Abstract
This study aimed at estimating the impact of red deer grazing on the productivity of meadows located in Pian Cansiglio, north-eastern Italian Pre-Alps. These meadows (185 ha; average elevation 1000 m asl are managed for hay/silage production (1–2 cuts per season) and are included in a protected area that hosts a high density of deer (around 30 heads/100 ha). In 2008 and 2010, dry matter (DM) production and loss due to deer grazing were estimated with exclusion cages (1 m2; 48 exclusion cages in 2008 and 52 in 2010). Night counts with spotlights were conducted to index deer use of meadows plots. DM production inside the cages was fairly good for the area (first-second cut: 4963-2297 kg DM/ha in 2008, and 4145-2475 kg DM/ha in 2010). DM production outside the cages was significantly lower (first-second cut in 2008: 4199-1378 kg DM/ha, and in 2010: 3376-2052 Kg DM/ha). Therefore, the magnitude of losses was of 15–20% at the first and 25–40% at the second cut. DM losses in the different meadow plots were positively correlated with index of deer use, which in some plots was as high as 7–8 heads/ha. Deer grazing reduced also crude protein (CP) content of forage (15.6±4.4% DM inside exclusion cages and 13.8±3.5% DM outside), with losses being greater where CP content was higher. This study demonstrates that high densities of grazing deer may seriously impact on forage production and quality.
Introduction
In the last decades, wild red deer (Cervus elaphus Linnaeus, 1758) populations have greatly expanded in range and numbers in Europe and in Italy, as a result of a number of factors including land use and climate changes, protection from poaching and implementation of sustainable harvest plans, and reintroduction programmes (CitationMattioli et al., 2001; CitationMilner et al., 2006; CitationCarnevali et al., 2009). Due to its body size, feed intake, and adaptability to a wide variety of habitats where it may reach high population densities, red deer is considered as a keystone species, i.e. a species that may exert serious impacts on the structure and functioning of agro-forestry ecosystems (CitationCôté et al. 2004; CitationGordon et al., 2004). These impacts include changes in vegetation structure and composition that influence plant richness and biodiversity (CitationRussell et al., 2001; CitationSchütz et al., 2003; CitationGoetsch et al., 2011), and may result in damages to forest regeneration and productivity (CitationAmmer, 1996; CitationMotta, 1996; CitationMotta et al., 2003; CitationTremblay et al., 2007). In addition, they may have cascading effects on the communities of birds (CitationHolt et al., 2011; CitationMartin et al., 2011), invertebrates (CitationRambo and Faeth, 1999; CitationMelis et al., 2005) and other mammals (CitationSmit et al., 2001; CitationMuñoz et al., 2008).
Although it is generally recognised that red deer impacts affect mainly forest ecosystems (CitationPutman and Moore, 1998; CitationRooney and Waller, 2003), the recent expansion of the species resulted in an increased use of cultivated areas, and claims of damages to agricultural crops are rising. Red deer is an intermediate feeder (CitationHofmann, 1989), and as such it tends to browse selectively but has also the ability to graze grass swards (CitationGebert and Verheyden-Tixier, 2001). Therefore, crops damaged may include fruit orchards and woody ornamental plants (CitationCaslick and Decker, 1979; CitationPorter, 1983; Fargione et al., 1991), cereals and oilseed crops (CitationDecalesta and Schwenndeman, 1978; CitationWilson et al., 2009), but also grasslands (CitationTrdan and Vidrih, 2008; CitationWilson et al., 2009; CitationKamei et al., 2010). These damages are linked to the seasonal availability of crops as respect to natural sources of food and to the deer density around and inside crop plots, which in turn is influenced by human interferences as hunting and disturbance, by seasonal displacements, and by the availability of cover (CitationPutman and Moore, 1998; CitationTrdan and Vidrih, 2008;CitationWilson et al., 2009; CitationKamei et al., 2010).
This study aimed at assessing the impact of red deer grazing on grass production of meadows located in a protected area of the eastern Italian Pre-Alps. In this area red deer has increased greatly in the last decade, causing significant damages to forest (CitationCaudullo et al., 2003). Recently, an impact on grasslands has also been suggested (CitationMearns et al., 2007), but a comprehensive assessment is lacking. Therefore, specific objectives were to estimate the extent of the reduction of grass production due to deer grazing, and to verify whether this reduction: i) varied from spring to summer; ii) was related to deer use of meadow plots and to distance from forest edge; iii) resulted in changes of chemical composition of forage.
