
Abstract
Whole mature seeds of eight selected varieties, subspecies and accessions of three Vicia L. species grown in Tunisia were investigated for their fatty acid (FA) profile. The FA composition ranged from lauric (C12:0) to lignoceric (C24:0) acids. The total FA content was 1235.14 to 1580.34 mg 100 g–1 dry matter (DM). Linoleic acid (C18:2 c 9c 12; 647.87 to 801.93 mg 100 g–1 DM, i.e. >50% of total FA), oleic acid (C18:1 c 9; 181.32 to 346.79 mg 100 g–1 DM, i.e. 13.2 to 24.6% of total FA) and α-linolenic acid (C18:3 c 9c 12c 15; 42.01 to 97.72 mg 100 g–1 DM, i.e. 3.4 to 7.1% of total FA) were the most abundant unsaturated FA. Palmitic acid (C16:0; 189.86 to 281.07 mg 100 g–1 DM, i.e. 15.4 to 17.8% of total FA) and stearic acid (C18:0; 24.35 to 52.75 mg 100 g–1 DM, i.e. 2.0 to 4.0% of total FA) were the major saturated ones. The sum of all other FA did not exceed 3.0% of TFA. The favourable FA profile of the studied vetch seeds makes them interesting cheap diet components to be used in the nutrition of ruminants and non-ruminants reared in the dryland agricultural regions of Mediterranean countries.
Introduction
The genus Vicia L. belongs to the Leguminosae (Fabaceae) family and comprises about 190 species, mainly distributed in temperate areas of both hemispheres. Mediterranean and Irano-Turanian regions represent primary vocation areas for the growth of these plants (van de Wouw et al., Citation2001). Positive agronomic attributes of vetches may be ascribed to their high fodder quality (Larbi et al., Citation2010a) and their ability to preserve/improve soil fertility (Rejili et al., Citation2012).
In Tunisia, three annual species, namely Vicia sativa L., Vicia villosa Roth. and Vicia narbonensis L., are largely cultivated in different bioclimatic areas (semi-arid for V. sativa, sub-humid for V. villosa, and from sub-humid to arid for V. narbonensis), particularly in the North of the country and mainly in association with oats (Avena sativa L.), for food and fodder production (Hassen and Zoghlami, Citation2004; Haffani et al., Citation2013).
Vetch seeds are considered valuable sources of protein to be used in animal nutrition (Larbi et al., Citation2010b). Some studies conducted on the potential nutritional value of vetch seeds from several species and cultivars grown in Tunisia showed that their high protein content makes them a cheap natural valid alternative to the more expensive soybean and its derivatives (Selmi et al., Citation2010). The widespread use of vetch seeds makes them also noteworthy sources of lipids for the rations of ruminants and non-ruminants (Kökten et al., Citation2010). The importance of fatty acid (FA) analysis in plant seeds relies in the possibility to select taxa characterised by a favourable FA profile (e.g., high levels of beneficial unsaturated FA) (Ryan et al., Citation2007; Kuhnt et al., Citation2012) which may lead to animal derived food products of enhanced fat quality, and to provide characteristic phenotypic information for the chemotaxonomic characterisation and the phylogenetic relationships existing at different taxonomic levels (Bağci and Şahin, Citation2004; Koçak et al., Citation2011).
The aim of this study was to determine the FA composition of the seeds of selected varieties, subspecies and accessions of three Vicia L. species grown in the region of Mateur (North Tunisia) as, despite their extensive use and value as animal feed in the dryland agricultural regions of Mediterranean countries, no such data are currently available.
Materials and methods
Vicia seeds
The biological material consisted of fully matured non heat-treated whole seed samples from three Vicia L. species: i) V. sativa L. (common vetch, section Vicia), represented by three Tunisian varieties (commune, Languedoc, Mghila) and one subspecies (amphicarpa (Dorthes) Asch.); ii) V. villosa Roth (hairy vetch, section Cracca), represented by a Tunisian variety (Sejenane) and two accessions (2565 and 3615) introduced from and provided by the International Center for Agricultural Research in the Dry Areas (ICAR-DA) in the frame of a germoplasm exchange; and iii) V. narbonensis L. (narbon vetch, section Narbonensis). All the seeds were collected in June 2012 from certified material grown in the region of Mateur (North Tunisia) and stocked in cleaned form in the gene bank of the Tunisian National Institute of Agronomy Research (INRAT, Tunisia).
Chemical analysis
The samples were ground with a cutting mill (MLI 204; Bühler AG, Uzwil, Switzerland) and analysed for their dry matter (DM) content according to the AOAC Official Method 930.15 (AOAC, Citation2000).
