Abstract
The aim of this trial was to study the effect of supplementary feeding offered to farmed fallow deer does on milk characteristics and on male and female fawns performance. During two years, 40 females were divided in two groups and assigned to two homogeneous paddocks of the same pasture: SUP group received 0.5 kg/d per female of supplement that was raised to 0.7 kg/d from the third month of fawns age, while HER group was fed only on herbage. Herbage availability and composition were monitored by exclusion cages. Female performance (body weight, body condition score and milk composition) was recorded. Further, during the first year, performance and hematological parameters (non-esterified fatty acids and urea nitrogen) were assessed. Fawns were weighed every 45 days until the end of the experiment, when the characteristics of their carcasses were recorded. Supplementary feeding reduced herbage gathering and quality, but allowed a full recovery of body weight and condition of does. The milk provided by the HER does to their female offspring was richer in fat (16.1 vs 14.2 g/100 mL) and protein (8.76 vs 8.04 g/100 mL) than that furnished to male fawns; the opposite condition happened for SUP does (12.6 vs 13.3 g/100 mL for fat and 8.04 vs 8.35 g/100 mL for protein). At slaughter, SUP fawns showed higher carcass weight (14.10 vs 11.26 kg), greater conformation score (2.1 vs 1.6 points) and higher fat score (2.0 vs 1.2 points) than HER ones, moreover male fawns presented higher carcass weight than female (13.62 vs 11.74 kg).
Introduction
Fallow deer (Dama dama) is a strongly dimorphic deer species; in Italy it is found in many natural environments characterized by mild climatic conditions, since this species is negatively affected by a harsh winter climate. Further, fallow deer are often reared for commercial purposes under both extensive and intensive conditions, since they are able to exploit effectively less favored areas (Mattiello et al., Citation2009; Piasentier et al., Citation2005).
The supplementary feeding with concentrates is often necessary to meet the nutrient requirements of animals in winter, in case of adverse weather conditions and, in general, in periods of pastoral feed deficiency (Ru et al., Citation2003). When a pasture is heavily utilized, supplementary feeding should start early, in summer-autumn, to allow animals facing winter in good conditions (Reinken, Citation1990).
Some studies reported the effect of supplementary feeding on growth rates (Saccà et al., Citation2001) and on carcass and meat characteristics in fallow deer (Volpelli et al., Citation2002; Citation2003; Wiklund et al., Citation2005). Hutchinson et al. (Citation2012) showed higher body condition of non-pregnant fallow deer does when fed with concentrate. According to studies showing variations in hind phenotypic traits and milk composition in red deer (Cervus elaphus) (Gallego et al. Citation2005; Landete-Castillejos et al. Citation2005; Citation2003; Citation2000) as a function of feeding regimes, we expect important effects also in fallow deer. Such a study is relevant, since this species has been much less studied than red deer.
The objective of this trial was to study the effect of supplementary feeding on farmed fallow deer does on milk characteristics and on male and female fawns performance.
Materials and methods
The study was carried out in accordance with the rules provided by European Parliament and Council Directive 2010/63/EU for animal experiments.
Weather conditions, experimental design and feed characteristics
The experiment was conducted on a temperate pasture of Festuca arundinacea in northeast Italy (latitude 46°07’33’’ N, longitude 13°10’14’’E; altitude 170 m), at the facilities of Udine University, from early May to early November, over two years. The climatic condition for the area was comparable between years concerning temperature; an average daily temperature of 19°C (2.4°C minimum and 28.8°C maximum values) were recorded from June to early November. In contrast, year 1 was drier than year 2, which showed a more regularly distributed rainfall during the experimental period. Indeed, while in year 1, there were 20 days of rain with a total rainfall of approx. 300 mm, in year 2 the data were 30 days and approx. 550 mm, respectively. The difference in rainfall between years was essentially due to the dry weather in July and August of year 1.
In early May, 20 fallow deer (Dama dama) females, were assigned to two homogeneous paddocks of the same pasture of approximately 10,000 m2, according to a similar stocking density (500 m2/head) and the number of pregnant animals. Consequently, during the experimental periods, the first paddock was grazed by 11 animals, 7 lactating does (LAC) and 4 yearling females (NLC), the second by 9 animals, 6 LAC and 3 NLC. Every year, from first fawn birth (mid-June), the females of the two paddocks were submitted to a different feeding treatment, which was switched between paddocks in the two years. The first group (SUP) received 0.5 kg/d per female of supplement that was raised to 0.7 kg/d per female from the third month of fawns age, while the second group (HER) was fed only on grazed herbage.
