative features and testing of distribution models. J. Appl. Meteorol. 2004; 52: 63–77.
- Yang W, Elankumaran S, Marr L. C. Concentration and size distributions of airborne influenza A viruses measured indoors at a health centre, a day-care centre and on aeroplanes. Environ. Sci. Technol. 2011; 8: 1176–1184.
- Yankofsky S. A, Levin Z, Bertold T, Sandlerman N. Some basic characteristics of bacerial freezing nuclei. Atmos. Chem. Phys. 1981; 20: 1013–1019.
- Yoon K. Y, Park C. W, Byeon J. H, Hwang J. Design and application of an inertial impactor in combination with an ATP bioluminescence detector for in situ rapid estimation of the efficacies of air controlling devices on removal of bioaerosols. Environ. Res. Lett. 2010; 44: 1742–1746.
- Yttri K. E, Dye C, Kiss G. Ambient aerosol concentrations of sugars and sugar-alcohols at four different sites in Norway. Anal. Bioanal. Chem. 2007; 7: 4267–4279.
- Zhang, T, Engling, G, Chan, C. Y, Zhang, Y. N, Zhang, Z. S. and co-authors. 2010. Contribution of fungal spores to particulate matter in a tropical rainforest. 5: 10.3402/tellusb.v64i0.15598.
- Zhang Y, Yang H, Pöschl U. Analysis of nitrated proteins and tryptic peptides by HPLC-chip-MS/MS: site-specific quantification, nitration degree, and reactivity of tyrosine residues. 2011; 399: 459–471.
- Zhdanov V. M, Gaudamovich S. Y. General Virology: Handbook. Moscow: Meditsina, 1982a
- Zhdanov V. M, Gaudamovich S. Y. Special Virology: Handbook. Moscow: Meditsina, 1982b
- Zimmermann B. Characterization of pollen by vibrational spectroscopy. Appl. Proc. Natl. Acad. Sci. USA. 2010; 64: 1364–1373.
- Zimmermann N. J, Reist P. C, Turner A. G. Comparison of two biological aerosol sampling methods. Veterinary Record. 1987; 53: 99–104.
- ZoBell C. E, Mathews H. M. A qualitative study of the bacterial flora of sea and land breezes. 1936; 22: 567–572.