Abstract
Aim of the study was to achieve scientific information about body measures and milk production of Cabannina cattle, a local breed reared in northern Italy. Fourteen body measures and five morphologic indexes were recorded from 86 heads enrolled in the herd book. Low differences between males and females of the same age-class were shown. Body measures were generally greater than those reported in previous studies, probably due to recent crosses. With reference to milk production, 991 test-day records from 128 lactations of 59 cows were analysed. Average milk daily production was 8 kg/d in 1st lactation to 10.61 in 3rd (P<0.05); the parameters of the Wood equation draw atypical curves with the exception of curves from spring calving cows. Only 74.5% of lactations with an adjusted R2>0.75 showed a standard curve, with low persistence (7.7%), high value of d at peak (103 d) and peak production of 20.18 kg of milk. Moreover, 100 milk samples (40 to 220 d of lactation) were submitted to a granulometric survey by laser scatter technique in order to evaluate the dimensions of fat globules; then milk fat was analyzed by gas chromatography, and desaturase indexes were determined. Cabannina cows showed small fat globules with high specific surface. Furthermore mean diameter of milk fat globules decreased during lactation then rose. Milk fat contained high levels of cis-MUFA, and high desaturase indexes. In conclusion, the low size of Cabannina cattle orients for a limited meat production. Instead milk production has a higher economic potential, aimed at cheese production and human nutrition.
Introduction
In a global scenario, in which water, energy and economic resources will decrease more and more, agricultural system’s goal must be vegetal and animal biodiversity preservation and an efficient use of lands (FAO, Citation2009). In this context the European food system must produce sufficient, healthy and nutritious food, while drastic reducing CO2 emissions and regenerating landscapes and natural resources, in a new concept of innovative and environmentally sustainable agricultural panorama. The Common Agricultural Policy must enhance biodiversity and ecologically functional productions, by connecting traditional knowledge and modern techniques. Researches and projects that explore the potential of diverse and locally adapted production systems, while respecting the animal welfare, will be pivotal in forthcoming years.
Due to its north-to-south extension Italy is endowed by a wide variety of climate, and a lot of cattle breeds, approximately 30, among which only 16 are included in the Herd Book of Italian autochthonous cattle breeds (Associazione Italiana Allevatori, Citation2012b), had the possibility, over the centuries, to adapt themselves to different environmental situations. Local breeds are important for the maintenance of biodiversity in several territories, mainly in areas where an intensive animal production system is not applicable. However, it is necessary that their productions are known, enhanced and included in a market context, in order to find an economic rationale to their breeding.
The Cabannina cattle breed is an autochthonous breed that originated in the province of Genoa, in northern Italy and it is considered as a dual purpose cattle breed (Associazione Italiana Allevatori, Citation2012b). There are currently about 250 heads, enrolled in the Herd Book of Italian endangered cattle breeds, of which only 73 underwent during 2011 to the milk recording activity (Associazione Italiana Allevatori, Citation2012a). Cabannina is a rustic cattle breed, with a low size and with a medium-low milk production. Milk is used for the production of a local cheese, named U’Cabanin (Guida, Citation2011).
The aim of the study was to achieve new scientific information about this breed, in order to enhance the meat and milk production and milk quality in the Cabannina breed and to better introduce these items in the new breeding scenario.
Materials and methods
Body measures
The study was carried out in 10 farms in the province of Genoa, with tie stall rearing in the cold season and pasture in the warm season, during the period from January to July 2011. Rations for growing and adult animals were based on dry forages or pasture and concentrates given to meet the requirements of the animals in relation to the stage of growth. Body measures were taken from a sample of 86 individuals (75 females born between 1997 and 2011 and 11 males born between 2003 and 2011), enrolled in the Cabannina herd book. The sampled animals were the same for all the considered parameters. In relation to the number of animals six age-classes were defined:
| i) | from 1 d to 30 d (n=5); | ||||
| ii) | from 31 d to 60 d (n=4); | ||||
| iii) | from 61 d to 120 d (n=2); | ||||
| iv) | from 121 d to 365 d (n=6); | ||||
| v) | from 366 d to 730 d (n=11); | ||||
| vi) | >730 d (n=58). | ||||
Fourteen body measures (Meregalli, Citation1980) were taken by means of the meter stick and of the metric tape:
| - | height at withers (HW): from the withers to the ground; | ||||
| - | height at croup (HC): from the top of the croup to the ground; | ||||
| - | chest circumference (CC): behind the posterior edge of the shoulders at the point of least perimeter; | ||||
| - | length of the trunk (LTr): from the anterior edge of shoulder to the posterior edge of buttock; | ||||
| - | depth of chest (DC): from the withers to the sternum; | ||||
| - | chest width (CW): behind the shoulder; | ||||
| - | length of the thorax (LTh): from the tip of the shoulder to the rear edge of the last rib; | ||||
| - | rump width (RW1): distance between the two iliac tuberosities; | ||||
| - | rump width (RW2): between the two ischial tuberosities; | ||||
| - | rump width (RW3): between the hips; | ||||
| - | rump length (RL): from the tip of the hip to that of the buttock; | ||||
| - | cannon bone circumference (CBC): at the upper third of the metacarpal bone; | ||||
| - | length of head (LH): from the head top to the tip of the muzzle; | ||||
| - | width of head (WH): between the temples. | ||||
| - | Five morphologic indexes were then calculated from body measures (Magliano, Citation1950; Fernandez et al., Citation2007), namely: | ||||
| - | anamorphosis index (AI): CC2/HW; | ||||
| - | pelvic index (PI): RW1/RL; | ||||
| - | lateral corporal index (LCI): HW/LTr; | ||||
| - | corporal index (CI): LTr/CC; | ||||
| - | dactylo-thoracic index (DTI): CBC/CC. | ||||
Milk production
The study was carried out using the test day (TD) records for milk production (kg), fat and protein content (%) and somatic cell count (SCC, no.*1000) of Cabannina cows reared in 7 herds of the province of Genoa from July 2008 to December 2011. The cattle management system was the same in all herds, based on tie stalls rearing and traditional feeding (dry forages and concentrates given separately twice a day, with a forage/concentrate ratio of 60/40) during the cold season and daily pasture plus evening concentrates in stalls during the warm season. In all herds involved in the study an automatic milking system was adopted, also during the pasture period.
