
Abstract
To incorporate metabolic, bioremedial functions into the performance of buildings and to balance generative architecture's dominant focus on computational programming and digital fabrication, this text first discusses hybridizing Maturana and Varela's biological theory of autopoiesis with Andy Clark's hypothesis of extended cognition. Doing so establishes a procedural protocol to research biological domains from which design could source data/insight from biosemiotics, sensory plants, and biocomputation. I trace computation and botanic simulations back to Alan Turing's little-known 1950s Morphogenetic drawings, reaction-diffusion algorithms, and pioneering artificial intelligence (AI) in order to establish bioarchitecture's generative point of origin. I ask provocatively, Can buildings think? as a question echoing Turing's own, "Can machines think?"
Thereafter, I assemble a research scaffold responsive to input along lines Baluška and Mancuso discuss where: “biological systems actively experience environment, both abiotic and biotic, and… memorize… the obtained information in… embodied knowledge.”Citation4 Sense-making, as in bacterial intelligence is then posited as capable of impacting generative design via plant and cellular organisms.
I do not contend that metabolic architectural intelligence will be like human cognition. Rather, I suggest that biological research involving sense-making bacteria, plants, synthetic biology, and algorithmic life define approaches that generative architecture should view for categorizing and sourcing new forms of life deployable as ecologically bioremediating, architecture-to-environment interfaces. I call the research protocol autopoietic-extended design, theorizing it as a design methodology bridging generative, bioresponsive architecture, technology, and plant science.
Hybridizing Autopoiesis + Extended Cognition
This is not a pure application of biological autopoiesis. It is a hybridized expression taking into account theoretical driftCitation64 and adaptation experienced by Humberto Maturana and Francisco Varela's 1980Citation41 publication Autopoiesis and Cognition.Citation24,27,68 My intention is to shape an autopoietic protocol based in biology and computation that brings to architectural research and experimentation core definitions of the physical and phenomenological properties required for living systems. In this pursuit, research investigating plant intelligence is significant for theory questioning types of intelligence in a context where botanic and single-cellular intelligences may be deployed to building and material performance. I outline this Autopoietic-Extended Design quest where:
Extension denotes multi-directional communication between organisms, matter, and force resolved in fields of agency, perception, cognition, and environment.
Nature is viewed as biologically/phenomenologically continuous (nondualistic) and autopoietic.
Animal architectures (including human) are classified as extended phenotypes.
The workings of cognition and environment unite, bonded by the theory of extended cognition to enable metabolic design visualization as inseparable from nature.
I situate autopoiesis in an educational scaffolding serving research in bioarchitecture directed through nature/biology/technology. The scaffold supports criteria, protocols, and methods for detecting and defining life and cognition via autopoietic precepts. To give Maturana and Varela's theory extension — reach — into the physical world, I pair it with a collaborator host. The resulting scaffolding/protocol helps users hypothesize and communicate sets of requirements, procedures, and relationships. Consequently, it aids observing phenomenological/biochemical intelligence relevant to sense-making (cellular intelligence) and bioremedial architectural tasks.
The designation of a particular order of extended cognition is based on its theorist's inclusion of unique animate, extended phenotypic, and object communication capable of factoring in cognition, non-human sense-making, design/construction, and environment. The theorist, cognitive scientist/philosopher Andy Clark explained to The New York Times:
It is possible that sometimes at least, some of the activity that enables us to be the thinking, knowing, agents that we are occurs outside the brain.... Minds like ours are the products not of neural processing alone but of the complex and iterated interplay between brains, bodies, and the many designer environments in which we increasingly live and work.Citation15
In 2008s Supersizing the Mind: Embodiment, Action, and Cognitive Extension, Clark wrote:
It matters that we recognize the very large extent to which individual human thought and reason are not activities that occur solely in the brain… it drives home the degree to which environmental engineering is also self-engineering. In building our physical and social worlds, we build (or rather, we massively reconfigure) our minds and our capacities of thought and reason.Citation14
Autopoiesis is more problematic to encapsulate than extended cognition because of its evolved and evolving ontology, its various adaptations, and its wide-scope scholarly interpretations. I position it here in the following 5 linked quotations:
[Autopoiesis is] a singularity among self-organizing concepts… provid[ing] the decisive entry point into the origin of individuality and identity… connecting… into the phenomenological realm.Citation68
This positioning is premised on theoretical adaptability where:
… autopoietic theory moves from being [solely] a theory of (all) the living to being a moment that allows us to grasp the phenomena of life and mind.”Citation26
More broadly documented:
Autopoiesis (from Greek αὐτo- (auto-), meaning “self,” and πoίησις (poiesis), meaning “creation, production”) literally means “self-creation” and expresses a fundamental dialectic among structure, mechanism, and function. The term was introduced in 1972 by Chilean biologists Humberto Maturana and Francisco Varela.Citation69
In Life in Mind, Evan ThompsonCitation49 gave the theory greater precision while maintaining original traits:
Autopoietic systems form a subset of self-organizing systems… interact[ing] with each other in non-linear ways to produce the emergence and maintenance of [their] structured global order…. [where] constituent processes “(i) recursively depend on each other for their generation and their realization as a network, (ii) constitute the system as a unity in whatever domain they exist, and (iii) determine a domain of possible interactions with the environment.”Citation55
From Thompson I turn to Randall D. Beer because he, like Ezequiel Di Paolo,Citation24,26 recognized that autopoiesis must not only evolve, but must accommodate adaptations, including algorithmic programming, to be relevant in biodigital and phenomenological environments. Beer's paper, “Autopoiesis and Cognition in the Game of Life” begins:
Autopoiesis is a network of component-producing processes with the property that the interactions between the components generate the very same network of processes that produce them, as well as constituting it as a distinct entity in the space in which it exists. The paradigmatic example of autopoiesis is a cell, in which the components are molecules, the interactions are chemical reactions and the cell membrane serves as a physical boundary that spatially localizes these reactions into an entity (or “unity”) distinguishable from its environment…. Maturana and Varela offer a view of life as a specific organization of physical processes that has as its principal product the maintenance of its own organization.Citation7
My analysis postulates an autopoietic presence in architectural learning and research — predominantly digital and biogenerative — mediated by science, computation, and technology and sourced in nature. I conclude that such a role, instantiated by researchers and designers as thinking agents, is inseparable from the “reality of the external world, [and is] hence part of nature,” to use Weber and Varela'sCitation68 construction of humans vis-à-vis nature. Georges Canguilhem came to a similar conclusion supported by Descartes:
[W]e arrive at the point where the machine is seen as a fact of culture, expressed in mechanisms that are themselves nothing more than an explainable fact of nature. In a celebrated text in “Principles of Philosophy,” Descartes writes, “It is certain that all the rules of mechanics belong to physics, to the extent that all artificial things are thereby natural...” [T]hey are thus the direct or indirect products of a technical activity that is as authentically organic as the flowering of trees.Citation11
By having a theoretic account of life/cognition in autopoiesis, developments in fields such as synthetic biologyCitation66,67 and plant neurobiologyCitation4,9,30,38,48,56 may be interrelated, sometimes revealing “hidden potentialities that remained unexpressed in the theory of autopoiesis: the possibility of adaptive autopoiesis.”Citation26 So too, “hidden potentialities” wait in plant signaling and living technologyCitation5,6 conceivably applicable to autopoiesis. Artificial Life's characterization of living technology, in such a case, is:
… based on the powerful core features of life… explained and illustrated with examples from artificial life software, reconfigurable and evolvable hardware, autonomously self-reproducing robots, chemical protocells, and hybrid electronic-chemical systems.Citation5
Those features, along with existing technological and computational systems — AI and Alife (AL) simulations, for example — contribute cultural and scientific data able to inflect autopoietic logic and rule-sets for plotting: Can buildings think?
