Abstract
The aim of this study was to estimate the requirement for digestible lysine for broilers from 35 to 49 days of age. Two hundred and forty chicks with a mean weight of 44±1 g were used in a completely randomized design, made up of male and female chicks and 6 digestible lysine levels. Experimental diets were formulated to be isoenergetic and isonitrogenous. Fitted broken lines on different responses indicated break points at 0.93, 0.93 (for body weight), 0.98, and 0.92 for feed conversion ratio for male and female, respectively. The results showed that the digestible lysine requirement of male broilers for maximum breast yield percentage, plasma free lysine and antibody titer against Newcastle disease virus exceeded the range of lysine levels tested. Dietary lysine had a significant effect in increasing the plasma free lysine, albumin, total protein, immunoglobulin, antibody titer against sheep red blood cell, Newcastle disease virus and heterophil to lymphocyte ratio. In conclusion, lysine requirements of broilers for performance were lower than breast yield percentage and immune responses. Broken-line analysis showed that the concentrations of plasma free lysine were useful physiological indicators for determining the digestible lysine requirement of male and female broilers. Our results suggest that the estimated requirements based on exponential response curves were higher than estimated requirements obtained using a broken-line model.
Introduction
Genetic selection by primary breeding companies has resulted in greatly improved growth rate and breast meat yield of broiler chickens (CitationDozier et al., 2008a,Citationb). Therefore, today broilers require higher dietary amino acid concentrations to optimize performance and breast meat yield than in the past (CitationDozier et al., 2008b).
Market weights of broiler chickens have increased to meet the demand for breast fillets and value-added products. Breast meat is relatively high in lysine (Lys) (7%) compared with other amino acids (AA). AAs/crude protein has a strong impact on the cost of broiler diet; diet costs represent approximately 65% of total live production costs. Formulating broiler diets on a digestible AA basis and utilizing the economically advantageous commercial AA supplements (methionine, Lys and threonine sources) result in diets with marginally less crude protein that support equal broiler growth compared to broilers fed diets containing higher crude protein with excess AAs. Digestible AA values are more beneficial than requirements expressed on a total basis. Lys is used as the reference AA for the ideal AA profile because more information is known about its concentration in feedstuffs. Lys is also the second-limiting AA in poultry diets (after methionine) its main purpose being protein synthesis (CitationBaker, 1994). CitationAyasan and Okan (2011) randomly assigned broilers to five dietary treatment groups: 0.70 (control diet), 0.75, 0.80, 0.85 and 0.90% of threonine. The results obtained showed that the highest body weight gain occurred at 0.75% threonine; the tendency towards a better feed conversion ratio in the group was reflected with 0.80% threonine for female broilers. Their results suggest that the requirement calculated according to the feed conversion ratio is higher than that calculated according to body weight gain (CitationAysan and Okan, 2011).
As all AAs are in relation to Lys in an ideal ratio, it is important to have an accurate Lys digestible requirement for the group of animals for which the diets are being formulated. If errors are made in the determination of the Lys requirement, all other AAs will also be estimated incorrectly. A review of the literature revealed that no research has been conducted to determine the digestible Lys requirement for broiler chicks based on immune response, breast meat yield and blood parameter. Unfortunately, few data are available on the digestible Lys requirements based on immune response, breast meat yield and blood parameters. This study aimed to determine the digestible Lys requirement for male and female broilers during the finisher period based on body weight, feed conversion ratio, immune responses, breast meat yield and blood parameters. It has become a common practice to include synthetic AAs such as Lys and methionine in rations for young broilers. The supply of optimal digestible Lys levels will reduce feed costs and decrease nitrogen excretion (CitationBaker, 1994; CitationFirman, 2001). CitationMehri et al. (2010) conducted an experiment to determine the digestible lysine requirement. They formulated basal diet for straight-run broilers from 15–28 days of age based on wheat, triticale and corn gluten meal, and provided 6 treatments: 0.60, 0.72, 0.84, 0.96, 1.08, and 1.20%. They estimated digestible lysine requirements by using a linear broken-line 0.95 and 1.08% for body weight gain and feed conversion ratio, respectively. The fundamental difference between our experiment and theirs was the type of response. We estimated digestible lysine requirement based on blood parameter, breast meat yield and immune response.
The primary objective of this research was to compare the determined digestible Lys requirement of male broilers for maximum performance (body weight gain and feed conversion ratio), immune response and breast meat yield in the finisher period using practical diets formulated on a digestible AA basis.