The seed FA composition was assessed using a combined direct transesterification and solid-phase extraction method as described by Alves et al. (Citation2008). Fatty acid methyl esters were separated and quantified by a high resolution gas chromatograph (Shimadzu GC 2010 Plus; Shimadzu, Kyoto, Japan) equipped with a flame-ionisation detector, and a CP-Sil 88 capillary column (100 m×0.25 mm ID, 0.20 µm film thickness; Varian Inc., Palo Alto, CA, USA). Injections were made in on-column mode and the injection volume was 0.5 µL. The temperatures of the injector and the flame-ionisation detector were maintained at 250 and 280°C, respectively. The column temperature was held at 45°C for 5 min, then raised 20°C min–1 up to 195°C and maintained for 65 min. Peaks were identified by comparing retention times to pure standards (Restek Corporation, Bellefonte, PA, USA) and by comparison with published chromatograms (Alves et al., Citation2008). Quantification was assessed by using heptadecanoic acid (C17:0) as internal standard. The results are expressed in absolute values as mg 100 g–1 DM and as percentages of total detected FA. All analytical determinations were performed in triplicate.
Statistical analysis
The statistical analysis was performed using IBM SPSS Statistics v.20 for Windows (SPSS Inc., Chicago, IL, USA). Data were subjected to one-way analysis of variance according to the following model:
Xij=µ+αi+εij
where: Xij is the observation; µ is the overall mean; αi is the effect of variety/subspecies/accession; and εij is the residual error. The Kolmogorov-Smirnov test was used to check dependent variables for normality. Pairwise multiple comparisons (Tukey’s test) were performed to test the difference between each pair of means. Significance was declared at P≤0.05.
Results and discussion
Differences among the analysed vetch seeds
The DM content and the FA composition of the seeds are reported in . The considered taxa showed a very similar DM content (P>0.05). The FA composition of the seeds ranged from lauric (C12:0) to lignoceric (C24:0) acids. Linoleic (C18:2 c 9c 12), oleic (C18:1 c 9) and palmitic (C16:0) acids were the most abundant ones. Unsaturated fatty acids (UFA) largely predominated over saturated fatty acids (SFA). The UFA/SFA ratio varied from 3.34 to 4.44 and was significantly different among the considered seeds (P≤0.001). The concentration of total polyunsaturated fatty acids (PUFA) was from 2.2- to 4.6-fold higher than that of total monounsaturated fatty acids (MUFA).
Polyunsaturated fatty acids
Linoleic acid was predominant, comprising more than 50% of total fatty acids (TFA) in all the samples. V. sativa subsp. amphicarpa and V. villosa accessions 2565 and 3615 showed higher concentrations of linoleic acid compared to V. narbonensis (P≤0.01), while intermediate values were detected for the other seeds.
Alpha-linolenic acid (C18:3 c 9c 12c 15) was also well represented, being the third most abundant UFA, after linoleic and oleic acids, in all the samples here analysed. The concentration of α-linolenic acid was comparable among the studied seeds, with the exception of V. narbonensis which showed approximately half values as much as all the other vetches.
Besides linoleic and α-linolenic acids, other detected PUFA were γ-linolenic (C18:3 c 6c 9c 12), eicosadienoic (C20:2 c 11c 14) and arachidonic (C20:4 c 5c 8c 11c 14) acids. All of them were detected only in traces. Stearidonic acid (C18:4 c 6c 9c 12c 15), a promising precursor of the endogenous synthesis of long-chain n3 FA in both animals and humans (Kuhnt et al., Citation2012), was not detected. In the current study, γ-linolenic acid was detected in all the seeds with the exception of V. narbonensis. The concentration of y-linolenic acid in V. sativa subsp. amphicarpa was significantly higher if compared to V. sativa var. commune and var. Mghila (P≤0.01); the other seeds showed intermediate values. Eicosadienoic acid was detected in all the analysed seeds and its concentration significantly varied among the considered taxa (P≤0.001). The highest amount was found in V. sativa var. Mghila, being different from the concentrations recorded in V. villosa var. Sejenane, acc. 2565 and acc. 3615, and V. sativa var. commune; the latter showed the lowest absolute concentration. Arachidonic acid was detected only in V. sativa subsp. amphicarpa and V. villosa var. Sejenane with relatively low and comparable concentrations.
No significant differences among the considered seeds were found in the n6/n3 PUFA ratio, with the exception of V. narbonensis which showed almost doubled values than the other seeds (P≤0.001). The n6/n3 FA ratio is commonly used to assess the nutritional value of lipids for human consumption. A strong imbalance towards high dietary intakes of n6 FA at the expense of n3 FA is positively correlated with a number of widespread human diseases. An optimal n6/n3 FA ratio should vary between 1:1 and 4:1, but Western diets may reach ranges of 10:1 to 20:1 (Simopoulos, Citation2011). None of the studied vetch seeds fell within the above-mentioned optimum recommended values.
Monounsaturated fatty acids
Compared to all other detected FA, oleic acid showed the greatest differences among the studied seeds. It ranked second after linoleic acid in the seeds of V. narbonensis and V. villosa accessions 2565 and 3615 the latter showing the highest absolute concentration. The seeds of V. sativa var. commune, var. Languedoc, subsp. amphicarpa and V. villosa var. Sejenane showed significantly lower concentrations of oleic acid if compared to V. narbonensis and V. villosa accessions 2565 and 3615 (P≤0.001). Moreover, V. villosa acc. 2565 and V. sativa var. Mghila showed significantly lower values than V. villosa acc. 3615. No significant differences were instead observed in the concentration of oleic acid between V. narbonensis and V. villosa acc. 2565, or between the latter and V. sativa var. Mghila. The oleic/linoleic FA ratio was always less than one, ranging from 0.23 to 0.47.