The supplement utilized was a commercial concentrate mixture (comprising maize meal, wheat bran, maize gluten feed, soybean meal, sunflower seeds meal, dried brewers’ grain, wheat middlings, barley meal, dried lucerne meal, molasses, minerals and vitamins) and was fed at once in daily meals. The concentrate offered was made up of crude protein 19.6% dry matter (DM), ether extract 3.4% DM, ash 10.1% DM, crude fibre 11.5% DM, and 12.45 MJ metabolizable energy (ME) /kg organic matter (OM).
The herbage mass was evaluated using exclusion cages (Meijs et al., Citation1982) cutting the herbage at about 3 cm above the ground level within three cages of 1 m2 (1×1 m) and within three other grazed areas close to the cages. The cages were randomly located and moved every three weeks. The differences between the herbage quantities collected inside (excluded areas) and outside (grazed areas) the cage were considered as an estimate of the herbage gathered by animals while grazing for a three-week period. The samples of herbage collected on the excluded areas and a monthly sample of the concentrate were dried at 65 °C in a forced oven draught, and analysed for crude protein (CP) and ether extract (EE) according to AOAC (Citation2000), and for neutral detergent fibre (NDF) according to Goering and Van Soest (Citation1970). The nutritive value, expressed as megajoules of metabolizable energy per kg of organic matter (MJ EM/kg OM), was estimated in accordance with the INRA standards (Vermorel, Citation1988).
Animal measurements
During the first year, females were captured in early May and mid September (together with their fawns), using a standard deer crush with a drop floor which allowed us to weigh animals [body weight (BW) accurate to 0.1 kg], to assess the body condition score (BCS), and to perform milk and blood collection. The BCS was assessed by two independent evaluators, according to a 5-point scale method, as an estimate of the nutritional condition of does (Flesch et al., Citation2002). The day after birth, the fawns were weighed (birth weight) using a dynamometer (accurate to 0.05 kg). For capturing newborns, systematic observations were performed from vantage points. In some cases, parturition was observed or the doe showed signs of recent delivery. Females were always checked in the morning and evening, so that we could establish if parturition had occurred. Most captures were performed after observation of the fawn, but in some cases, fawns were detected upon systematic search. Fawns were captured using a long-handled (150 cm) circular net (diameter 80 cm), as described by Galli et al. (Citation2008). To avoid desertion by the mother, we waited the day after delivery before capturing the newborn and latex gloves were always used when handling animals. During the capture, fawns were also sexed and marked with a numbered plastic ear-tag.
The first year, the females were captured in the crush every 45 days from the beginning of lactation, to assess BW and BCS and to perform blood and milk collection. In the same year, fawns were also weighed in the crush every 45 days until slaughtering, at 135 days of age. Immediately after slaughter the hot carcass weight of fawn was recorded and the dressing percentage calculated. On the same day, the carcasses were graded for conformation and fatness on the basis of Commission Regulations (EEC) No. 461/93 and Council Regulation (EEC) No. 2137/92. In particular, three parts of the carcasses (shoulder, back, round) were independently assessed according to a scale ranging from 1 (poor) to 5 (excellent) and the average was used to assign a conformation score to each animal. On the other hand, the fat score for each animal was calculated as mean of 3 scores obtained by the independent visual assessment of the amount of external fat, feathering and thoracic fat, kidney knob and channel fat according to a scale ranging from 1 (very lean) to 5 (very fat). Carcasses were first kept for 6 h at room temperature (12-15 °C), and then chilled for 18 h at 4°C.
Milk collection and analysis
Milk collection was performed on lactating does by hand after an intramuscular injection of 12 U of oxytocin (Zoovet Srl, Cesena, FC, Italy). Each milk sample (about 23 mL) was added to 200 L of Bronopol (Rohm and Haas, Philadelphia, PA, USA) and stored at -20°C until analysis. An infrared MilkoScan System 6000 (Foss Electric, Hillerød, Denmark) was used to determine fat, protein and lactose using water buffalo standards. The results were expressed as g/100 mL milk. Milk was also analysed for total solids, according to AOAC (Citation2000), and the results were expressed as g/100 mg milk.