Evolving Turing's “Can machines think?”
The seemingly banal question, Can buildings think? riffs Alan Turing's 1950 “Can machines think?”Citation58 framing his pursuit of machine intelligence (AI),Citation58 biochemical symmetry-breaking,Citation59 and botanic and embryonic simulation.Citation29,50,60 Yet, before impacting generative architecture, Turing's inquests enabled Chomsky'sCitation12,29 generative grammars and subsequently, Turing's equations and Chomsky's theory contributed to Lindenmayer'sCitation29,37 L-systemsCitation23,29,50 for coding digital organisms. Here, I venture that organic and digital:
Organisms are a wave of matter and energy, they are bound by the laws of physics but not fully determined by them as their destiny is not attached to any particular material configuration but they ride from one configuration to another.Citation27
In this lineage, aligning Maturana and Varela with Turing may be defended when Turing's botanic drawings,Citation28 reaction-diffusion calculations,Citation49 and Richards'sCitation51 embryological (diatom) simulations are seen as ancestry for Maturana, Varela, and Uribe's cellular automata (CA).Citation25,62 Their CA prototype was programmed after Conway's Game of LifeCitation8,Citation31 to demonstrate digital autopoiesis in a rule-based, computational environment.Citation62,63 In each, we encounter relevant overlap: theoretical biology-to-autopoietic AL from Maturana, Varela, and Uribe — and observation of flowers/fircones for drawings, calculations, and simulations from Turing.Citation28,51,53,54
Turing's biocomputational plant and embryological research,Citation53,54 what he called Morphogenesis,Citation59,60 is frequently overshadowed by his universal machine,Citation57 his WWII role at Bletchley Park deciphering Enigma,Citation18 and the Turing Test.Citation58 Therefore it may be helpful to note that from the late 1940s to his death in 1954 he concentrated on biological input for algorithmic formulation.Citation28,59,60 John Reinitz wrote in Nature that the importance of Turing's revolutionary paper, “The Chemical Basis of Morphogenesis” (1952) is:
… difficult to overstate. It was a transition point form the era of analytical mathematics to that of computational mathematics…. Turing's paper contains the first computer simulation of [plant and animal] pattern formation… and is possibly the first openly published case of computational experimentation.Citation50
Nobel Prize laureate, Sidney Brenner came to a similar conclusion:
“The Chemical Basis of Morphogenesis” explored the hypothesis that patterns are generated in plants and animals by “chemical substances called morphogens, reacting together and diffusing through a tissue.” Using differential equations Turing set out how instabilities in a homogeneous medium could produce wave patterns that might account for the processes such as the segregation of tissue types in the developing embryo.Citation10
Computational life/intelligence as sparked in programming and scripting, in conjunction with observational tactics like those Turing used to draw and calculate plants and nature, here anchors the specialized origin point I assign to biogenerative architecture.Citation28,Citation57-60 That origin is mapped through autopoietic-extended design. Its parameters establish the route for a biology-driven, computationally coded, and metabolically expressed architectural lineage.Citation50 Maturana and Varela'sCitation41 theory now critically articulates a means for projecting future-possible metabolic/intelligent objects and buildingsCitation24-26 consistent with that lineage.