Except for oleic acid, all other detected MUFA [trans-3-hexadecenoic acid (C16:1 t 3), palmitoleic acid (C16:1 c 9), cis-vaccenic acid (C18:1 c 11) and eicosenoic acid (C20:1 c 11)] were present only in traces in the seeds. Their sum accounted for approximately 1% of TFA. Even if at low levels, they were detected in all the analysed samples. V. villosa acc. 3615 was significantly richer in trans-3-hexadecenoic acid than the other taxa (P≤0.001). V. villosa acc. 3615 showed significantly higher levels of cis-vaccenic acid with respect to the other seeds (P≤0.001). The lowest absolute concentration of cis-vaccenic acid was observed in the seeds of V. sativa subsp. amphicarpa, being significantly different from those recorded for V. villosa accessions 2565 and 3615 and V. sativa var. Mghila. Palmitoleic acid did not show any significant difference among the considered seeds. Regarding eicosenoic acid (a n9 very long chain MUFA), the seeds of V. sativa subsp. amphicarpa, V. villosa var. Sejenane and V. narbonensis showed very similar concentrations, which were significantly higher if compared to those of V. sativa var. commune. The other seeds showed intermediate amounts.
Erucic (C22:1 c 13) and nervonic (C24:1 c 15) acids were not detected in the seeds analysed in this study.
Saturated fatty acids
Considering all detected FA, palmitic acid ranked second after linoleic acid (in V. sativa var. commune, var. Languedoc, var. Mghila, subsp. amphicarpa and V. villosa var. Sejenane) or third after linoleic and oleic acids (in V. narbonensis and V. villosa accessions 2565 and 3615). The concentration of palmitic acid in V. villosa acc. 3615 significantly differed from that of all other seeds (P≤0.001), with the exception of V. villosa acc. 2565. The latter showed a concentration of palmitic acid which significantly differed only from that recorded in V. narbonensis.
The second most abundant SFA was stearic acid (C18:0) in all the seeds. V. sativa var. Mghila and V. villosa var. Sejenane showed significantly higher values of stearic acid if compared to V. narbonensis and V. villosa accessions 2565 and 3615 (P≤0.001). V. narbonensis showed the lowest absolute concentration, being significantly different from all the other seeds except for V. villosa acc. 3615.
The sum of all other detected SFA [lauric (C12:0), myristic (C14:0), arachidic (C20:0), behenic (C22:0) and lignoceric (C24:0) acids] did not exceed 23.29 mg 100 g–1 DM, that is 1.74% of TFA. Low molecular weight SFA, such as lauric and myristic acids, were found in all the samples. Odd-chain SFA [pentadecanoic (C15:0), heptadecanoic (C17:0) and nonadecanoic (C19:0) acids] were not detected in the current study. Among the considered seeds, no significant differences were observed in the concentration of lauric acid. On the contrary, myristic acid varied significantly: V. sativa var. Mghila showed the absolute highest concentration being significantly different (P≤0.05) from the concentrations recorded in V. villosa acc. 2565 and V. narbonensis. The other vetch seeds showed intermediate amounts.
Long-chain SFA levels significantly differed among the considered vetches. The concentration of arachidic acid was significantly higher (P≤0.001) in the seeds of V. sativa var. commune, var. Languedoc, var. Mghila, subsp. amphicarpa and V. villosa var. Sejenane if compared to V. narbonensis and V. villosa accessions 2565 and 3615. The highest and lowest absolute concentrations of behenic acid were observed in V. sativa subsp. amphicarpa and V. narbonensis. Lignoceric acid was not detected in the seeds of V. narbonensis and V. villosa accessions 2565 and 3615. The other vetch seeds showed significant differences (P≤0.01) in the concentration of lignoceric acid. The highest value was detected in V. villosa var. Sejenane, being twice as much as that recorded in V. sativa var. commune. The latter showed the absolute lowest concentration.
Compared to the other taxa, the seeds of V. narbonensis showed a significantly lower total SFA concentration (P≤0.01).
It is known that several factors, such as genetics, geographical location, climatic settings, growing conditions and post-harvest treatments, may affect the content of FA in the seed oils of many plants (Johansson et al., Citation2000; Khan et al., Citation2012). Environmental-based factors are likely not to be significant contributors of the observed variations in seed FA among the analysed Vicia taxa, as all the seeds were collected in a short period of time from the same geographical region and grew under similar climatic conditions and soil features. Therefore we conclude that genetic predisposition had a major impact on the observed variations. V. narbonensis provided the most considerable differences among the studied taxa, despite the lower taxonomic distance (based on morphological, cytological, biochemical, and molecular approaches) existing between V. sativa and V. narbonensis (both belonging to subgenus Vicia) if compared to those existing between V. sativa or V. narbonensis and V. villosa (the latter belonging to subgenus Cracca) (Mirali et al., Citation2007; Leht, Citation2009; Schaefer et al., Citation2012). Such hypothesis seems to be also confirmed by the results obtained in other studies where vetch seeds were collected in restricted geographical areas (Kokten et al., Citation2010; Emre et al., Citation2011).