Materials and methods
Birds, experimental design and bird diets
For this experiment, 240 35-day-old broiler chicks were sorted according to gender and randomly assigned to the dietary treatments so that each pen had a similar initial weight (the initial body weight averaged 44±1 g for both male and female chicks) and weight distribution. The birds received the experimental diets consisting of 6 graded levels of digestible Lys. Each dietary treatment had 4 replicates each for male and female chicks. There were 15 birds in each pen in the battery studies. Feed and water was available ad libitum, and a 24-h constant light schedule was maintained. For blood parameters and immune evaluation, 3 birds per pen were randomly selected to be wing-banded. Venous blood samples were collected at the end of the experiment. A corn-soybean meal-corn gluten meal diet was formulated to meet or exceed the CitationNational Research Council (1994) recommendations for all nutrients except Lys (). Graded levels of L-Lys-HCl were added to the diet. Changes in the dietary ingredients were made to ensure that all diets were isoenergetic, isonitrogenous, and equal in electrolyte balance (DCAD).
Table 1 Composition of experimental diets.
Levels of digestible Lys for the experiment were 0.77, 0.84, 0.91, 0.98, 1.05 and 1.12%. The true AA digestibility of the basal diets was determined using CitationRhone-Poulenc feed (1993). Diets were analyzed for nitrogen (the Kjeldahl method) (CitationAOAC, 2006), crude protein (CP), dry matter (DM) (by drying in an oven at 103°C for 8 h) and acid insoluble ash (CitationAOAC, 2006). All values were expressed according to DM. Samples of feed used for laboratory analysis were ground to pass through a 1 mm mesh in a micro-Wiley mill.
Measurement of performance parameters
Individual body weights, feed consumption and feed conversion ratio per pen were determined weekly and at 49 days of age. Body weight gain, feed intake, and feed conversion ratio measurements were taken during the 35–49 day period. No mortality was observed for any of the treatments; therefore, feed intake was not adjusted for mortality to calculate feed conversion ratio.
At the end of the assay, birds were individually weighed. The 3 birds within each pen that were closest to the mean body weight of that pen were killed with CO2, and breast meat (pectoralis major, without skin) was excised and weighed; abdominal fat, including fat surrounding the gizzard, was also removed and weighed (CitationChaiyapoom et al., 2006; CitationDozier et al., 2008a).
At the end of the experiment, on Day 49, all birds were starved for 3 h, and then 3 birds from each pen were killed by cervical dislocation for carcass chemical composition and breast meat yield measurements. Abdominal fat was also weighed. Carcass weight, abdominal fat weight and breast meat weight were expressed as absolute weights and relative to live body weight at processing.
Blood chemical parameters
At 49 days, a blood sample was collected with a heparinized syringe from the wing vein of the chickens that had been wing banded. Blood samples were collected, placed on ice, centrifuged and stored as plasma at -18°C until the time of analysis. Parameters measured were total protein, creatinine, albumin, and globulin ((CitationHiller et al., 1927) uric acid (CitationFossati et al., 1980; CitationDonsbough et al., 2010). Levels were measured spectrophotometrically (Shimadzu, UV 2100, Kyoto, Japan) using commercial kits.
Blood samples were collected in non-heparinized blood collection tubes to obtain the serum levels. The total number of leukocytes in the blood was assessed using a hemocytometer. Blood samples were put into heparinized tubes at the end of the experiment and from each sample, 4 smears were prepared and air-dried. The smears were fixed in glutaraldehyde-acetone solution, pH 4.8 for 3 min at −10°C. Two of the smears were stained with May Grunwald-Giemsa and peripheral blood leuckocyte percentages of the samples were determined by counting 200 leukocytes on each specimen (CitationCampbell, 1988; CitationPost et al., 2003). The results were expressed as percentages. Serum chemical parameters were analyzed using test-kits supplied by Merck (Darmstadt, Germany): total protein and hemoglobin (CitationDacie and Lewis, 1984). Hematocrit was determined by using heparinized capillaries for blood sampling and a micro-hematocrit centrifuge. Heterophils and total lymphocytes were counted microscopically. One hundred cells were counted to each ratio. Heterophil/lymphocyte (H/L) ratios were determined by dividing the number of heterophils by the number of lymphocytes (CitationGross and Siegel, 1983).