This ancestry, understood in the context of cognition-to-environmentCitation13-16 generation, underwrites architects' contemplating design sense-making for autonomousCitation22a buildings. Di Paolo writes, “sense-making is the engagement of a cognitive system with its world in terms of significance or value.”Citation26 Elsewhere, he contends, “sense-making of living systems… [I]s meaningful in terms of its consequences for the conservation of a way of life. In the living system this is ultimately the conservation of its autopoietic organization, its own survival and viability.”Citation27 For plants, and I think for future buildings, we may ask “Do capabilities such as intelligence, pain perception, learning, and memory require the existence of a brain.. ?”Citation48 Hereafter, although Maturana and Varela provided no formulation for constructing intelligent life (beyond their CA.Citation62,70 See also McMulling and Varela's 1997 recreationCitation44), they offered repeated encouragement for its meta-conceptualization:
Machines are generally viewed as human made artifacts with completely known deterministic properties which make them, at least conceptually, perfectly predictable. Contrariwise, living systems are a priori frequently viewed as autonomous, ultimately unpredictable systems, with purposeful behavior similar to ours. If living systems were machines, they could be made by man and, according to the view mentioned above, it seems unbelievable that man could manufacture a living system. This view can be easily disqualified, because it either implies the belief that living systems cannot be understood because they are too complex for our meager intellect and will remain so, or that the principles which generate them are intrinsically unknowable; either implication would have to be accepted a priori without proper demonstration.Citation41
Population-Based Adaptation and BioDigital Models
Eighteen years after Maturana and Varela's above text, Christopher Langton discussed the animation of machines. I find his AI/AL focus compatible with autopoieitc-extended design through his account of ground-up self-organization:
… living organism are nothing more than complex biochemical machines. However, they are different from the machines of our everyday experience. A living organism is not a single, complicated biochemical machine. Rather it must be viewed as a large population of relatively simple machines. The complexity of its behavior is due to the highly nonlinear nature of the interactions between all of the members of this polymorphic population. To animate machines, therefore, is not to “bring” life to a machine; rather it is to organize a population of machines in such a way that their interactive dynamic is “alive.”Citation36
From Langton'sCitation36 population-based proposition, I entrust Maturana and Varela'sCitation41 theory of life and cognition as it factors in aggregate populations. In practice here, autopoietic components and unities fill the job Langton gave to aggregations. Once identified, component populations may be extrapolated from for attributes to “organize a population of machines”Citation36 (or rule-sets) composed through direct and/or technologically enhanced agent observation and prediction. To do this, mechanisms from autopoiesis involve components, unities, domains, structural coupling, border creation, and operational closure.Citation55 Concatenated, those workings are recursively generative within or around autopoietic-extended design's protocol fostering research and highlighting structural coupling of agents, technology/objects and nature.Citation29
New conceptualizations of biochemical memory and plant intelligenceCitation1,4,9,38,48 hint at sense-making options for bioremedial building infrastructure and architectural performance. I speculate that materials and agent populations conceptualized through autopoietic-extended design are to be recognized first in biology, then (via Turing), expressed as digital code amenable to bioprinting and machine fabrication not currently found in design fablabs, but existing in medical, laserjet-printed skin and 3D bioprinted organs, kidneys for example.Citation3,35,45
In, I have fabricated a demonstration model for which anticipatory and reactive botanic metabolism, intelligence, and morphology could be investigated to generate ideas, methodology, and prototypes as components for intelligent buildings. This is the heart of the autopoietic-extended design protocol — the integration of smart, non-polluting, bioreactive sensing and response in architecture. The model becomes a physical manifestation of the autopoietic-extended design scaffold that generated it. In this way, it carries embedded data as a cognitive-state object giving it potential to join next phase experiments.
Figure 1. Shape-Shifting e-Tree. Dennis Dollens. Digitally hybridized tree simulation generated in Xfrog (L-systems), exported to Rhino, and built as an STL model. The structure tests computationally evolved tree branches intersecting each other to create an abstract, trusslike component, capable of varying degrees of environmentally responsive bending and twisting. Xfrog/Rhino/3DS Max.

As presented, the stereolithographic (STL) model was 3D printed from a file using L-systemsCitation37 (Xfrog software) to digitally simulate tree branching by revisualizing and drastically manipulating phyllotaxis, parastichy, gravitropism, and phototropism. The model's branching then displays alternative anatomical performance delivered in mutual self-bracing structures I call eTrees.
The eTrees' differing configurations populate a new typology of trusslike, digitally simulated structures, matrices, substrates, and armatures. At present, the eTree exhibits a limited range of performative abilities such as flex, stretch, and twist intended (at the next stage), to engage sensed aerodynamic, seismic, and tropic input/output responses. The properties are clearly not foreign to plants or animals, but animate sensing and responsiveness — biointelligence and biomotor skills for buildings — are only now coming into focus for the models. As the physical expressions of an idea, the STL armatures require astuteness — self-maintaining abilities — that include recognition and anticipation.Citation13-16,Citation24-27
To go beyond biomimetics and toward living, self-maintaining, sense-making organisms, is the goal. To do that, I contemplate replacing the current model's material with a living, 3D printable bioresin or, secondly, hybridizing a cellular-intelligent system into the current L-systems' skeletal matrix. In, the second option, involving a cellular-intelligent, membrane-clad structure is visualized with leaflike performance. These are working, concept models in primary stages of process/structure integration. For architectural tasks they initiate discussion and illustrate research direction — predicated for example, on plant or bacterial sense-making and response. In a research practice the models necessitate fresh relationships through which design accesses new typologies not only redeploying life and intelligenceCitation4,9,19,38 but begin an ontological protocol to deal with dwelling in an environmental crisis.
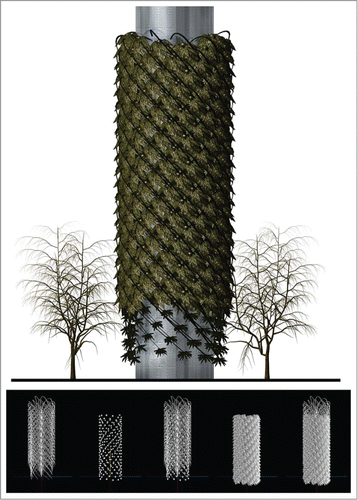
Figure 2. L-Systems and Generative BioAlgorithmic Structure. BioTower. Dennis Dollens. Hypothetical performative leaf movement activated by metabolic controllers for plantlike filtration and sensor/monitor bioremedial systems. Multiple Xfrog-grown e-trees (bottom) illustrating, from left to right: 1) intersecting structural branching (truss), 2) housing for metabolic controllers (pods), 3) branching and pods, 4) leaves, and 5) assembled components. Xfrog/Rhino/3DS Max.
Evolving Autopoiesis
Within this quest, it is useful to understand that autopoiesis evolves as life and intelligence expand and evolve. This process began almost immediatelyCitation65 (note 1) after publication in Autopoiesis and Cognition: The Realization of the Living.Citation41 Di PaoloCitation24,26 proposes that “the theory of autopoiesis in itself should be critically assessed and, if necessary, reinterpreted or extended.”Citation25 And, it is Di Paolo's adaptations I trigger, in conjunction with those of Weber and Varela.Citation68 Once reinterpreted, autopoiesis (in relation to self-organization, adaptation, and extended cognition) helps order living properties for tactical observation of components essential in living organisms/environments. Relational living systems, comprised of or by components, unities, structural coupling, and operational closure then sustain autopoieticCitation41 domains to register member behavior appropriate to the construction of rule-sets.
