
Abstract
Thyroid hormones (TH) are the primary endocrine stimulators of non-shivering thermogenesis and are known to stimulate lactation. Triiodothyronine (T3) is the bioactive form, mainly derived by deiodination of thyroxine (T4), and the free quote (unbound to plasma proteins) is immediately bioavailable. This study aimed to evaluate potential relationships among TH in the blood, triiodothyronine in the milk (T3M), milk yield and environmental temperature in March to July for 8 lactating donkeys. Milk yield and blood TH concentrations changed significantly over time, whereas T3M was rather stable among individuals and not affected by time of sampling. Free T3 was not correlated with free T4 or with total TH in the blood, but it was weakly correlated with T3M. No relationship was found between blood TH and milk yield, which was negatively correlated with T3M. Thus, the absolute quantity of bioactive hormone in milk secretion is maintained. Milk yield was positively correlated with the free/total T3 and free T3/free T4 ratios, thus in turn with the relative quote of the circulating bioactive hormone. Circulating T3/T4 ratios were negatively correlated with environmental temperature. It is concluded that environmental temperature, in the range of the present study (-2 to 35°C), does not significantly entrain thyroid gland activity, which is affected more by other factors, such as inter-individual variations and physiological status (i.e., stage of lactation). However, increases in environmental temperature most likely induce decreases in deiodinase activity at the peripheral tissue level, as indicated by the decrease in the T3/T4 ratios in the blood.
Introduction
The thyroid hormones (TH), tetraiodothyronine or thyroxine (T4) and 3-3’-5-triiodothyronine (T3), act stimulating oxygen utilization and heat production in every cell of the body. The overall effects of these hormones are to increase the basal metabolic rate, make more glucose available to cells, stimulate protein synthesis, increase lipid metabolism and stimulate cardiac and neural functions. The thyroid gland, as such, secretes mostly T4, which is monodeiodinated to T3, the active form that binds to receptors at the target cells and exerts its biological effect (Capen and Martin, Citation1989). More than 99% of TH circulates in the blood bound to plasma proteins. Only the free hormone is responsible for the biological activity, and protein-bound hormones function as a promptly utilizable source, delaying the effects of decreased thyroid secretion and buffering against sudden increases in the thyroid’s secretory activity (Bartalena, Citation1990; Utiger, 1995). Thyroid hormones are the primary endocrine stimulators of non-shivering (facultative or adaptive) thermogenesis, thus regulating body temperature (Silva, Citation2005). A major exogenous regulator of thyroid gland activity is environmental temperature (Dickson, Citation1993); therefore, an inverse relationship between ambient temperature and blood TH concentrations often has been found in many animal species, such as domestic ruminants (Todini, Citation2007). Thyroid hormones are known to stimulate lactation (Tucker, 2000) and play a permissive role for galactopoietic hormones, such as prolactin and somatotropin (Kahl et al., Citation1995). During lactation, TH (together with insulin and cortisol) drives nutrient partitioning toward milk production and visceral protein synthesis (Motil et al., Citation1994). On the other hand, active T3 in colostrum and milk could play various physiological roles, such as serving as a source of systemic T3 and as a paracrine action that supports lactogenesis in the mother; on the other hand, physiological effects on the suckling offspring may be systemic and/or occur within the gastrointestinal tract (Sheard and Walker, Citation1988; Murray and Luba, Citation1993; Slebodzinski et al., Citation1995).
The ancient traditional utilization of donkeys (Equus asinus) as dairy animals excited a renewed popular and substantial scientific interest with regard to human nutrition in recent years. Clinical evidence shows that donkey milk is well tolerated by infants who are allergic to cows’ milk (Monti et al., Citation2007; Vita et al., 2007), and its use is reported to be useful in the treatment of human immune-related diseases and in the prevention of atherosclerosis (Tafaro et al., Citation2007). Donkey milk gross composition is similar to human milk, except for the low and variable average total solids and fat content (Salimei, Citation2011). Donkey mammary secretions contain compounds that can exert biological activities (i.e,. IGF-1, leptin, ghrelin and T3) (Salimei and Fantuz, Citation2012). Moreover, Todini et al. (Citation2012) reported that in donkey milk, the concentration of T3, a human-like bioactive compound, can be affected by the supply of dietary trace elements.