Humoral immune response
Antibody titers against sheep red blood cell
For humoral immune response, on Day 49, primary antibody response to sheep erythrocytes was measured in 3 randomly selected birds from each pen. Birds were intravenously injected with a 10% solution of sheep erythrocytes at Day 42. The blood samples from all the sheep red blood cell (SRBC) injected birds were collected on Day 5 post inoculation for HA antibody titer estimation by the microtiter hemagglutination procedure (CitationSiegel and Gross, 1980;CitationDeng et al., 2005).
Antibody titers against Newcastle disease
Birds were vaccinated with commercial NDV La Sota vaccine on Day 4 via eye drop and on Day 21 intramuscularly (inactive vaccine). The dose and vehicle of vaccine was used according to the manufacturer’s instructions. Each bird received the same amount of NDV vaccine. At 49 days of age, blood samples were collected from 3 birds from each pen (CitationNanthakumar et al., 2000). Serum antibodies to NDV were determined by a hemagglutination inhibition test (micromethod) (CitationIsakov et al., 2005). Antibody of serum samples was read as the highest dilution that can inhibit agglutination. Titer is expressed as Geometric Mean Titer (log2) to simplify numerical writing.
Statistical analysis
The statistical analysis was carried out with PROC GLM CitationSAS (2007) with the fixed effect of sex and digestible Lys when significant interaction was also considered. Differences between treatments were tested for significance using the Duncan test. A P value <0.05 was considered significant.
To estimate the digestible AA requirements, multiple regression models (quadratic, exponential and one-slope broken-line; CitationRobbins et al., 2006) were developed:
Exponential:
Y= a + b (1- e -c(x-d))
One-slope broken line:
Y= L + U (R- XLR) when Lys < Lys0, and Y= constant, when Lys > Lys0
where Y = BW, feed conversion ratio, plasma Lys, SRBC, H/L; all variables were analyzed (AT IG, etc.).
a = Y at the basal dietary Lys level;
b = maximum response to supplemental dietary Lys;
c = curvature steepness;
d = Lys content of basal diet (g/kg);
x = dietary Lys content (g/kg).
Values of R2 were calculated to describe the goodness of fit.
Results and discussion
No mortality was observed between Days 35 and 49. There was a significant quadratic, broken-line and exponential response to increasing dietary levels of digestible Lys based on BWG and feed conversion ratio for females (). Subjecting the growth data to broken-line analysis indicated that the digestible Lys requirement for maximum body weight was 0.93% for both males and females (). The requirement for optimum feed conversion ratio was higher in males: 0.98% for males and 0.94% for female broilers (). When comparing males and females, there was some evidence that males and females differed in their digestible Lys requirement estimates. Ninety-five per cent of the asymptote in body weight was achieved at digestible Lys concentrations of 1.036 and 1.009% in males and females, respectively. CitationMehri et al. (2010) estimated digestible lysine requirements by using a linear broken-line 0.95 and 1.08% for body weight gain and feed conversion ratio, respectively (regardless of gender parameter). A higher digestible Lys requirement for FCR than for BWG has been reported (CitationMack et al., 1999; CitationBaker et al., 2002). Body composition and growth differences in the birds used in those studies could explain the discrepancy (CitationKidd et al., 2004). Lys limit protein accretion and increase fat accretion of broilers at low levels of supplementation (CitationMoran and Bilgili, 1990) Another explanation for the effect of Lys on feed conversion ratio is the higher proportion of energy spent for maintenance. The poor growth rate (BW) and feed efficiency of chickens fed the diets including 0.77 and 0.84% digestible Lys at this period, indicated that the diets were, indeed, deficient in Lys. The decrease in performance and efficiency at the highest experimental digestible Lys level is due to less efficient use of AAs above the requirements for protein synthesis (imbalance). In contrast, the lower growth and high FRC at low experimental digestible Lys levels may be attributed to limiting supplies of the essential AAs. These results support the general principle that chick diets should be formulated to provide sufficient amounts of all AA corresponding subtle requirement for protein synthesis (CitationSklan and Plavnik, 2002). Lys and Arg are involved in the release of the growth hormone, insulin like growth factor I (IGF-I) and modulate bone growth by differentiation of osteoblastosis and collagen synthesis. New evidence has been provided by CitationSakomura and Coon (2003) and by CitationNonis and Gous (2008).
Table 2 Results of estimation for the digestible lysine requirement base on different parameters.
