

Abstract
Streptozotocin (STZ) is used to induce diabetes in experimental animals. It has a variety of adverse effects, ranging from nausea, emesis, and weight loss to liver damage, renal failure, and metabolic acidosis. STZ also has effects on the immune system, being associated with lymphopenia in rodents, the mechanism of which is not fully understood. We present data on a significant STZ-associated reduction in lymphocyte count in nonhuman primates. We report a significant reduction in absolute lymphocyte count; in 2 monkeys, the lymphopenia persisted for >100 d. However, a significant increase in absolute monocyte count was noted. Furthermore, an increase in serum monocyte chemoattractant protein-1 (MCP-1) was observed. The reduction in lymphocyte numbers may contribute to immunomodulation that may be beneficial to a subsequent islet graft, and may reduce the need for immunosuppressive therapy. The increase in monocytes and MCP-1, however, may be detrimental to the islet graft. Studies are warranted to explore the mechanism by which STZ has its effect.
Abbreviations:
| STZ | = | streptozotocin |
| WBC | = | white blood cell |
Introduction
Streptozotocin (STZ) is a glucosamine derivative of nitrosourea that is preferentially toxic to pancreatic β-cells.Citation1 It is used as a chemotherapeutic agent in the treatment of (a) metastatic pancreatic islet cell tumors,Citation2,3 (b) metastatic carcinoid tumors and syndrome,Citation4 and (c) pancreatic adenocarcinoma.Citation5 STZ is also employed to induce diabetes in experimental animals.Citation6-8
STZ has a broad toxicity profile affecting many organs and tissues. Elevated liver enzymes, raised serum creatinine, a protein-losing nephropathy, and renal failure have been noted in animals receiving STZ.Citation9 Histological evaluations of organs have demonstrated renal tubular necrosis and hepatic steatosis.Citation7,10,11 Other reported toxicities include nausea, emesis, weight loss, interrupted growth, hormone imbalance, lipid metabolism derangements, metabolic acidosis, and pulmonary edema.Citation11,12
In rodents, STZ has been reported to affect the immune system, being associated with bone marrow suppression, derangements in leukocyte count (leukopenia), and lymphocyte depletion in the blood and spleen, the mechanisms of which are not completely understood.Citation13-16 Direct and irreversible damage to the bone marrow and the early precursors of T cells is seen in STZ-treated mice.Citation17 Levine et al studied the toxicological effects of STZ in mice and in large animals (dogs and rhesus monkeys), and reported necrosis of lymphoid organs, and bone marrow hypoplasia.Citation18 The present report consists of our observations in cynomolgus monkeys.
Results
Metabolic monitoring
STZ administration resulted in marked hyperglycemia in all monkeys (). Before STZ, the mean fasting blood glucose was 57 ± 14 mg/dl (range 27–85 mg/dl). During the two weeks after STZ, it rose to 148 ± 36 mg/dl (range 80–204 mg/dl) despite insulin therapy.
Figure 1. Fasting blood glucose (mean+standard error) during the first month after streptozotocin (STZ) administration in cynomolgus monkeys (n = 14).
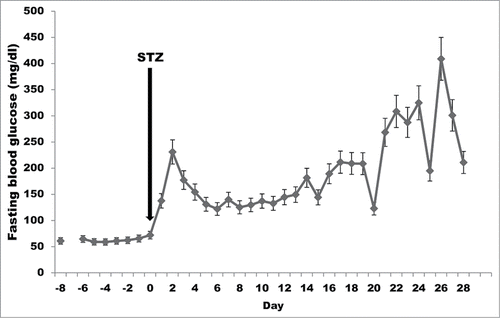
The mean HbA1c in non-diabetic cynomolgus monkeys has been reported to be 3.5–5.0%.Citation20 In the present study, before STZ the mean HbA1c was 4.7 ± 0.44%. Two weeks after STZ, the mean HbA1c was 5.9 ± 0.77% in the 5 animals tested. HbA1c fluctuated between 8.2% at 48 d (in one monkey) and 8.9%, 6.4%, and 7.6% at 61, 77, and 103 d (in a second monkey). Despite insulin treatment, a third monkey had an HbA1c of 9.9% at 153 d after STZ administration.