Lactations with less than 5 controls were removed from the dataset, so the analysis was conducted on 991 monthly TD records (conducted in 145 different days) from 128 lactations of 59 cows. Out of the 128 lactations, 27 were first, 20 were second and 81 third or higher lactation; 34 started in spring, 28 in summer, 22 in autumn and 44 in winter (respectively, from March to May, from June to August, from September to November and from December to February). Fourty-four 7-day classes plus one class for lactations over 308 d were considered. Data for fat and protein production (g/d) were calculated from milk production and percentage composition. Because SCC data were not normally distributed, SCC was transformed into somatic cell score (SCS), according to Shook and Schutz (Citation1994) by means of the following equation: SCS=3+log2(SCC/100,000).
Milk fat globule granulometry
A total number of 100 milk samples were taken from 10 lactating Cabannina cows (5 in second and 5 in third lactation) from one herd. Milk samples were collected from the whole udder by a lactometer during the morning milking from animals in early and mid lactation, ranging from 40 to 220 days in milk (DIM), with a 20 d sampling interval. Milk samples were forwarded to the laboratory at 4°C and analyzed within 2 h.
Distributions of fat globules diameter were performed on 10 mL samples using a granulometer laser scatter, according to the indirect method proposed by Lopez (Citation2005). Size distributions were characterized by volume weighted diameter of the globules (d43, in μm) and by specific surface area (SSA, in cm2 mL–1).
Fatty acid extraction, derivatization and desaturase indexes
The remaining part of the samples was stored at -20°C until analysis. Lipid extraction was performed according to a modified Bligh and Dyer method (Manirakiza et al., Citation2001) with chloroform/methanol mixtures, chloroform and micro-filtered distilled water. The samples (7.25 mL) were thus placed in centrifuge at 2000 rpm for 10 min at room temperature (20°C) in order to separate the solution into three phases. The deepest phase (solution of chloroform containing the lipids) was drawn and placed in a glass tube in order to be dried under a slight nitrogen flow. The solution previously dried was derivatized according to Molto-Puigmartì et al. (Citation2007), using solutions of NaOCH3/MeOH 0.5M and BF3. After complete derivatization, 0.5 L of solution containing lipids was finally injected and analyzed by gas chromatograph Trace GC 2000 (Thermo-Fisher Scientific, Waltham, MA, USA) equipped with a TR-CN100 column (100 m, 0.25 mm i.d. x 0.20 μm film thickness) and a 5 m precolumn (i.d. 0.25 mm) deactivated (TeknoKroma, Barcelona, Spain). The oven temperature program was: 70°C held for 3 min, then from 70°C to 240°C at 2.5°C min–1, held for 10 min. Carrier gas was nitrogen at 0.5 mL min–1. Fatty acids were identified using external standards (Standard containing 37 fatty acids, FAME Mix 37, Supelco, Bellefonte, PA, USA) and two CLA standards (Matreya, Pleasant Gap, PA, USA) and quantified using C19:0 (nonadecanoic acid) as internal standard. Peak areas were corrected according to the theoretical relative FID response correction factors (TRFs) published by Ackman (Citation2002). The results are presented as g/100 g fatty acids (% by weight).
The percentage of the contribute of each fatty acid was calculated on the total of the area under the known peaks. Desaturase indexes (Δ) were calculated according to Schennink et al. (Citation2008) and the total desaturase index (TDI) was calculated according to Mele et al. (Citation2007) on C14, C16 and C18 fatty acids; briefly, the individual Δ was calculated as Cx:1/(Cx+Cx:1)*100, where x is the number of carbons of fatty acid. The total desaturase index was calculated as [(C14:1 + C16:1 + C18:1)/(C14 + C14:1 + C16 + C16:1 + C18 + C18:1) * 100)].