The percentages of breast and weight of abdominal fat were significantly affected by digestible Lys levels and breast meat increased with increasing digestible Lys concentration (). The responses suggested that the digestible Lys requirement for maximal breast yield differed greatly from that predicted from the feed efficiency data (). Broken-line analysis suggests that the digestible Lys requirement for optimum body abdominal fat is 0.98 and 0.96% for male and female, respectively () There was a great difference according to sex in the digestible Lys requirement estimates based on breast meat yield (). In order to achieve 95% of the asymptote in breast meat weight in male chicks, digestible Lys need was higher than experimental digestible Lys levels. For optimized BW gain, feed conversion ratio and breast meat yield, male broilers had approximately 2.66, 2.81 and 3.84 g of digestible Lys intake per day, respectively. The amounts of digestible Lys intake necessary to optimize growth performance and breast meat yield of female broilers were 2.19 and 2.57 g per bird per day, respectively. CitationUrdaneta-Rincon et al. (2005) reported that both protein synthesis and breakdown increased at levels of dietary Lys and CP above those required for maximum growth.
Table 3 Effects (interaction sex × lysine) of digestible lysine levels on carcass parameters of male and female broilers.
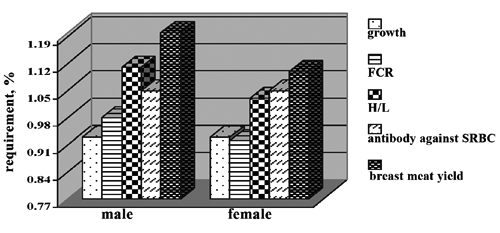
The results of CitationNasr and Kheiri (2011) suggested that additional lysine at a level of 120% of NRC in starter and grower diets optimized body weight gain, carcass and breast percentage in Arian broiler, whereas reductions in lysine level reduced growth and live weight (CitationNasr and Kheiri, 2011; CitationKerd et al., 1998). Breast muscle (BM) constitutes the greatest portion of edible meat in broilers and, depending upon market conditions, it is generally the most valuable part of the carcass. The way in which the yield of BM changes as a bird grows is of considerable importance in deciding the optimal weight to slaughter, estimating accurate nutrient requirements, and evaluating nutritional effects (CitationGous et al., 1999). In recent years, the importance of quality in meat production has increased. In this, some believe that feed is an important factor (CitationAcar et al., 1993; CitationGous et al., 1999; CitationScheuermann et al., 2003) and diet composition (CitationMoran, 1999), especially protein and Lys levels, influences BM yield. It was believed that breast meat was affected by digestible Lys level in diet (CitationFarmer, 1999), but there have not been any reports on the determined requirement of digestible Lys based on breast meat yield. Immune response, including antibody against Newcastle and SRBC, H/L and immunoglobulin (Ig) of both males and females, followed a significant quadratic and broken-line response to increasing dietary levels of digestible Lys (). Males had a higher digestible Lys requirement estimate for maximum antibody titration against Newcastle than experimental digestive Lys levels. However, based on antibody against SRBC, the estimates were similar for both sexes. In females, the estimated digestible Lys requirement by quadratic model based on immune response was higher than that based on breast meat yield ().
Table 4 Comparison of digestible lysine (in percentage) requirement estimate base on performance, blood parameters and immune response.
The plasma Lys concentration data could not be fitted either to a quadratic or exponential response curve. Therefore, an estimation of the digestible Lys requirement for plasma Lys concentration was obtained only by broken-line; although, based on this parameter, digestible Lys requirement estimates of males were slightly higher than that of experimental digestible Lys levels. Regardless of sex, the digestible Lys requirement based on plasma Lys concentration was consistently higher than that based on feed conversion ratio (FCR). Males had higher feed intake, BW, concentration plasma of creatinin and urea ( and ), and breast meat yield than females. From the results of this experiment, there appeared to be more effect of sex on the digestible Lys requirement during this period. Additions of Lys improved plasma Lys concentration and other blood parameters (). Likewise, total protein accretion is correlated with breast muscle yield. Optimizing the AA supply in diet formulation is one of the important tools to restrict wide range in N excretion of animals (CitationFirman, 2001; CitationDozier et al., 2008b; CitationDonsbough et al., 2010). Other serum chemical parameters which are indicative of the protein synthesis capacity of the liver are total protein and the albumin. The low correlation between urea concentration and predicted protein deposition implied that the former was not a useful physiological indicator of the latter (CitationCameron et al., 2003). They also improved immune response (antibody titration against Newcastle, SRBC, H/L and total Ig) and hemoglobin in both sexes (). Determination of serum titers to NDV after regular vaccination is a method which is often used to evaluate immunomodulating effects. There was no significant change in hemotocrit and feed intake according to differing levels of digestible Lys ( and ). The H:L ratio, as an indicator of stress, was found to be highly heritable and under the influence of few genes (CitationAL-Murrani et al., 2002). An inadequate supply of Lys would reduce antibody response and cell-mediated immunity in chickens (CitationGeraert and Mercier, 2010).