Materials and methods
Study area
The study was conducted in the Cansiglio forest, located in the north-eastern Italian Pre-Alps (Longitude 12°20’-12°29’E; Latitude 46°01’-46°08’N), between the provinces of Belluno, Treviso and Pordenone (). The Cansiglio Forest covers approximately 7000 ha and is owned by the Italian Government. It has a characteristic bowlshaped morphology, where a central basin lying at an elevation of about 1000 m asl is surrounded by mountain chains (Monte Pizzoc: 1565 m; Monte Millifret: 1581 m; Monte Croseraz: 1694 m; Monte Cavallo: 2250 m). This morphology and karst phenomena are responsible of a thermal inversion. Average annual temperatures are close to 6.8°C, which is 2–3 degrees less than in other pre-Alpine areas at the same elevation (CitationCaudullo et al., 2003). Annual precipitations are abundant and close to 2000 mm (average±SD=1946±476 mm/year, from 1993 to 2010) with highest values in May and November (CitationARPAV, 2011).
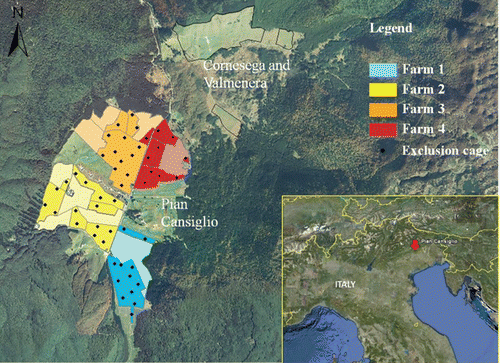
Vegetation in the Cansiglio Forest is mainly beech (Fagus sylvatica L.) dominated woodland, with minor areas of Norway spruce (Picea abies (L.) Karst) and silver fir (Abies alba Miller). In the central basin, three areas, i. e. Pian Cansiglio, Cornesega and Valmenera (), are covered by meadows and pastures for a surface of approximately 383 ha. The whole forest is Site of Community Importance according to directive 92/43/EEC Habitat and Special Protection Area according to directive 79/409/EEC Birds (Natura 2000 identification site code: IT3230077).
Grassland management
The grasslands in Pian Cansiglio are managed by four dairy farms, three of which are organic. The total surface managed is 305 ha, with an average farm size of 81 ha (SD=11). Meadows are in total 185 ha, with an average size per farm of 46 ha (SD=11), and pastures are 120 ha, with an average size per farm of 30 ha (SD=3). During the study period the four farms had a total of 339 livestock units (LU), with an average herd size of 81 LU (SD=31). The average stocking rate (LU/ha) was 1.03 (SD=0.25).
Meadows are normally cut two times per season, the first between mid and late June and the second around mid August. However, some plots are cut only once and then grazed.
Grasslands in Valmenera and Cornesega are managed by a single farm (meadows, 15 ha; pasture, 63 ha), with 34 LU and a stocking rate of 0.47. The few hectares of meadows are cut only once, after July 15, due to limitations imposed for the protection of corncrake (Crex crex) nesting. Due to these particularities, this farm was not included in the study.
According to botanical composition estimated on 79 sample sites (CitationWildi and Orloci, 1996), the meadows in Pian Cansiglio can be assigned to the two groups of high productive meadows. The most represented group cannot be classified as natural according to phytosociological criteria, because it includes mainly few species, as Poa trivialis, Phleum pratense, Festuca pratensis, and Dactylis glomerata, which have a high forage value and result from past actions of re-seeding and high inputs of fertilizers. The second group is less influenced by management practices and can be classified as a transition between Centaureo transalpinae – Trisetum flavescentis (Marshall 1974 corr. CitationPoldini and Oriolo 1994) and Centaureo carniolicae – Arrhenatheretum elatioris (CitationOberdorfer 1964 corr. CitationPoldini and Oriolo 1994). Small, localised areas are affected by Deschampsia caespitosa.