Configured, autopoietic-extended design supports researchers by aiding ontological procedures to codify ways of being (goal-seeking behavior, identity generation, perception) biotechnologically engaging recognition, pairing, and prediction.Citation16 Recognition, pairing, prediction, with visualization consequently surrounds autopoietic components, unities, and domains as they engage and bond agency + nature + technology. Prediction, now driving segments of foresight and visualization in design is configured for autopoietic-extended design through procedures ClarkCitation13–16 hypothesizes in extended cognition. In relation, I match Clark'sCitation16 prediction to questions Di PaoloCitation26 forefronts when he asks “How can we invest artifacts with a… form of sense-making.”Citation27 Significantly, Maturana and VarelaCitation41 identified predictive mechanisms in play early on for autopoiesis:
A prediction is a statement of a case within a relational matrix; it is a cognitive statement, and as such it takes place within a descriptive domain. Thus, unless, mistakes are made, if all the relations that define the particular matrix within which the prediction is made are properly taken, the prediction is valid. Errors of interpretation may arise only by misapplication, that is, by pretending that the observer makes a prediction in one matrix when he is making it in another. In particular, predictions in the physical space are possible, because a description, as an actual behavior, exists in a matrix of interactions which (by constitution) has a logical matrix necessarily isomorphic with the substratum matrix within which it takes place, not because we have an absolute knowledge of the universe.Citation41
Metaphorically, autopoietic-extended design convenes a cognition-to-practice ecotone — a domain hosting one-off forms/species of intelligence potentially nurturing evolution for a metabolic species of architecture existing nowhere else. Those cultivated requisites sustain designer/researcher subjectively deploying observation, search, monitoring, and identification in service to physical responses synchronized with environments, organisms, objects, data, and/or matter.Citation29 Animate properties, living organization, and morphology subsequently produce feedback for interpretation — witnessed early on, for example, in Turing's research drawings. Today, those drawings ontologically illustrate thought evolved from observation to further understand/express intelligences found in nature and extend those findings to visualizations, code, and computer simulationsCitation28 ( and ).
Flower-to-Bee-to-Building
For an example of data transference, Dominic Clarke et al.Citation17 have identified, “floral electrical-fields” used by bees and flowers to communicate. Their research, announced in Science,Citation17 documents localized interspecies messaging with chemical and electrical signals and complements other research in plant signaling and behavior.Citation9,38,43 While years away from practical development, animal/plant signal decoding and biochemical plant memory,Citation4,9,17,38,43,46,48 — do, here-and-now, illustrate experimental routes compatible with technology, computation, and bioremediation.
Plant signalingCitation17 and modes of biochemical communication and memory researched by Baluška and Mancurso,Citation4 Brenner et al.,Citation9 Gagliano et al.,Citation30 Mousavi et al.,Citation40 and TrewavasCitation56 are also channels for studying and adapting intelligence sequestered in plant/bacteria sensing, signaling, and performance.Citation4,9,46 For design research, plant sensing/signaling opens envisioning consistent with AI and metabolic (but not brain-based) intelligence, discussed here as it edges toward bioresponsive buildings.Citation5,6 — How, you may wonder?
Clarke et al.'sCitation17 bee-to-flower/flower-to-bee signaling is a case-in-point. The team's research could be narrated as: cognitive social organism (bee), communicates with animate (but differently or non-human) intelligent plant/animal. Here, intelligence in plants is defined by Brenner et al.Citation9 as “an intrinsic ability to process information from both abiotic and biotic stimuli that allows optimal decisions about future activities in a given environment.” Similarly, Anthony Trewavas wrote the hallmark of plant intelligence is “variable growth and development in the lifetime of the individual…[requiring an] organism able to compute and the right environmental circumstances to elicit that computation.”Citation56
The bee-to-flower narrative could be rewritten with a building occupying the role of the flower and programmed agents — animats, biorobots, or biomaterial agents standing in the role of the bees. In this scenario, with a building as living organisms, we may contemplate a near-field (NFC) communications network with biochemical/electrochemical receptors/actuators incorporating intelligence and goals that position architecture as a subset of extended phenotypes.Citation20,34 Hereby, autopoietic-extended design emerges as a scaffold and protocol for research. Such a scenario opens theoretical as well as bioremedial performance to integrate ecology and plant intelligence with systems, communication, technology, and nature.Citation14,17,27,41,68
Intelligent Objects, Extracranial Environment, Extended Phenotypes
Chris Gosden set an adaptive course in the context of archeology when he wrote: “the house is an intelligent object.”Citation33 As a synonym for architecture, I evolve the designation of house as intelligent. By extending his proto-autopoietic insight to dwelling/shelter/house/tool I identify future buildings as candidates for construction behavior partnered by intelligencesCitation36 and biotechnologically coupled with nature, (or even as), organisms.Citation25 Gosden's assertion does not cite autopoiesis, but it references extended cognition in the framework of “The Cognitive Life of Things: Recasting the Boundaries of Mind”Citation33 to which Clark also contributed. From Gosden's text, a house/dwelling is a cognitive-state object participating here within the boundaries of autopoiesis as an extended phenotype.Citation20,34 Mike Hansell, calling to account “all construction behavior,”Citation34 opens a conceptual conduit to extended phenotypes in Animal Architecture:
… different senses in which the term tool is applied in human and animal biology, makes a distinction which incidentally but rightly includes all construction behavior. The distinction is between constructions that conform to the Dawkins (1982) concept of extended phenotype (a product of the animal's genotype, externally expressed), and those that result from a mental plan.Citation34
Hansell makes a further important point since “genotype, externally expressed,” by necessity (to at least some degree), involves “a mental plan”Citation34 guiding agency, construction behavior, material, and site/environment implementation. Still, in a recent email exchange Hansell stressed “that, whereas the concept of extended phenotypes has a relevance to discussions on human constructions, that relevance needs to be heavily quailfied.”Citation34a
Gosden argued that extended cognition was receptive to intelligence arising in things and objects:
Current attempts to rethink mind have extended the spatial metaphor, so that intelligence arises in and through things as well as people…. These ideas see mind as not located in our heads, but something which comes about through the interactions of the whole human organism with its world, so that intelligence resides in action as much as thought and in the social use people make of the object world.Citation33
Thinking “the house is an intelligent object,”Citation33 in relation to generative architecture and plant sense-making, I contemplate differently hybridizing matter and intelligence for eventual material/building resolution. This controversial proposition requires additional research and specialized categorization of electrochemical, biochemical, and phenomenology of mind, memory, and intelligence as elemental forces of nature on the order argued by Thomas Nagel in Mind and Cosmos.Citation47 Nagel's view of thought/intelligence, phenomenologically conceived as a force of nature, seems to me an enabling view tacitly backing research into plant sense-making as parallel, perhaps equivalent, to neurologically enabled intelligence. To this point: Stefano Mancuso was quoted in a New Yorker profile titled “Plant Intelligence”Citation48 saying “neurons perhaps are overrated.... They're really just excitable cells.”