Table 5 The effects (interaction sex × lysine) of digestible lysine levels on growth performance of male and female broilers.
Table 6 Effects (interaction sex × lysine) of digestible lysine levels on blood parameters of male and female broilers.
Table 7 Effects (interaction sex × lysine) of digestible lysine levels on blood parameters and immune response of male and female broilers.
Overall, these results indicate that digestible Lys levels required for males were higher for breast meat yield than growth performance and immune response variables ().
Serving as a precursor for protein synthesis, the most significant functions of Lys appear to be as a substrate for the synthesis of nitric oxide, polyamines and various hormones, all of which are capable of modulating immune responses (CitationWu and Morris, 1998). CitationKidd et al. (2001) reported that other than lymphoid organ weights, antibody responses were influenced by dietary Lys and Arginine varying around the CitationNRC (1994) recommended level in commercial broilers. Dietary manipulations may also exert carry-over effects on immune responses in birds.
The higher levels of Lys may be required for the synthesis of Ig antibodies or perhaps for thymus derived T-cell helper function (CitationTsiagbe et al., 1987). The antibody production against SRBC inoculation was increased with the level of digestive Lys in the present study. Similarly, CitationTakahashi et al. (1997) reported significant improvement in the magnitude of antibody production to SRBC antigen. The dose-related antibody titers to SRBC inoculation in broilers, given different levels of Lys and methionine, were also reported by CitationDunnington et al. (1994).
In general the bursa (source of B lymphocytes that produce antibody) responds well to Lys deficiency by increasing its ability to take up Lys from the blood (relative to the abilities of other tissues, such as muscle) and maintains normal cell numbers. In contrast, the thymus (source of T lymphocytes that regulate immune function and kill virus-infected cells) responds poorly, does not augment its ability to compete for Lys, and the number of cells in the thymus decreases. This suggests that animals preserve the antibody-preserving arm of the immune system during malnutrition while the regulatory, T-lymphocyte arm of the immune response is diminished (CitationKlasing, 2007). The requirements of the immune system have been met using a the direct approach by summing up the components of the immune system (cells and accessory proteins) and estimating the amounts of nutrients in them relative to the rest of the body (CitationKlasing, 1998, Citation2003; CitationBarnes et al., 2002). CitationKlasing (2007) estimated that rates of AA incorporation into leukocytes, Igs and accessory proteins indicate that the resting immune system utilizes only about 1.2% of the Lys intake in a healthy growing broiler chick. However, during a robust immune response, production of protective accessory proteins by the liver increases (hepatic acute phase response). These anabolic processes increase nutrient use by the immune system by almost 6-fold in the case of Lys (from 1.2 to 6.7% of Lys intake). For nutrients for which the immune system is most vulnerable due to a low priority for acquisition, requirements set based on maximal weight gain or FCR are likely to be inadequate for optimal disease resistance. Low immune response is possibly due to reduced protein availability for liver protein synthesis associated with immune response or antibody production (CitationMurwani, 2008).
The results of previous experiments by other researchers indicated that the digestible Lys requirements of male broilers with purified diet only achieve maximum body weight gain and feed conversion ratio in the starter period (CitationZaghari et al., 2002). Nevertheless accuracy of estimates is critical for applying data in practice to broiler diet formulation. Therefore, usual feed ingredient was used for practical evaluation of the determined digestible Lys requirements of male and female broilers by comparing the performance of chicks, immune response and breast meat yield.
CitationNRC (1994) recommendations are based on experiments (total requirement) that were, in part, conducted more than 20 years ago, and it is questionable whether they are still applicable to modern breeds. Changes in the rate and composition of growth, namely fat and protein concentration in gained body weight, affect feed conversion ratio and, consequently, the required Lys concentration in the diet. Based on the breast meat yield for the finisher period, we recommend a dietary digestible Lys concentration of 1.10%, or 2.57 g/day and more than 1.12% for female and male, respectively. A comparison with the current CitationNRC (1994) recommendations indicates that the recommendations for the other AAs should be urgently revised in order to adjust and change to digestible requirement for modern genotypes.