As part of the search for potential factors that entrain thyroid gland activity and TH bioavailability in donkeys, the present study aimed to identify relationships among TH concentrations in lactating donkeys, milk production and environmental temperatures.
Materials and methods
Location, animals and diet
The animal care procedures followed the European recommendations for the protection of experimental animals (EU Directive 2010/63/EU; European Commission, Citation2010).
The study was carried out on a farm in the Emilia Romagna region, Italy, at an altitude of 317 m a.s.l. and at latitude 44.38 °N, where approximately 500 donkeys are bred for dairy purposes.
Eight lactating, pluriparous jennies (6-11 years old), at 34-57 days post partum at the beginning of the study, were used to provide blood and milk samples. The day of samplings (every two weeks) they were weighted and their body condition scores were measured (on a 1 to 5 scale: Martin Rosset, Citation1990). Animals had continuous access to fresh water and to meadow hay and were fed 2.5 kg of mixed feed (12.6% crude protein as fed basis), divided into two meals. Jennies were housed with foals, which were separated from the dams 3 h before mechanical milking (Salimei et al., Citation2004).
Environmental temperatures were measured by a meteorological station near the farm and recorded by the Parma station. The following data were considered: maximum, minimum and mean temperatures of the sampling day, and the respective means for the 7 days preceding the samplings.
Samplings and assays
Individual milk and blood samples were collected every 2 weeks from 9 March to 1 June 2009 and then on 9 July 2009 (eight sampling times) at 1100 a.m. during the mechanical milking session (Salimei, Citation2011). Individual milk yield was recorded by graduated bulk, and the values were also expressed for yield relative to body weight (mL/kg body weight/milking). Because foals were approaching weaning, milk production at the last sampling date was scanty and was not considered for elaboration. Aliquots of milk samples were utilized for iodothyronine extraction with alkaline ethanol at low temperature, as described by Slebodzinski et al. (Citation1998) and by Todini et al. (Citation2010). Briefly, 2 mL of cold (-20°C) alkaline ethanol (pH 9.0 with NH4OH) were added to 1 ml of whole donkey milk, mixed thoroughly with a glass rod, vortexed and left in the freezer (-20°C) for 24 h. Then, the milk-ethanol mixture was vortexed, left in the freezer for a further 24 h and then centrifuged at 3500 g at 0°C for 30 min. The supernatant was collected and stored at -20°C. Twenty-four hours before the assay, these supernatant samples were diluted 1:1 with the assay buffer.
Blood samples were collected by jugular venipuncture in evacuated tubes containing K3-EDTA as anticoagulant (Venoject; Terumo Europe NV, Leuven, Belgium). Tubes were immediately centrifuged (2500 g for 15 min), and the plasma aliquots were stored at -20°C until assayed. The total concentrations of T3 in the extracted milk samples (T3M), total T3 and T4 in blood plasma (TT3P and TT4P), and free (unbound) T3 and T4 in blood plasma (FT3P and FT4P) were assayed using commercially available enzyme immunoassay (EIA) kits that are competitive ELISAs designed for clinical use in humans (T3KT1EW, T4KT2EW, FT3KT0037EW, FT4KT7EW; Radim, Rome, Italy). Assays were performed using the automated processor Brio 2 reader (Seac, Firenze, Italy). The assay kits were specifically validated for donkey species, as previously described (Todini et al., Citation2010). Intra- and inter-assay coefficients of variations were all between 1.9 and 8.4%. For each hormone, all samples from one animal were analyzed within the same assay session. Hormone concentrations were determined as the mean of duplicate determinations. The ratios of free/total hormones were also calculated, as well as the ratios of T3/T4 (both free and total). As a part of a larger study on lactating donkeys, data about changes of total TH with time have been partially published elsewhere (Todini et al., Citation2012).