In the framework of biological research considering sensory ecology, signaling, biosemiotics, cognition, and plant neurobiology,Citation48 I detect an opportunity to conjure dialectical propositions of intelligence(s)-in-the-environmentCitation47 suggesting new paradigms of architecture, technology, science, and nature without dualistic interference. What I mean by working propositions are conceptualizations, experiments, and models participating in translated sense-making from biological sensors, communicators, and actuators setting off biological responses in a species of architectural intelligence.Citation64 At the far end of such a research spectrum, translating sense-making to buildings, will likely result in some of these processes entering the realm of living technologyCitation5,6 “where a living system is also potentially a sense-maker.”Citation27 And, from living technologyCitation5,6 I want to call attention to Leroy Cronin's ongoing (and I believe compatible) protocell research where:
Buildings would have a cellular structure with living inorganic components that would allow the entire structure to self-repair, to sense environmental changes, establish a central nervous system, and even use the environment to sequester water, develop solar energy systems, and regulate the atmosphere.Citation19
For autopoietic-extended design, functions understood as sensed and bioresponsive might include networked communications, channeled photosynthesis, organic (water/air/noise/light) filtration, aerodynamic shape shifting, resource management, urban signaling, and toxic biochemical cleanup; tasks responsible beyond today's appliqué of greenwalls and greenroofs. And, for clarity, I'm not talking about buildings that look like plants or trees, I contemplate buildings ( and ) that mobilize and metabolize responses to their environment that live like plants, trees, or bacteria.
Biologically re-coding buildings from within nature, as part of nature — autopoiesis, extended cognition, and metabolic performance signals drive, action, response functions to prototype into living structures. Intelligence as sense-making hybridized in materials and structures mingles nature and buildings when envisioned as physiological and environmental monitors programmed to remediate toxins. I point out that monitor/mediator roles are native for many biochemical plant processes. Observed plant/organism responses may then result in materials and structures that inform building performance with native plant logic.Citation64 That logic, identified by first consulting plant signaling and behavior as data, is similar to Turing's botanic and embryological decoding for biosimulations.Citation57–60 HansellCitation34 (above) and ClarkCitation13 (below) chart related aspects of cognition-to-nature-to-technology that interface designer agency with nature. I have followed their work to substantially — but nowhere near exclusively — conceptualize and visualize tools, plans, materials, actions, and metabolic architectures as extended phenotypes, keeping in mind:
[The] power and beauty of the brain's role is that it acts as a mediating factor in a variety of complex and iterated processes which continually loop between brain, body, and technological environment. And it is this larger system which solves… problem[s]. We thus confront the cognitive equivalent of Dawkins's (1982) vision of the extended phenotype.Citation13
Through the above “mediating factor,”Citation13 Clark looks to environments and objects as co-participants in thinking and doing — what I've called cognitive-state objects.Citation29 Thereby, he allows for distributed (in the manner of Langton above) environmental intelligence to validate pedagogical sets of rules guiding in-field observation and research.
Machinic Phylum holds the Conclusion
Design equations, arising through autopoiesis,Citation41 living technology,Citation5,6 and sense-making,Citation24,26 do not challenge or mimic human intelligence — however, they overwrite Maturana and Varela'sCitation41 denial of teleology in autopoiesis. As adaptive,Citation24 teleology, enables biological urges, drive, habits, responses, and decisionsCitation2,19,27,52 — it is therefore considered here “grounding teleology,”Citation27 — a driving force in metabolic systems. Di PaoloCitation27 has herewith outlined a startling potential:
[N]ew lifeforms may relate to the metabolic substrate and other lifeforms in a variety of ways, calling for a veritable topology of processes of identity generation (intersecting, embedded, hierarchical, shared, etc.). It is also an open possibility that the dependence on a form of life so much modifies the basic autonomy of metabolism that the higher identity essentially intervenes in the very condition of operational closure of autopoiesis.Citation27
It will probably not come as a surprise that I imagine Di Paolo's “new lifeforms”Citation27 to include bioremediating architecture. In this framework, and with human constructions including architectures classified as extended pheno- types,Citation20,34 intelligent architectures are ultimately and only ecologically resolvable in an equation such as:
architecture=nature
To this end, I locate Di Paolo's definition of cognition — subjectively “brought forth”Citation25 or constructed — as compatible with metabolic architectural potential:
… an embodied engagement in which the world is brought forth by the coherent activity of a cognizer in its environment…. this engagement involves the structuring of the immediate milieu with the consequent building of regularities, which feed back to the organism itself.Citation25
Di Paolo's idea of “embodied engagement” is utilized for architecture, so that, through autopoietic-extended design, the “cognizer” will be a structure, material, object, building, or society of buildings based outside of human thinking, yet modeled on, and incorporating, biological intelligence.
I figure, that in an age where machines have driftedCitation42 toward artificial lifeCitation5,6,19 and have begun to learn and modify themselves and their networks (e.g. neuromorphic chipsCitation39), that research and pedagogical scaffolding extending ideas of life and cognition be adopted. Such a scaffold needs to brace research between science, philosophy, and technology for contingent metabolic architectures. Unfolding intelligent architecture from sense-making in bacteria or plants — for example, within Clark'sCitation13–16 powerful siting of cognition partially taking place “out of our brains” — is a shift countering dualism.Citation32 The opposition corrects what Thompson described as, “the dualistic separation of consciousness and life [that] makes it impossible to understand coconsciousness in its basic form …”Citation55 But, furthermore, by implication, dualistic separations perpetuate the impossibility of considering plants as intelligent at all.