In general, the requirement of Lys is higher for increased immunity than for growth. The competition for limited resources may contribute to a negative relationship between growth and immunity. Similarly, several authors (CitationLiu et al., 1995; CitationParmentier et al., 1996) found that body weight and antibody titers are negatively correlated. Also, more immune competent birds have poor nutrient utilization ability. The results from the present study indicate that the Lys requirement for maximum antibody response was greater than for maximum growth for broilers (CitationKlasing, 2007).
Results showing significant differences between sexes in all traits such as immune parameters may result from differences in sex steroid hormones. This hypothesis is supported by the findings of CitationDaynes et al. (1990), who reported that dehydroepiandrosterone (a weak androgen) is an important enhancer of interleukin 2 (IL2) by helper T cells. IL2 is the most powerful growth factor as well as activating factor for lymphocytes. It promotes T-cell proliferation and activates B and T cells (CitationKhajavi et al., 2003).
Conclusions
Based on the results from this study, high-yield male broilers should be fed a minimum of digestible Lys 3.18 g/day (8.5 g/kg) from 35 to 49 days of age. In conclusion, supplementing L-Lys to meet the CitationNRC (1994) recommended requirement for growth selectively decreased serum antibody and Ig levels. This effect probably increased mortality. However, BW, feed conversion ratio, plasma Lys concentration and other blood parameters, such as albumin, total protein, creatinin, uric acid, urea nitrogen, and antibody responses to SRBC were improved by adding digestible Lys level. The underlying mechanisms and the practical implication of these findings in disease resistance merit further studies, especially involving cytokines. Economically, the most important criterion is the breast muscle yield which clearly responded to Lys in our study. A 1% improvement in breast meat yield occurred among the treatments. This signifies an increase in revenue for a broiler company.
References
- AcarN. MoranE.T.Jr. MulvaneyD.R. 1993 Breast muscle development of commercial broilers from hatching to twelve weeks of age Poultry Sci 72 317 325
- AL-MurraniW.K. AL-RawiI.K. RaofN.M. 2002 Genetic resistance to Salmonella typhimurium in two lines of chickens selected as resistant and sensitive on the basis of heterophil/lymphocyte ratio Brit. Poultry Sci 43 501 507
- AOAC 2006 Official Methods of Analysis 18th ed. Association of Official Analytical Chemists Arlington, VA, USA
- AyasanT. OkanF. 2011 Threonine requirement of female broilers from 22 to 42 days of age Süleyman Demirel Üniversitesi Ziraat Fakültesi Dergisi 6 15 21
- BakerD.H. 1994 Ideal Amino acid profile for chicks during the first three weeks posthatching Poultry Sci 73 1441 1447
- BakerD.H. BatalA.B. ParrT.M. AugspurgerN.R. ParsonsC.M. 2002 Ideal ratio (relative to lysine) or tryptophan, threonine, isoleucine and valine for chicks during the second and third weeks post hatch Poultry Sci 81 485 494
- BarnesD.M. SongZ. KlasingK.C. BottjeW. 2002 Protein metabolism during an acute phase response in chickens Amino Acids 22 15 26
- CameronN.D. McCulloughE. TroupK. PenmanJ.C. 2003 Serum urea concentration as a predictor of dietary lysine requirement in selected lines of pigs Anim. Sci 81 91 100
- CampbellT.W. 1988 Avian haematology and cytology Iowa State University Press Ames, IA, USA
- ChaiyapoomB. TakawanS. RatchadapornC. 2006 Effect of adding methionine hydroxy analogue as methionine source at the commercial requirement recommendation on production performance and evidence of ascites syndrome of male broiler chicks fed corn-soybean based Int. J. Poultry Sci 5 744 752