Hematologic and immunologic monitoring
The mean total WBC count did not change significantly after STZ ( and ), and neither did the mean absolute neutrophil count. However, a significant reduction was documented in the mean absolute lymphocyte count (pre-STZ 6.1 × 109/L vs. post-STZ 4.7 × 109/L; p < 0.05) and in the mean percentage of lymphocytes (pre-STZ 55.12% vs. post-STZ 44.36%; p < 0.05). In contrast, a statistically significant increase was noted in the mean absolute monocyte count (pre-STZ 0.55 × 109/L vs. post-STZ 0.77 × 109/L; p < 0.05), and in the percentage of monocytes (pre-STZ 4.9% vs. post-STZ 7.4%; p < 0.05).
Table 1. Mean total and differential white blood cell (WBC) counts pre- and post-streptozotocin (STZ) administration in cynomolgus monkeys (n = 14)
Figure 2. Total white blood cell (WBC) count and absolute numbers of neutrophils, lymphocytes, and monocytes in cynomolgus monkeys (n = 14) pre- and post-streptozotocin (STZ).
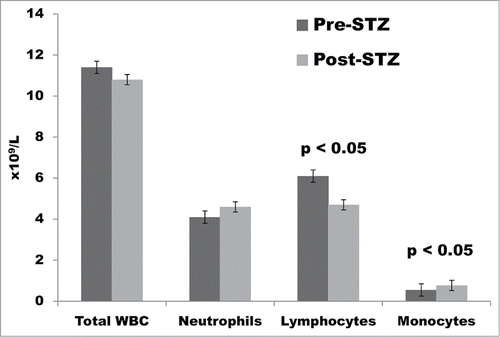
The reduction in absolute lymphocyte count after STZ was maintained until the time of islet transplantation, i.e., for a mean period of 27 d (range 2–156 d) with 2 monkeys demonstrating persisting lymphopenia (2.21 and 2.77 × 109/L, respectively) for >100 d. This remarkably prolonged effect may have been a result of the hyperglycemia that developed after STZ, rather than a continuing direct effect of STZ itself (discussed below).
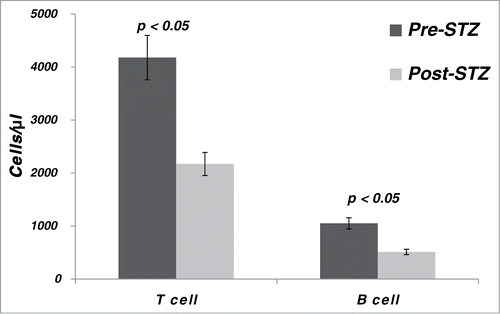
In the 4 monkeys tested, flow cytometry analysis of lymphocyte subgroups showed a proportionate reduction in both T and B cells after STZ administration (). The mean absolute T cell count decreased from 4,005 ± 1,334 cells/μl to 2,172 ± 890 cells/μl (p < 0.05) and the mean B cell count decreased from 963 ± 261 cells/μl to 512 ± 198 cells/μl (p < 0.05).
Figure 3. T and B lymphocyte counts in cynomolgus monkeys (n = 4) pre- and post streptozotocin (STZ).
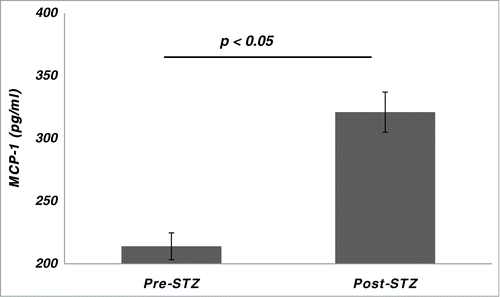
The 6 serum cytokines measured all increased after STZ, but a statistically significant increase was seen only in mean MCP-1, which (6 ± 4 d after STZ) increased from 214 ± 82 pg/mL to 321 ± 105 pg/mL (p < 0.05) ().
