
Abstract
Survival is one of the most important traits in genetic improvement programmes in livestock. The objective of this study is to determine sex, inbreeding, and birth type effects as well as their interactions on birth survival in lambs of two breeds. Records of 6356 Polish Merino (PM) breed and 9143 Wielkopolska sheep (WS) were taken into the analysis. Relationships between survival and three binary variables (sex, inbreeding, type of birth) were estimated by the use of logistic regression. Interactions among these variables were also included in the model. Thus, the odds ratios and synergy coefficients were estimated. Lowest birth mortality was registered for inbred females from single birth. Differences in survival between the studied breeds were observed. Negligible single effects of sex, inbreeding and birth type for PM were estimated, whereas for WS these effects were significant (P<0.01). Opposite dependencies were obtained for interactions among these variables. In the case of PM, synergy between birth survival and joint effects of sex-inbreeding, sex-birth type, sex-inbreeding-birth type were highly significant, but for WS only inbreeding-birth type interaction was considerable. Hence, a generalisation of the obtained results seems to be difficult: they exhibit a very complex background of lamb survival.
Introduction
Survival can be perceived as one of the most important traits for both livestock and wild animal species for at least two main reasons. Firstly, this character plays a key role in small endangered populations. Over the last decades, sheep population has strongly decreased in a majority of countries in Europe, whereas the breed structure is relatively conserved (Barczak, Citation2012). This leads to a reduction of breed vs genetic group size with unfavourable accompanying effects (e.g. increase of inbreeding rate). Secondly, birth survival is important from the economic perspective. It should be noted that it is also determined by chromosomal abnormalities, and (semi-)lethal loci associated with diseases. Survival may be reduced by the sex chromosome chimerism and the freemartin syndrome. Dixon et al. (Citation2007) reported that individual potential offspring may be lost without a total loss of pregnancy in multiple pregnancies. Probability of these abnormalities also increases with the inbreeding level, as documented by many authors (e.g. Wooliams et al., Citation1983). Differences for early mortality in lambs have been registered among breeds (Maxa et al., Citation2009). Although this trait is genetically determined, heritability estimates in the literature are generally low (Hatcher et al., Citation2010). It implies a low efficiency of the genetic improvement programme for this trait. On the other hand, considerable influences of some factors (sex, birth type of lambs, etc.) on lamb survival have been reported in the literature (Piwczyński et al., Citation2010). Two sheep breeds were studied. Polish Merino (PM) is an old breed originated from West Europe in the 19th century. Average fecundity of ewes was 135%. It is perceived as a relatively highly genetically consolidated breed. Currently, PM contributes to about 20% (43.6 thousands individuals) of the total sheep population in Poland. Wielkopolska sheep (WS) was created in the years 1948-1976 by several steps crossing of old local (Swiniarka and Leszczynska) and international breeds (Merino and Romney Marsh, East Fresian sheep, Berrichonne du Cher). Average fecundity of this breed is higher (140%) compared to PM. However, its participation in current population is relatively small – approximately 0.2% (about 500 individuals). At the moment, WS is included into a national programme of conservation of genetic resources. It should be noted that both breeds play a considerable role in sheep production in the Wielkopolska region. Hence, sizes of both these populations are relatively large in the region.
By contrast to many continuous sheep performance characters, a survival is a binary. Hence, it is troublesome in statistical analysis, especially when relationships with other binary characters are studied. The classical approaches are usually based on contingency tables. However, in this case, a statistical inference is very limited. Recently, new methods have been developed to analyse this kind of data. One of them is the analysis of synergy factors (SFs). It is successfully implemented mainly for epidemiological data (Cortina-Borja et al., Citation2009) in human populations, when binary factors are studied. The method has a number of advantages. It is simple in calculations and allows assessment of binary interactions. To our knowledge, this approach has not been employed for livestock population.
The objective of this study is to test a new method to estimate the effect of sex, inbreeding, and birth type as well as their interactions on lambs’ birth survival of two very different (from a genetic perspective) sheep breeds.