- DacieJ.V. LewisS.M. 1984 Estimation of plasma haemoglobin Practical haematology 6th ed. Churchill Livingstone Ed. London, UK
- DaynesR.A. DudleyD.J. AraneoB.A. 1990 Regulation of marine lymphokine production in vivo 2. Dehydropiandrostron is a natural enhancer of interleukin- 2 synthesis by helper T cell Eur. J. Immunol 20 793 802
- DengK. WongC.W. NolanJ.V. 2005 Long-term effects of early life L-arginine and L-lysine supplementation on growth performance, lymphoid organs and immune responses in Leghorn-type chickens Brit. Poultry Sci 46 318 324
- DonsboughA.L. PowellS. WaguespackA. BidnerT.D. SouthernL.L. 2010 Uric acid, urea, and ammonia concentrations in serum and uric acid concentration in excreta as indicators of amino acid utilization in diets for broilers Poultry Sci 89 287 294
- DozierW.A.3rd CorzoA. KiddM.T. SchillingM.W. 2008a Dietary digestible lysine requirements of male and female broilers from forty-nine to sixty-three days of age Poultry Sci 87 1385 1391
- DozierW.A.3rd KiddM.T. CorzoA. 2008b Dietary amino acid responses of broiler chickens J. Appl. Poultry Res 17 157 167
- DunningtonE.A. ZulkifliI. SiadakD. LarsenA.S. SiegelP.B. 1994 Dietary methionine and antibody response in broiler cockerels Arch. Geflugelkd 58 125 129
- FarmerL.J. 1999 Poultry meat flavor RichardsonR.I. MeadG.C. Poultry Meat Science CAB Int. Wallingford, UK
- FirmanJ.D. 2001 Formulation of diets on a digestible amino acid basis and factors affecting digestibility of feedstuffs. ASA Technical Bulletin No. AN32 Melbourne, Australia
- FossatiP. PrencipeL. BertiG. 1980 Use of 3,5-dichloro-2- hydroxybenzenesulfonic acid/4-aminophenazone chromogenic system in direct enzyme assay of uric acid in serum and urine Clin. Chem 26 227 231
- GeraertP.A. MercierY. 2010 Amino Acids: Beyond the Building Blocks! Available from: http://www.thepoultryfederation.com/public/userfiles/files/3-1%20Thr%20-%20Pierre-Andre%20Geraert%20-%20AA%20Beyond%20Building%20Blocks.pdf
- GousR.M. MoranJ.R. StilbornE.T. BradfordH.R. EmmansG.C. 1999 Evaluation of the parameters needed to describe the overall growth, the chemical growth, and the growth of feathers and breast muscles of broilers Poultry Sci 78 812 821
- GrossW.B. SiegelH.S. 1983 Evaluation of the heterophil/ lymphocyte ratio as a measure of stress in chickens Avian Dis 27 972 979
- HillerA. McIntoshD.F. Van SlykeD.D. 1927 The excretion of albumin and globulin in nephritis J. Clin. Invest 4 235 242
- IsakovN. FeldmannM. SegalS. 2005 The mechanism of modulation of humoral immune responses after injection of mice with SRBC Immunology 128 969 975
- KerdB.J. KiddM.T. HalpinK.M. McWardG.W. QuarlesC.L. 1998 Lysine level increases live performance and breast meat yield in male broilers J. Appl. Poultry Res 8 381 390
- KhajaviM. RahimiS. HassanZ.M. KamaliM.A. MousaviT. 2003 Effect of feed restriction early in life on humoral and cellular immunity of two commercial broiler strains under heat stress conditions Brit. Poultry Sci 44 490 497
- KiddM.T. McDanielC.D. BrantonS.L. MillerE.R. BorenB.B. FancherB.I. 2004 Increasing amino acid density improves live performance and carcass yields of commercial broilers J. Appl. Poultry Res 13 593 604
- KiddM.T. PeeblesE.D. WhitmarshS.K. YeatmanJ.B. WidemanR.F. 2001 Growth and immunity of broiler chicks as affected by dietary arginine Poultry Sci 80 1535 1542
- KlasingK.C. 1998 Nutritional modulation of resistance to infectious diseases Poultry Sci 77 1119 125
- KlasingK.C. 2003 Effect of lysine deficiency on immune response to vaccination against Newcastle disease in broilers Avian Dis 47 1346 1351
- KlasingK.C. 2007 Nutrition and the immune system. Gordon Memorial Lecture. Brit Poultry Sci 48 525 537
- LiuG. DunningtonE.A. SiegelP.B. 1995 Correlated responses to long-term divergent selection for eight-week body weight in chickens: Growth, sexual maturity, and egg production Poultry Sci 74 1259 1268