Figure 4. Serum levels of monocyte chemo-attract protein (MCP)-1 in cynomolgus monkeys (n = 8) pre- and post-streptozotocin (STZ).
Discussion
When used in clinical practice, e.g., in the treatment of patients with metastatic pancreatic islet cell, carcinoid, or pancreatic adenocarcinomas,Citation2-5 STZ is administered at a dose of 500 mg/m2 daily for 5 d every 6 weeks or as a single dose of 1000 mg/m2 once weekly for 2 weeks.Citation21 Further treatment is then determined based on the toxicity/adverse effects seen in the patient. STZ has a high toxicity profile, being associated with severe nausea, vomiting, nephrotoxicity, and myelosuppression.Citation21 Myelosuppression can manifest as anemia, leukopenia, neutropenia, and/or thrombocytopenia,Citation21 though an absolute lymphopenia does not appear to have been reported. Metabolic adverse effects, e.g., mild-to-moderate reversible abnormalities of glucose tolerance, have been reported in some patients,Citation21 but are possibly less common because of the different dosing regimen when compared with that used in experimental animals.
The diabetogenic effect of STZ was first reported in rats by Rakieten in 1963.Citation22 STZ is a cytotoxic glucose analog that exerts its effects via interaction of its glucose moiety with the GLUT-2 glucose transport protein on pancreatic β-cells.Citation23 In experimental animals such as mice, high-dose STZ (150–200 mg/kg body weight) causes cellular DNA damage by alkylation or fragmentation of DNA or by generation of nitric oxide,Citation15 resulting in β-cell death. STZ also targets mitochondrial DNA and impairs mitochondrial function, resulting in inhibition of insulin secretion in response to glucose stimulation.1 Low-dose STZ (40–50 mg/kg body weight), given repeatedly, elicits an inflammatory/immune response that mimics autoimmunity and type 1 diabetes mellitus in mice.Citation24 As STZ acts via the GLUT-2 receptor, toxicity is logically seen to affect organs/tissues that express GLUT-2, i.e., mainly liver and kidneys.
The hyperglycemic state of diabetes by itself has an immunosuppressive effect. Acute hyperglycemia causes lymphopenia,Citation16,25 and a chronic insulin-deficient diabetic state is associated with decreased proliferation and differentiation of lymphoid cells.Citation26 The STZ-induced immunosuppressive effect and that associated with hyperglycemia/diabetes may thus be additive. In the present study, hyperglycemia was observed in all monkeys after STZ administration, and within two weeks appeared to be associated with some increase in HbA1c. Therefore, a role for hyperglycemia specifically inducing and/or maintaining lymphopenia cannot to be excluded.
Alloxan, unlike STZ, does not cause a direct depletion of lymphocytes and thymocytes in mice,Citation27 but the hyperglycemia it causes may result in lymphopenia. Diabetes induction using other non-STZ agents, e.g., diphtheria toxin in rat insulin promoter diphtheria toxin receptor (RIP-DTR) transgenic mice, has been associated with a reduction in leukocytes and splenocytes.Citation16
STZ-induced changes in the haematopoietic and reticulo-endothelial systems of mice, dogs, and monkeys were documented by Levine et al.Citation18 The maximum dose of STZ used on monkeys in their study was 80 mg/kg, which is less than that used in most nonhuman primate islet transplant models. The median lethal dose of STZ for monkeys in Levine's study (80 mg/kg) was found to cause necrosis of lymphoid tissue, bone marrow hypoplasia, anemia, and neutrophilia. Toxic effects of STZ were, however, seen even at very low doses (5 mg/kg). Similar findings of lymphopenia, lymphoid tissue necrosis, and bone marrow hypoplasia, anemia and neutrophilia were also noted in dogs.Citation18
The dosage of STZ used to induce diabetes in mice, however, is much higher than the maximum dose of STZ used by Levine et al in monkeys. Single-dose STZ in the range of 100–300 mg/kg body weight is used in different strains of mice.Citation28
The present study also used a dose of STZ similar to that used in mice. However these doses are much higher than those used in human patients.Citation21 STZ is associated with an absolute lymphopenia and, in particular, depletion of T lymphocytes, which is seen in both the blood and spleen.Citation16 The present study documented a statistically significant decrease in absolute lymphocyte count in the blood, and a proportionate reduction in both T and B cell numbers.