Statistical analysis
The body measurements and morphologic indexes data were processed by analysis of variance, by using the following general linear model (SAS, Citation2008):
where
yijkl = dependent variable (body measure or morphologic index);
μ = overall mean;
Hi = fixed effect of herd (10 levels);
Sj = fixed effect of sex (2 levels);
Ak = fixed effect of age-class (6 levels);
SAjk = interaction between sex and age-class (12 levels);
εijkl = residual error.
Limitedly to HW and CC, the growth curves were then calculated, through the application of the non-linear regression analysis to fit the Gompertz equation in the following form:
where
P is the body measure (cm) at the age t (d); b0 is the measure at maturity, b1 is the rate of growth and b2 the age at maximum growth. The described curve is a continue sigmoid curve (France et al., Citation1996).
Data from milk TD controls were submitted to analysis of variance, using the following mixed model (SAS, Citation2008):
where
yijklmno = dependent variable;
μ = overall mean;
TDi = fixed effect of the day of test (145 levels);
Hj = fixed effect of herd (7 levels);
Lk = fixed effect of lactation number (3 levels, 1st, 2nd and ≥3rd);
Sl = fixed effect of season of calving (4 levels, winter, spring, summer, autumn);
DIMm = fixed effect of class of days in milk (44 seven days classes plus one class for lactations over 308 d);
Cn(Hj) = random effect of individual cow within herd (59 levels);
εijklmno = residual error.
The lactation curve parameters for milk production were calculated by means of the non linear regression analysis to fit the Wood (Citation1967) model:
where yn is the production (kg/d) at day n, and A, B and C are the equation parameters (initial yield or scale factor, and slope during the ascendant and the descendent phase of the curve, respectively). The linear form of the model (Macciotta et al., Citation2005) was used:
The equation parameters were estimated for the whole sample or by lactation number and calving season. The model was also fitted to TD records of each individual lactation; the adjusted R-square (ADJRSQ) was calculated for each individual curve and when ADJRSQs were >0.75, then the B and C parameters sign were highlighted, so to define three different shapes of the lactation curve: standard, continuously decreasing and reversed (Macciotta et al., Citation2005).
For standard curves with ADJRSQ >0.75, the mean equation parameters were combined to calculate the persistency (-(B+1)*lnC), the peak day (B/C) and the peak production (A*(B/C)B*e–B) (Rekaya et al., Citation2000). Data concerning the size of fat globules and the analysis of fatty acids were submitted to analysis of variance by means of the following mixed model (SAS, Citation2008):
where
yijkl = dependent variable;
μ= overall mean;
Li = fixed effect of lactation number (2 levels, 2nd and 3rd);
DIMj = fixed effect of stage of lactation (ten 20 d classes from 40 to 220 d of lactation), tested until the 4th power;
Ck = random effect of individual cow (10 levels);
εijkl = residual error.
Results and discussion
Body measures and morphologic indexes
The study involved about 34% of the total population of Cabannina cattle, so it can be considered as a good indicator of the actual status of the breed. The analysis of variance applied to body measures and morphologic indexes of Cabannina cattle () revealed, along with a predictable and always highly significant (P<0.001) effect of age-class (with the exception of CI; P>0.05), also a significant herd effect in 9 out of 14 body measures and in 3 out of 5 morphologic indexes. This finding is probably related to selective choices by individual farmers, who, while trying to adhere to the breed standard, sometimes select to improve some specific traits, related to the production of meat (in particular high HW, LTr, DC) (Stålhammar et al., Citation1995) or milk (in particular high RL, low HC) (Strapak et al., Citation2011). Sex resulted as a source of variability for LTr, CC, RW2 (P<0.05) and for the sizes of the head (P<0.01). The interaction between sex and age-class was found to be significant for some dimensions of the rump (P<0.05 and P<0.01), as far as for LH (P<0.001), CBC (P<0.001), PI (P<0.05) and DTI (P<0.001). High proportion of variability (R2>0.75) was explained by the model for all parameters, but not for CI (R2=0.41).
shows the least squares means of the interaction between sex and age-class for body measures and morphologic indexes. Low differences between males and females of the same age-class have been found, though the results could be affected by the low number of animals in some classes. When differences were significant, generally females showed lower values than males of the same age-class. An exception was in a measure of the width of the croup (RW3), where class 3 females showed a significant higher value than males (which then reversed over time). Males with age >730 d were higher (about 6 cm) and longer (about 13 cm) than females of the same age class; they had a greater CC (about 18 cm), a wider rump, a more robust skeleton (greater CBC, approximately 2.5 cm) and a longer and wider head (P<0.05). Moreover, starting from 1 year of age, males showed a significantly (P<0.05) higher value of the anamorphosis index than females, that indicates a greater dimensional development, as reported also by Fernandez et al. (Citation2007). The differences between males and females were highest at two years of age (class 5) and then reduced (class 6).