Blood collection and analysis
Blood was collected from the jugular vein (10 mL) into sterile heparinized tubes. Within 1 h from collection, plasma was separated by centrifugation (10 min) at 2000 g and stored at -20°C until analysis. Non-esterified fatty acids (NEFA) (Wako Chemicals GmbH, Neuss, Germany) and urea nitrogen (BUN) (Eurokit Srl, Gorizia, Italy) were analysed.
Statistical analysis
The statistical analyses were performed using SPSS version 17 software (SPSS Inc., Chicago, IL, USA). Normality of data distribution was tested by the Kolmogorov-Smirnov test. The characteristics of the pasture were analyzed using GLM repeated measures procedure considering diet (SUP and HER group) as between-subject factors, time of sampling (T) as a within-subject factor and year as a block. The two-year performance of does and fawns were analysed by analysis of variance, considering year as a block, and taking into account the following factors and their interactions: diet and physiological stage (lactating LAC vs non-lactating NLC) or fawn gender (male vs female). The year effect was not included in the tables because it never reached a level of significance.
The variables monitored only in year one were analysed using the repeated measures procedure of the general linear model (GLM), considering the recording time as a within-subject factor and the other, already mentioned factors (diet, physiological stage or fawn’s gender), as between-subject factors. For multiple comparisons.
Bonferroni adjustments were made.
Two-way ANOVA was performed for carcass quality, composition, and meat composition of fawn considering diet and gender as fixed effects.
A probability level of P≤0.05 was established for statistical significance; and P<0.10 indicated a trend.
Results and discussion
Herbage availability, body condition and blood metabolites
reports the available herbage mass and composition. In both years and without significant differences between them, the paddock grazed by the SUP group showed a higher forage availability than HER group (232 vs 131 g OM/m2; P<0.01). This condition was determined by a different herbage intake of the experimental herds, which was roughly estimated by the exclusion cage technique. Indeed, the average herbage removed in a three week-period by the animals tended to be higher in HER than SUP group (101 vs 65 g OM/m2; P=0.07). This result is in close agreement with the findings of Ru et al. (Citation2003), which showed that herbage intake decreases with the increase in the supplement level. There was a significant pattern of pasture biomass availability during the experimental period (T effect, P<0.01). The forage allowance followed a trend that was affected by feeding treatment (D×T effect, P<0.01). In particular, as depicted in , after starting from a homogenous pasture availability of 175 g OM/m2, herbage allowance in the control paddock showed a tendency to a steadily decline to a residual level of 91 g OM/m2. On the contrary, the OM available in the supplemented paddock increased during summer to levels over 250 g OM/m2, and then declined to a final residual value of approx. 200 g OM/m2. These results were probably due to a seasonal trend and to the increasing of intake by the growing fawn in the last six weeks.
The composition of the herbage sampled in the exclusion cages is an estimate of the forage available for animal selection during grazing. It was relatively poor in protein, high in fibre fractions and it had a mean nutritional value of 10.2 MJ ME/kg OM. The herbage of SUP group presented slightly lower CP (12.2 vs 12.7 %DM; P=0.10) and higher NDF level (63.0 vs 58.7% DM; P<0.01) than HER group. The different chemical composition between paddocks was related to the grazing intensity, higher in HER than SUP group. Ru and Fortune (Citation1996) reported that grazing intensity enhanced the protein level of subterranean clover favouring the re-growths of plant and changing the leaf/steam ratio. In general, many authors argue that plant nutritional value is the result of a complex interaction between grazing intensity and management, species and plant’s physiological stage (Ru and Fortune, Citation2000; Ru et al., Citation2003). During the trial, the chemical composition of herbage was variable (P<0.01), and reached the best nutritive conditions in late spring (10.98 MJ ME/kg OM) and early September (10.44 MJ ME/kg OM).