Materials and methods
Data
The data were extracted from databases of the Regional Association of Sheep and Goat Breeders in Poznan (Poland). For each of the two breeds, a single record consisted of the following: animal identity, sire identity, dam identity, sex, flock code, birth type, lambing date, birth survival and inbreeding level. Routine performance system evaluation included birth body weight and consecutive measurements of body weight. Two local breeds were analysed: PM and WS.
After preliminary editing of the data, 6748 complete records for PM and 9450 complete records for WS were extracted. A majority of them originated from first lambing (94.19 and 96.75% for PM and WS, respectively), and only these animals were analysed. Finally, 6356 (for PM) and 9143 records (for WS) from 1995-2008 were taken into the analysis. The PM individuals were kept in 181 flocks, whereas the other breed in 158 flocks. In general, climatic and management conditions across the flocks were similar.
The average inbreeding levels of recorded individuals were 0.005 (for PM) and 0.012 (for WS). In the case of inbred animals the mean inbreeding coefficients were as follows: 0.031 (for PM) and 0.047 (for WS). The completeness of pedigrees measured by the Cassel coefficient in both populations studied gradually increased up to 95% (Barczak, Citation2012). As in Cerchiaro et al. (Citation2007) we categorised inbreeding in two classes, grouping into one class animals with inbreeding.
A preliminary analysis of the data showed a significant influence of flock on the survival in both breeds. Because it was impossible to introduce flock as a variable into the model, a uniform subset of stocks was taken from the data. It consisted of 40 flocks for PM (2017 records) and 43 flocks for WS (5619 records). The inclusion of a subsequent factor (e.g. flocks) into the analysis would have destabilised the data structure with some empty subclasses. Also from a biological perspective, the effects of flocks are usually perceived as only a disturbed factor. Hence, the analysis included also reduced data with homogenous birth survival across flocks. It should be stressed that negligible trends in survival were registered over the years (Barczak, Citation2012). Hence, the birth year was omitted in this analysis.
The material is divided according to the following three criteria: sex, inbreeding [two classes – animals with (F) vs without (NF) inbreeding], birth type [single (S) vs multiple (M) birth]. Description of lambing survival in the above mentioned subclasses for both populations studied is shown in . Since there is one empty subclass in the reduced set of data for PM, a value of 0.5 is added to each subclass following Breslow (Citation1981).
Methods
To establish the relationship between the analysed binary variables, sex, inbreeding and type of birth and their influence on survival, the logistic regression model and SF analysis related to it are used. Let θ(x) be the probability of survival, where x=(x1, x2, x3) denotes the vector of three binary analysed variables. Then, θ(x)/( 1-θ(x)) denotes the corresponding odds. In what follows the notion of odds ratio (OR) will also be used. The regression model has the following form:
By estimating and testing the significance of β coefficients, the influence of analysed variables as well as their interactions on lamb survival at birth can be established. In fact, the estimates of the β’s are defined by logarithms of SFs. The SF is a function of ORs. It assumes value 1 if the two traits act independently. When synergy is greater than 1 it indicates a positive interaction (synergy) and in the opposite case there is a negative interaction (antagonism).
Let θijk be the probability of survival, when x1, x2, x3 take the values i, j, k respectively, and as these are binary variables i, j, k € {0,1}. Then, it is easy to show that the estimates of the regression model coefficients have the form:
where
In practice, the ORs are calculated based on corresponding frequencies of lambs from different classes (). An extension of the results of Berrington de Gonzales and Cox (Citation2005) for a case of three variables gives the following formula for standard deviation:
where nl are numbers of lambs in different classes. As shown by Hosmer and Lemeshow (Citation2000) and Cortina-Borja et al. (Citation2009), a normal approximation may be used to the logarithm of SF divided by its standard deviation. In this way it is possible to test the hypotheses about the significance of regression coefficients as well as the hypotheses about the significance of ln(SF). This result may also be used to derive confidence intervals.
