Abstract
Two forms of ide Leuciscus idus (L.) spawners: wild-coloured and ornamental: yellow-coloured were kept at three various temperature regimes shortly before spawning at optimal temperature regimes (group 1), under natural temperature conditions (group 2) and in rapidly increasing temperature (group 3). The quality and quantity of collected semen, ovulation rate and survival rate of embryos to the eyed-egg-stage were recorded. The quality of semen from group 3 (where the temperature increased over the thermal spawning optimum) was the worst (46 and 51% motility of spermatozoa for the wild-coloured and yellow form, respectively). The quantity of collected semen also was the lowest in the same groups (1.1 and 1.0 cm3 kg−1 for the wild-coloured and yellow form, respectively). Increasing the temperature to 16°C also caused a decreasing percentage of ovulated females (70% and 60% of ovulation for wild-coloured and yellow form, respectively) and biological quality of eggs (48.9 and 47.8% embryo survival for wild-coloured and yellow form, respectively). Fluctuations of temperature at a level of 8–14°C (group 2) did not negatively affect spawning results, except for a longer latency time (over 44 hrs). The results suggest that the temperature regime shortly before controlled reproduction of ide plays an important role influencing reproductive success.
Introduction
In recent years, many fish populations have become endangered or vulnerable. This has mostly been caused by anthropogenic factors where many valuable biotopes were transformed, especially rivers which are substantially changed by regulations and dam construction (CitationPenczak and Kruk, 2000). It has caused the degradation of many habitats suitable for spawning grounds and larvae growth. Moreover, with ongoing climate change it is now much more difficult to predict the potential effect of unpredictable thermal fluctuations in the wild. This especially concerns freshwaters, where the temperature is much more dependent on atmospheric conditions and all irregularities have a very strong impact on the fish life cycle.
Sustainable aquaculture plays a very important role in the preservation of biodiversity in open waters (CitationRoss et al., 2008). It is a major branch of restitution programmes, where millions of fry are restocked every year. One of the bottlenecks in this area is controlled reproduction and its synchronization (CitationTargońska et al., 2010;Citation2011), which is an essential condition for further effective activities, including controlled rearing. The synchronization of ovulation under controlled conditions in many fish species is possible, mainly through hormonal treatment. Many fish were artificially reproduced using hormonal stimulation. In freshwater cyprinid reproduction, many kinds of spawning agents were applied, i.e. common carp Cyprinus carpio (L.) pituitary homogenate, hCG or GnRH analogues combined with dopamine antagonist (CitationKrejszeff et al., 2009).
Ide Leuciscus idus (L.) has a huge economic value in Poland, as well as many other European countries, because its production is very high in comparison to other rheophilic cyprinids and it is produced as a market and sport fish. In addition, the ornamental form of this species (golden orfe) is also cultured. For example, between 2000 and 2002, the production of ide summer fry for restocking was about 4,700,000 specimens. One-year-old fish production (2004) was about 27,000 kg, which is 69% and 91% of the total production of riverine cyprinids for 2000 and 2002, respectively, including barbel, Barbus barbus (L.), asp Aspius aspius (L.), chub Leuciscus cephalus (L.), nase, Chondrostoma nasus (L.) and vimba bream, Vimba vimba (L.). Ide production was based on artificial reproduction, in which many different hormonal stimulants were tested. Various differences in the spawning results of wild and domesticated stock have been described (CitationKrejszeff et al., 2009).