Comparison with the literature data
The DM content of the analysed seeds was comparable to previously reported literature data (Yu et al., Citation2001; Seifdavati et al., Citation2013).
A comparison among the mean FA percentages obtained in this study for V. sativa and V. narbonensis with data found by other authors is presented in . To the best of our knowledge, for V. villosa no literature data of the seed FA profile is currently available.
Higher UFA than SFA, as well as higher PUFA than MUFA levels, were reported in the seeds of various wild and cultivated legumes in different ecological and geographical areas (Grela and Günter, Citation1995; Maestri et al., Citation2002; Bağci et al., Citation2004; Bağci and Şahin, Citation2004; Bağci, Citation2006; Yoshida et al., Citation2007; Pastor-Cavada et al., Citation2009a, Citation2009b; Kökten et al., Citation2010; Koçak et al., 2011).
The percentages of total UFA were comparable to those previously reported for other species of the genus Vicia (71.0 to 92.2%) (Bağci et al., Citation2004; Pastor-Cavada et al., Citation2009b; Kökten et al., Citation2010; Emre et al., Citation2011), including the species studied here. The obtained percentages were also comparable to those of the seeds of other related genera of the tribe Fabeae, such as Lathyrus L. (56.1 to 86.7%) (Bağci et al., Citation2004; Bağci and Şahin, 2004; Pastor-Cavada et al., Citation2009a; Emre et al., Citation2010), Lens Mill. (73.7 to 82.5%) (Ryan et al., Citation2007; Pastor-Cavada et al., Citation2009a) and Pisum L. (75.9 to 85.3%) (Ryan et al., Citation2007; Yoshida et al., Citation2007; Pastor-Cavada et al., Citation2009a; Renna et al., Citation2012), which are used as a protein source in animal and human nutrition. The seeds of some vetches grown in the Sivas region of Turkey (namely V. cracca, V. hyrcanica, V. galilaea and V. faba) were however reported to contain <60% of total UFA (Akpinar et al., Citation2001).
The obtained percentages of total SFA were similar to those reported by Kökten et al. (Citation2010) for six vetch species (18.0 to 22.4%), but slightly higher if compared to the 10-20% total SFA levels generally found by Bağci et al. (Citation2004) for legume seeds.
Linoleic-oleic, linoleic-palmitic and linoleicoleic-palmitic types FA patterns are known to be typical of many leguminous genera (Bağci et al., Citation2004). This was also confirmed by the preponderance of these three fatty acids in the analysed Tunisian vetch seeds.
Polyunsaturated fatty acids
In the current study, the observed variations in the linoleic acid percentages among vetch seeds were less pronounced (50.75 to 57.53% of TFA) if compared to those reported in other published works. Pastor-Cavada et al. (Citation2009b) and Bağci et al. (Citation2004) reported more than double levels of linoleic acid (28.7 to 66.3% of TFA and 20 to 50% of TFA, respectively) among the vetch species considered in their respective studies. On a whole, linoleic acid was usually found to be the most abundant FA in vetches (Bağci et al., Citation2004; Yoshida et al., Citation2008; Pastor-Cavada et al., Citation2009b; Kökten et al., Citation2010), with few exceptions reported (Akpinar et al., Citation2001; Bağci et al., Citation2004; Pastor-Cavada et al., Citation2009b). High levels of linoleic acid are also known to be typical of the seeds of many other legumes (Maestri et al., Citation2002; Bağci et al., Citation2004; Yoshida et al., Citation2007; Pastor-Cavada et al., Citation2009a; Emre et al., Citation2010; Koçak et al., Citation2011).
Alpha-linolenic acid was found to be one of the most variable FA components in legume seeds (Bağci et al., Citation2004). It was reported as the major FA in V. michauxii var. stenophylla, but more usually as the third most abundant UFA (after linoleic and oleic acids) in other vetch species (Bağci et al., Citation2004), as also occurred in the current study. Many vetches were reported to contain less than 15% α-linolenic acid in their seeds (Akpinar et al., Citation2001; Bağci et al., Citation2004; Pastor-Cavada et al., Citation2009b). Exceptions regarded few species or varieties such as V. articulata (16.6% of TFA) and V. pubescens (16.6%) (Pastor-Cavada et al., Citation2009b), V. ervilia (19.7%) and V. hybrida (22.0%) (Kökten et al., Citation2010) and particularly V. michauxii var. stenophylla (39.1%) (Bağci et al., Citation2004). As found in the analysed Tunisian V. narbonensis seeds, quite low α-linolenic acid levels (3-4% of TFA) in such species were also previously obtained by other authors (Pastor-Cavada et al., Citation2009b; Kökten et al., Citation2010).
The absence of stearidonic acid in the analysed vetch seeds confirms previously published data for V. sativa and V. narbonensis oilseeds. On the contrary γ-linolenic acid, which is known to possess a therapeutic value (being able to modulate inflammatory responses) (Kapoor and Huang, Citation2006), was not reported in vetch seeds in previously published papers, but it was found in traces in V. sativa and V. villosa oilseeds in the current study.