The yearling not lactating females were included in the experiment because they are normal components of the herd, they allowed balancing the stocking rate between paddocks and, mainly, they permit a comparison of the effect of diet on females in different physiological states. NLC group, had lower body weight (; 33.6 vs 44.4 kg on average; P<0.01) than LAC at early-May, when groups were formed; however the former deer were younger (23 vs 42 months, data not reported in Table) than the latter. Consequently, NLC had not reached the adult weight during the trial, when they continue their growth at a rate that was positively influenced by feeding supplementation (6.4 vs 4.7 kg in SUP and HER groups, respectively; P<0.05). LAC, which were selected at a comparable body weight, underwent a diet dependent weight variation during the experimental period, indeed while supplemented lactating does (SUP, LAC) showed a full recovery of body weight, not supplemented lactating does (HER, LAC) did not (), even in early-fall (), likely as a consequence of the already described herbage shortage (). By contrast, the lactating does receiving the concentrate recover their weight () and they continued their positive body variation even in autumn (), likely favoured by the reduction of milk yield in late lactation. Landete-Castillejos et al. (Citation2000) considering hinds (Cervus elaphus hispanicus) showed a reduction of body weight during lactation; instead Loudon et al. (Citation1984) hypothesized that hinds are able to recover body weight from mid to late lactation. The trend of body weight was confirmed by that of body condition. Indeed, starting from a comparable condition score in early-May (2.75 points, ), SUP and HER groups showed a different evolution during the experimental period reflecting the availability of energy for fallow deer. The HER, LAC group lost half of a point of body score during lactation without any recovering during fall. Instead, SUP, LAC group underwent a smaller initial loss (a quarter of a point) that they completely regained before the end of lactation, during which they preserved a higher body condition than HER group (). These results are in agreement with Wiklund et al. (Citation2005) who showed higher BCS in fallow deer supplemented with barley than those raised on pasture. Asher et al. (Citation2000) considering 36 hinds across two years, found that BCS decreased and increased during lactation in the first and second year respectively, suggesting that the pasture condition is able to modify the BCS variations. Following the suggestions of Mulley and Flesch (Citation2001), who recommend a BCS of 3 points, and at least higher than 2, to avoid compromising the reproductive performances of fallow deer, the body condition of the HER, LAC does at the beginning of the mating season was not adequate, suggesting to practise an earlier weaning in these conditions. Conversely, herbage consumption provided enough feed to HER,NLC yearlings to face successfully their incoming first reproductive season.
For a better understanding of diet effect on animals’ physiological stage, the evolution of some blood metabolites was monitored and reported in . It is well known that NEFA are related to ruminant energy balance, and in particular to fatty acid mobilization (Dunshea and Bell, Citation1989). LAC does showed higher plasma NEFA (P<0.01) than yearlings in accordance to their high-energy requirements for lactation and their body fat mobilization. Moreover, in both LAC groups, NEFA concentration was significantly affected by recording time (). Indeed, NEFA decreased during the experimental period from a common initial value of 0.79 mmol/L, particularly during late lactation when a reduction of milk yield could be assumed. However, in early-November, in agreement with the body changes described above, HER, LAC does presented the highest plasma levels of NEFA: 0.62 vs 0.47 mmol/L of the other groups of fallow deer females.
The mean plasma urea nitrogen level was 8.67 mmol/L (). Other studies have found BUN ranging from 5.4 to 11.1 mmol/L (Polji ak-Milas et al., Citation2006, Citation2009; Vengušt and Bidovec, Citation2002). The level of BUN is influenced by the rumen degradable protein intake, ruminal nitrogen balance, urea recycling, and by the tissue catabolism (Saltz et al., Citation1995). Considering the slight fluctuation of herbage CP content, and despite BUN, up to a value of 10.7 mmol/L, is considered a suitable indicator of CP intake (Sun and Christopherson, Citation2005), a recording time effect was observed (P<0.01). Indeed, both in LAC and NLC group BUN increased during season, from an average value of 6.6 mmol/L in early August to 11.5 mmol/L in early November. Waid and Warren (Citation1984) in white-tailed deer suggested that nutritional stress could increase the BUN level by affecting nitrogen recycling.
In our study, considering the body weight variation previously discussed, a nutritional stress should not be hypothesised. Most likely, the BUN increasing could be due to the ruminal nitrogen to energy balance, and in particular to the reduction of dietary fermentable sugar.