In males between 1 and 2 years of age and in individuals of both sexes older than of 2 years, nine out of twelve body measures were greater than those reported in the breed standard (Associazione Italiana Allevatori, Citation2012b) and in previous studies (Usai, Citation1974; Orefice, Citation1978). This tendency to an increased size is probably related to recent crosses with higher sized breeds, such as Italian Brown, aimed at a higher milk (Sieber et al., Citation1988) and meat production, rather than to selection. Such behaviour by farmers represents a danger to the breed, because it affects a fundamental morphologic aspect. In fact, the small size is a positive feature for Cabannina cattle exploiting the pasture in a hill-mountain environment, because, lowering the centre of gravity, it allows them to be more stable in the sloping pastures. In addition, small-sized animals have lower maintenance requirements; in fact, when cattle is bred under a pasture regime, systems utilizing small cows are more profitable (Long et al., Citation1975). It could be therefore positive to return quickly to the typical morphological features (low size) by means of selection, abandoning cross-breeding.
The analysis of growth curves for CC and HW showed a great sexual dimorphism in adults ( and ), but the time when the dimorphism became apparent was different, depending on the measure concerned. It is evident that for CC, young males differed from females, while in the case of the HW, the two curves were parallel up to about 500 d of age and then they divided sharply. As previously stated, this also affected the value of the anamorphosis index, that well describes the morphological type of animals and their aptitude: typically, high values of the anamorphosis index indicate the ability for meat production (brachymorphic type) (Araújo et al., Citation2006) and reduced values the ability for milk production (dolichomorphic type) (Beretti et al., Citation2010).
Milk production
The study involved over 80% of cows of the breed that underwent the milk productivity controls, the remaining part of the cow’s population being distributed in several small sized herds. The analysis of variance conducted on TD data () revealed a highly significant effect (P<0.001) of day of test and of the individual on all the production parameters considered; the SCC showed a less important effect of the herd (P<0.05) and a non-significant effect of the class of DIM (P>0.05). Compared to the SCS, the SCC presented lower F values and, consequently, also a smaller R2: this is probably due to the fact that SCC was not normally distributed; it is therefore more correct to use the SCS value for the analysis. The lactation number showed a significant effect only on milk and protein daily production (P<0.001), while the calving season was significant for the percentage content of protein (P<0.01). This result was common to other cattle breeds (Reggiana, Valdostana, Grigio Alpina, Italian Friesian and Bianca Val Padana) under Italian environmental conditions (Sabbioni et al., Citation2003, Citation2008, Citation2011, Citation2012, Citation2013) and for Holsteins in Arizona (Ray et al., Citation1992). The R2 was always higher than 0.55, less than for the SCC (0.44). This finding is supported by previous observations on other Italian breeds (Valdostana and Grigio Alpina) (Sabbioni et al., Citation2008; Citation2011), and enforces the above mentioned preference to the use of SCS rather than SCC for the analysis of the qualitative characteristics of the milk.
The average Cabannina breed milk daily production () was particularly low (8 kg/d in 1st lactation to 10.61 in 3rd; P<0.05); because the lactation length was, on average, 290 d, the production of milk fluctuated between 2320 and 3077 kg. The fat (3.83% in 1st lactation to 3.55 in 3rd; P>0.05) and protein content (3.34% in 1st lactation to 3.26% in 3rd; P>0.05) were good and in line with those reported by Matassino et al. (Citation2011) for the same breed (3.79% fat; 3.37% protein). The same authors also showed that the fat content of Cabannina breed milk is significantly higher than that of other Italian autochthonous breeds (Burlina, Bianca Val Padana, Castana), reared under the same environmental conditions, and not significantly different from that of Agerolese. The official controls on the productivity of milk in Italy (Associazione Italiana Allevatori, Citation2012a) showed an average production of milk of 2924 kg for Cabannina breed, with 3.60% fat and 3.20% protein.
The parameters of the Wood equation for milk production for the whole sample, or relating to different lactation orders and seasons of calving are in . The table shows that generally the parameters draw atypical curves, characterized by negative values of the parameter B, and then by the absence of the peak of lactation (continuously decreasing curves) (Macciotta et al., Citation2005). Only in correspondence of spring calving cows lactations, mean parameters fit standard curves. This finding is common to other autochthonous Italian cattle breeds, as Bianca Val Padana, in which the selection for milk production has never taken place (Sabbioni et al., Citation2012).
Out of 128 lactations, 81 (63.3%) showed an ADJRSQ <0.75 and 47 (36.7%) an ADJRSQ >0.75. By limiting the survey to the latter, the data presented in () were obtained. Standard curves were 74.5%, continuously decreasing 21.3% and reversed standard 4.3%. No continuously increasing curves were detected. Standard curves with ADJRSQ >0.75 have shown a high decreasing rate (persistence = 7.7%), a very high value of days at peak (103 d) and a peak production of 20.18 kg of milk.