Before closing, I reference Deleuze and Guattari's notion of machinic phylum.Citation22 Machinic phylum, configured as a toolbox housing intelligence, morphology, and metabolism provisions retooling and then reorienting the trail left by Turing's universal machines,Citation57 algorithms/drawings,Citation28 and reaction-diffusion equations and simulations.Citation59,60 Deleuze and Guattari write:
… the machinic phylum is materiality, natural or artificial, and both simultaneously; it is matter in movement, in flux, in variation, matter as conveyor of singularities and traits of expression.Citation22
De Landa sees it as an:
… abstract reservoir of machinelike solutions, common to physical systems diverse as clouds, flames, rivers and even the phylogenetic lineages of living creatures… called the “machinic phylum” — a term that would indicate how nonlinear flows of matter and energy spontaneously generate machinelike assemblages.Citation21
Elsewhere I discuss the machinic phylum accommodating organisms and varieties of intelligence and sense-making.Citation29 Here I stress that the analogy between Turing's, can machines think? and my, can buildings think? requires that they be considered parallel. Both questions are occupants in human-built domains as thoughts — cognitive and/or linguistic extensions leading to physical sheltering tools (huts-to-buildings for example) that ultimately transforms (if nothing more than conceptually at first) nature from a domain of raw materials to a participatory domain of intelligence. In that domain of intelligence architecture and technology are manifested as qualified, atypical,Citation34a extended phenotypes.Citation20,34,34a,Citation61
By referencing a machinic phylum housing “machinelike assemblages”Citation21 (buildings) on the way to metabolic intelligence, we acknowledge a category in nature different from machines such as self-driving cars or attack drones. Neither self-driving cars nor drones strive toward synthesis with nature or bioremedial autonomy. It then seems to me, we may contemplate our largest built machines (architectures) in Canguilhem's words as “an explainable fact of nature”Citation11 moving toward explanations found in extended phenotypes and metabolic integration with nature to ask: can buildings think? From that query, a protocol is evolving we as animals build, extending our thoughts as inseparable from nature.Citation13-16,Citation24-27,Citation68 This autopoietic-extended design context of theory, technology, biology, and agency therein sets up 2 dialectical stages where:
Stage 1) Establishes processes for identifying and then translating metabolic and intelligent functions from nature in order to replicate desirable traits as elements of generative computational-to-bioperformative architecture. (How the building should act, what it should do.) At this stage, decisions are made concerning how, why, and in what configuration, AI,Citation36 synthetic biology,Citation67 plant signaling,Citation4,9,17,30 and/or living technologyCitation5,6 will be introduced into research and development.
Stage 2) Positions generative architecture in a research/education context around autopoiesis, extended cognition, technology, algorithmic computation, and biological systems related through Turing's morphogenetic drawings,Citation59 digital simulations,Citation60 and computational programming. This lineage is then foundational for implementing and prototyping ideas and processes (Stage 1) as theory, models, materials, and simulations.
Coordinating stages 1 and 2 is an exercise in hybridizing biological theory,Citation41 cognitive science,Citation14 and holdings in autopoietic-extended design's machinic phylum.Citation21,22,29 Once established, the dialectical interface aids orienting research and extending morphology, metabolism, and biology as components of generative architecture. In this scenario, architecture is argued as a human extended phenotypeCitation20,34,34a with the understanding that technology represented, for example, by AI and sensors, is approaching a point to be matched or bypassed for use in experimental technology and architecture by animate, sense-making organisms currently researched and theorized via plant intelligence, signaling, and behavior.Citation4,9,17,19,38,43,46,48
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Note
1. Even when conflicting publication dates suggest otherwise, the English-language publication of Autopoiesis and Cognition: The Realization of the Living is the version of record. Maturana and Varela's original only found a publisher 7-years after the publication in Chile of Carmen Cienfuegos's 1973 Spanish-language translation, De Máquinas y Seres Vivos: Una teoría de al organizatión biológica.Citation40,65
Bibliography
- Adamatzky, Andrew. (2013) “Slimeware: Engineering Devices with Slime Mold.” Artificial Life. Nineteen. 317-30; PMID:23834592; http://dx.doi.org/10.1162/ARTL_a_00110
- Armstrong, Rachel. (2012) Living Architecture: How Synthetic Biology Can Remake Our Cities and Reshape Our Lives. (Kindle Single). Amazon Single. New York. TED Books.
- Atala, Anthony. (2011) “Printing a Human Kidney.” TED. Accessed: 23 2014. www.ted.com/talks/anthony_atala_printing_a_human_kidney.
- Baluška, František. Mancuso, Stefano. (2009) “Deep evolutionary origins of neurobiology.” Communicative & Integrative Biology. Two: 1, 60-5; 2009; http://dx.doi.org/10.4161/cib.2.1.7620
- Bedau, Mark A, McCaskill, John S, Packard, Norman H, Rasmussen, Steen. (2010) “Living Technology: Exploiting Life's Principles in Technology.” Artificial Life. Cambridge, MA. The MIT Press. Sixteen:1. 89-97. Accessed: 2012. http://www.mitpressjournals.org/toc/artl/16/1.
- Bedau, Mark A. McCaskill, John S. Packard, Norman H. Rasmussen, Steen. (2013) “Introduction to Recent Developments in Technology.” Artificial Life. Cambridge, MA. The MIT Press. Nineteen. 291-8. Accessed: 2013. http://www.mitpressjournals.org/toc/artl/19/3_4.
- Beer, Randall D. (2004) “Autopoiesis and Cognition in the Game of Life.” Artificial Life. Ten: 309-26; PMID:15245630; http://dx.doi.org/10.1162/1064546041255539
- Beer, Randall D. (2014) “The Cognitive Domain of a Glider in the Game of Life.” Artificial Life. Twenty: 183-206; PMID:24494612; http://dx.doi.org/10.1162/ARTL_a_00125
- Brenner, Eric D. Stahlberg, Rainer. Mancuso, Stefano. Vivanco, Jorge. Baluška, František. van Volkenburgh, Elizabeth. (2006) “Plant Neurobiology: An Integrated View of Plant Signaling.” Trend Plant Sci. Eleven:8. 413-9. Accessed 15 February 2014; PMID:16843034; http://dx.doi.org/10.1016/j.tplants.2006.06.009
- Brenner, Sydney. (2012) “Life's Code Script.” Nature. 482. 461.23 2012
- Canguilhem, Georges. Cohen, Mark. Cherry, Randall. Tr. (1992) “Machine and Organism.” 44-69. In: Crary, Jonathan & Kwinter, Stanford. Eds. Incorporations. New York. Zone.
- Chomsky, Noam. (1959) “On Certain Formal Properties of Grammars.” Information and Control. 2. 137-67; http://dx.doi.org/10.1016/S0019-9958(59)90362-6
- Clark, Andy. (2001) “Reasons, Robots, and the Extended Mind: Rationality for the New Millennium.” Mind and Language. 16: 2. 121-45; ; http://dx.doi.org/10.1111/1468-0017.00162
- Clark, Andy. (2008) Supersizing the Mind: Embodiment, Action, and Cognitive Extension. New York. Oxford University Press.