Considering the vetch seeds studied by Bağci et al. (Citation2004), eicosadienoic acid was detected only in one out of six species analysed, with a percentage (0.1% of TFA) comparable to those obtained in our trial. Conversely, Akpinar et al. (Citation2001) did not detect eicosadienoic acid in the seeds of V. hybrida, but they found a large variation in the levels of this FA (0.38 to 10.9% of TFA) among the remaining seven studied vetch species. These authors reported 9.25% eicosadienoic acid in the seeds of V. sativa, a value notably higher if compared to the range values (0.06 to 0.13% of TFA) found in our study. The same authors also reported notable amounts of arachidonic acid (1.23 to 6.83% of TFA) in the seeds of all examined species, which contrasts with the relatively low levels of this FA found in just two Tunisian vetch seeds in the current trial.
Monounsaturated fatty acids
Oleic acid was found to be the most abundant FA in the seeds of V. cassubica, V. cracca, V. hyrcanica, V. peregrina, V. hybrida, V. sativa, V. galilaea and V. faba by Akpinar et al. (Citation2001) and in the seeds of V. articulata by Pastor-Cavada et al. (Citation2009b). However, the oleic/linoleic FA ratio was usually reported to be less than one in the seeds of many species of the genus Vicia (Pastor-Cavada et al., Citation2009b) or other genera of the Leguminosae family (Maestri et al., Citation2002). As obtained in the current study, high levels of oleic acid in V. narbonensis seeds were already detected in different Mediterranean regions (Bağci et al., Citation2004; Pastor-Cavada et al., Citation2009b; Kökten et al., Citation2010; Emre et al., Citation2011).
The other monoenoic FA detected in the current study were either not reported (C16:1 t 3), found in traces (C16:1 c 9 and C20:1 c 11) or only in small amounts (C18:1 c 11) in the seeds of legume species, including those belonging to the genus Vicia (Maestri et al., Citation2002; Bağci et al., Citation2004; Bağci, Citation2006; Pastor-Cavada et al., Citation2009a, Citation2009b; Kökten et al., Citation2010; Koçak et al., Citation2011). The presence of trans-3-hexadecenoic acid was previously found to occur in the seeds of some Asteraceae (Hopkins and Chisholm, Citation1964; Morris et al., Citation1968) and, in general, in photosynthetic systems (Harwood and James, Citation1975). As occurred in our study, cis-vaccenic acid was usually found at higher levels if compared to palmitoleic and eicosenoic acids in different legume seeds (Bağci et al., Citation2004; Bağci, Citation2006).
The occurrence of erucic acid in vetch seeds was previously reported by Akpinar et al. (Citation2001), who found percentages varying from 0.23 (in V. hyrcanica) to 3.01% of TFA (in V. hybrida), with V. sativa presenting levels slightly less than 1% of TFA. Bağci et al. (Citation2004) revealed the occurrence of low erucic acid levels in some legumes, but not in vetch seeds. In accordance with the latter authors, erucic acid was not detected in the Tunisian vetch seeds here analysed. Such a result seems to be of quite importance as erucic acid was reported to exert negative effects on animal and human metabolism, so that the government regulation of the European Union limits its levels for human consumption to a maximum of 5% (Kuhnt et al., Citation2012). Nervonic acid, another n9 very long chain MUFA known to derive from erucic acid, was never reported as lipid constituent in vetch seeds in previously published works, a result which is also confirmed in our study.
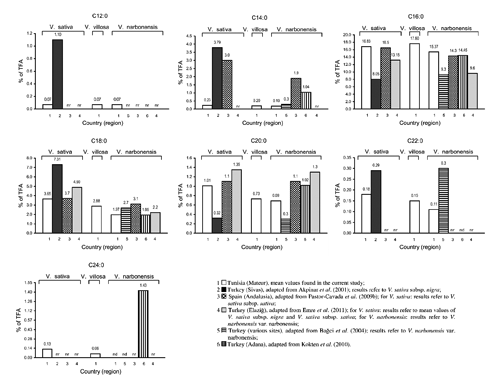
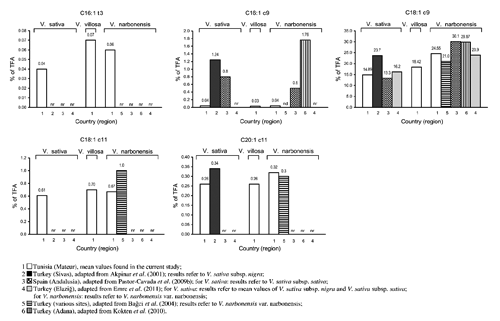
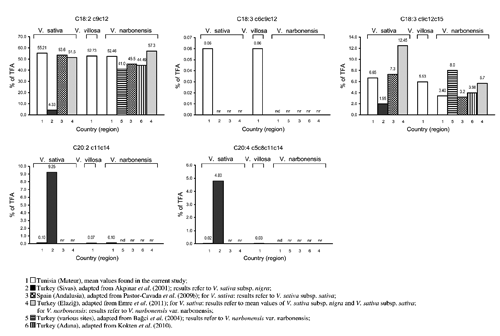
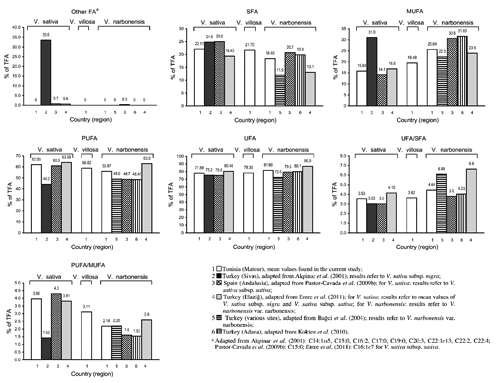
Table 1. Dry matter content and fatty acid composition of the seeds of selected Vicia L. taxa grown in Tunisia.