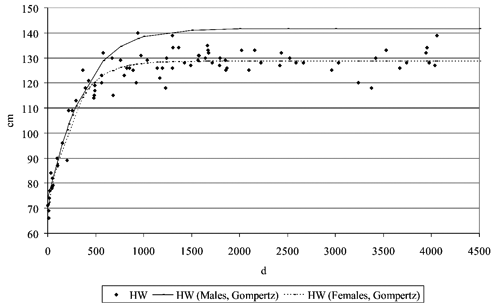
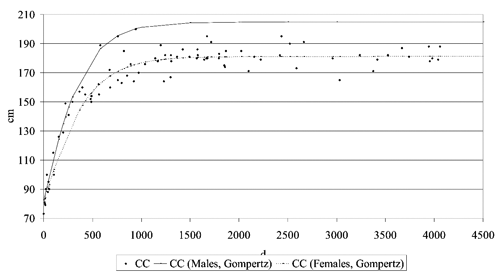
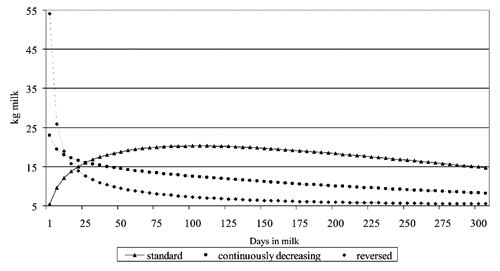
Table 1. Analysis of variance (F values) of body measures and morphologic indexes.
Table 2. Least squares means of body measures in cm and morphologic indexes in Cabannina cattle.
Table 3. Analysis of variance (F values) of milk production and quality in Cabannina cattle.
Table 4. Least squares means (±SE) of milk production and quality as affected by lactation number and calving season.
Table 5. Parameters (±SE) of the Wood’s equation for milk production in kg/d, as affected by lactation number and calving season.
Table 6. Frequency of lactation curves with different shapes and mean values (±SD) of parameters calculated only from individual curves with adjusted R-square >0.75 in Cabannina cattle.
Table 7. Ls means for d43 (volume weighted diameter), specific surface area, fatty acids and desaturase indexes of milk fat in Cabannina cows.
Fat globules size and fatty acids content
Means of Cabannina fat globules size and specific surface area are summarized in . The data confirmed the results shown in a recent survey by Communod et al. (Citation2012), in which Cabannina breed showed smaller fat globules than Friesian cows, and, consequently, the highest specific surface area. The specific surface area were twofold in Cabannina than in Friesian fat globules; this means that small fat globules contains twice as much fat globule membrane material (trilayered biological membrane) per unit triglyceride content, as reported by Michalsky et al. (Citation2005).
It has been reported (Communod et al., Citation2012; Martini et al., Citation2013) that in cow and in ewe’s milk the globule diameter decreases during lactation. In our study, this reduction is more evident in the first part of lactation, slowing down until it reaches a plateau at the 5th month. This remarkable initial decrease in fat globule diameter during the first lactation period could be explained by the maturation of the mammary gland that progressively provides more membrane to cover milk fat core, as suggested in a recent study by Michalsky et al. (Citation2005). In the second part of lactation, the results of the actual research are in contrast with those obtained by Communod et al. (Citation2012), whereas they confirm the results by Michalsky et al. (Citation2005), who described a subsequent increase in mature fat globule size during later months of lactation, probably due to the increased fat content and limited available membrane coverage.
The size of milk fat globules is directly related to the physiology of the mammal female producing milk (Lopez, Citation2011). The secretion of small milk fat globules is a biological process that requires the production of high amounts of membrane material to cover the whole surface of the lipid droplets and thus a high synthesis activity of the lactating cells (Lopez, Citation2011). Cabannina cows that secrete small fat globules could have a potential higher mammary metabolic activity than those secreting large fat globules (Communod et al., Citation2012). Having more abundant milk fat globule membrane (MFGM) could have important effects on cheese making processes (Martini et al., Citation2008), such as better incorporation of MFGM in cheese, higher resistance to churning, and more extensive water retention by the membrane, resulting in less firmness but also better storage ability (Michalski et al., Citation2002).
The high mammary activity is suggested also by high desaturase indexes, if compared to literature (Schennink et al., Citation2008; Heck et al., Citation2009; Garnsworthy et al., Citation2010), especially Δ14, Δ16 and Δ18 (); these indexes could also explain the ability of Cabannina cows to produce more unsaturated fatty acids than Friesian cows (Schennink et al., Citation2008, Communod et al., Citation2012).
In the last years the importance of milk fat in human and animal nutrition has come rapidly to the fore. Several nutritionist Associations recommend the intake of higher levels of cis-MUFA in milk despite of SFA levels (Kris-Etherton, Citation1999). The content in cis-MUFA found in the present research is quite similar to data reported in the recent literature (Mansson, Citation2008; Soyeurt et al., Citation2008; Heck et al., Citation2009; Gaspardo et al., Citation2010), with some exceptions: in particular, the C14:1cis9 and C16:1cis9 levels were higher, whereas C18:1cis9 showed slight lesser levels than those reported by Mansson (Citation2008), Soyeurt et al. (Citation2008) and Gaspardo et al. (Citation2010). Also the desaturase indexes overlap with those reported by other authors, with slightly reduced Δ14 levels (Schennink et al., Citation2008; Heck et al., Citation2009; Garnsworthy et al., Citation2010).