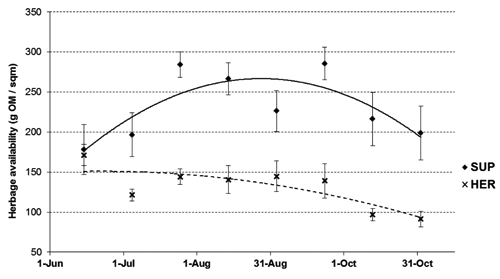
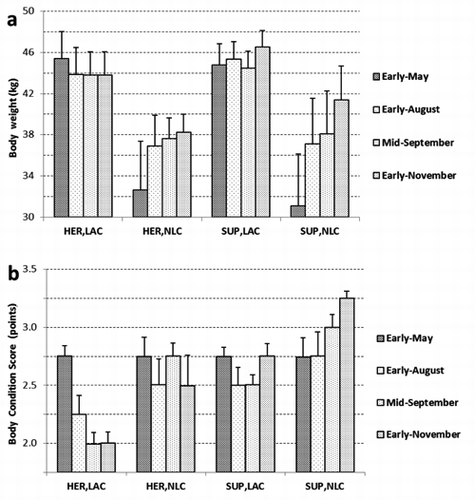
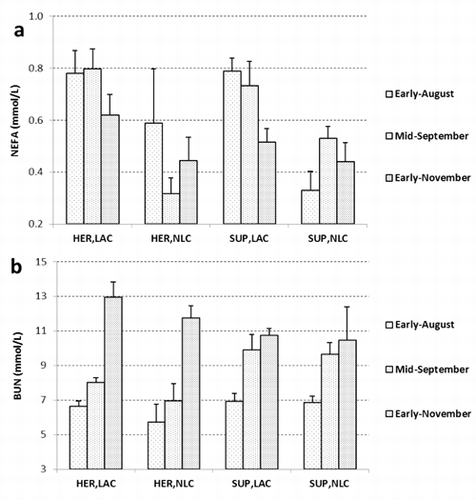
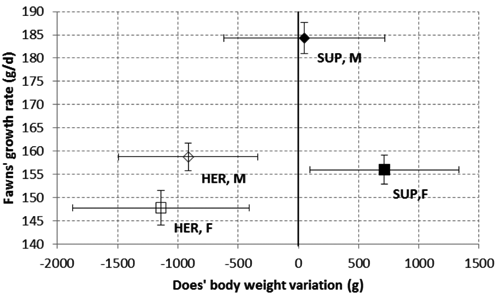
Table 1. Effect of diet on herbage availability and composition, evaluated for two years every three weeks from the beginning of the trial
Table 2. Effect of diet and physiological stage on body weight and body condition score of fallow deer.
Table 3. Effect of diet and fawn’s gender on milk composition of lactating fallow deer recorded 90 days after parturition and fawns’ performance
Table 4. Evolution of milk composition of lactating fallow deer (n=13) recorded 45 (early), 90 (mid) and 135 (late) days after parturition (first year)
Table 5. Effect of fawn’s gender and does diet on carcass quality of fawns (first year)
Milk composition and fawn performance
The milk nutrients concentration, which followed in the range of values reported by Pisani et al. (Citation2006) and Reinken et al. (Citation1990) in fallow deer, was significantly affected by the experimental factors, that showed interesting interactions between them (). Actually, the milk provided by the HER does to their female offspring was richer in fat (16.1 vs 14.2 g/100 mL) and protein (8.76 vs 8.04 g/100 mL) than that furnished to male fawns; the opposite condition happened for SUP does (12.6 vs 13.3 g/100 mL and 8.04 vs 8.35 g/100 mL, respectively for fat and protein in female and male offspring). The milk of the SUP group revealed higher lactose (4.5 vs 4.2 g/100 mL) content than HER milk without difference between genders. Moreover, as described in , fat content increased during lactation, with a linear effect of time that doubled its concentration between early and late suckling period (from 10.0 to 20.4 g/100 mL). In parallel, while the protein content of milk, in agreement with fat, significantly increased from 5.66 to 9.29 g/100 mL, the lactose followed an opposite trend, decreasing with a quadratic path, from 4.91 to 4.27 g/100 mL, during lactation. Shahbazkia et al. (Citation2010) showed a strong positive correlation between lactose yield and milk volume (r=0.958). Indeed, one of the main functions of lactose is to determine the osmolarity of milk. The above-cited authors explained that lactose favors the increase of water in milk with the result of increasing the volume and dilute the nutrients content. Taking together, these results suggest that from 45 to 135 days of lactation, as physiologically expected, fallow deer does experienced a steady reduction of milk yield but also that they would have produced more milk in SUP instead of HER feeding treatment and when they were suckled by male instead of female offspring. Moreover, while the HER does would have provided their male offspring of a poorer milk than their female fawns, the SUP mothers would have fed their male sons with a richer milk than the female ones. These hypotheses justify the observed growth rate of the different classes of fawns (). Considering a similar birth weight (4.42 kg), the SUP-male fawns showed the highest growth rate (186 g/d vs 159 g/d in the first 90 days), because of a positive interaction between gender and feeding treatment of the mother.