During the trial, fresh bulk milk samples were analyzed by IR for crude protein, fat, and lactose content, and gravimetrically for total solids and ash content (Milkoscan, Foss, Hillerod, Denmark). The health of the mammary glands and milk hygiene were also monitored by somatic cell count (Fossomatic 360, Foss, Hillerod, Denmark) and total bacteria count (Bactoscan 8000, Foss, Hillerod, Denmark) in fresh bulk milk samples.
Statistical analysis
Statistical analyses were performed with SPSS 12.0 (SPSS Inc., Chicago, IL, USA; SPSS, Citation2003). All the investigated variables were checked for normal distribution using the Kolmogorov—Smirnov test. To meet ANOVA assumptions (i.e., normal distribution and homogeneity of variance) data were logarithmically (FT3P, TT4P, TT3P, T3M) or square-root (FT3P/TT3P, FT3P/FT4P, FT4P/TT4P, TT3P/TT4P) transformed for statistical elaboration. The results presented in tables and figures are least square means ± SE from the original data. The data were processed by analysis of variance for repeated measures to evaluate the effects of time of sampling. Differences between means were analyzed by least significant difference. Associations among variables were examined by calculating simple linear correlation (Pearson coefficient). Significant correlations were declared as strong (r>0.7), moderate (r from 0.3 to 0.7) or weak (r<0.3).
Results
At the beginning of the experiment the jennies have mean body weights of 205±32 kg and 3.45±0.39 body condition score, both of which did not significantly change during the trial.
Data from bulk milk samples indicated that the health of the udders (somatic cell count 8200/mL), milk hygiene (colony forming unit 3600/mL) and milk gross composition (crude protein 1.74%, fat 0.24%, lactose 7.05%, ash 0.40%) were within the ranges previously reported for donkey milk (Salimei and Fantuz, Citation2012). In agreement with previous studies, the milk protein content significantly (P<0.05) decreased during the trial (from 1.87 to 1.59%) but the fat and lactose content remained constant (Salimei and Fantuz, Citation2012).
Means for milk yield and TH concentrations are reported in and the respective trends during the study are described in . There was a significant (P<0.05) effect of time on milk yield and blood TH concentrations, whereas T3M was not affected. The free/total hormone concentrations ratios were affected by time (P<0.01 and P<0.05, for T3 and T4, respectively). The rates of bound circulating hormones ranged from 99.9991 to 99.9999% for T3 and from 99.9897 to 99.9997% for T4.
Table 1. Mean milk yield and thyroid hormone concentrations by eight jennies, and statistical significance.
Table 2. Correlations among thyroid hormone concentrations and milk yield by eight jennies.
Table 3. Correlations among thyroid hormone concentrations and milk yield by eight jennies with environmental temperatures.
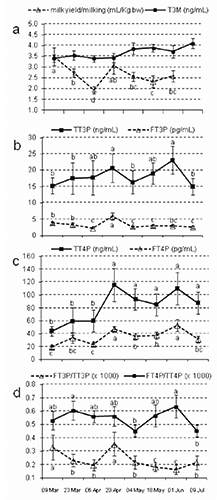
Correlations among hormone concentrations and milk yield are reported in . Total T3 and T4 in the blood were strongly correlated (r=0.8, P<0.0001). Free T4 in blood was strongly correlated with TT4P (r=0.72, P<0.0001) and TT3P (r=0.72, P<0.0001). Free T3 in blood was not correlated with TT3P, TT4P or FT4P, but it was weakly correlated with T3M. None of the hormone concentrations in the blood (TT3P, TT4P, FT3P, FT4P) were correlated with milk yield/milking (actual or expressed for body weight), but T3M was negatively correlated with the actual milk yield per milking. Milk yield/kg body weight was positively correlated with the FT3P/TT3P and FT3P/FT4P ratios.