The enhancement of the cis-MUFA in milk is desirable for the human consumption, as reported by Givens (Citation2008). Higher intakes of cis-MUFA and a reduction in short fatty acids reduce the plasma insulin levels, the total plasma cholesterol and the LDL-cholesterol concentrations, therefore reducing the risks of coronary heart disease. The positive effects of cis-MUFA can exert also on the udder health. During mastitis, for example, an enhancement in lipase activity can be appreciated, with increase in free fatty acids, mainly short chain fatty acids (Randolph and Erwin, Citation1974); several fatty acids are endowed of a good antibiotic power, that can be expressed via inhibition of enzyme/fatty acid synthesis/nutrient uptake, cell lysis, metabolites leakage, disruption of electron transport chain, interference with oxidative phosphorylation and lipid peroxidation (Desbois and Smith, Citation2010). By enhancing the activity of stearoyl CoA desaturase, the nutritional value of milk would be positively ameliorated, but a simple up regulation of its activity seems to be limited, as reported in a comprehensive milk lipid synthesis model (Shorten et al., Citation2004).
In our study, Cabannina cows showed both higher levels of cis-MUFAs and of desaturase indexes if compared with recent literature (Schennink et al., Citation2008; Garnsworthy et al., Citation2010; Communod, Citation2012). These features could be linked to genetics, by complex interaction in gene/allele expressions, as evidenced by Schennink et al. (Citation2008). Another important characteristic taken into account was the oleic acid content, in relation to stearic acid content. C18:1cis/C18:0 trend increases with the lactation stage. Recent studies (Schwartz et al., Citation2008; Dipasquale et al., Citation2010) showed that oleic acid introduced by diet is the most important substrate to produce a lipid amide by enterocytes, involved in the induction of satiety. This compound is oleoylethanolamide (OEA) and, as reported by Dipasquale et al. (Citation2010), has several characteristics of satiety factor: it inhibits feeding by prolonging the interval to next meal. OEA synthesis is regulated by nutrient availability and its levels undergo circadian fluctuations.
Conclusions
The study confirmed the low milk production of Cabannina cattle and highlighted that the lactation curves are frequently far from a standard shape, and that, also when the shape is in the standard type, the peak is very late and the persistence is low.
Smaller dimensional aspect of Cabannina milk globules could become an important reason to recover this autochthonous breed, because this peculiar feature could assign to milk fat globules a potential higher digestibility, influencing pancreatic lipase activity. Small globule dimension highlights the crucial role of the MFGM in the efficient digestion of milk fat globules and brings new perspectives for the design of dairy products and infant formulas. Moreover, the results suggest that it is interesting to assume that from a breed biodiversity could derive a milk biodiversity, showing in the forthcoming years peculiar nutritional and nutraceutical aspects in this different milk and dairy products derived.
Meat production has a lower importance because the breed is small sized: nevertheless, males can be an important source of income. Males reach the highest differences in size with females at approximately two years; moreover, crosses with larger breeds should be avoided in future, to maintain the breed efficiency in pasture systems.
Further studies should be conducted at farm level to optimize feeding and pasture quality, in order to improve the potentiality of the breed.
Acknowledgments
The authors kindly acknowledge Mr. Marco Rizzo from the Provincial Breeders Association (APA), Genova, Italy, and all farmers for their collaboration.
References
- AckmanR.G. 2002. The gas chromatograph in practical analyses of common and uncommon fatty acids for the 21st century. Anal. Chim. Acta 465:175-192.
- AraújoJ P.MachadoH.CantalapiedraJ.IglesiasA.Petim-BatistaF.ColaçoJ.SánchezL. 2006. Biometric and liveweight analysis of Minhota beef cows. pp 15-32 in Proc. 8th World Congr. Genetics Appl. Livest. Prod., Belo Horizonte, Brazil.
- Associazione Italiana Allevatori, 2012a. Bollettino dei Controlli della Produttività del Latte. Available from: http://bollettino. aia.it/bollettino/bollettino.htm
- Associazione Italiana Allevatori, 2012b. Norme tecniche del Registro Anagrafico delle razze bovine autoctone a limitata diffusione. Available from: http://www.aia.it/aia-website/it/attivita/area-tecnica/ufficiosviluppo/razze-bovine-autoctone
- BerettiV.SimonazziD.PainiV.SuperchiP.SabbioniA. 2010. Body measures in Reggiana cattle and their correlations with morphological evaluations. Ann. Fac. Med. Vet. Parma 30:89-100.
