
Abstract
As a widely used warm-season turfgrass in landscapes and golf courses, bermudagrass encounters multiple abiotic stresses during the growth and development. Physiology analysis indicated that abiotic stresses induced the accumulation of ROS and decline of photosynthesis, resulting in increased cell damage and inhibited growth. Proteomic and metabolomic approaches showed that antioxidant enzymes and osmoprotectant contents (sugar, sucrose, dehydrin, proline) were extensively changed under abiotic stress conditions. Exogenous application of small molecules, such as ABA, NO, CaCl2, H2S, polyamine and melatonin, could effectively alleviate damages caused by multiple abiotic stresses, including drought, salt, heat and cold. Based on high through-put RNA seq analysis, genes involved in ROS, transcription factors, hormones, and carbohydrate metabolisms were largely enriched. The data indicated that small molecules induced the accumulation of osmoprotectants and antioxidants, kept cell membrane integrity, increased photosynthesis and kept ion homeostasis, which protected bermudagrass from damages caused by abiotic stresses.
Abbreviations
| ABA | = | abscisic acid |
| NO | = | nitrogen oxide |
| ROS | = | reactive oxygen species |
| TCA | = | tricarboxylicacid |
| GO | = | gene ontology |
| RCAR | = | regulatory component of ABA receptors |
| PYR/PYL | = | pyrabactin resistance/PYR like |
| SnRK2 | = | SNF1-related protein kinases 2 |
| NCED | = | 9-cis-epoxycarotenoid dioxygenase |
| JA | = | jasmonate |
| JAZ | = | jasmonate ZIM-domain |
Bermudagrass (Cynodon dactylon) is a warm-season turfgrass and widely used on home lawns, golf courses and sport fields. Bermudagrass is adapted to cultivation in a wide range of climate condition. Analysis of natural variations of drought stress tolerance revealed that different bermudagrass cultivars exhibited varied tolerance to drought stress.Citation1-3 Exogenous application of small molecules could effectively alleviate damages caused by multiple abiotic stresses, including drought, salt and cold.Citation4-8 Exogenous ABA treatment improved drought stress tolerance in ‘TifEagle’ cultivar through induction of H2O2 and NO, which, in turn, enhanced antioxidant enzyme activities.Citation4 Our studies indicated that exogenous CaCl2, polyamines, H2S, and NO treatments were effective to induce bermudagrass tolerance to drought, salt, and cold stresses.Citation5,6,8
Abiotic stress tolerance is a complex trait, therefore plants develop different approaches to cope with adverse environmental conditions. About half century ago, Barnett and Naylor found that water stress induced a 10- to 100-fold accumulation of free proline and a 2- to fold6- accumulation of free asparagines.Citation9 Further studies showed that abiotic stress treatments changed antioxidant enzyme activities, ROS contents, osmoprotectant contents (sugar, sucrose, dehydrin, proline), and photosynthesis.Citation1-2,Citation5-6,Citation10-14
Recently, proteomics and metabolomics approaches have been successfully used to detect omic level changes during bermudagrass stress responses. Under water-deficit condition, 32 proteins had increases in the abundance and 22 proteins exhibited decreases in the abundance, which were mainly involved in metabolism, energy, cell growth/division, protein synthesis and stress defense.Citation15 In previous study, we found that chilling treatment modulated the abundance changes of 28 proteins and in total 51 proteins were regulated by CaCl2 treatment. These proteins were enriched in redox, tricarboxylicacid cycle, glycolysis, photosynthesis, oxidative pentose phosphate pathway, and amino acid metabolisms.Citation8 Additionally, we identified 39 proteins with significantly changed abundance after drought stress treatment in leaves and stems of Yukon and Tifgreen cultivars.Citation3 Among them, proteins involved in photosynthesis, glycolysis, N-metabolism, TCA and ROS pathways were enriched. Pretreatment of exogenous polyamine conferred increased salt and drought tolerances in bermudagrass. Comparative proteomic analysis revealed that 36 commonly regulated proteins by at least 2 types of polyamines in bermudagrass. Among them, proteins involved in ROS, electron transport and energy pathways were largely enriched.Citation5 Metabolomics analyses confirmed that amino acids, organic acids, sugars, and sugar alcohols were significantly changed after stress treatments.Citation8,16 All these results indicated that abiotic stresses or exogenous small molecule treatments extensively regulated down-stream protein/enzyme and metabolite biosynthesis.