Temperature plays an important role throughout a fish’s life, including cyprinids. This is based on many studies into the effect of temperature on fish (CitationKucharczyk et al., 1998; CitationDavies and Bromage, 2002; CitationHilder and Pankhurst, 2003; CitationAngius and Canavate, 2005; CitationDaufresne and Boët, 2007; CitationKupren et al., 2011). This is clearly visible during the most important phase of fish development: e.g. embryonic (CitationKucharczyk et al., 1997; CitationKupren et al., 2010;Citation2011) or larval development (CitationKucharczyk et al., 1998; CitationKupren et al., 2010;Citation2011). The effect of temperature is species-dependent, which is caused by its thermal preferences. In addition, temperature is one of the most important factors during finale gametes’ maturation and spawning. Ide is a species which usually spawn at a temperature of about 10 – 14°C during early spring (CitationMann, 1996; CitationKupren et al., 2011) when water temperatures are rising. Due to this, ide spawners should be kept at this temperature regime before controlled reproduction. In a typical protocol for applying hormonal stimulation of this species, the temperature increased slightly from 10–11°C to 12°C after the primary injection and up to 13.5–14.5°C after the resolving injection (CitationKucharczyk et al., 2008; CitationKrejszeff et al., 2009; CitationŻarski et al., 2009). However, the temperature effect on spawning success in ide has not yet been considered. Personal observations, conducted on fish farms over several spawning seasons indicate that high and rapid changes in temperatures affected spawning effectiveness in rheophilic cyprinids, i.e. ide, dace (Leuciscus leuciscus) and chub, defined as gamete quality and the percentage of ovulation and spermiation. A similar situation was also noted for asp (CitationTargońska et al., 2010) and burbot (Lota lota) (CitationŻarski et al., 2010). These changes were usually caused by a rapid change of weather during spawning and incubation periods which influenced the water temperature of rivers. These rivers are very often the main source of water for commercial hatcheries where spawners are kept before spawning at this time. In the present paper, the influence of different temperature regimes (including short-term increases shortly before hormonal induction of spawning) was studied on wild and cultured ide.
Materials and methods
Ide spawners (wild form) were obtained from lakes and rivers in Warmia (north-eastern Poland) a few months before the natural spawning period. The collected fish were kept in a pond on the Janowo Fish Farm near Szczytno (north-eastern Poland). The golden form (cultured) of ide (orfe) were collected from the Oleśnica Fish Farm near Piła (southwestern Poland). Both forms were kept under natural photo-thermal conditions during the winter. At the beginning of the spawning season, fish from both stocks were selected according to the following criteria: the belly of the females had to be fully distended and bulging, soft and resilient to touch; males had to be slightly spermiating (CitationKucharczyk et al., 2008). The size of the wild spawners ranged from 0.8 to 1.2 kg, whereas the cultured fish size was much smaller (0.2–0.3 kg). All fish were individually marked (using floy tags), weighed and oocytes were taken from females using a catheter. The oocytes were sampled in vivo and placed in Serra’s solution for five minutes.
After clarification of the cytoplasm, the position of the germinal vesicle was determined according to a four-stage scale described by CitationBrzuska (1979) for common carp.
Following the clarification of the cytoplasm, the position of the germinal vesicle was determined in oocytes according to a four-degree scale:
stage 1 - germinal vesicle in the central position
stage 2 - early migration of germinal vesicle (less than half of the radius)
stage 3 - late migration of germinal vesicle (more than half of the radius)
stage 4 - periphery germinal vesicle or germinal vesicle breakdown (GVBD)
For further experiments, only females with oocytes at stage 2 or 3 were taken for the spawning experiment. The number of males and females in all groups was 10. There were no statistical differences between the weights of fish from the different origin groups: wild (average weight 982±86 g) and cultured yellow-coloured (average weight 223±29 g) fish. The selected males and females from each group were kept in a hatchery in separate 1000 dm3 tanks with controlled temperature and photoperiod. The maximum fish load was 25 kg m−3. Fish from both stocks were randomly divided into three (wild fish) or two groups (cultured fish). All fish were kept in the same type of tanks. In each group, a different thermal regime before spawning was tested:
Group 1: wild and cultured fish, were kept in a model temperature regime for ide artificial reproduction (hatchery of Department of Lake and River Fisheries - wild and yellow form of ide) (CitationKucharczyk et al., 2008). The temperature was gradually increased from 10 to 14°C ().
Figure 1 Temperature changes during the experiment. Arrow indicates the moment of applying hormonal stimulation. 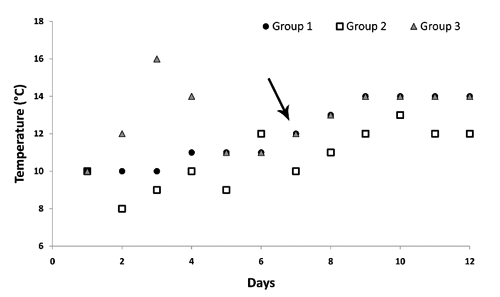
Group 2: wild form only, which were kept indoors under natural temperature conditions (hatchery Fish Farm Janowo); water was taken directly from the Sawica River. The temperature was variable and oscillated between 8 and 14°C ().