Saturated fatty acids
Palmitic acid is a steady lipid constituent in the seeds of various genera of the Leguminosae family (Bağci et al., Citation2004; Koçak et al., Citation2011). Confirming this, the range of palmitic acid variation among the seeds analysed in the current study was also relatively low (15.37 to 17.79% of TFA).
As occurred in the considered Tunisian vetch seeds, various other vetches were previously found to contain stearic acid as second most abundant SFA in their seeds (Akpinar et al., Citation2001; Bağci et al., Citation2004; Pastor-Cavada et al., Citation2009b; Emre et al., Citation2011). The majority of the species of the genus Vicia were reported to contain less than 6.0% stearic acid, with some exceptions such as V. pubescens (7.5% of TFA), V. cracca (13.2%), V. hyrcanica (19.4%), V. peregrina (7.26%), V. hybrida (9.13%), V. sativa (7.31%), V. galilaea (15.94%) and V. faba (9.03%) (Akpinar et al., Citation2001; Pastor-Cavada et al., Citation2009a). The percentages of stearic acid obtained in our study were also similar to those previously reported for the seeds of other leguminous genera which can be used in animal and human nutrition, such as Hedysarum, Lathyrus, Gonocytisus, Lupinus, Trigonella, Onobrychis, Lens, Pisum and Astragalus (Bağci et al., Citation2004; Bağci, Citation2006; Pastor-Cavada et al., Citation2009a; Renna et al., Citation2012).
Low molecular weight SFA, such as lauric and myristic acids, were found in all the samples analysed, as previously reported by Akpinar et al. (Citation2001). The presence of odd-chain FA was noticed in some vetch seeds in other trials (Akpinar et al., Citation2001; Pastor-Cavada et al., Citation2009b). Lauric, myristic and pentadecanoic acids were not usually found or found only in traces in the seeds of other leguminous genera (Bağci et al., Citation2004).
Long-chain SFA (arachidic, behenic and lignoceric acids) were not usually found or found at low levels (<1.5% of TFA) in vetch seeds (Bağci et al., Citation2004; Pastor-Cavada et al., Citation2009b; Kökten et al., Citation2010), with only few species (mainly V. cracca, V. peregrina, V. hybrida and V. galilaea) showing more than double amounts (Akpinar et al., Citation2001). Such findings are interesting from a nutritional point of view as oils with high levels of long-chain SFA were reported to be difficult to digest in both humans and animals (Akpinar et al., Citation2001).
On a whole, Figures show that great differences exist among the studies regarding both the number of detected FA and the relative percentage of each FA relative to the TFA content. Such discrepancies may be partly explained by the different levels of accuracy in FA analysis applied in the studies. Variations in the ecological and geographical zones where the seeds were collected may also have exerted a key role as it is known, as above mentioned, that the environment can significantly affect the synthesis of FA in plants (Akpinar et al., Citation2001; Mao et al., Citation2012).
Conclusions
In the studied vetch seeds the major FA ranked in the following order: C18:2 c 9c 12 > C16:0 > C18:1 c 9 [or C18:1 c 9 > C16:0, depending on the considered subspecies/variety/accession] > C18:3 c 9c 12c 15 > C18:0, which is consistent with data reported in the available literature for leguminous seeds. From a qualitative perspective, oleic, stearic, linoleic and α-linolenic acids (among individual FA) and total MUFA (among FA groups), were the most useful parameters for highlighting interspecies variability among the seeds. Arachidic acid, expressed as percentage of total detected FA, seems to be helpful to show up intraspecies variability for the three varieties/accessions of V. villosa. Characteristic phenotypic information was provided by i) arachidonic acid, which was only detected in the seeds of V. sativa subsp. amphicarpa and V. villosa var. Sejenane, and ii) lignoceric acid, which was not detected in the seeds of V. villosa acc. 2565 and acc. 3615. The seeds of V. narbonensis drew away from those of the other studied vetches, essentially due to i) their high levels of oleic acid, total MUFA, UFA/SFA and n6/n3 PUFA ratios, ii) their low levels of palmitic acid and total SFA, and iii) the absence of γ-linolenic acid.