Red deer population
As a State property since 1871, the Cansiglio Forest is a protected area and hunting is forbidden. Red deer in Cansiglio and in all the surrounding areas became extinct in the mid 1800s. The species reappeared in the forest in the late 1980s, when a few individuals escaped from a fenced area (CitationMattioli et al., 2001). The population, at that time demographically isolated, increased slowly and in 1997 the estimated number of red deer was of about 200 heads (CitationStiz, 1997). Since then, also as a result of immigration from the expanding populations of the surrounding areas, the population growth was impressive. In the red deer management unit comprising the forest plus a buffer area for a total of approximately 85 km2, the population size between 2008 and 2009 has been estimated at around 2800–3000 heads, with a population density higher than 30 heads/km2 (CitationBottazzo and Nicoloso, 2010). This density is very high, as compared to that of the species in other Alpine areas (CitationCarnevali et al., 2009), and is favoured by both the protection status and the good habitat suitability of the Cansiglio forest. The seasonal displacements of the population are not known in details. However, population density decreases in winter due to partial migration towards wintering areas at lower elevations, while it increases again in spring and summer, to peak probably during the rutting season between September and October (CitationBottazzo and Nicoloso, 2010). In the forest there are no large predators, and other wild ungulate species include roe deer (Capreolus capreolus, L. 1758) and fallow deer (Cervus dama, L. 1758). In contrast with red deer, for these species there are no recent standardized counts, but population densities are considered much lower than those of red deer.
Data collection
The study was conducted in the summers of 2008 and 2010 on the meadows of the four farms located in Pian Cansiglio (). Pastures were excluded, because of mixed grazing by livestock and deer. Grass production and consumption by deer were estimated using exclusion cages, made of wire mesh, with a size of 1.0×1.0×1.2 (height) m. The cages (48 in 2008 and 52 in 2010) were distributed in order to cover all the meadow plots of each farm, and to sample different distances from the forest edge, as shown in . Their location was geo-referenced with a portable GPS to allow re-positioning after each cut and in different years, and implemented into ArcGiS 10 (CitationESRI 2010) to calculate their distance (m) from the forest edge, using an ortophotograph (year 2006; 1:10.000) of the area. The cages were placed in April, before the resumption of vegetative growth. Immediately before the first cut, the grass grown inside the cages was manually harvested by cutting at an approximate height of 5 cm from the ground. The same procedure was followed for a sample area of 1×1 m chosen randomly at a distance of 1–2 m from the cages. The cages where then removed to allow mowing, and re-positioned with the aid of the GPS immediately after harvest. The sampling procedure was repeated before the second cut.
Samples collected inside and outside each cage were oven dried at 65°C until constant weight, ground, and analysed for residual dry matter (DM) content (CitationAOAC, 1990) in order to estimate the DM produced and that removed by deer. In addition, the contents of crude protein (CP), ash (CitationAOAC, 1990), and neutral detergent fibre (NDF), acid detergent fibre (ADF), and acid detergent lignin (ADL) (CitationVan Soest et al., 1991) were determined in 130 samples in 2008 (34 cages x 2 positions (inside and outside) at the first cut and 31 cages × 2 positions at the second) and in 138 samples in 2010 (41 cages × 2 positions (inside and outside) at the first cut and 28 cages x 2 positions at the second).
The DM production inside the cages was used as an estimate of meadows productivity, while that outside the cages gave an estimate of the residual production left after deer grazing. The DM loss was calculated as the difference between them. DM productions and losses were expressed as kg/ha. The loss of CP was calculated as the difference between the CP content (% DM) of the samples collected inside and those collected outside the cages.
Index of deer use of meadow plots
During the summer 2008, night counts of deer with spot-lights were conducted from vehicles driving three transects that covered the meadow plots surveyed. The three transects were covered simultaneously by three teams over 1–2 h, starting at least 3 h after sunset. Counts were replicated on 29 May, 19 June, and 24 July. The aim of these counts was to obtain an index of the use of the different meadow plots by red deer, roe deer and fallow deer. To this purpose, the number of deer in each group observed for each species was recorded, and its location was plotted on a map (1:10.000). Based on this location, the deer counted were then assigned to each plot using ArcGIS 10 (CitationESRI 2010). The index of deer use was expressed for each plot as number of deer/ha. Given the very low numbers of roe deer and fallow deer counted (see results), the index was calculated only for red deer.
Statistical analysis
Forage productions (DM, kg/ha) and chemical composition (% DM) were analysed with the PROC MIXED procedure of CitationSAS (2006), using a mixed linear model with the fixed effects of farm (4 levels), cut (2 levels), year (2 levels), sampling position (2 levels: inside vs outside of the cage) and the interaction between cut, year and sampling position. Cage nested within farm was used as random effect.