Group 3: fish (both forms) were kept initially in rapidly rising temperature from 10 to 16°C (during the first three days) (hatchery of the Department of Lake and River Fisheries - both colour form of ide). Three days before hormonal stimulation, the temperature was set according to group 1.
All of the thermal regimes applied are presented in . The photoperiod in all groups was the same during the experiment: 12 h light - 12 h dark. The oxygen level in tanks throughout the experiment was over 6 ppm and pH ranged from 7.4 to 7.7. The final gametes’ maturation was achieved using hormonal stimulation. Injections with Ovopel (Unic-trade, Budpest, Hungary) (one pellet contains 18–20 µg a mammalian GnRHa and 810 mg dopamine antagonist: metoclopramide) were applied intraperitoneally at the base of the ventral fin. The spawning agent was used in two successive doses: 0.2 and 1.0 pellet kg−1 given 24 h apart. Before manipulations, ripe gamete donors were anaesthetized in a solution of 2-phenoxyethanol (0.5 mg dm−3) (Sigma-Aldrich, Munich, Germany). Semen was stripped using a catheter to avoid urine and collected with plastic syringes (±0.05 cm3) and kept at 4°C before further treatment. The volume of semen collected from each male was recorded. The sperm motility was recognized subjectively under a light microscope (500 X). The sperm was diluted in 0.5% NaCl solution in a 1:1 ratio. Females were checked every three hours between 30 and 48 h after the second injection. Eggs were collected to plastic vessels individually. Eggs portions with at least 10% of white, dead eggs were noted as over-matured. For egg fertilization, only sperm samples showing more than 70% spermatozoa motility were taken for egg fertilization. Eggs samples were placed on Petri dishes. Three egg samples (150–200 eggs each) from each female were mixed with 0.05 mL of pooled semen sample collected from a minimum of 5 males. Later, 5 cm3 of water (15°C) was added to each sample to the gametes for their activation. After 3 min, the water was changed. Eggs were incubated at 15°C, which was the best temperature for ide embryonic development (CitationKucharczyk et al., 2008). Oocyte samples from non-ovulated females were taken with a catheter after the experiment and their maturity stage was recorded.
Survival of the embryos was checked at the eyed-egg-stage (7th day of incubation). Statistical differences between groups (incubation success, semen characteristics) were analyzed with an analysis of variance (ANOVA) and post-hoc Tukey’s test (P<0.05) for wild-coloured ide and a t-test for orfe (P<0.05). Normal distribution of the data was checked using a Shapiro-Wilk test. Statistical analysis was done using Statistica 8.0 (StatSoft) software.
Results
In the present study, differences in percentage of spermiation, ovulation, semen quantity and gamete quality were observed ( and ) in relation to temperature. The percentage of spermiation in males from group 3 was the lowest, as well as the sperm volume and spermatozoa motility in both cases of wild and cultured fish. The results obtained in group 3 were statistically different from other groups.
Table 1 Results of the artificial propagation of wild-coloured ide in various temperatures.
Table 2 Results of the artificial propagation of yellow coloured ide (orfe) in various temperatures.
The obtained results of female reproduction also showed the impact of temperature on successful reproduction. The percentage of ovulated females was different in all groups ( and ). The highest was observed in group 1, where eggs were stripped from all females. In ide, there were no differences in the quality of eggs in groups 1 and 2, expressed as embryo survival. The lowest percentage of ovulated females was noted in group 3 (70% and 60% for wild- and yellow-coloured form respectively). In the same group in the wild-coloured form, over 40% of the obtained eggs (for 3 from 7 females) were of very low quality: the eggs were over-matured and they were not used for the fertilization process. The percentage of ovulation and quality of eggs, as well as the embryo survival from the eggs obtained (and fertilised) from the rest of the females in this group was the lowest. However, the latency time was the shortest in this group (32–36 h), but was close to group 1. Oocytes from non-ovulated females were in stages 3 and 4 for fish from group 2 and during resorption in fish from group 3.