Results
The percentage of survived lambs within particular subclasses is listed in . In general, the differences in survival between the breeds analysed are confirmed. Lower birth mortality is registered for WS compared to PM. However, the survival percentages varied across sex, inbreeding levels, and type of birth. It should be noted that highest values of survival were obtained for inbred animals. As it has been already mentioned, the inbreeding levels in both populations were relatively low. The relations between percentages in different subclasses for complete and reduced set of data are similar, except the case of animals with inbreeding. On the other hand, for inbred individuals differences between both breeds were the smallest. This can indicate non-inbreeding effects on this trait under the homozygosity level in the populations studied. It seems that a positive impact of negligible inbreeding rate can be hypothesed. The optimistic results are probably influenced by gene pools of both populations.
Lowest survival (49%) is registered for multiple birth in PM, whereas for the other breed the value achieved 72%. As expected, survival of females is higher than males. For the eight combinations the smallest survival was registered for female-non-inbred-multiple birth groups for PM and male-non-inbred-multiple birth groups for WS. Hence, these groups were assumed as reference ones and the estimated values of odds and OR refer to them. Consequently, the analysed variables take the following values:
0, Dead female(PM) male(WS), inbred=0 multiple birth
1, Survived male(PM), female(WS), inbred>0, single birth
The odds and ORs for lambs of both breeds within factorial subclasses are shown in . By definition these values of odds and ORs are proportional. For both breeds, the coefficients are highest for female-inbred-multiple birth. They indicate positive joint effects of the variable classes on lamb survival. For a majority of variable combinations, the highest values of odds were estimated for WS. This indicates larger survival potential of this breed compared to PM. As said, WS originated from a long crossbreeding process. However, over the last two decades the WS breed was considerably reduced. Hence, the population has been covered by the national genetic resource programme. This can bring about an increase of the inbreeding rate since the number of sires and dams were reduced as well. Fortunately, the current inbreeding level in these populations seems to be safe. Positive inbreeding effect on survival likely resulted from accumulation of desirable genotypes.
Estimates of logistic regression coefficients and SF coefficients for inbreeding, sex, and birth type and their two and three step combination on survival within both breeds are listed in . For our considerations it is important to notice that in the case of both breeds the results are similar for nonreduced and reduced sets of data. Generally, these estimates considerably varied across breeds. Indeed in the case of PM with one exception, all model components are highly significant. Sex seems to have a non significant influence on survival. For WS the main effects are significant, whereas the interaction coefficients, again with one exception, are non significant. It shows that inbreeding and birth type significantly influence survival in both breeds, whereas the sex only influences survival in WS. Additionally, it should be noted, that the SFs, with one exception, are less than one, indicating antagonism of the analysed variables. In the analysis of SFs we should recall that for example SF110 will equal 1 if the increase of survival odds when changing from females to males is the same in a group with inbreeding and in a group without inbreeding for lambs from multiple birth. In our case this hypothesis is rejected for PM. Visualisations of ORs under all studied scenarios are given in Figures 1-3. The comparison of the first and third column in shows the significance of SF110 in the case of PM and its non significance in the case of WS. In other words, for PM from multiple birth, the sex acts differently in a group with inbreeding and in a group without inbreeding. Similarly, SF101 will be one if the increase of survival odds when changing from females to males and from multiple birth to single birth will be the same for non-inbred individuals. Again the comparison of the first and third column in shows the significance of SF101 in the case of PM and its non significance in the case of WS. Practically, it means, that for PM individuals without inbreeding, type of birth acts differently on survival for males and females.
illustrates the significance of SF011 which measures the increase of survival odds when changing from non-inbred to inbred and multiple birth to single birth for females in PM and males in WS. It should be noted that all these parameters are less than one proving the antagonism of analysed variables.
The significance of SF111 in the case of PM is illustrated for example in . For individuals from single birth, the influence of sex in inbred and non inbred groups is similar (columns 5 and 7) in contradiction to multiple birth (columns 1 and 3). In other words, in the case of multiple birth, in a group without inbreeding, males show higher survival than females; in a group with inbreeding the relation is the reverse. In the case of single birth, sex does not differentiate survival. In the case of the same comparisons in WS the tendencies are similar (see whole data set columns in ). Again it should be stressed, that the foregoing conclusions are valid for the whole and reduced set of data proving the non important influence of different flocks.