- MackS. BercoviciD. DegrooteG. LeclercgB. LippensM. PackM. SchutteJ.B. Van CauwenbergheS. 1999 Ideal amino acid profile and dietary lysine specification for broiler chickens of 20 to 40 days of age Brit. Poultry Sci 40 257 265
- MehriM. Nassiri-MoghadamH. KermanshahiH. Danesh-MesgaranM. 2010 Digestible lysine requirements of straight-run broiler chickens from fifteen of twenty-eight days of age J. Anim. Vet. Adv 9 2321 2324
- MoranE.T. 1999 Live production factors influencing yield and quality of poultry meat RichardsonR.I. MeadG.C. Poultry Meat Science CABI Int New York, NY, USA
- MoranE.T. BilgiliS.F. 1990 Processing losses, carcass quality, meat yields of broiler chickens receiving diets marginally deficient to adequate in lysine prior to marketing Poultry Sci 69 702 710
- MurwaniR. 2008 Effect of corn or sorghum in combination with soybean meal or mungbean as feed ingredients on the serum antibody titres to NDV vaccine in broiler chickens Int. J. Poultry Sci 7 497 501
- NanthakumarT. TiwariA.K. KatariaR.S. ButchaiahG. KatariaJ.M. GoswamiP.P. 2000 Sequence analysis of the cleavage site-encoding region of the fusion protein gene of Newcastle disease viruses from India and Nepal Avian. Pathol 29 603 607
- NasrJ. KheiriF. 2011 Effect of different lysine levels on Arian broiler performances Ital. J. Anim. Sci 10 e32
- National Research Council 1994 Nutrient Requirements of Poultry 9th rev. ed. National Academy Press Washington, DC, USA
- NonisM.K. GousR.M. 2008 Threonine and lysine requirements for maintenance in chickens S. Afr. J. Anim. Sci 38 75 82
- ParmentierH.K. NieuwlandM.G.B. RijkeE. de Vries ReilinghG. SchramaJ.W. 1996 Divergent antibody responses to vaccines and divergent body weights of chicken lines selected for high and low humoral responsiveness Avian Dis 40 634 644
- PostJ. RebelJ.M.J. ter HuurneA.A.H.M. 2003 Automated blood cell count: A sensitive and reliable method to study corticosterone-related stress in broilers Poultry Sci 82 591 595
- Rhône-Poulenc Animal Nutrition 1993 Rhodmet Nutrition Guide 2nd ed. Rhône-Poulenc Animal Nutrition Publ Antony Cedex, France
- RobbinsK.R. SaxtonA.M. SouthernL.L. 2006 Estimation of nutrient requirements using broken-line regression analysis Anim. Sci 84 150 155
- SakomuraN.K. CoonN.C. 2003 Amino acid requirements for maintenance of broiler breeder pullets 280 281 in Proc. 14th Eur. Symp. on Poultry Nutrition Lillehammer, Norway
- SAS 2007 User's Guide Statistics Ver. 9.1.3, SAS Inst. Inc. Cary, NC, USA
- ScheuermannG.N. BilgiliS.F. HessJ.B. MulvaneyD.R. 2003 Breast muscle development in commercial broiler chickens Poultry Sci 82 1648 1658
- SiegelP.B. GrossW.B. 1980 Production and persistence of antibodies in chickens to sheep erythrocytes.1. Directional selection Poultry Sci 59 1 5
- SklanD. PlavnikI. 2002 Interactions between dietary crude protein and essential amino acid intake on performance in broilers Brit. Poultry Sci 43 442 449
- TakahashiK. OrtaN. AkibaY. 1997 Influence of dietary methionine and cystine on metabolic responses to immunological stress by Escherichia coli lipopolysaccharide injection and mitogenic response in broiler chickens Brit. J. Nutr 78 815 821
- TsiagbeV.K. CookM.E. HarperA.E. SundeM.L. 1987 Efficacy of cysteine in replacing methionine in the immune responses of broiler chicks Poultry Sci 66 1138 1146
- Urdanneta-RinconM. de LangK. Pena-OrtegaL. LeesonS. 2005 Lysine requirements of young broiler chickens are affected by level of dietary crude protein Can. J. Anim. Sci 85 195 204
- WuG. MorrisS.M.Jr. 1998 Arginine metabolism: nitric oxide and beyond Biochem. J 336 1 17
- ZaghariM. ShivazadM. KamyabA. NikkhahA. 2002 Digestible lysine requirement of arian male and female broiler chicks during the 6-21 days of age J. Agric. Sci. Technol 4 111 117