Discussion
The results obtained in this study confirmed the influence of temperature on final oocyte maturation and spawning effectiveness in fish. Because other environmental parameters, such as photoperiod, dissolved oxygen level or pH might mask the influence of temperature on successful spawning, all measured environmental factors were similar (as well as the maturation stage of spawners) in order to demonstrate the decisive effect of temperature. Reproductive processes in fish (defined in aquaculture as the percentage of spermiated and ovulated spawners, gametes quality and quantity) is controlled by environmental factors, among which the photoperiod and water temperature are the most important (CitationStacey, 1984). In many teleost fish, the photoperiod is the most important factor in the reproductive cycle (CitationBromage et al., 2001) and the temperature is mainly important in the final gamete maturation, ovulation and spawning (CitationHilder and Pankhurst, 2003; CitationAngius and Canavate, 2005; CitationTargońska et al., 2010; CitationŻarski et al., 2010). In rheophilic cyprinid fish, such as asp, dace, ide or chub, shortly before spawning under controlled conditions, the temperature usually increases slightly (CitationKucharczyk et al., 2008; CitationKrejszeff et al., 2009). This suggests an important role of temperature in final gamete maturation. Temperature not only strongly influences the process of reproduction in fish (CitationBromage et al., 2001), but also the gamete quality (CitationHilder and Pankhurst , 2003; CitationKupren et al., 2010, Citation2011).
During artificial reproduction of ide, similar results as in the case of group 3 were noted in out-of-season reproduction of ide (CitationKucharczyk et al., 2008). The obtained results suggest that only four days of increasing temperature before reproduction might have a strong influence on spawning effectiveness. In spiny dam-selfish, Acanthochromis polyacanthus (Bleeker) only three days of rising temperature was enough to affect spawning results (CitationHilder and Pankhurst, 2003). A similar situation was noted for a domesticated strain of ide (CitationKrejszeff et al., 2009). Generally in cyprinids, females without hormonal stimulation failed to ovulate and their oocytes did not mature. A similar observation was made for asp, common bream, chub, dace, nase, ide, common carp and many other fish species (CitationKrejszeff et al., 2009; Targońska et al., 2010). This indicates that stimulation with environmental conditions alone, without hormonal stimulation, is not enough to cause final gamete maturation and ovulation under controlled conditions. In the present study, the results of the reproduction of ide males were similar to those obtained earlier during the spawning season (CitationKucharczyk et al., 2008), although males kept under natural temperature conditions produced a smaller volume of semen (group 2). In addition, there were no differences between either forms of ide (wild or cultured), although in other cases some differences during artificial reproduction between wild and domesticated strain were recorded (CitationKrejszeff et al., 2009). Temperature-dependent reproduction effectiveness of ide females was confirmed by the data in this study. The data are quite similar to these obtained earlier (CitationKucharczyk et al., 2008) for both in- and out-of-season ide spawning. In captivity, the strong influence of temperature on artificial reproduction was also noted for spiny damselfish (CitationHilder and Pankhurst, 2003) and Senegal sole, Solea senegalensis (CitationAngius and Canavate, 2005). In the case of the latter species, keeping the fish under natural temperature conditions caused a longer reproduction period. In the present study, keeping the fish under natural conditions only influenced elongation of the latency time and asynchronous ovulation, but did not decrease the biological quality of eggs. A similar observation was made by Targońska et al. (unpublished) for dace. In the present work, temperature fluctuations up to the highest tested level occurred at decreased levels of all measured parameters. The same tendency, with a much stronger decrease in the percentages of ovulation and embryo survival, was noted for asp after a sudden increase in temperature during the spawning season (CitationTargońska et al., 2010).
Conclusions
The present study shows that the temperature regime in ide artificial reproduction should be kept as optimal (CitationKucharczyk et al., 2008), especially for artificial reproduction of wild populations (CitationŻarski et al., 2009). A decrease in temperature to the level of 12°C or below caused only by longer latency time along with an increase in temperature to about 16 °C decreased the number of ovulated females and quality of gametes. Rapid changes in water temperature or an excessively high pre-spawning temperature probably cause overly rapid gamete maturation and adversely affects their quality. The temperature shortly before reproduction should not exceed the thermal optimum for each species. For example, it was about 13°C for asp (CitationTargońska et al., 2010) and about 16°C for ide (CitationKucharczyk et al., 2008; present paper). It creates the necessity for much more attentive observations on the influence of temperature on the reproductive performance of various fish species during controlled reproduction.
Acknowledgments:
this research was financed under UWM project No. 528-0808-0801 and project “Innovations in finfish aquaculture with special references to reproduction”, Operational Programme “Sustainable Development of the Fisheries Sector and Coastal Fishing Areas 2007-2013” (OR14-61724-OR1400003/09/10/11).