- Clark, Andy. (2010) “Out of Our Brains.” The New York Times. New York. 12 December 2010. http://opinionator.blogs.nytimes.com/2010/12/12/out-of-our-brains/.
- Clark, Andy. (2013) “Whatever Next? Predictive Brains, Situated Agents, and the Future of Cognitive Science.” Behavioral and Brain Sciences. 36:03. 181-253; PMID:23663408; http://dx.doi.org/10.1017/S0140525X12000477
- Clarke, Dominic. Whitney, Heather. Sutton, Gregory. Robert, Daniel. (2013) “Detection and Learning of Floral Electric Fields by Bumblebees.” Science Magazine. . 21 February 2013. Accessed: 25 2013. http://www.sciencemag.org/content/early/recent; PMID:23429701
- Copeland, Jack B. (2012) Turing: Pioneer of the Information Age. Oxford. Oxford University Press.
- Cronin, Leroy. (2011) “Defining New Architectural Design Principles With ‘Living’ Inorganic Materials.” In: Spiller, Neil. & Armstrong, Rachel. Eds. (2011) “Protocell Architecture.” AD: Architectural Design. Wiley. 81:2. 34-43. 2011.
- Dawkins, Richard. (1982) The Extended Phenotype: The Long Reach of the Gene. New York. Oxford University Press.
- De Landa, Manuel. (1992) “Nonorganic Life.” 128-167. In: Crary, Jonathan. & Kwinter, Stanford. Eds. Incorporations. New York. Zone.
- Deleuze, Gilles. Guattari, Félix. Massumi, Brian. Tr. (1987) A Thousand Plateaus: Capitalism and Schizophrenia. Minneapolis. University of Minnesota Press.
- Dennett, Daniel C. (1998) Brainchildren: Essays on Designing Minds. London. Penguin Books.
- Di Paolo, Ezequiel. (2005) “Autopoiesis, Adaptivity, Teleology, Agency.” Phenomenology and the Cognitive Sciences. Four: 42952. DOI: 10.1007/s11097-005-9002-y C _ Springer 2005.
- Di Paolo, Ezequiel. (2008) “Extended Life." Topoi. 28. 9-21. Downloaded author's website: 20 April 2013. http://ezequieldipaolo.wordpress.com/publications/journal-papers/
- Di Paolo, Ezequiel. (2009) “Overcoming Autopoiesis: An Enactive Detour on the Way from Life to Society. In: Olivas-Lujan, Miguel R. & Bondarouk, Tanya. Eds. (2009) Autopoiesis in Organization Theory and Practice, V6. Bingley, UK. . 6. 43-68.
- Di Paolo, Ezequiel. (2010) “Robotics Inspired in the Organism." Intellectia. 2010 1-2:53-54. 129-162
- Dollens, Dennis. (2014) “Alan Turing's Drawings, Autopoiesis and Can Buildings Think.” Leonardo: The International Society for the Arts, Sciences and Technology. Cambridge, MA. The MIT Press. 47:3. 249-53. Abstract: http://www.mitpressjournals.org/doi/abs/10.1162/LEON_a_00766#.VBunXEi3Ans.
- Dollens, Dennis. (2014a) “Autopoietic-Extended Architecture: Can Buildings Think?” PhD Diss. Edinburgh School of Architecture and Landscape Architecture. University of Edinburgh.
- Gagliano, Monica. Renton, Michael. Depczynski, Martial. Mancuso, Stefano. (2014) “Experience Teaches Plants to Learn Faster and Learn Slower in Environments Where it Matters.” Oecologia. 5 2014; PMID:24390479.
- Gardner, Martin. (1970) “Mathematical Games.” Scientific American. 1970. 223. 120-3; http://dx.doi.org/10.1038/scientificamerican1070-120
- Gibson, James J. (1986/1979) The Ecological Approach to Visual Perception. Hillsdale, New Jersey. Lawrence Erlbaum Associates. Publishers.
- Gosden, Chris. (2010) “The Death of the Mind.” p39-46. In: Malafouris, Lambros. & Renfrew, Colin. 2010. The Cognitive Life of Things: Recasting the Boundaries of the Mind. Cambridge. McDonald Institute for Archaeological Research. University of Cambridge.
- Hansell, Mike. (2005) Animal Architecture. Oxford. Oxford University Press.
- Hansell, Mike. (2014) Email to Dennis Dollens. 9 October 2014.
- Jorgensen, Ellen. (2012) “Biohacking — You Can Do It Too.” TED Global. Accessed: 2 2014. http://www.ted.com/talks/ellen_jorgensen_biohacking_you_can_do_it_too.
- Langton, Christopher G. (1988) “Artificial Life." In: Langton, Christopher G. (1988) Artificial Life. Santa Fe, NM. Addison Wesley & The Santa Fe Institute. 6:1-47.
- Lindenmayer, Aristid. (1968) “Mathematical Models for Cellular Interaction in Development. II.” Journal of Theoretical Biology. 18. 300-15. 1968; PMID:5659072; http://dx.doi.org/10.1016/0022-5193(68)90080-5
- Mancuso, Stefano. (2010) “The Roots of Plant Intelligence.” TED Global http://www.ted.com/talks/stefano_mancuso_the_roots_of_plant_intelligence.html
- Markoff, John. (2014) “IBM Develops New Computer Chip Designed to Work Like the Brain.” The New York Times. 7 August 2014. Accessed: 7 2014. http://www.nytimes.com/2014/08/08/science/new-computer-chip-is-designed-to-work-like-the-brain.html?hp&type=Homepage&&-region&-region&WT.av=pocket-region
- Maturana, Humberto, Varela, Francisco, Carmen Cienfuegos. Tr. (1973) De Máquinas y Seres Vivos: Una teoría de al organizatión biológica. Editiorial Universitaria, Santiago de Chile. 1973.
- Maturana, Humberto. Varela, Francisco. Carmen Cienfuegos. Tr. (1980) Autopoiesis and Cognition: The Realization of the Living. Dordrecht, Holland. D. Reidel Publishing Company.
- Maturana, Humberto. Varela, Francisco. Paolucci, Robert. Tr. (1987) The Tree of Knowledge: The Biological Roots of Human Understanding. Boston. Shambhala.