Discussion
The frequency of lamb birth survival is low in both population studied (63 and 75% for PM and WS, respectively). In the case of PM population (in other region of Poland) Piwczy ski et al. (Citation2010) reported lamb survival has varied over time (from 89.34% in 1999 to 73.66% in 2004 year). Whereas Gama et al. (Citation1991) estimated that survival from birth to weaning ranged from 73.7 to 80%. Higher survival level than in the present study was also estimated by Matos et al. (Citation2000). However, it should be recalled that highest mortality is registered in the first day after birth. This case is frequently classified as birth death. Generally, lamb birth survival varies across breeds and genetic groups.
This study established that the lamb sex, inbreeding and birth type affected survival. Furthermore, interactions among these variables were also registered. There were considerable differences between lamb survival from the two breeds. In general the obtained results correspond partially with some reports. The origins of these breeds are quite different. As previously mentioned, the PM breed was formed by very long term selection. It is one of the branches of Merino sheep kept in a number of countries around the world. For this breed, the estimates of regression coefficients (and in consequence ORs) are statistically non-significant. It should be recalled that the WS can be perceived as a crossbreeding product because it was formed by the use of many breed components. In fact, over the last four decades WS has been kept as purebred. However, from this point of view the genetic variability would be likely larger than that of PM population. On the other hand, the size of population WS is considerably decreased. Differences in risk of lamb death among breeds have been shown by many authors. Indeed Nguti et al. (Citation2003) indicated differences between two local sheep purebreds whereas they did not show the influence of heterosis effects on survival at consecutive points of life. Some authors (e.g. Gudex et al., Citation2005) observed lamb survival among genetic groups (lines, and half-, and full sibs) within breeds. It could suggest a large genetic variability, and in consequence high heritability of survival. Thus, an effective selection could be performed. Unfortunately, a number of reports explicitly indicate a low heritability of this trait (Olivier et al., Citation1998; Riggio et al., Citation2008). On the other hand, it must be noted that Cloete et al. (Citation2009) concluded that sustained genetic gain in lamb survival due to indirect selection is focused on some correlated traits which are connected with the dam’s ability to rear multiples. However, this does not apply to the genetic improvement programme in birth survival. So, low genetic variation has led to the suggestion that the improvement of birth lamb survival should be done by modification of management conditions. It is well known that survival is determined by a number of genetic and non-genetic factors. Besides the previously mentioned additive genetic variance, another genetic source of variability seems to be responsible for birth mortality. Cloete et al. (Citation2009) reported relatively high maternal heritability for this trait whereas the covariance between direct and maternal genetic effects is usually negative. It implies some barriers in the construction of efficient genetic improvement programmes. Prenatal and postnatal death may also be affected by congenital defects and chromosomal abnormalities. However, their scale is relatively small and from an economic perspective they have a marginal context. Some authors (Maxa et al., Citation2009) suggested considerable impacts of other effects such as: age of ewe, season of birth, lambing year, herds, sex of lamb, birth type, birth weight, lambing difficulty, parity, and inbreeding level. It is obvious that some of the factors are strongly correlated. For instance, the litter size is associated with lamb birth body weight. Thus, the number of analysed effects can be reduced.
In the present study, the logistic function approach is employed to detect the influence of three factors (inbreeding level, lamb sex and birth type) and their interactions on lamb birth survival. Unfortunately, from the perspective of the method used, the number of included factors (variables) has been limited, especially when two- and three step interactions between them were estimated. As shown in , the current structure of the data is considerably unbalanced. The number of inbred males from multiple pregnancies is relatively small (only 11 individuals for PM) compared to the number of inbred females from single birth (3988 animals for PM). As it already mentioned, the analysis included also reduced data with homogenous birth survival across flocks. These results obtained for both (whole and reduced) populations were similar. It seems that the approach can be recommended when many numbers of subclasses are incorporated into the analysis. Thus, a homogenous subset can be perceived as a representative sample of the population studied. On the other hand Hatcher et al. (Citation2009) estimated small negligible differences among Australian Merino flocks for birth survival. For the same reason, the year effects were also omitted in this study.