At slaughter, SUP fawns showed higher carcass weight than HER ones (, 14.10 vs 11.26 kg; P<0.01), and an ordinal interaction between diet and fawn’s gender was found (P<0.05). The dressing percentage was higher in SUP than HER animals (52.0 vs 47.9%; P<0.01); the average dressing percentage was similar to those reviewed by Russo et al. (Citation2005) in fawns slaughtered at 27 kg of BW. Despite the fact that male fawns presented higher carcass weight than female (P<0.05), the fawn gender has not influenced the carcass killing out (P>0.05). Also Summer et al. (Citation1997) slaughtering 33 males and 44 females of fallow deer at different live weights have not found differences in the dressing percentage.
The SUP group presented fawns with greater conformation score (2.1 vs 1.6 points; P<0.01) and higher fat score (2.0 vs 1.2 points; P<0.01) than HER group. Volpelli et al. (Citation2002) showed greater muscle development and fat deposition in carcasses of fallow deer that received a daily amount of feed supplement equal to 1.25 % of their initial weight than those fed only on herbage. As for carcass weight, the interaction between experimental factors was significant for conformation and fat score (P<0.05), but ordinal from the perspective of diet. It means that, as for growth rate, the positive effect of the mother diet supplementation was greater in male than female litter.
Even if the experiment was not specifically designed to test this hypothesis, the results seem to be consistent with the theory of maternal investment (Trivers and Willard, Citation1973) that explains how mothers of sons produced greater yields of milk with higher protein content than daughters’ mothers. In fact, this investment could be done in different not alternative ways: differences in sex ratio; sex differences in pre- and post-natal growth; and sex differences in suckling behaviour (Byers and Moodie, Citation1990; Clutton-Brock, Citation1991; Pelabon et al., Citation1995). Some studies carried out in Sweden have shown that the adjustment of the sex ratio according to ability to invest does not occur in fallow deer (Birgersson et al., Citation1988b), while the different suckling behaviour of male fawns allowed them receiving more milk and growing more than females. In addition to this, mothers who raised sons accumulated less body mass than those raising daughters, during the period between late gestation and the end of lactation (Birgersson, Citation1988a). In the four does experimental conditions were scattered in a bi-dimensional plot, defined by the does’ body mass variation against the fawns’ growth. The HER does did not present a positive body mass variation raising either male or female fawns, probably because the herbage allowance was not able to fully satisfy the nutritional requirements of the herd (Asher, Citation1993). Conversely, the SUP treated does showed a differential maternal investment. Indeed, while the does suckling a son preferentially addressed the incremental nutritive resources for a faster offspring growth, the does suckling a daughter preferentially addressed them for improving her fitness for the incoming mating season.
Conclusions
This trial shows that several beneficial effects can be observed by supplementing with concentrate (25/30% of DM intake) grazing lactating fallow deer. Specifically, the supplementation improves the body condition score of does, increases the nutrients content of milk, comprising lactose, which implies a higher milk yield. Moreover, it allows the production of heavier fawns with higher performance at slaughter in terms of dressing percentage, conformation and fatness of carcasses.
Furthermore, taking into account our results, a different maternal investment could be hypothesized. Indeed, supplementing grazing lactating fallow deer would preferably address the incremental nutritional resources of does towards males fawn growth instead of maternal body condition recover.
References
- AOAC, 2000. Official methods of analysis, 17th ed. Association of Official Analytical Chemistry, Gaithersburg, MD, USA.
- AsherG.W. 1993. Growth and feeding management of fallow deer in New Zealand. pp 67-72 in Proc. 1st World Forum on fallow deer farming. Ruakura Agricultural Centre, Hamilton, New Zealand.
- AsherG.W. MulleyR.C. O’NeillK.T. ScottI.C. 2000. Studies on the nutritional requirements of adult red deer hinds during late pregnancy. pp 97-104 in Proc. 17th Nat. Conf. Deer Branch of the New Zealand Veterinary Association, Queenstown, New Zealand.