The analysed vetch seeds are a valuable source of UFA (both mono- and polyunsaturated ones), whose levels are comparable to those of other edible seeds. Such a favourable FA profile and the high protein levels make these seeds interesting cheap diet components for animal nutrition. Due to the higher concentration of the sum of linoleic, a-linolenic and γ-linolenic acids (about 890 mg 100g–1 DM), the seeds of V. sativa subsp. amphicarpa and V. villosa accession 3615 may be the most effective, among the studied ones, in improving the quality of the lipid fraction of ruminantderived food products (raise in the content of beneficial FA such as vaccenic and conjugated linoleic acids).
Acknowledgments
This research was supported by Italian Ministry of Education, University and Research (MIUR, Italy) grants (ex 60%) and by Laboratory of Economy and Food Technology (INAT, Tunisia). The authors gratefully acknowledge their colleague Vanda Malfatto for her careful technical assistance.
References
- AkpinarN. AkpinarM.A. TürkoğluŞ., 2001. Total lipid content and fatty acid composition of the seeds of some Vicia L. species. Food Chem. 74:449-453.
- AlvesS.P. CabritaA.R.J. FonsecaA.J.M. BessaR.J.B., 2008. Improved method for fatty acid analysis in herbage based on direct transesterification followed by solid-phase extraction. J. Chromatogr. A 1209:212-219.
- AOAC, 2000. Official methods of analysis. 17 th ed., Association of Official Analytical Chemists, Gaithersburg, MD, USA.
- BağciE., 2006. Fatty acid composition of some Astragalus species from Turkey. Chem. Nat. Compd. 42:645-648.
- BağciE. BruehlL. ÖzçelikH. AitzetmullerK. VuralM. §ahinA., 2004. A study of the fatty acid and tocochromanol patterns of some Fabaceae (Leguminosae) plants from Turkey I. Grasas Aceites 55:378-384.
- BağciE. §ahinA., 2004. Fatty acid patterns of the seed oils of some Lathyrus species L. (Papilionideae) from Turkey, a chemotaxonomic approach. Pak. J. Bot. 36:403-413.
- Emreİ. ŞahinA. ÔkkeşYılmaz, Ö. GençH., 2010. Compositions of seed fatty acids in some Lathyrus taxa from Turkey. Acta Bot. Gallica 157:241-246.
- Emreİ. ŞahinA. YılmazÖ. GençH. BahşiM., 2011. Compositions of seed fatty acids in some Vicia L. taxa from Turkey. Acta Bot. Gallica 158:493-498.
- GrelaE.R. GünterK.D., 1995. Fatty acid composition and tocopherol content of some legume seeds. Anim. Feed Sci. Tech. 52:325-331.
- HaffaniS. MezniM. SlamaI. KsontiniM. ChaïbiW., 2013. Plant growth, water relations and proline content of three vetch species under water-limited conditions. Grass Forage Sci. ( In press).
- HarwoodJ.L. JamesA.T., 1975. Metabolism of trans-3-hexadecenoic acid in broad bean. Eur. J. Biochem. 50:325-334.
- HassenH. ZoghlamiA., 2004. Ressources génétiques des espèces spontanées de légumineuses fourragères et pastorales en Tunisie. In: FerchichiA. FerchichiA. ( eds.) Réhabilitation des pâturages et des parcours en milieux méditerranéens. CIHEAM Publ., Zaragoza, Spain, pp 375-377.
- HopkinsC.Y. ChisholmM.J., 1964. Occurrence of trans-3-hexadecenoic acid in a seed oil. Can. J. Chem. 42:2224-2227.
- JohanssonA. LaineT LinnaM.-M. KallioK., 2000. Variability in oil content and fatty acid composition in wild northern currants. Eur. Food Res. Technol. 211:277-283.
- KapoorR. HuangY.S., 2006. Gamma linolenic acid: an antiinflammatory omega-6 fatty acid. Curr. Pharm. Biotechno. 7:531-534.
- Khan R. Srivastava R. Khan M.A. Alam P. Abdin M.Z. Mahmooduz zafar, 2012. Variation in oil content and fatty acid composition of the seed oil of Acacia species collected from the northwest zone of India. J. Sci. Food Agr. 92:2310-2315.
- KoçakA. KöktenK. BağciE. AkçuraM. HaytaS. BakogluA. KilicO., 2011. Chemical analyses of the seeds of some forage legumes from Turkey. A chemotaxonomic approach. Grasas Aceites 62:383-388.
- KöktenK. KoçakA. BağciE. AkçuraM. ÇelikS., 2010. Tannin, protein contents and fatty acid compositions of the seeds of several Vicia L. species from Turkey. Grasas Aceites 61:404-408.
- KuhntK. DegenC. JaudszusA. JahreisG., 2012. Searching for health beneficial n-3 and n-6 fatty acids in plant seeds. Eur. J. Lipid Sci. Tech. 114:153-160.
- LarbiA. HassanS. KattashG. Abd El-MoneimA.M. JammalB. NabilaH. NakkulH., 2010a. Annual feed legume yield and quality in dryland environments in north-west Syria: 1. Herbage yield and quality. Anim. Feed Sci. Tech. 160:81-89.