Direct toxicity of STZ on CD8+ T and B cells has been reported in an in vitro model.Citation15,16 STZ is also associated with a relative increase in T regulatory cell (CD4+CD25+Foxp3+) numbers in the blood and with an immunosuppressive effect in vitro.Citation16 These changes in the T regulatory cell numbers after STZ were seen despite strict blood glucose control (by using insulin pumps or transplant of syngeneic mice islets under the kidney capsule of the STZ-induced diabetic mice).Citation16 The direct immunosuppressive effect of STZ is demonstrated by its ability to prevent development of autoimmune diabetes in young non-obese diabetic (NOD) mice.Citation15 This ability to prevent development of autoimmune diabetes was abrogated when the mice were depleted of CD8+ T cells, suggesting a role for CD8+ T regulatory cells in mediating the immunological effects of STZ.Citation15 Bone marrow cells from STZ-diabetic mice are unable to reconstitute gamma-irradiated normal syngeneic mice.Citation17 Islet and skin allografts also survive longer in STZ-induced diabetic mice compared to naïve mice.Citation16 Greater numbers of Foxp3+ T regulatory cells were found within the grafts in STZ diabetic mice compared to naïve mice.Citation16 Although STZ acts via GLUT-2, the effects of STZ on lymphocytes must be mediated through other (unknown) mechanisms as lymphocytes do not express GLUT-2.Citation16
In the present study, a statistically significant increase in absolute monocyte count (by 40%; p < 0.05) was seen (which does not appear to have been reported previously), and may possibly be detrimental to islet graft outcome.Citation29,30 The increases in cytokines seen after administration of STZ, and specifically MCP-1, may be indirect evidence of an inflammatory response to the agent. Islets constitutively express MCP-1 in the absence of inflammatory infiltration.Citation30,31 Furthermore, a high level of MCP-1 has been associated with poor engraftment of transplanted islets.Citation31 Thus, a significant increase in MCP-1 after STZ administration may be detrimental to subsequently transplanted islets.
Despite multiple studies on STZ and its metabolic implications in large animals,Citation7,Citation9-11 to our knowledge there has been no published study to date on the immunomodulatory effect of STZ in pigs or nonhuman primates, although Wu et al reported a statistically significant reduction in total WBC count in STZ-treated male cynomolgus monkeys compared to naïve ones.Citation12
In summary, the reduction in lymphocyte numbers may provide a degree of immunosuppression (and therefore be beneficial to a subsequent islet transplant), and influence the subsequent dosage of immunosuppressive therapy that needs to be administered. However, the increase in monocyte numbers and MCP-1 may be detrimental to survival of a subsequent islet graft. More detailed studies are needed in STZ-treated nonhuman primates to determine the mechanism by which STZ has its effect.
Methods
Induction of diabetes in cynomolgus monkeys
Cynomolgus monkeys (n = 14; Three Springs Scientific, Perkasie, PA) that were rendered diabetic in a study of pig-to-cynomolgus monkey islet xenotransplantation were included in this analysis. STZ (Zanosar, Sicor Pharmaceuticals, Irvine, CA; 125–150 mg/kg i.v.) was administered and blood samples were drawn at various intervals of time before and after STZ administration (with post-STZ follow-up for 2 days to 156 days [mean 27 days] before islet transplantation was performed).