The effect of distance from forest edge on DM losses was tested with a mixed linear model where year was used as fixed effect, distance (m) of the cage from the forest edge as a covariate, and cage as a random effect.
Finally, simple Pearson correlation analyses were used to test whether in the different meadow plots there was any relation between the index of deer use (average of all counts) and the DM losses (sum of losses at the first and second cut), and whether the CP content of grass inside the different cages was related to the CP losses.
Results
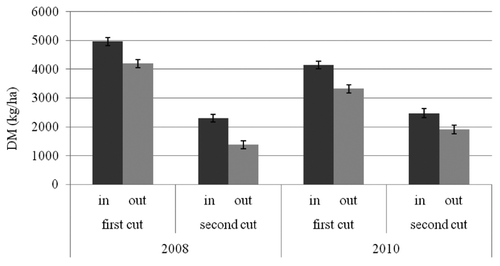
The analysis of variance of DM production showed highly significant effects for year (F=7.04; df=1; P<0.01), cut (F=532.60; df=1; P<0.001), sampling position (F=69.85; df=1; P<0.001), farm (F=20.65; df=3; P<0.001), and the interaction between cut, year and sampling position (F=10.99; df=4; P<0.001). The LS means for this interaction are given in . The DM production inside the cages at the first and second cut was 4963±139 kg/ha and 2297±136 kg/ha in 2008, and 4145±133 kg/ha and 2475±153 kg/ha in 2010. The residual DM production outside the cages was remarkably lower in both cuts, with 4199±139 kg/ha at the first and 1378±136 kg/ha at the second in 2008, and 3320±133 kg/ha and 1913±153 kg/ha in 2010.
The distance from forest edge significantly affected the DM losses in 2008 (b=−1.69; t=−2.28; SE=0.74; df=42; P<0.05), but not in 2010 (b=−0.48; t=−0.65; SE=0.48; df=42; P=0.52). For each m of increasing distance from forest, DM losses decreased of approximately 1.7 kg DM/ha in 2008, while did not change in 2010.
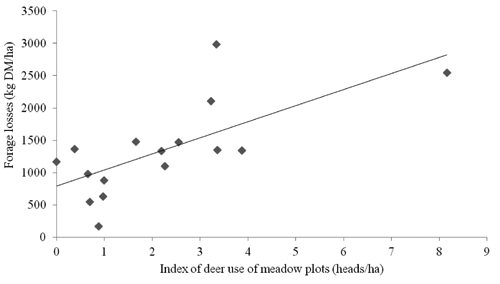
The maximum number of red deer observed during the night counts was 686 heads, while the maximum numbers of roe deer and fallow deer were 3 and 20 heads respectively. The index of meadows use by red deer varied widely between plots, without a clear temporal trend (average 2.1±2.0 heads/ha). There was a significant correlation with total DM losses (n of meadow plots=15; r=0.70; P<0.01 ).
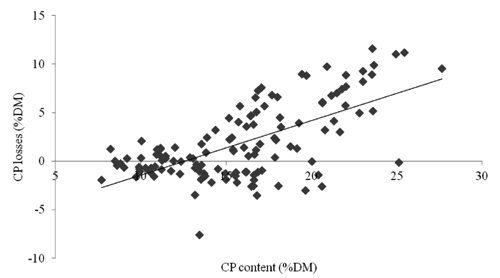
The chemical composition of forage is given in . On average, forage quality was good, with CP and NDF contents of 13–15% DM and 56–60% DM at the first cut and 18–19% DM and 51% DM at the second. The statistical analysis showed significant differences for chemical composition. Crude Protein content was affected by cut (F=153; df=1; P<0.001),), farm (F=24.76; df=1; P<0.001), sampling position ((F=24.76; df=1; P<0.001), and the three-way interaction between year, cut and sampling position (F=3.25; df=4; P<0.05). In all cuts and year, CP content of forage was 1–3% lower outside than inside the cages. However, CP losses depended on the initial quality of the forage, as indicated by a strong positive correlation between the CP content of the samples inside of the cages and the corresponding CP losses (n=128; r=0.66; P<0.001; ).
Table 1 LS means (SE) of chemical composition (% DM) per year, cut and sampling position.
Also for ADL, ADF, and Ash contents the three-way interaction was statistically significant, but post-hoc tests showed no significant differences between sampling position. This interaction was not significant for NDF (F=2.02; df=4; P=0.092).