- BirgerssonB. 1988a. Male-biased maternal expenditure and associated costs in fallow deer. Behav. Ecol. Sociobiol. 43: 87-93.
- BirgerssonB. TillbomM. EkvallK. 1988b. Male-biased investment in fallow deer: an experimental study. Anim. Behav. 56:301-307.
- ByersJ.A. MoodieJ.D. 1990. Sex-specific maternal investment in pronghorn, and the question of a limit on differential provisioning in ungulates. Behav. Ecol. Sociobiol. 26:157-164.
- Clutton-BrockT.H. 1991. The evolution of parental care. Princeton University Press, Princeton, NJ, USA.
- Clutton-BrockT.H. HarveyP.H. 1978. Mammal resources and reproductive strategies. Nature 273:191-195.
- DunsheaF.R. BellA.W. 1989. Relations between plasma non-esterified fatty acid metabolism and body fat mobilization in primiparous lactating goats. Brit. J. Nutr. 62:51-61.
- FleschJ.S. MulleyR.C. AsherG.W. 2002. Development of a body condition scoring system for farmed fallow deer (Dama dama). pp 20-26 in Proc. 5th Int. Congr. on Deer Biology, Québec City, Canada.
- GallegoL. Landete-CastillejosT. GarciaA. SánchezP.J. 2005. Seasonal and lacta-tional changes in mineral composition of milk from Iberian red deer (Cervus elaphus hispanicus). J. Dairy Sci. 89:589-595.
- GalliM.A. FocardiS. IseppiM. PiasentierE. 2008. Age determination in fallow deer Dama dama neonates. Wildlife Biology 14:386-390.
- GoeringH.K. Van SoestP.J. 1970. Forage fibre analyses (apparatus, reagents, procedures, and some applications). Agricultural Handbook N. 379, US Department of Agriculture Publ., Washington, DC, USA.
- HutchinsonC.L. MulleyR.C. WiklundE. FleschJ.S. 2012. Effect of concentrate on instrumental meat quality and sensory characteristics of fallow deer venison. Meat Sci. 90:801-806.
- Landete-CastillejosT. GarcíaA. GόmezJ.A. GallegoL. 2003. Lactation under food constraints in Iberian red deer (Cervus elaphus hispanicus). Wildlife Biol 9: 131-139.
- Landete-CastillejosT. GarcíaA. Lόpez SerranoF.R. GallegoL. 2005. Maternal quality and differences in milk production and composition for male and female Iberian red deer calves (Cervus elaphus hispanicus). Behav. Ecol. Sociobiol. 57: 267-274.
- Landete-CastillejosT. GarcíaA. MolinaP. VergaraH. GardeJ. GallegoL. 2000. Milk production and composition in captive Iberian red deer (Cervus elaphus hispanicus): effect of birth date. J. Anim. Sci. 78:2771-2777.
- LoudonA.S.I. DarrochA.D. MilneJ.A. 1984. The lactation performance of red deer on hill and improved species pastures. J. Agr. Sci. 102:149-158.
- MattielloS. 2009. Welfare issues of modern deer farming. Ital. J. Anim. Sci. 8:205-217.
- MeijsJ.A.C. WaltersR.J.K. KeenA. 1982. Sward methods. In: LeaverJ.D. ( ed.) Herbage intake handbook. British Grassland Society Publ., Berkshire, UK, pp 11-36.
- MulleyR.C. FleschJ.S. 2001. Nutritional requirements for pregnant and lactating red and fallow deer. Report N. 01/0095. Rural Industries Research and Development Corporation Publ., Canberra, Australia.
- PelabonC. GaillardJ.M. LoisonA. PortierC. 1995. Is sex-biased maternal care limited by total maternal expenditure in polyg-ynous ungulates? Behav. Ecol. Sociobiol. 47:311-319.
- PiasentierE. BovolentaS. VilianiM. 2005. Wild ungulate farming systems and product quality. Vet. Res. Comm. 29 (Suppl.2):65-70.
- PisaniG.M. MalacarneM. SoffiantiniC.S. FranceschiP. FormaggioniP. PiasentierE. SummerA. MarianiP. 2006. Gross composition and protein fractions of milk from fallow deer (Dama dama). Page 51 in Proc. 6th Int. Congr. on Deer Biology, Prague, Czech Republic.