All animal care procedures were in accordance with the Principles of Laboratory Animal Care formulated by the National Society for Medical Research and the Guide for the Care and Use of Laboratory Animals prepared by the Institute of Laboratory Animal Resources, and published by the National Institutes of Health (NIH publication No. 86-23, revised 1985).
Metabolic, hematologic, and immunologic monitoring
Fasting blood glucose was monitored using a Precision Xtra Glucometer (Abbott Laboratories, North Chicago, IL). Blood glucose control was also assessed by determination of glycosylated hemoglobin (HbA1c) using a DCA Vantage analyzer (Siemens Healthcare Diagnostics, Deerfield, IL). Total and differential WBC counts were carried out at the laboratory of the Children's Hospital of Pittsburgh using a Beckman Coulter LH780 (Beckman Coulter, Brea, CA). A diurnal variation in the white blood cell (WBC) count, especially in neutrophils, is known to occur.Citation19 To eliminate this bias, all blood samples were drawn between 8–9 a.m. In 4 of the 14 monkeys, pre- and post-STZ blood samples were analyzed for T and B lymphocyte numbers by flow cytometry. Assays for the measurement of cytokines (interleukin [IL]-6, IL-8, IL-10, tumor necrosis factor [TNF]-α, interferon [IFN]-γ and monocyte chemo-attractant protein [MCP]-1) were carried out in serum from 8 monkeys using Milliplex Map kit (Millipore, Billerica, MA) at the Luminex core facility of the Hillman Cancer Center, University of Pittsburgh Medical Center, Pittsburgh, PA.
Statistical analyses
The arithmetic mean for each parameter was calculated and statistical significance was determined by the Student t test, using Microsoft Excel 2010 (Redmond, WA).
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Funding
Work on xenotransplantation at the University of Pittsburgh has been supported in part by Department of Defense grant W81XWH-06-1-0317, JDRF grant 6-2005-1180, NIH grants #U19 AI090959-01, #U01 AI068642, and #R21 A1074844, and by Sponsored Research Agreements between Revivicor, Blacksburg, VA, and the University of Pittsburgh. This project used the Luminex Core Laboratory of the UPCI Cancer Biomarkers Facility that is supported in part by NIH grant P30CA047904.
References
- Lenzen S. The mechanisms of alloxan- and streptozotocin-induced diabetes. Diabetologia 2008; 51:216-26; PMID: 18087688; http://dx.doi.org/10.1007/s00125-007-0886-7
- Broder LE, Carter SK. Pancreatic islet cell carcinoma. II. Results of therapy with streptozotocin in 52 patients. Ann Intern Med 1973; 79:108-18; PMID: 4352784; http://dx.doi.org/10.7326/0003-4819-79-1-108
- Murray-Lyon IM, Eddleston AL, Williams R, Brown M, Hogbin BM, Bennett A, Edwards JC, Taylor KW. Treatment of multiple-hormone-producing malignant islet-cell tumour with streptozotocin. Lancet 1968; 292:895-8; http://dx.doi.org/10.1016/S0140-6736(68)91058-1
- Schein P, Kahn R, Gorden P, Wells S, Devita VT. Streptozotocin for malignant insulinomas and carcinoid tumor: report of 8 cases and review of the literature. Arch Intern Med. 1973; 132:555-61; PMID: 4355081; http://dx.doi.org/10.1001/archinte.1973.03650100069013
- Moertel CG, Reitemeier RJ, Schutt AJ, Hahn RG. Phase II study of streptozotocin (NSC-85998) in the treatment of advanced gastrointestinal cancer. Cancer Chemother Rep 1971; 55:303-7; PMID: 5115851
- Szkudelski T. The mechanism of alloxan and streptozotocin action in β-cells of the rat pancreas. Physiol Res. 2001; 50:537-46; PMID: 11829314
- Dufrane D, van Steenberghe M, Guiot Y, Goebbels RM, Saliez A, Gianello P. Streptozotocin-induced diabetes in large animals (pigs/primates): role of GLUT2 transporter and beta-cell plasticity. Transplantation 2006; 81:36-45; PMID: 16421474; http://dx.doi.org/10.1097/01.tp.0000189712.74495.82