Discussion
Grass production differed slightly between the two years, most probably as a result of climatic variability, and was in line with what expected from these groups of high productive mountain meadows (CitationZiliotto et al., 2004; CitationScotton et al., 2005). Also the difference between the first and the second cut was predictable (CitationGruber et al., 1999; CitationSpanghero et al., 2003). A significant source of difference was the farm. This effect was expected and was included in the model because sward improvements and fertilizers inputs vary between the four farms studied.
The production losses observed in this study were remarkable and fairly constant throughout the study period, varying within a narrow range of around 800–1000 kg DM/ha in both cuts and years. These amounts account for approximately 15–20% of the forage productivity at the first cut, and for 25–40% of that at the second cut. The negligible occurrence of other deer species observed during the night counts, and the positive correlation between the index of deer use and the production lost in the different plots, confirmed that these losses are a direct consequence of red deer grazing. In general, the losses observed confirm preliminary indications obtained in the same area by CitationMearns et al. (2007), while are higher than those reported by CitationTrdan and Vidrih (2008) and CitationWilson et al. (2009). There may be many reasons why losses in this study were so remarkable and did not vary between cuts and years. Red deer density in Cansiglio is high because the species, which is very sensitive to hunting disturbance (CitationPedrotti et al., 2007; CitationKamei et al., 2010), is attracted by protected areas (CitationPedrotti et al., 2007). In addition, the absence of risks to be shot while in open areas encouraged deer to use grasslands, as is also suggested by the fact that the distance from forest edge had a limited effect on forage losses. In fact, losses decreased with increasing distance in 2008 but not in 2010, most likely as a result of a progressive habituation by deer to using open areas. This might be an important implication, because other studies, which were conducted in proximity of forest edges, suggested that forage consumption by deer was higher in plots closer to cover (CitationTrdan and Vidrih, 2008; CitationWilson et al., 2009; CitationKamei et al., 2010). Another reason for the high losses is the continuous presence of deer throughout summer. In general, in mountain areas red deer use meadows in early spring, at the beginning of grass re-growth, but then, with the advancing of summer, migrate to higher elevations where forage quality is better (CitationPettorelli et al., 2005; CitationLuccarini et al., 2006; CitationBocci et al., 2010). However, in Cansiglio the density of deer is constant, or even growing, from late spring to autumn, probably because the second cut of the meadows maintains an offer of good quality forage provided by the grass re-growth (CitationMearns et al., 2007).
This study demonstrates also that red deer grazing, in addition to reducing the production of forage, might have a negative impact on its quality. The chemical composition of grass, especially for CP content, was slightly better than that observed in previous studies in the same area (CitationAndrighetto and Ramanzin, 1987; CitationXiccato et al., 1998), but was significantly affected by deer grazing. In fact, there were no differences in CP content between the samples collected inside and outside the cages when CP content inside was around 12–13% DM, but the reduction in CP content outside reached an average of around 5% DM when CP content inside was around 20–25% (). This effect most probably reflects a selective feeding of deer on plant species and/or morphological parts that are richer in CP content. To this regard, an increase in the proportion of grasses as respect to that of legumes in the samples collected outside the cages might explain why in these samples CP decreased while NDF did not change, since grasses are poorer in CP than legumes, while are richer in NDF. In this study the botanical and morphological compositions of the samples collected inside and outside the cages were not compared. However, based on the feeding behaviour of other deer species grazing pastures of different quality (CitationBryant et al., 1981; CitationWeckerly, 1994), it is plausible to hypothesize that red deer in Cansiglio were able to feed more selectively when sward was younger and richer in leaves.
The results obtained in this study have also important management implications. In the most severely damaged plots, the farmers might decide to shift to pasture after the first instead than the second cut, because its labour and mechanisation costs would not be compensated by the low residual production. In addition, the loss in protein content increases the costs of forage supplementation, and might again discourage the farmers from investing in management practices designed to improve forage quality. The losses in quantity and quality of the forage produced on farm are especially negative for the organic farmers, who might be unable to fulfil the share of the forage budget that must be produced on farm as required by organic regulations (EU Regulation N. 834/2007), and, in any case, have to face higher costs when purchasing forage and supplements from the market.