Significant influences of sex, inbreeding and birth type were observed for WS and PM (except the sex effect). However an impact of the individual’s sex on survival is well documented in literature. Many authors evidently showed that longevity of females is larger than of males (Molińska-Glura et al., Citation2008). The dependencies are strongly associated with birth vigor, and in consequence it affects a larger survival at birth. Higher female than male birth survival was also registered in the present study. Furthermore, this analysis indicates that better survival was registered for lambs from single pregnancies compared to twins. However, by contrast to some species sheep multiple pregnancies have a relatively negligible effect on survival.
There are many reports on inbreeding effects on traits in both livestock and wild animals. Although from the perspective of a genetic programme strategy the mating of relatives is perceived as undesirable, some authors reported a positive influence of inbreeding on survival. Lamberson et al. (Citation1982) observed higher survival for the progeny of inbred ewes. Positive inbreeding effects for this trait were also reported for two local sheep breeds by Boujenane and Chami (Citation1997). However, Wiener et al. (Citation1992) indicate a negative impact of increasing homozygosity on lamb survival. It can be noted that the estimates of inbreeding level have been relatively low in sheep populations (including both breeds studied). Moreover, some studies performed on small populations of wild animals as well as the history of formation of some breeds suggest both bad and good inbreeding. Some breeds have been created due to positive effects of the mating of relatives. As already mentioned, estimated significant SF effects (for PM) strongly exhibited the complex action of inbreeding, sex and birth type. Hence, further analysis of risk factors should be focused on analytical evaluation of their interaction, which must not correspond with single effects of the studied variables.
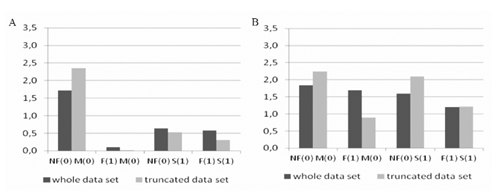
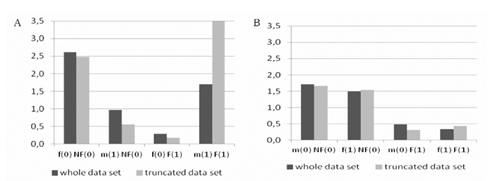
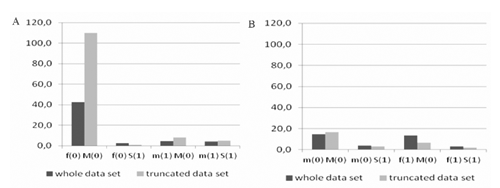
Table 1. The description of data structure used in the analysis.
Table 2. Percentage of survived lambs of both breeds.
Table 3. Odds and odds ratios.
Table 4. Estimates of regression coefficients and the corresponding synergy factors.
Conclusions
All the three studied factors (inbreeding level, lamb sex and birth type) and an interaction among them affected survival at birth. The joint effects can vary across different compositions of factors. It should be stressed that considerable differences in the estimated parameters were registered for both breeds. Therefore, a generalisation of the obtained results is seemingly difficult. It exhibits a very complex background of lamb survival dependent on specific conditions of particular populations.
References
- Barczak E., 2012. Individual and maternal inbreeding level and its effects on performance traits in sheep. PhD Diss., Poznan University, Poland.
- Berrington de GonzalezA. Cox D.R., 2005. Additive and multiplicative models for the joint effect of two risk factors. Biostatistics 6:1-9.
- Boujenane I. Chami A., 1997. Effects of inbreeding on reproduction, weights and survival of Sardi and Beni Guil sheep. J. Anim. Breed. Genet. 114:23-31.
- Breslow N.E., 1981. Odds ratio estimators when the data are sparse. Biometrika 68:73-84.
- Cerchiaro I. Cassandro M. Dal ZottoR. Camier P. Gallo L., 2007. A field study on fertlity and purity of sex-sorted cattle sperm. J. Dairy Sci. 90:2538-2542.