To further detect up-stream changes in bermudagrass upon stress treatment, macroarray analysis was performed and 189 drought responsive candidate genes were identified, of which 120 were up-regulated and 69 were down-regulated. Functional annotation analysis suggested that upregulated genes were mainly involved in proline biosynthesis, signal transduction pathways, protein repair systems, and removal of toxins, while downregulated genes were mostly related to basic plant metabolism such as photosynthesis and glycolysis.Citation17
More recently, high throughput RNA seq analysis showed that exogenous melatonin treatment conferred improved salt, drought, and cold stress tolerances in bermudagrass through modulation of 3933 genes (2361 up-regulated and 1572 down-regulated).Citation7 Melatonin (N-acetyl-5-methoxytryptamine) is a well-known animal hormone which is also found to play important roles in plant development and abiotic stress responses.Citation18-19 Exogenous melatonin treatment alleviated ROS burst and cell damage induced by abiotic stress in bermudagrass. This was consistent with transcriptomic data which showed that redox-related genes were enriched.Citation7 Moreover, pathway and GO term enrichment analyses indicated that 8 pathways were over-represented after melatonin pre-treatment, including nitrogen metabolism, major carbohydrate metabolism, TCA/org transformation, transport, hormone metabolism, metal handling, redox, and secondary metabolism. Interestingly, several key genes involved in ABA (RCAR/PYR/PYL, SnRK2, and NCED3) and JA (JAZs) signaling were significantly changed after melatonin pre-treatment, which might be contributed to increased abiotic stress tolerance in bermudagrass.Citation7
Conclusions
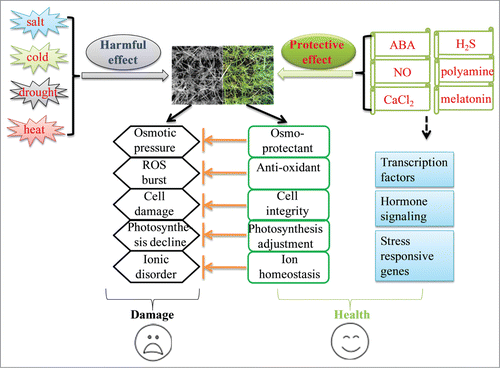
Based on these observations, a possible model that depicting bermudagrass abiotic stress responses was proposed (). Abiotic stress treatments severely inhibited plant growth and development through osmotic pressure, ROS burst, cell damage, photosynthesis decline, and ionic disorder. Exogenous application of small molecules, including ABA, NO, CaCl2, H2S, polyamine and melatonin, induced the accumulation of osmoprotectants and antioxidants, kept cell membrane integrity, increased photosynthesis and kept ion homeostasis. These treatments might also induce the expressions of transcription factor and hormone signaling related genes to activate down-stream stress responsive pathways, resulting in stress resistance in bermudagrass.
Figure 1. A possible model showing mechanisms involved small molecules induced bermudagrass stress resistance.
Acknowledgments
We thank Prof. Frantisek Baluska for the invitation for this addendum.
Funding
This research was supported by “the Hundred Talents Program” (Grant Nos. 54Y154761O01076 and Y329631O0263) to Zhulong Chan, and by the Knowledge Innovative Key Program of Chinese Academy of Sciences (No. 55Y455446O0544) to Haitao Shi.
Related Research Data
References
- Lu S, Peng X, Guo Z, Zhang G, Wang Z, Wang C, Pang C, Fan Z, Wang J. In vitro selection of salinity tolerant variants from triploid bermudagrass (Cynodon transvaalensis x C. dactylon) and their physiological responses to salt and drought stress. Plant Cell Rep 2007; 26:1413-20; PMID:17377793; http://dx.doi.org/10.1007/s00299-007-0339-2
- Shi H, Wang Y, Cheng Z, Ye T, Chan Z. Analysis of natural variation in bermudagrass (Cynodon dactylon) reveals physiological responses underlying drought tolerance. PLoS ONE 2012; 7:e53422; PMID:23285294; http://dx.doi.org/10.1371/journal.pone.0053422
- Shi H, Ye T, Chan Z. Comparative proteomic responses of two bermudagrass (Cynodon dactylon (L). Pers.) varieties contrasting in drought stress resistance. Plant Physiol Biochem 2014b; 82:218-28; http://dx.doi.org/10.1016/j.plaphy.2014.06.006
- Lu S, Su W, Li H, Guo Z. Abscisic acid improves drought tolerance of triploid bermudagrass and involves H2O2- and NO-induced antioxidant enzyme activities. Plant Physiol Biochem 2009b; 47:132-8; http://dx.doi.org/10.1016/j.plaphy.2008.10.006
- Shi H, Ye T, Chan Z. Comparative proteomic and physiological analyses reveal the protective effect of exogenous polyamines in the bermudagrass (Cynodon dactylon) response to salt and drought stresses. J Proteome Res 2013a; 12:4951-64; http://dx.doi.org/10.1021/pr400479k
- Shi H, Ye T, Chan Z. Exogenous application of hydrogen sulfide donor sodium hydrosulfide enhanced multiple abiotic stress tolerance in bermudagrass (Cynodon dactylon (L). Pers.). Plant Physiol Biochem 2013b; 71:226-34; http://dx.doi.org/10.1016/j.plaphy.2013.07.021
- Shi H, Jiang C, Ye T, Tan D, Reiter RJ, Zhang H, Liu R, Chan Z. Comparative physiological, metabolic and transcriptomic analyses reveal mechanisms of improved abiotic stress resistance in bermudagrass (Cynodon dactylon (L). Pers.) by exogenous melatonin. J Exp Bot 2015; 66:681-94; http://dx.doi.org/10.1093/jxb/eru373.