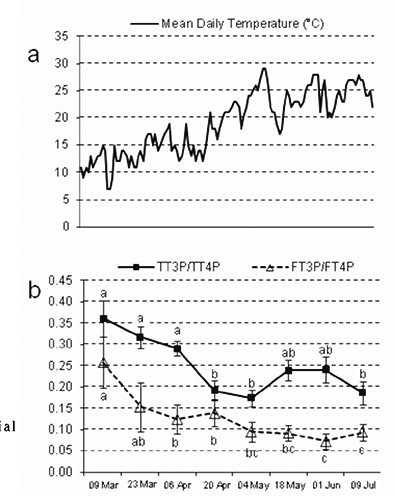
shows the mean daily environmental temperature and the ratios of T3/T4 in the blood. There was a significant effect of time on circulating T3/T4 ratios (P<0.05 and P<0.01 for TT3P/TT4P and FT3P/FT4P, respectively).
The correlations between thyroid hormone concentrations and milk yield with environmental temperatures are reported in . Milk T3 concentration showed moderate positive correlations with all the temperature parameters. Free T3 in blood was negatively correlated with the mean and and maximum temperatures of the day of sampling. The ratios TT3P/TT4P and FT3P/FT4P were negatively correlated with almost all the environmental temperatures, moderately but sometimes very significantly. Milk yield was negatively correlated with some temperature parameters.
Discussion
In the present study, total hormone concentrations in the blood showed widespread variations among individuals, with highest individual means being 4- to 6-fold higher than the lowest means, and the variability of the free fractions concentrations was lower (). In Racehorse, T4 values also showed large individual variations, while both free and total T3 were relatively stable among individuals (Bayly et al., Citation1996). The hypothesis is that the large variability in thyroid gland secretory activity (affected by both individual and exogenous factors) could be buffered by the differential binding affinities of the plasma proteins, in particular for the active hormone T3, which also showed a very wide range in the free/total hormone ratio (). The plasma concentrations of free T3 were not correlated with the concentrations of total TH and free T4 (). This could indicate that the effective amount of quickly bioavailable hormone is finally regulated and dependent on other factors mainly at tissue level, beyond the thyroid gland activity. Both T3 and T4 free/total ratios () showed profiles similar to those of the total hormone concentrations ( and ), with the respective peaks often coinciding; this may indicate that a certain degree of saturation of the plasma binding protein can occur. The percentages of bound fractions, which were well above 99.98% for both hormones, were similar to those found in pregnant and barren donkeys, by the same ELISA methods (Fazio et al., Citation2012).
It is noteworthy that the concentration of active T3 in milk was less variable among individuals and did not significantly change with time (). In mare milk lower levels than in our donkeys have been obtained by RIA: 0.5-0.76 ng/mL, associated with a mean of 0.9 ng/mL in maternal serum, giving an approximate colostrum/serum ratio of 0.82 (Slebodzinski et al., Citation1998). At a mean colostral T3 concentration of 0.456 mg/L and a daily intake of colostrum by a foal of about 4 L, the daily T3 consumed by a suckling foal is approximately 1.8 mg, which may likely have some stimulatory effects within the intestinal tract (Slebodzinski et al., Citation1998). Triiodothyronine concentration in milk is considered mainly dependent on deiodinase activity at the mammary level, as in mares they are strongly correlated (Slebodzinski et al., Citation1998). The correlation between free T3 in the blood and T3 in the milk () may indicate a certain transfer of active T3 from blood to milk, although no conclusions can be advanced from the current study.