- Graham ML, Janecek JL, Kittredge JA, Hering BJ, Schuurman HJ. The streptozotocin-induced diabetic nude mouse model: differences between animals from different sources. Comp Med. 2011; 61:356-60; PMID: 22330251
- Rood PP, Bottino R, Balamurugan AN, Smetanka C, Ezzelarab M, Busch J, Hara H, Trucco M, Cooper DK. Induction of diabetes in cynomolgus monkeys with high-dose streptozotocin - adverse effects and early responses. Pancreas 2006; 33:287-92; PMID: 17003651; http://dx.doi.org/10.1097/01.mpa.0000235307.04110.a2
- Koulmanda M, Qipo A, Chebrolu S, O’Neil J, Auchincloss H, Smith RN. The effect of low versus high dose of streptozotocin in cynomolgus monkeys (Macaca fascilularis). Am J Transplant 2003; 3:267-72; PMID: 12614280; http://dx.doi.org/10.1034/j.1600-6143.2003.00040.x
- Graham ML, Mutch LA, Kittredge JA, Rieke EF, Robinson NA, Zolondek EK, Faig AW, Dufour TA, Munson JW, Schuurman HJ. Management of adverse side-effects after chemotherapy in macaques as exemplified by streptozotocin: case studies and recommendations. Lab Anim 2012; 46:178-92; PMID: 22577153; http://dx.doi.org/10.1258/la.2012.011077
- Wu D, Zou C, Yue F, Li X, Li S, Zhang YA. The effect of long-term streptozotocin-induced diabetes mellitus (STZ-DM) on cynomolgus (Macaca Fascicularis) monkeys. J Med Primatol 2009; 38:15-22; PMID: 19018946; http://dx.doi.org/10.1111/j.1600-0684.2008.00300.x
- Nichols WK, Spellman JB, Vann LL, Daynes RA. Immune responses of diabetic animals: direct immunosuppressant effects of streptozotocin in mice. Diabetologia 1979; 16:51-7; PMID: 153868; http://dx.doi.org/10.1007/BF00423151
- Takayama Y, Ichikawa T, Maki T. Effect of STZ administration on islet isograft and allograft survival in NOD mice. Diabetes 1993; 42:324-9; PMID: 8425668; http://dx.doi.org/10.2337/diab.42.2.324
- Koulmanda M, Qipo A, Auchincloss HJ Jr, Smith RN. Effects of streptozotocin on autoimmune diabetes in NOD mice. Clin Exp Immunol 2003; 134:210-6; PMID: 14616779; http://dx.doi.org/10.1046/j.1365-2249.2003.02293.x
- Muller YD, Golshayan D, Ehirchiou D, Wyss JC, Giovannoni L, Meier R, Serre-Beinier V, Puga Yung G, Morel P, et al. Immunosuppressive effects of streptozotocin-induced diabetes result in absolute lymphopenia and a relative increase of T regulatory cells. Diabetes 2011; 60:2331-40; PMID: 21752956; http://dx.doi.org/10.2337/db11-0159
- Nichols WK, Vann LL, Spellman JB. Streptozotocin effects on T lymphocytes and bone marrow cells. Clin Exp Immunol 1981; 46:627-32; PMID: 6461461
- Levine BS, Henry MC, Port CD, Rosen E. Toxicologic evaluation of streptozotocin (NSC 85998) in mice, dogs and monkeys. Drug Chem Toxicol 1980; 3:201-12; PMID: 6452996; http://dx.doi.org/10.3109/01480548009108283
- Sennels HP, Jørgensen HL, Hansen AL, Goetze JP, Fahrenkrug J. Diurnal variation of hematology parameters in healthy young males: the Bispebjerg study of diurnal variations. Scand J Clin Lab Invest 2011; 71:532-41; PMID: 21988588; http://dx.doi.org/10.3109/00365513.2011.602422
- Marigliano M, Casu A, Bertera S, Trucco M, Bottino R. Hemoglobin A1c percentage in nonhuman primates: a useful tool to monitor diabetes before and after porcine pancreatic islet xenotransplantation. J Transplant 2011; 965605; http://dx.doi.org/10.1155/2011/965605
- Weiss RB. Streptozocin: a review of its pharmacology, efficacy, and toxicity. Cancer Treat Rep 1982; 66:427-38; PMID: 6277485
- Rakieten N, Rakieten Ml, Nadkarni MV. Studies on the diabetogenic action of streptozotocin (NSC-37917). Cancer Chemother Rep 1968; 29:91-8.