- Poljičak-MilasN. MarenjakT.S. SlavicaA. JanickiZ. FilipoviN. SrukV. 2009. Comparative hematological and biochemical values in pregnant and non-pregnant red, (Cervus elaphus), and fallow deer, (Dama dama), females. Folia Zool. 58:36-44.
- Poljičak-MilasN. SlavicaA. JanickiZ. MarenjakT.S. KoliE. 2006. Comparison of serum biochemical parameters between red (Cervus elaphus) and fallow deer (Dama dama L.) in Moslavina region of croatia. Vet. Arhiv. 76:229-238.
- ReinkenG. HartfielW. KörnerE. 1990. A pratical guide to german techniques. Farming Press Books, Ipswich, UK.
- RuY.J. FortuneJ.A. 1996. Hard grazing during spring growth improved nutritive value of subterranean clover. Page 423 in Proc. 21st Biennal Nat. Conf. Australian Society of Animal Production, Brisbane, Queens -land, Australia.
- RuY.J. FortuneJ.A. 2000. The effect of grazing intensity and cultivars on morphology, phenology and nutritive value of subterranean clover. II. Nutritive value during the growing season. Aust. J. Agric. Res. 51:1047-1055.
- RuY.J. FischerM. GlatzP.C. WyattS. SwansonK. FalkenbergS. 2003. Forage intake and nutrient requirements of fallow weaner deer in Southern Australia. Asian Austral. J. Anim. 16:685-692.
- RussoC. 2005. Carcass and meat quality in fallow deer (Dama dama). A review. Ann. Fac. Med. Vet. Pisa 58:207-212.
- SaccàE. VolpelliL.A. BovolentaS. PiasentierE. PinosaM. 2001. Effect of supplementary feeding on pasture utilization by growing fallow deer. Zootec. Nutr. Anim. 27:33-39.
- SaltzD. WhiteG.C. BartmannR.M. DelGiudiceG.D. RiggsM.R. MechL.D. SealU.S. 1995. Assessing animal condition, nutrition, and stress from urine in snow: a critical view and response. Wildlife Soc. B. 23:694-704.
- ShahbazkiaH.R. AminlariM. TavasoliA. MohamadniaA.R. CravadorA. 2010. Associations among milk production traits and glycosylated haemoglobin in dairy cattle; importance of lactose synthesis potential. Vet. Res. Commun. 34:1-9.
- SummerA. SussiC. MartuzziF. CatalanoA.L. 1997. Rilievi alla macellazione, prove di sezionamento e composizione chimica della carne di daino (Dama dama) e di cervo (Cervus elaphus). Ann. Fac. Med. Vet. Parma. 17:253-262.
- SunS. ChristophersonR.J. 2005. Urea kinetics in wethers exposed to different ambient temperatures at three dietary levels of crude protein. Asian Austral. J. Anim. 18:795-801.
- TriversR. L. WillardD.E. 1973. Natural selection of parental ability to vary the sex ratio of offspring. Science 179:90-92.
- VenguštG.A. BidovecA. 2002. Some serum chemistry values of fallow deer (Dama dama l.) in Slovenian hunting enclosures. Vet. Arhiv. 72:205-212.
- VermorelM. 1988. Nutrition energetique. pp 57-74 in JarrigeR. ( ed.) Alimentation des Bovins, Ovins, Caprins. Institut National Recherche Agronomique Publ., Paris, France.
- VolpelliL.A. ValussoR. MorganteM. PiasentierE. 2003. Meat quality in male fallow deer (Dama dama): effects of age and supplementary feeding. Meat Sci. 65:555-562.
- VolpelliL.A. ValussoR. PiasentierE. 2002. Carcass quality in male fallow deer (Dama dama): effects of age and supplementary feeding. Meat Sci. 60:427-432.
- WaidD.D. WarrenR.J. 1984. Seasonal variations in physiological indices of adult female white-tailed deer in Texas. J. Wildlife Dis. 20:212-219.
- WiklundE. HutchisonC. FleschJ. MulleyR. LittlejohnR.P. 2005. Colour stability and water holding capacity of M. longis-simus and carcass characteristics in fallow deer (Dama dama) grazed on pasture or fed barley. Rangifer 25:97-105.