- LarbiA. HassanS. KattashG. Abd El-MoneimA.M. JammalB. NabilaH. NakkoulH., 2010b. Annual feed legume yield and quality in dryland environments in north-west Syria: 2. Grain and straw yield and straw quality. Anim. Feed Sci. Tech. 160:90-97.
- LehtM., 2009. Phylogenetics of Vicia (Fabaceae) based on morphological data. Feddes Repert. 120:379-393.
- MaestriD.M. FortunatoR.H. GuzmánC.A. TorresM.M. LamarqueA.L., 2002. Seed compositional studies of some species of Papilionoideae (Leguminosae) native to Argentina. J. Sci. Food Agr. 82:248-251.
- MaoZ. FuH. NanZ. WangJ. WanC., 2012. Fatty acid content of common vetch (Vicia sativa L.) in different regions of Northwest China. Biochem. Syst. Ecol. 44:347-351.
- MiraliN. El-KhouriS. RizqF., 2007. Genetic diversity and relationships in some Vicia species as determined by SDS-PAGE of seed proteins. Biol. Plantarum 51:660-666.
- MorrisL.J. MarshallM.O. HammondE.W., 1968. The trans-3-enoic acids of Aster alpinus and Arctium minus seed oils. Lipids 3:91-95.
- Pastor-CavadaE. JuanR. PastorJ.E. AlaizM. VioqueJ., 2009a. Chemical composition and nutritional characteristics of the seed oil of wild Lathyrus, Lens and Pisum species from Southern Spain. J. Am. Oil Chem. Soc. 86:329-335.
- Pastor-CavadaE. JuanR. PastorJ.E. AlaizM. VioqueJ., 2009b. Fatty acid distribution in the seed flour of wild Vicia species from Southern Spain. J. Am. Oil Chem. Soc. 86:977-983.
- RejiliM. MahdhiM. FterichA. DhaouiS. GuefrachiI. AbdeddayemR. MarsM., 2012. Symbiotic nitrogen fixation of wild legumes in Tunisia: soil fertility dynamics, field nodulation and nodules effectiveness. Agr. Ecosyst. Environ. 157:60-69.
- RennaM. CornaleP. LussianaC. MalfattoV. FortinaR. MimosiA. BattagliniL.M., 2012. Use of Pisum sativum (L.) as alternative protein resource in diets for dairy sheep: Effects on milk yield, gross composition and fatty acid profile. Small Ruminant Res. 102:142-150.
- RyanE. GalvinK. O’ConnorT.P. MaguireA.R. O’BrienN.M., 2007. Phytosterol, squalene, tocopherol content and fatty acid profile of selected seeds, grains, and legumes. Plant Food. Hum. Nutr. 62:85-91.
- SchaeferH. HechenleitnerP. Santos-GuerraA. Menezes de SequeiraM. PenningtonR.T. KenicerG. CarineM.A., 2012. Systematics, biogeography, and character evolution of the legume tribe Fabeae with special focus on the middle-Atlantic island lineages. BMC Evol. Biol. 12:250.
- SeifdavatiJ. TaghizadehA. JanmohammadiH. Abbas RafatS. AlijaniS., 2013. In situ ruminal degradability and in vitro intestinal digestibility of crude protein from some minor legume species. Afr. J. Biotechnol. 12:2293-2302.
- SelmiH. Gasmi-BoubakerA. Mosquera LosadaR. AbdouliH. RekikB. RouissiH. HassenH. Rigueiro-RodriguezA., 2010. Natural vetch economic alternative to soybean meal in Tunisia. In: PorquedduC. RíosS. ( eds.) The contributions of grasslands to the conservation of Mediterranean biodiversity. CIHEAM/CIBIO/FAO/SEEP Publ., Zaragoza, Spain, pp 63-66.
- SimopoulosA.P., 2011. Importance of the omega-6/omega-3 balance in health and disease: evolutionary aspects of diet. In: SimopoulosA.P. ( ed.) Healthy agriculture, healthy nutrition, healthy people. Karger, Basel, Switzerland, pp 10-21.
- van de WouwM. EnnekingD. RobertsonL.D. MaxtedN., 2001. Vetches (Vicia L.). In: MaxtedN. BennettS.J. ( eds.) Plant genetic resources of legumes in the Mediterranean. Kluwer Academic Publ., Dordrecht, The Netherlands, pp 134-158.
- YoshidaH. TomiyamaY. TanakaM. MizushinaY., 2007. Distribution of fatty acids in triacylglycerols and phospholipids from peas (Pisum sativum L.). J. Sci. Food Agr. 87:2709-2714.
- YoshidaH. TomiyamaY. YoshidaN. SaikiM. MizushinaY., 2008. Lipid classes, fatty acid distributions and triacylglycerol molecular species of broad beans (Vicia faba). J. Am. Oil Chem. Soc. 85:535-541.
- YuP. SpragueM. EganA.R. CastlemanG.H. LeuryB.J., 2001. Comparison of raw and roasted narbon beans (Vicia narbonensis) on performance and meat sensory attributes of lambs fed a roughage-based diet. Anim. Feed Sci. Tech. 92:1-16.