References
- AngiusV. CanavateJ.P. 2005 Spawning of captive Senegal sole (Solea senegalensis) under a naturally fluctuating temperature regime Aquaculture 243 133 145
- BromageN. PorterM. RandallC. 2001 The environmental regulation of maturation in farmed finfish with special reference to the role of photoperiod and melatonin Aquaculture 197 63 98
- BrzuskaE. 1979 The in vivo method of estimating the stages of oocyte maturation in carp Cyprinus carpio L Acta Hydrobiol 21 423 433
- DaufresneM. BoëtP. 2007 Climate change impacts on structure and diversity of fish communities in rivers Global Change Biol 13 2467 2478
- DaviesB. BromageN. 2002 The effects of fluctuating seasonal and constant water temperatures on the photoperiodic advancement of reproduction in female rainbow trout, Oncorhynchus mykiss Aquaculture 205 183 200
- HilderM.L. PankhurstW. 2003 Evidence that temperature change cues reproductive development in the spiny damselfish, Acanthochromis polycanthus Environ. Biol. Fish 66 187 196
- KrejszeffS. TargońskaK. ŻarskiD. KucharczykD. 2009 Domestication affects spawning of the ide (Leuciscus idus) — preliminary study Aquaculture 295 145 147
- KucharczykD. LuczynskiM. KujawaR. CzerkiesP. 1997 Effect of temperature on embryonic and larval development of bream (Abramis brama L.) Aquat. Sci 59 214 224
- KucharczykD. LuczynskiM. KujawaR. KaminskiR. UlikowskiD. BrzuzanP. 1998 Influences of temperature and food on early development of bream (Abramis brama L.) Arch. Hydrobiol 141 243 256
- KucharczykD. TargońskaK. ŻarskiD. KujawaR. MamcarzA. 2008 A review of the reproduction biotechnology for fish from the genus Leuciscus Arch. Pol. Fish 16 319 340
- KuprenK. MamcarzA. KucharczykD. 2010 Effects of temperature on survival, deformation rate and selected parameters of newly hatched larvae of three rheophilic cyprinids (genus Leuciscus) Pol. J. Nat.Sci 25 299 312
- KuprenK. MamcarzA. KucharczykD. 2011 Effect of variable and constant thermal conditions on embryonic and early larval development of fish from the genus Leuciscus (Cyprinidae, Teleostei) Czech J. Anim. Sci 56 70 80
- MannR.H.K. 1996 Environmental requirements of European non-salmonid fish in rivers Hydrobiologia 323 223 235
- PenczakT. KrukA. 2000 Threatened obligatory riverine fishes in human modified Polish Rivers Ecol. Freshw. Fish 9 109 117
- RossL.G. Martinez PalaciosC.A. MoralesE.J. 2008 Developing native fish species for aquaculture, the interacting demands of biodiversity, sustainable aquaculture and livelihoods Aquac. Res 39 675 683
- StaceyN.E. 1984 Control of the timing of ovulation by exogenous and endogenous factors PottsG.W. WoottonR.J. Fish Reproduction: Strategies and Tactics Academic Press London, UK 207 222
- TargońskaK. KucharczykD. KujawaR. MamcarzA. ŻarskiD. 2010 Controlled reproduction of asp, Aspius aspius (L.) using luteinizing hormone releasing hormone (LHRH) analogues with dopamine inhibitors Aquaculture 306 407 410
- TargońskaK. KucharczykD. ŻarskiD. CejkoB.I. KrejszeffS KuprenK. KrólR. DrylK. KowalskiR.K. GlogowskiJ. 2011 Artificial reproduction of wild and cultured barbel (Barbus barbus, Cyprinidae) under controlled conditions Acta Vet. Hung 59 363 372
- ŻarskiD. KucharczykD. SasinowskiW. TargońskaK. MamcarzA. 2010 The influence of temperature on successful reproduction of burbot Lota lota L. under the hatchery conditions Pol. J. Nat. Sci 25 93 105
- ŻarskiD. KucharczykD. TargońskaK. JamrózM. KrejszeffS. MamcarzA. 2009 Application of Ovopel, Ovaprim and their combination in artificial reproduction of two rheophilic cyprinid fishes Pol. J. Nat. Sci 24 235 244