During the current experiment, thermal conditions were not extreme and did not fall in the values commonly considered for heat stress (Silanikove, Citation2000). On many occasions, the maximum daily temperature was above 30°C, but the minimum during the night was always under 20°C (). Nonetheless, milk yield was negatively correlated with some parameters of environmental temperature (), which rose along with advancing season and stage of lactation (). Therefore, under the conditions of the present experiment, the potential effect of relatively high temperature on the milk yield decrease, as observed with advancing lactation, remains to be demonstrated. Milk yield showed no correlation with hormone concentrations in the blood, but was positively correlated with the FT3P/TT3P and FT3P/FT4P ratios (), thus with the relative quote of the bioactive hormone, according the well known stimulatory role of general metabolism and lactogenesis in particular (Tucker, 2000). The negative correlation between milk T3 concentrations and milk yield per milking () could either be due to a simple dilution effect or suggest a homeostatic control for maintaining an absolute quantity of active hormone in milk secretion. Therefore, concentrations of T3 in milk tended to increase when milk yield was decreasing, and this corresponded with advancing lactation and the spring season, and thus, to increasing environmental temperatures. We can only hypothesize such a mechanism as an explanation for the positive correlations found between milk T3 and temperature parameters ().
In the mild conditions of the present study, environmental temperature had weak or no effect on blood hormone concentrations, which mainly reflect gland secretion. Serum T4 showed no correlation with ambient temperature down to -31°C in Icelandic horses (Mejdell and Boe, Citation2005), and Cymbaluk (Citation1994) had previously also reported that horses do not respond to chronic cold exposure by increasing thyroid hormone secretion, which instead is stimulated more by lengthening photoperiod. In the present study, the supposed inhibitory effect of the increasing environmental temperature may have been overcome by the increasing day length. In fact, treatment with artificial photoperiod cycles clearly affected concentrations of circulating TH (Todini et al., Citation2006) and the light-induced increase in circulating hormones could play a role in sustaining advanced lactation. The galactopoietic effect of increasing day length have been demonstrated in dairy ruminants submitted to artificial photoperiod (Dahl et al., Citation2000, Citation2015). In goats, short-day treatment during the last third of pregnancy induced higher blood T3 afterwards during the next lactation, accompanied by higher milk yield (Mabjeesh et al., Citation2007). A retrospective study, based on a very large number of cow milk samples collected during several years, allowed to establish critical limits of temperature humidity index values for mammary health and milk composition (Bertocchi et al., Citation2014). The significant inverse relationship between environmental temperature and T3/T4 ratios (both free and total) () indicates that regulation of peripheral deiodination allows enough prompt and fine adjustments of bioavailability and activity at the target level, without significant changes in thyroid gland secretory activity. In ruminants, even in conditions of heat stress, the response of the thyroid gland is rather slow and the effects, such as the decline of blood TH concentrations, are only visible after several days (Silanikove, Citation2000). Also, the stimulatory effect of day length on blood T4 in goats was characterized by a long latency of several weeks (Todini et al., Citation2006). In this view, the positive correlations between T4 and minimal environmental temperatures () can be explained; the adjustments of thyroid gland secretory activity, for example in case of cold exposure, are likely preceded by a short-term stimulation of peripheral deiodination, which consumes circulating thyroxine, as reflected by the proportional increases of T3 and T3/T4 ratios. Vice-versa, in animals facing a hot climate, an inhibition of deiodinase activity occurs and less T4 is activated to T3. Therefore, whereas FT3 was negatively correlated with the daily mean and maximum temperature, such an inverse relationship was much more evident and constant for the T3/T4 ratios, for both free and total hormone levels ().
Conclusions
In conclusion, the concentrations of the bioactive triiodothyronine in donkey milk are not negligible and do not vary between individuals and over time, unlike the hormone concentrations in the blood. Moreover, the inverse relationship between milk T3 concentration and milk yield may indicate that adequate amounts of the hormone are maintained and can be acquired by the suckling foal, even when milk yield decreases, i.e., with advancing lactation and toward weaning.