Conclusions
This study confirms that high densities of red deer seriously impact on forage production by mountain meadows. In addition, demonstrates that this heavy grazing might negatively influence the crude protein content of the residual forage, most likely as a result of selective feeding by deer. These damages increase substantially production costs for the local dairy farms, and might require adjustments in management practices of meadows and in feeding strategies. The magnitude of the effects observed in this study is likely the result of a series of combined circumstances: the study area is protected and therefore attracts high densities of the species, the intensive management to which meadows are subjected maintains an offer of good quality forage throughout summer, and the forest surrounding grasslands provides cover when needed. Therefore, the findings of this study cannot be generalised to other areas where deer density is lower and human disturbance is stronger. However, red deer is rapidly expanding in many European Countries, and this, in combination with the abandoning of agriculture and the depopulation of mountain regions, will increase the presence of the species into rural areas. Hence, the occurrence of conflicts with agriculture will also increase. Finally, the study concentrated on the damages to meadows, but it is also important to stress that this problem must be addressed with an ecosystem and holistic approach. The co-existence of deer and cattle in the same meadows might cause cross-transmission of etiological agents and have sanitary implications (CitationGortazár et al., 2007). To avoid any actual and potential conflict, meadows could be fenced to exclude deer. However, this practice could be too expensive for large areas, and could not be acceptable in a Nature 2000 site because of potential impacts on other species. In addition, it would concentrate all feeding by deer in the forest, where the impacts on biodiversity are already high. Red deer culling might reduce population density, and provide an additional source of income to the local communities, but it would also reduce the touristic appeal of the area, which is now often visited especially for deer-watching, and would be strongly contrasted by part of the public opinion. In conclusion, this paper is also an example of how, in marginal areas, farming will increasingly need to be integrated into a multi-purpose ecosystem management.
Acknowledgments:
This study was funded by Belluno province in 2008 and by Veneto Agricoltura in 2010.
The authors thank Federica Sancin for botanical analyses, and the personnel of Veneto Agricoltura for helping in field work. Preliminary results were presented at the 19th ASPA Congress, 7-10 June 2011, Cremona, Italy.
References
- AmmerC. 1996 Impacts of ungulates on structure and dynamics of natural regeneration of mixed mountain forests in the Bavarian Alps Forest. Ecol. Manag 88 43 53
- AndrighettoI. RamanzinM. 1987 Caratteristiche chimiche e digeribilità dell'erba di un pascolo posto in Pian Consiglio Agricoltura delle Venezie 9 535 542
- AOAC 1990 Official methods of analysis 15th ed. Association of Official Analytical Chemist Arlington, VA, USA
- ARPAV 2011 Available from: http://www.arpa.veneto.it/
- BocciA. MonacoA. BrambillaP. AngeliniI. LovariS. 2010 Alternative strategies of space use of female red deer in a mountainous habitat Ann. Zool. Fenn 47 57 66
- BottazzoM. NicolosoS. 2010 Piano di controllo del cervo nel comprensorio del Cansiglio Veneto Agricoltura Ed Venezia, Italy
- BryantF.C. TaylorC.A. MerrillL.B. 1981 White-tailed deer diets from pastures in excellent and poor range condition J. Range Manage 34 193 200
- CarnevaliL. PedrottiL. RigaL. TosoS. 2009 Ungulates in Italy. Status, distribution, abundance, management and hunting of Ungulate populations in Italy. Report 2001-2005 Biol. Cons. Fauna 117 1 168
- CaslickJ.W. DeckerD.J. 1979 Economic feasibility of a deer-proof fence for apple orchards Wildlife Soc. B 7 173 175