- Schnedl WJ, Ferber S, Johnson JH, Newgard CB. STZ transport and cytotoxicity. Specific enhancement in GLUT2-expressing cells. Diabetes 1994; 43:1326-33; PMID: 7926307; http://dx.doi.org/10.2337/diab.43.11.1326
- Paik SG, Fleischer N, Shin SI. Insulin-dependent diabetes mellitus induced by subdiabetogenic doses of streptozotocin: obligatory role of cell-mediated autoimmune processes. Proc Natl Acad Sci USA 1980; 77:6129-33; http://dx.doi.org/10.1073/pnas.77.10.6129
- Luo B, Chan WF, Lord SJ, Nanji SA, Rajotte RV, Shapiro AMJ, Anderson CC. Diabetes induces rapid suppression of adaptive immunity followed by homeostatic T-cell proliferation. Scand J Immunol 2007; 65:22-31; PMID: 17212763; http://dx.doi.org/10.1111/j.1365-3083.2006.01863.x
- Ishibashi T, Kitahara Y, Harada Y, Harada S, Takamoto M, Ishibashi T. Immunologic features of mice with streptozotocin-induced diabetes: depression of their immune responses to sheep red blood cells. Diabetes 1980; 29:516-23; PMID: 7380115; http://dx.doi.org/10.2337/diab.29.7.516
- Wellhausen SR. Definition of streptozocin toxicity for primary lymphoidal tissues. Diabetes 1986; 35:1404-11; PMID: 2945746; http://dx.doi.org/10.2337/diab.35.12.1404
- Deeds MC, Anderson JM, Armstrong AS, Gastineau DA, Hiddinga HJ, Jahangir A, Eberhardt NL, Kudva YC. Single dose streptozotocin induced diabetes: considerations for study design in islet transplantation models. Lab Anim 2011; 45:131-40; PMID: 21478271; http://dx.doi.org/10.1258/la.2010.010090
- Lee I, Wang L, Wells AD, Ye Q, Han R, Dorf ME, Kuziel WA, Rollins BJ, Chen L, Hancock WW. Blocking the monocyte chemoattractant protein-1/CCR2 chemokine pathway induces permanent survival of islet allografts through a programmed death-1 ligand-1-dependent mechanism. J Immunol 2003; 171:6929-35; PMID: 14662900; http://dx.doi.org/10.4049/jimmunol.171.12.6929
- Ehrnfelt C, Kumagai-Braesch M, Uzunel M, Holgersson J. Adult porcine islets produce MCP-1 and recruit human monocytes in vitro. Xenotransplantation 2004; 11:184-94; PMID: 14962280; http://dx.doi.org/10.1046/j.1399-3089.2003.00104.x
- Piemonti L, Leone BE, Nano R, Saccani A, Monti P, Maffi P, Bianchi G, Sica A, Peri G, Melzi R, et al. Human pancreatic islets produce and secrete MCP-1/CCL2: relevance in human islet transplantation. Diabetes 2002; 51:55-65; PMID: 11756323; http://dx.doi.org/10.2337/diabetes.51.1.55