We also conclude that environmental temperature, in the range of the present study (-2 to 35°C), does not significantly affect thyroid gland activity. However, an increase in environmental temperature most likely induces a decrease in the deiodinase activity at the peripheral tissue level, as indicated by the decrease in the T3/T4 ratios in the blood.
The hormone bioavailability and effects at the target level can be regulated by changes in thyroid gland secretion, activity of deiodinase enzymes, and plasma binding proteins. At each of these levels, many endogenous, as well as exogenous, regulatory factors can act. Considering that donkeys in the world are most commonly located in areas characterized by a harsh climate, thyroid hormone physiology is a main topic among the mechanisms that allow the animals to adapt to the surrounding environment, thereby maintaining their welfare and productions.
Acknowledgements
This paper was partly funded by the University of Camerino (FAR L. Todini, F. Fantuz, A. Malfatti). The authors gratefully acknowledge the valuable collaboration of Mr. D. Borghi, Mr. G. Borghi and Mrs. T. Faietti (Azienda Agricola Montebaducco, Salvarano di Quattro Castella, Reggio Emilia, Italy). The authors are also grateful to Mr. G. Lebboroni (University of Camerino) and Mr. A. El Jeddad (Az. Agr. Montebaducco) for technical assistance.
References
- BartalenaL., 1990. Recent achievements in studies on thyroid hormone-binding proteins. Endocr. Rev. 11:47–64.
- BaylyW. AndreaR. SmithB. StenslieJ. BergsmaG., 1996. Thyroid hormone concentrations in racing Thoroughbreds. Pferdeheilkunde 12:534–538.
- BertocchiL. VitaliA. LaceteraN. NardoneA. VariscoG. BernabucciU., 2014. Seasonal variations in the composition of Holstein cow’s milk and temperature-humidity index relationship. Animal 8:667–674.
- CapenC.C. MartinS.L., 1989. The thyroid gland. In: McDonaldL.E. PinedaM.H. (eds.) Veterinary endocrinology and reproduction. 4th ed. Lea and Febiger, Philadelphia, PA, USA, pp 58–91.
- CymbalukN.F., 1994. Thermoregulation of horses in cold, winter weather. Livest. Prod. Sci. 4:65–71.
- DahlG.E. BuchananB.A. TuckerH.A., 2000. Photoperiodic effects on dairy cattle: a review. J. Dairy Sci. 83:885–893.
- DahlG.E. TaoS. ThompsonI.M., 2015. Lactation biology symposium: effects of photoperiod on mammary gland development and lactation. J. Anim. Sci. 90:755–760.
- DicksonW.M., 1993. Endocrine glands. In: SwensonM.J. ReeceW.O. (eds.) Duke’s physiology of domestic animals. 11th ed. Comstock Publishers Association, Ithaca, NY, USA, pp 629–664.
- European Commission, 2010. Directive of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes, 2010/63/EU. In: Official Journal, L 276/33, 20-10-2010.
- FazioE. MedicaP. CravanaC. FerlazzoA., 2012. Total and free iodothyronines profile in the donkey (Equus asinus) over a 12-month period. Acta Vet. Brno 81:239–244.
- KahlS. CapucoA.V. BinelliM. VanderkooiW.K. TuckerH.A. MoseleyW.M., 1995. Comparison of growthhormone-releasing factor and somatotropin: thyroid status of lactating, primiparous cows. J. Dairy Sci. 78:2150–2158.
- MabjeeshS.J. Gal-GarberO. ShamayA., 2007. Effect of photoperiod in the third trimester of gestation on milk production and circulating hormones in dairy goats. J. Dairy Sci. 90:699–705.
- Martin RossetW., 1990. L’alimentation des chevaux. INRA Edition, Paris, France.