- CaudulloG. De BattistiR. ColpiC. VazzolaC. Da RonchF. 2003 Ungulate damage and silviculture in the Cansiglio forest (Veneto Prealps, NE Italy) J. Nat. Conserv 10 233 241
- CôtéS. RooneyT.P. TremblayJ.P. DussualtC. WallerD.M. 2004 Ecological impacts of deer overabundance Annu. Rev. Ecol. Evol. Syst 35 113 147
- DecalestaD.S. SchwendemanD.B. 1978 Characterization of deer damage to soybean plants Wildlife Soc. B 6 250 253
- Environmental Systems Resource Institute 2010 ArcMap 10. Redlands, CA, USA Available from: http://www.esri.com/
- FargioneM.J. CurtisP.D. RichmondM.E. 1992 Resistance of woody ornamental plants to deer damage Hort Impact 92 1 3
- GebertC. Verheyden-TixierH. 2001 Variations of diet composition of Red Deer (Cervus elaphus L.) in Europe Mammal Rev 31 189 201
- GoetschC. WiggJ. RoyoA.A. RistauT. CarsonW.P. 2011 Chronic over browsing and biodiversity collapse in a forest understory in Pennsylvania: results from a 60 year-old deer exclusion plot J. Torrey Bot. Soc 138 220 224
- GordonI.J. HesterA.J. Festa-BianchetM. 2004 The management of wild large herbivores to meet economic, conservation and environmental objectives J. Appl. Ecol 41 1021 1031
- GortazárC. FerroglioE. HöfleU. FrölichK. VicenteJ. 2007 Diseases shared between wildlife and livestock: a European perspective Eur. J. Wildlife Res 53 241 256
- GruberL. SteinwidderA. StefanonB. SteinerB. SteinwenderR. 1999 Influence of grassland management in Alpine regions and concentrate level on N excretion and milk yield of dairy cows Livest. Prod. Sci 61 155 170
- HofmannR. R. 1989 Evolutionary steps of ecophysiological adaptation and diversification of ruminants: a comparative view of their digestive system Oecologia 78 443 457
- HoltC.A. FullerR.J. DolmanP.M. 2011 Breeding and post-breeding responses of woodland birds to modification of habitat structure by deer Biol. Conserv 144 2151 2162
- KameiT. TakedaK. KohK. IzumiyamaS. WatanabeO. OhshimaK. 2010 Seasonal pasture utilization by wild sika deer (Cervus nippon) in a sown grassland Grassland Sci 56 65 70
- LuccariniS. MauriL. CiutiS. LambertiP. ApollonioM. 2006 Red deer (Cervus elaphus) spatial use in the Italian Alps: home range patterns, seasonal migrations, and effects of snow and winter feeding Ethol. Ecol. Evol 18 127 145
- MarshallE.J.P. ThomasC.F.G. JoenjeW. KleijnD. BurelW. LeCoeurD. 1994 Establishing vegetation strips in contrasted European farm situations BoatmanN.D. Field margins: integrating agriculture and conservation The British Crop Protection Council Publ Farnham, UK 335 340
- MartinT.G. ArceseP. ScheerderN. 2011 Browsing down our natural heritage: deer impacts on vegetation structure and songbird populations across an island archipelago Biol. Conserv 144 459 469
- MattioliS. MeneguzP. G. BrugnoliA. NicolosoS. 2001 Red deer in Italy: recent changes in range and numbers Hystrix It. J. Mamm 12 27 35
- MearnsG.C. Da RonchF. BernasconiL. De BattistiR. DestroM. LupanoS. MazzoccoM. OngaratoM. PiccinA. 2007 Pabular preferences and feeding sites of the red deer (Cervus elaphus Linneaus, 1758) in Cansiglio (NE Italy) 44 in Proc. 1st Int. Conf. on Genus Cervus Primiero, TN, Italy
- MelisC. BusetA. AarrestadP.A. HansenO. MeisingsetE.L. AndersenR MosknesA. RøskaftE. 2005 Impact of red deer Cervus elaphus grazing on bilberry Vaccinium myrtillus and composition of ground beetle (Coleoptera, carabidae) assemblage Biodivers. Conserv 6 2049 2059
- MilnerJ.M. BonenfantC. MysterudA. GaillardJ.M. CsànyiS. StensethN.C. 2006 Temporal and spatial development of red deer harvesting in Europe: biological and cultural factors J. Appl. Ecol 43 721 734
- MottaR. 1996 Impact of wild ungulates on forest regeneration and tree composition of mountain forests in the Western Italian Alps Forest Ecol. Manag 88 93 98
- MottaR. 2003 Ungulate impact on rowan (Sorbus aucuparia L.) and Norway spruce (Picea abies (L.) Karst) height structure in mountain forests in the eastern Italian Alps Forest Ecol. Manag 181 139 150