- Mazzolai, Barbara. Laschi, Cecilia. Dario, Paolo. Mugnai, Sergio. Mancuso, Stefano. (2010) “The Plant as a Biomechatronic System.” Plant Signaling & Behavior. 5:2 90-3. 2010; PMID:20023403; http://dx.doi.org/10.4161/psb.5.2.10457
- McMulling, Barry, Varela, Francisco J. (1997) “Rediscovering Computational Autopoiesis.” SFI (Santa Fe Institute) Working Paper 97-02-012. Accessed 26 2013. www.santafe.edu/media/workingpapers/97-02-012.pdf
- Moon, Mariella. (2014) What You Need to Know About 3D-Printed Organs. Engadget. (New York Times). Accessed: 20 2014. www.endgadget.com/2014/06/20/3d-printed-organ-explainer/.
- Mousavi, Seyed A. R., Chauvin Adeline, Pascaud, François, Kellenberger, Stephan, Farmer, Edward E. (2013) “Glutamate Receptor-Like Genes Mediate Leaf-to-Leaf Wound Signaling.” Nature. 500. 422-6. 22 2013; PMID:3969459; http://dx.doi.org/10.1038/nature12478
- Nagel, Thomas. (2012) Mind & Cosmos: Why the Materialist Neo-Darwinian Conception of Nature is Almost Certainly False. New York. Oxford University Press. Kindle edition paginated on an iPad Mini.
- Pollan, Michael. (2013) “The Intelligent Plant: Scientists Debate a New Way of Understanding Flora.” The New Yorker. 23 & 30 2013. 92-105
- Prigogine, Ilya. Nicolis, Grégoire. (1967). “On Symmetry-Breaking Instabilities in Dissipative Systems.” The Journal of Chemical Physics. 46:9. 3542; http://dx.doi.org/10.1063/1.1841255.
- Reinitz, John. (2012) “Pattern Formation.” Nature. 482. 464. 23 2012; PMID:22358813.
- Richards, Bernard. (2013/1954) “Radiolaria: The Results of Morphogenesis.” In: Cooper, S. Barry & van Leeuwen, Jan. Eds. (2013) Alan Turing: His Work and Impact. Amsterdam. Elsevier. 765.
- Spiller, Neil. Armstrong, Rachel. Eds. (2011) “Protocell Architecture.” AD: Architectural Design. Wiley. 81:2. 34-43. March/April 2011
- Swinton, Jonathan. (2004) “Watching the Daisies Grow: Turing and Fibonacci Phyllotaxis.” 477-498. In: Teuscher, Christof. Ed. (2004) Alan Turing: Life and Legacy of a Great Thinker. Berlin. Springer-Verlag.
- Swinton, Jonathan. (2013) “Turing, Morphogenesis, and Fibonacci Phyllotaxis: Life in Pictures.” 834-849. In: Cooper, Barry. & van Leeuwen, Jan. Eds. (2013) Alan Turing: His Work and Impact. Amsterdam. Elsevier.
- Thompson, Evan. (2007) Mind in Life: Biology, Phenomenology, and the Science of Mind. Cambridge, MA. Harvard University Press.
- Trewavas, Anthony. (2003) “Aspects of Plant Intelligence.” Annals of Botany. 92. 1-20; PMID:12740212; http://dx.doi.org/10.1093/aob/mcg101
- Turing, Alan M. (1936) “On Computable Numbers, With an Application to Entscheidungsproblem.” London. Proc. London Maths. Soc. Ser.2: 42. 230-265. http://plms.oxfordjournals.org/content/s2-42/1/230.full.pdf+html?ijkey=bvNIrAXLJ7n4ODP&keytype=ref.
- Turing, Alan M. (1950) “Computing Machinery and Intelligence.” Mind, New Series, 59:236 (Oct., 1950). 433-460. See also: Turing, Alan M. (1950) “Computing Machinery and Intelligence.” In: Evans, C.R., Robertson, A.D.J. (1966) Key Papers: Brain Physiology and Psychology. London. Butterworths.
- Turing, Alan M. (1952) “The Chemical Basis of Morphogenesis.” Philosophical Transactions of the Royal Society B. 237. 37-72. Reprinted in: Saunders, P.T. (1992) Collected Works of A.M. Turing: Morphogenesis. London. North-Holland.
- Turing, Alan M. (1953) “Morphogen Theory of Phyllotaxis”. In: Saunders, P.T. (1992) Collected Works of A.M. Turing: Morphogenesis. London. North-Holland.
- Turner, Scott J. (2000) The Extended Organism: The Physiology of Animal-Build Structures. Cambridge, MA. Harvard University Press.
- Uribe, Ricardo B. (1981) “Modeling Autopoiesis.” In: Zeleny, Milan. Ed. (1981) Autopoiesis: A Theory of Living Organization. New York. North Holland. 51-62.
- Varela, Francisco J. Maturana, Humberto R. Uribe, Ricardo. (1974) “Autopoiesis: The Organization of Living Systems, its Characterization and a Model.” BioSystems. . 55:4 PMID:4407425; http://dx.doi.org/10.1016/0303-2647(74)90031-8
- Varela, Francisco J. Thompson, Evan. Rosch, Eleanor. (1993) The Embodied Mind: Cognitive Science and Human Experience. Cambridge, MA. The MIT Press.
- Varela, Francisco J. Thompson, Evan. Rosch, Eleanor. (1996) “The Early Days of Autopoiesis: Heinz and Chile.” Systems Research. 1313:3. 407-16; http://dx.doi.org/10.1002/(SICI)1099-1735(199609)13:3<407::AID-SRES100>3.0.CO;2-1
- Venter, Craig. (2012) “What is Life? A 21st Century Perspective.” Lecture. Dublin. Trinity College Dublin. http://www.tcd.ie/Communications/news/news.php?headerID=2606&vs_date=2012-07-24.
- Venter, Craig (2013) Life at the Speed of Light: From the Double Helix to the Dawn of Digital Life. [Kindle Edition.] New York. Little Brown Book Group.
- Weber, Andreas. Varela, Francisco. (2002) “Life after Kant: Natural Purposes and the Autopoietic Foundations of Biological Individuality.” Phenomenology and the Cognitive Sciences. 1. 97-125; http://dx.doi.org/10.1023/A:1020368120174
- Wikipedia. (2013) “Autopoiesis.” Accessed: 9 March 2013. http://en.wikipedia.org/wiki/Autopoiesis.
- Zeleny, Milan. Ed. (1981) Autopoiesis: A Theory of Living Organization. New York. North Holland.