- Cloete S.W.P. Misztal I. Olivier J.J., 2009. Genetic parameters and trends for lamb survival and birth weight in a Merino flock divergently selected for multiple rearing ability. J. Anim. Sci. 87:2196-2208.
- Cortina-Borja M. Smith A.D. Combarros O. Lehmann D.J., 2009. The synergy factor: a statistic to measure interactions in complex diseases. Available from: www.bio-medcentral.com/content/pdf/1756-0500-2105.pdf
- Dixon A.B. Knights M. Winkler J.L. Marsh D.J. Pate J.L. Wilson M.E. Dailey R.A. Seidel G. Inskeep E.K., 2007. Patters of late embryonic and fetal mortality and association with several factors in sheep. J. Anim. Sci. 85:1274-1284.
- Gama L.T. Dickerson G.E. Young L.D. Leymaster K.A., 1991. Effects of bred, heterosis, age of dam, litter size, and birth weight on lamb mortality. J. Anim. Sci. 69:2727-2743.
- Gudex B.W. Hickford J.G.H. Frampton C.M., 2005. Sire line differences in lamb losses from starvation-exposure. Proc. New Zeal. Soc. An. 65:186-190.
- Hatcher S. Atkins K.D. Safari E., 2009. Phenotypic aspects of lamb survival in Australian Merino sheep. J. Anim. Sci. 87:2781-2790.
- Hatcher S. Atkins K.D. Safari E., 2010. Lamb survival in Australian merino sheep: a genetic analysis. J. Anim. Sci. 88:3198-3205.
- Hosmer D.W. Lemeshow S., 2000. Applied logistic regression. John Wiley & Sons, Hoboken, NJ, USA.
- Lamberson W.R. Thomas D.L. Rowe K.E., 1982. The effects of inbreeding in a flock of Hampshire sheep. J. Anim. Sci. 55:780-786.
- Matos C.A.P. Thomas D.L. Young L.D. Gianola D., 2000. Genetic analyses of lamb survival in Rambouillet and Finnsheep flocks by linear and threshold models. Anim. Sci. 71:227-234.
- Maxa J. Sharifi A.R. Pedersen J. Gualy M. Simianer H. Norberg E., 2009. Genetic parameters and factors influencing survival to twenty-four hours after birth in Danish meat sheep breeds. J. Anim. Sci. 87:1888-1895.
- Molńska-Glura M. Szwaczkowski T. Moliński K., 2008. Hazard rates for clustered population of David’s Deer (Elaphurus davidianus Milne Edwards 1866). Anim. Sci. Pap. Rep. 26:141-151.
- Nguti R. Janssen P. Rowlands G.J. Audho J.O. Baker R.L., 2003. Survival of Red Maasai, Dorper and crossbreed lambs in the subhumid tropics. Anim. Sci. 76:3-17.
- Olivier W.J. Snyman M.A. van WykJ. B. Erasmus G.J., 1998. Genetic parameters for fitness traits in Merino sheep. Livest. Prod. Sci. 56:71-77.
- Piwczyński D. Mroczkowski S. Sitkowska B., 2010. Survival analysis of lambs using Cox’s proportional hazards’ modeling and Kaplan-Meier method. Rocz. Nauk. Pol. Tow. Zoot. 6:81-89.
- Riggio V. Finocchiaro R. Bishop S.C., 2008. Genetic parameters for early lamb survival and growth in Scottish Blackface sheep. J. Anim. Sci. 86:1758-1764.
- Wiener G. Leet G.J. Woolliams J.A., 1992. Effects of rapid inbreeding and of crossing of inbred lines on the body weight growth of sheep. Anim. Prod. 55:89-99.
- Wooliams C. Wiener G. Macleod N.S.M., 1983. The effects of breed, breeding system and other factors on lamb mortality. 2. Factors influencing the incidence of delayed birth, dystokia, congenital defects and miscellaneous causes of early death. J. Agr. Sci. 100:553-561.