- Shi H, Ye T, Zhong B, Liu X, Chan Z. Comparative proteomic and metabolomic analyses reveal mechanisms of improved cold stress tolerance in bermudagrass (Cynodon dactylon (L). Pers.) by exogenous calcium. J Integ Plant Biol 2014; 56:1064-79; http://dx.doi.org/10.1111/jipb.12167
- Barnett NM, Naylor AW. Amino Acid and protein metabolism in bermudagrass during water stress. Plant Physiol 1966; 41:1222-30; PMID:16656387; http://dx.doi.org/10.1104/pp.41.7.1222
- Lu S, Chen C, Wang Z, Guo Z, Li H. Physiological responses of somaclonal variants of triploid bermudagrass (Cynodon transvaalensis x Cynodon dactylon) to drought stress. Plant Cell Rep 2009a; 28:517-26; http://dx.doi.org/10.1007/s00299-008-0649-z
- HuL, Wang Z, Du H, Huang B. Differential accumulation of dehydrins in response to water stress for hybrid and common bermudagrass genotypes differing in drought tolerance. J Plant Physiol 2010; 167:103-9; PMID:19716198; http://dx.doi.org/10.1016/j.jplph.2009.07.008
- Carmo-Silva AE, Keys AJ, Andralojc PJ, Powers SJ, Arrabaca MC, Parry MA. Rubisco activities, properties, and regulation in three different C4 grasses under drought. J Exp Bot 2010; 61:2355-66; PMID:20363871; http://dx.doi.org/10.1093/jxb/erq071
- Carmo-Silva AE, Powers SJ, Keys AJ, Arrabaca MC, Parry MA. Photorespiration in C4 grasses remains slow under drought conditions. Plant Cell Environ 2008; 31:925-40; PMID:18331589; http://dx.doi.org/10.1111/j.1365-3040.2008.01805.x
- Carmo-Silva AE1, Keys AJ, Beale MH, Ward JL, Baker JM, Hawkins ND, Arrabaca MC, Parry MA. Drought stress increases the production of 5-hydroxynorvaline in two C4 grasses. Phytochemistry 2009; 70: 664-71; PMID:19359022; http://dx.doi.org/10.1016/j.phytochem.2009.03.001
- Zhao Y, Du H, Wang Z, Huang B. Identification of proteins associated with water-deficit tolerance in C4 perennial grass species, Cynodon dactylon x Cynodon transvaalensis and Cynodon dactylon. Physiol Plant 2011; 141:40-55; PMID:21029106; http://dx.doi.org/10.1111/j.1399-3054.2010.01419.x
- Du H, Wang Z, Yu W, Liu Y, Huang B. Differential metabolic responses of perennial grass Cynodon transvaalensis x Cynodon dactylon (C4) and Poa Pratensis (C3) to heat stress. Physiol Plant 2011; 141:251-64; PMID:21114672; http://dx.doi.org/10.1111/j.1399-3054.2010.01432.x
- Kim C, Lemke C, Paterson AH. Functional dissection of drought-responsive gene expression patterns in Cynodon dactylon L. Plant Mol Biol 2009; 70:1-16; PMID:19152115; http://dx.doi.org/10.1007/s11103-009-9453-6
- Posmyk MM, Bałabusta M, Wieczorek M, Sliwinska E, Janas KM. Melatonin applied to cucumber (Cucumis sativus L.) seeds improves germination during chilling stress. J Pineal Res 2009a; 46:214-23; http://dx.doi.org/10.1111/j.1600-079X.2008.00652.x
- Yin L, Wang P, Li M, Ke X, Li C, Liang D, Wu S, Ma X, Li C, Zou Y, et al. Exogenous melatonin improves Malus resistance to Marssonina apple blotch. J Pineal Res 2013; 54:426-34; PMID:23356947