- MuñozA. BonlR. DìazM. 2008 Ungulates, rodents, shrubs: interactions in a diverse Mediterranean ecosystem Basic Appl. Ecol 10 151 160
- OberdorferE. 1964 Der insubrische Vegetationskomplex, seine Struktur und Abgrenzung gegen die submediterrane Vegetation in Oberitalien und in der Südschweiz Beitr Naturk Forsch Sfidw Deut 23 141 187
- PedrottiL. AngeliF. LuchesaL. 2007 Red deer management in an Alpine protected area: the dilemma of natural regulation or culling 46 in Proc. 1st Int. Conf. on Genus Cervus Primiero, TN, Italy
- PettorelliN. MysterudA. YoccozN.G. LangvatnR. StensethN. C. 2005 Importance of climatological downscaling and plant phenology for red deer in heterogeneous landscapes Proc. R. Soc. B 272 2357 2364
- PoldiniL. OrioloG. 1994 La vegetazione dei prati da sfalcio e dei pascoli intensivi (Arrhenatheretalia e Poo-Trisetalia) in Friuli, NE Italy Studia Geobotanica 14 3 48
- PorterW.F. 1983 A baited electric fence for controlling deer damage to orchard seedlings Wildlife Soc. B 11 325 327
- PutmanR.J. MooreN.P. 1998 Impact of deer in lowland Britain on agriculture, forestry and conservation habitats Mammal Rev 28 141 164
- RamboJ.L. FaethS.H. 1999 Effect of vertebrate grazing on plant and insect community structure Conserv. Biol 13 1047 1054
- RooneyT.P. WallerD. 2003 Direct and indirect effects of white-tailed deer in forest ecosystems. Forest Ecol Manag 181 165 176
- RussellF.L. ZippinD.B. FowlerN.L. 2001 Effects of white-tailed deer (Odocoileus virginianus) on plants, plant populations and communities: a review Am. Midl. Nat 146 1 26
- SAS 2006 User's Guide SAS Inst. Inc Cary, NC, USA
- SchützM. RischA. C. LeuzingerE. KrüsiB. O. AchermannG. 2003 Impact of herbivory by red deer (Cervus elaphus L.) on pattern and processes in subalpine grasslands in the Swiss National Park Forest Ecol. Manag 181 177 188
- ScottonM. MariniL. PecileA. RodaroP. 2005 Tipologia dei prati permanenti del Trentino orientale Istituto Agrario di San Michele all'Adige Ed. Trento, Italy
- SmitR. BokdamJ. Den OudenJ. OlffH. Schot-OpschoorH. SchrijversM. 2001 Effects of introduction and exclusion of large herbivores on small rodent communities Plant Ecol 155 119 127
- SpangheroM. BoccalonS. GraccoL. GruberL. 2003 NDF degradability of hays measured in situ and in vitro Anim. Feed Sci. Tech 104 201 208
- StizG.P. 1997 Il cervo (Cervus elaphus) del Bosco del Cansiglio: risultati del censimento al bramito e note sulla distribuzione della popolazione nel territorio Rapporto inedito all'Osservatorio Faunistico Pordenone, Italy Available from: http://www.cansiglio.it/
- TrdanS. VidrihM. 2008 Quantifying the damage of red deer (Cervus elaphus) grazing on grassland production in southeastern Slovenia Eur. J. Wildlife Res 54 138 141
- TremblayJ.P. HuotJ. PotvinF. 2007 Density-related effects of deer browsing on the regeneration dynamics of boreal forests J. Appl. Ecol 44 552 562
- Van SoestP.J. RobertsonJ.B. LewisB.A. 1991 Methods for dietary fiber, neutral detergent fiber, and non starch polysaccarides in relation to animal nutrition J. Dairy Sci 74 3583 3597
- WekerlyF.W. 1994 Selective feeding by black-tailed deer: forage quality or abundance? J. Mammal 75 905 913
- WildiO. OrlociL. 1996 Numerical exploration of community patterns. A guide to the use of MULVA-5 2nd ed. SPB Academic Publ. Amsterdam, The Netherlands
- WilsonC.J. BrittonA.M. SymesR.G. 2009 An assessment of agricultural damage caused by red deer (Cervus elephus L.) and fallow deer (Dama dama L.) in Southwest England Wildl. Biol. Pract 5 104 114
- XiccatoG. Parigi BiniR. CarazzoloA. TrocinoA. CossuM.E. 1998 Wilting effect on fermentation characteristics and nutritive value of mountain permanent meadow grass silage Ann. Zootech 47 279 291
- ZiliottoU. AndrichO. LasenC. RamanzinM. 2004 Tratti essenziali della tipologia veneta dei pascoli di monte e dintorni Accademia Italiana di Scienze Forestali Ed. Venezia, Italy