- CommunodR. 2012. Bovine lactation and reproduction physiology: a survey on some fundamental features in autochthonous dairy breeds reared in northern Italy. Degree Diss., University of Milano, Italy.
- CommunodR.FaustiniM.ChiesaL.M.TorreM.L.LazzatiM.VigoD. 2012. Milk biodiversity: future perspectives of milk and dairy products from autochthonous dairy cows reared in northern Italy. In: AladjadjiyanA. ( ed.) Food production-Approaches, challenges and tasks. InTech On-line Publ., pp 169-184.
- DesboisA.P.SmithV.J. 2010. Antibacterial free fatty acids: activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotech. 85:1629-1642.
- DipasqualeP.RomanoA.CianciS.RighettiL.GaetaniS. 2010. Oleoylethanolamide: a new player in energy metabolism control. Role in food intake. Drug Discov. Today 7:e169-e174.
- FAO, 2009. How to feed the world in 2050. Available from: http://www.fao.org/fileadmin/templates/wsfs/docs/expert_paper/Ho w_to_Feed_the_World_in_2050.pdf
- FernandezE.N.MartinezR.D.GeneroE.R.BroccoliA.M. 2007. Indices zoometricos en bovinos criollos de origen patagonico y del noroeste argentino. Veterinaria (Montevideo) 42:23-27.
- FranceJDijkstraJThornleyJ.H.DhanoaM.S. 1996. A simple but flexible growth function. Growth Develop. Aging 60:71-83.
- GarnsworthyP.C.FengS.LockA.L.RoyalM.D. 2010. Heritability of milk fatty acid composition and stearoyl-CoA desaturase indices in dairy cows. J. Dairy Sci. 93:1743-1748.
- GaspardoB.LavrencicA.LevartA.Del ZottoS.StefanonB. 2010. Use of milk fatty acids composition to discriminate area of origin of bulk milk. J. Dairy Sci. 93:3417-3426.
- GivensD.I. 2008. Challenges facing the food industry in innovating for health: impact on CVD risk of modifying milk fat to decrease intake of SFA and increase intake of cis-MUFA. pp 419-427 in Proc. Nat. Symp. Nutrition Society of the University of Dublin, Dublin, Ireland.
- GuidaS. 2011. Caratterizzazione morfologica e studio della curva di lattazione nella razza bovina Cabannina. Degree Diss., University of Parma, Italy.
- HeckJ.M.L.ValenbergH.J.F.V.DijkstraJ.HooijdonkA.C.M.V. 2009. Seasonal variation in the Dutch bovine raw milk composition. J. Dairy Sci. 92:4745-4755.
- Kris-EthertonP.M. 1999. AHA Science advisory: monounsaturated fatty acids and risk of cardiovascular disease. J. Nutr. 129:2280-2284.
- LongC.RCartwrighT.C.FitzhughH.A.Jr. 1975. Systems analysis of sources of genetic and environmental variation in efficiency of beef production: cow size and herd management. J. Anim. Sci. 40:409-420.
- LopezC. 2005. Focus on the supramolecular structure of milk fat in dairy products. Reprod. Nutr. Dev. 45:497-511.
- LopezC. 2011. Milk fat globules enveloped by their biological membrane: Unique colloidal assemblies with a specific composition and structure. Curr. Opin. Colloid. In. 16:392-404.
- MacciottaN.P.P.VicarioD.Cappio-BorlinoA. 2005. Detection of different shapes of lactation curve for milk yield in dairy cattle by empirical mathematical models. J. Dairy Sci. 88:1178-1191.
- MaglianoA. 1950. Ezoognosia generale. Vallardi Ed., Milano, Italy.
- ManirakizaP.CovaciA.SchepensP. 2001. Comparative study on total lipid determination using Soxhlet, Roese-Gottlieb, bligh & dyer, and modified bligh & dyer extraction methods. J. Food Compos. Anal. 14:93-100.
- ManssonH.L. 2008. Fatty acids in bovine milk fat. Food Nutr. Res. 52:10-34.
- MartiniM.AltomonteI.SalariF. 2013. Evaluation of the fatty acid profile from the core and membrane of fat globules in ewe’s milk during lactation. LWT-Food Sci. Technol. 50:253-258.
- MartiniM.ScolozziC.CecchiF.MeleM.SalariF. 2008. Relationship between morphometric characteristics of milk fat globules and the cheese making aptitude of sheep’s milk. Small Ruminant Res. 74:194-201.
- MatassinoD.BaroneC.M.A.CastellanoN.GiganteG.GrassoM.ZulloA. 2011. Effect of autochthonous bovine genetic type on some quanti-qualitative characteristics of milk. Ital. J. Anim. Sci. 10(Suppl.1):13 ( abstr.).
- MacciottaN.P.P.SerraA.BuccioniA.PagnaccoG.SecchiariP. 2007. Stearoyl-Coenzyme A desaturase gene polymorphism and milk fatty acid composition in Italian Holsteins. J. Dairy Sci. 90:4458-4465.