- MejdellC.M. BoeK.E., 2005. Responses to climatic variables of horses housed outdoors under Nordic winter conditions. Can. J. Anim. Sci. 85:301–308.
- MontiG. BertinoE. MuratoreM.C. CosciaA. CresiF. SilvestroL. FabrisC. FortunatoD. GiuffridaM.G. ContiA., 2007. Efficacy of donkey’s milk in treating highly problematic cow’s milk allergic children: and in vivo and in vitro study. Pediatr. Allergy Immu. 18:258–264.
- MotilK.J. ThotathucheryM. MontandonC.M. HacheyD.L. BouttonT.W. KleinP.D. GarzaC., 1994. Insulin, cortisol and thyroid-hormones modulate maternal protein status and milk-production and composition in humans. J. Nutr. 124:1248–1257.
- MurrayM.J. LubaN.K., 1993. Plasma gastrin and somatostatin, and serum thyroxine (T4), triiodothyronine (T3), reverse triiodothyronine (rT3) and cortisol concentrations in foals from birth to 28 days of age. Equine Vet. J. 25:237–239.
- SalimeiE., 2011. Animals that produce dairy foods. Donkey. In: FuquayJ.W. FoxP.F. McSweeneyP.L.H. (eds.) Encyclopedia of dairy sciences. 2nd ed. Oxford Academic Press, Oxford, UK, pp 365–373.
- SalimeiE. FantuzF., 2012. Equid milk for human consumption. Int. Dairy J. 24:130–142.
- SalimeiE. FantuzF. CoppolaR. ChiofaloB. PolidoriP. VariscoG., 2004. Composition and characteristics of ass’s milk. Anim. Res. 53:67–78.
- SheardN.F. WalkerW.A., 1988. The role of breast milk in the development of the gastrointestinal tract. Nutr. Rev. 46:1–8.
- SilanikoveN., 2000. Effects of heat stress on the welfare of extensively managed domestic ruminants. Livest. Prod. Sci. 67:1–18.
- SilvaJ.E., 2005. Thyroid hormone and the energetic cost of keeping body temperature. Bioscience Rep. 25:129–148.
- SlebodzinskiA. Brzezinska-SlebodzinskaE. NowakJ. KowalskaK., 1998. Triiodothyronine (T3), insulin and characteristics of 5’-monodeiodinase (5’-MD) in mare’s milk from parturition to 21 days post-partum. Reprod. Nutr. Dev. 38:235–244.
- SlebodzinskiA.B. LipczakW. Brzezinska-SlebodzinskaE., 1995. Peroral administration of triiodothyronine (3T) affects absorption of immunolactoglobulins in calves. Reprod. Nutr. Dev. 35:387–393.
- SPSS, 2003. SPSS 12.0 applications guide. SPSS Inc., Chicago, IL, USA.
- TafaroA. MagroneT. JirilloF. MartemucciG. D’AlessandroA.C. AmatiL. JirilloE., 2007. Immunological Properties of donkey’s milk: its potential use in the prevention of atherosclerosis. Curr. Pharm. Design 13:3711–3717.
- TodiniL., 2007. Thyroid hormones in small ruminants: effects of endogenous, environmental and nutritional factors. Animal 1:997–1008.
- TodiniL. DelgadilloJ.A. DebenedettiA. ChemineauP., 2006. Plasma total T3 and T4 concentrations in bucks as affected by photoperiod. Small Ruminant Res. 65:8–13.
- TodiniL. MalfattiA. SalimeiE. FantuzF., 2010. Measurement of thyroid hormones in donkey (Equus asinus) blood and milk: validation of ELISA kits and evaluation of 3 sample collection, handling and storage. J. Dairy Res. 76:1–6.
- TodiniL. SalimeiE. MalfattiA. FerraroS. FantuzF., 2012. Thyroid hormones in milk and blood of lactating donkeys as affected by stage of lactation and dietary supplementation with trace elements. J. Dairy Res. 79:232–237.