- MeregalliA. 1980. Conoscenza morfo-funzionale degli animali domestici. Liviana Ed., Padova, Italy.
- MichalskiM.C.BriardV.MichelF.TassonF.PoulainP. 2005. Size distribution of fat globules in human colostrum, breast milk, and infant formula. J. Dairy Sci. 88:1927-1940.
- MichalskiM.C.CariouRMichelF.GarnierC. 2002. Native vs. damaged milk fat globules: membrane properties affect the viscoelasticity of milk gels. J. Dairy Sci. 85:2451-2461.
- Molto-PuigmartìC.CastelloteA.I.Lopez-SabaterM.C. 2007. Conjugated linoleic acid determination in human milk by fastgas chromatography. Anal. Chim. Acta 602:122-130.
- OreficeG. 1978. Studio dei caratteri morfologici e produttivi di una razza bovina autoctona della valle d’Aveto. Degree Diss., University of Pisa, Italy.
- RandolphH.E.ErwinR.E. 1974. Influence of mastitis on properties of milk.X. Fatty-acid composition. J. Dairy Sci. 57:865-868.
- RayD.E.HalbachT. J.ArmstrongD. V. 1992. Season and lactation number effects on milk production and reproduction of dairy cattle in Arizona. J. Dairy Sci. 75:2976-2983.
- RekayaR.CarabanoM.J.ToroM.A. 2000. Bayesian analysis of lactation curves of Holstein-Friesian cattle using a non linear model. J. Dairy Sci. 83:2691-2701.
- SabbioniA.BerettiV.SuperchiP. 2008. Curve di lattazione in bovine di razza Valdostana. Sci. Tecn. Latt. Cas. 59:345-349.
- SabbioniA.BerettiV.TardiniL.VezzaliS.PainiV.SuperchiP. 2012. Milk production and lactation curves of Bianca Val Padana and Italian Friesian dairy cows in relation to the management system. Ital. J. Anim. Sci. 11:e26.
- SabbioniA.BerettiV.TardiniL.VezzaliS.PainiV.SuperchiP. 2013. Frequenza delle curve di lattazione standard e atipiche per latte, grasso, proteine e lattosio in bovine di razza Bianca Val Padana. Sci. Tecn. Latt. Cas. (In press).
- SabbioniA.BerettiV.ZanonA.FranceschiP. 2003. Factors affecting the shape of the lactation curve in Reggiana cattle. Ital. J. Anim. Sci. 2(Suppl.1):278-280.
- SabbioniA.SuperchiP.BerettiV.MalacarneM.QuagliatiC.SummerA. 2011. Curve di lattazione in bovine di razza Grigio Alpina. Sci. Tecn. Latt. Cas. 62:301-308.
- SAS, 2008. User’s Guide: Statistics, ver. 9.2. SAS Inst. Inc., Cary, NC, USA.
- SchenninkA.HeckJ.M.L.BovenhuisH.ViskerM.H.P.W.ValenbergH.J.F.V.ArendonkJ.A.M.V. 2008. Milk fatty acid unsaturation: genetic parameters and effects of Stearoyl-CoA desaturase (SCD1) and Acyl CoA: diacylglycerol acyltransferase 1 (DGAT1). J. Dairy Sci. 91:2135-2143.
- SchwartzG.J.FuJ.AstaritaG.LiXGaetaniS.CampolongoP.CuomoV.PiomelliD. 2008. The lipid messenger OEA links dietary fat intake to satiety. Cell Metab. 8:281-288.
- ShookG.E.SchutzM.M. 1994. Selection on Somatic Cell Score to improve resistance to mastitis in the United States. J. Dairy Sci. 77:648-658.
- ShortenP.R.PleasantsT.B.UpretiG.C. 2004. A mathematical model for mammary fatty acid synthesis and triglyceride assembly: the role of stearoyl CoA desaturase (SCD). J. Dairy Res. 71:385-397.
- SieberM.FreemanA.E.KelleyD.H. 1988. Relationships between body measurements, body weight, and productivity in Holstein dairy cows. J. Dairy Sci. 71:3437- 3445.
- SoyeurtH.DeharengF.MayeresP.BertozziC.GenglerN. 2008. Variation of Δ9- desaturase activity in dairy cattle. J. Dairy Sci. 91:3211-3224.
- StålhammarH.HenningssonT.PhilipssonJ. 1995. Factors influencing ultrasonic scanning measures, muscularity scores and body measures in performance-tested dairy bulls and their usefulness as predictors of beef production ability in Friesian cattle. Acta Agr. Scand. A-An. 47:230-239.
- StrapakP.JuhasP.StrapakovaE. 2011. The relationship between the length of productive life and the body conformation traits in cows. J. Cent. Eur. Agr. 12:239-254.
- UsaiG. 1974. Ricerche biometriche sulla razza bovina Cabannina in provincia di Genova. Specialization Diss., University of Milano, Italy.
- WoodP.D.P. 1967. Algebraic model of the lactation curve in cattle. Nature 216:164-165.