
Abstract
DNA vaccination has emerged as an attractive immunotherapeutic approach against cancer due to its simplicity, stability, and safety. Results from numerous clinical trials have demonstrated that DNA vaccines are well tolerated by patients and do not trigger major adverse effects. DNA vaccines are also very cost effective and can be administered repeatedly for long-term protection. Despite all the practical advantages, DNA vaccines face challenges in inducing potent antigen specific cellular immune responses as a result of immune tolerance against endogenous self-antigens in tumors. Strategies to enhance immunogenicity of DNA vaccines against self-antigens have been investigated including encoding of xenogeneic versions of antigens, fusion of antigens to molecules that activate T cells or trigger associative recognition, priming with DNA vectors followed by boosting with viral vector, and utilization of immunomodulatory molecules. This review will focus on discussing strategies that circumvent immune tolerance and provide updates on findings from recent clinical trials.
Abbreviations
| TAAs | = | tumor-associated antigens |
| CT antigens | = | cancer-testis antigens |
| CEA | = | carcinoembryonic antigen |
| Id | = | idiotype |
| TLRs | = | Toll-like receptors |
| IRF | = | interferon regulatory factor |
| IFNs | = | interferons |
| TBK1 | = | Tank-binding kinase 1 |
| STING | = | stimulator of interferon genes |
| APCs | = | antigen presenting cells |
| MHC | = | major histocompatibility complex |
| CTLs | = | cytotoxic lymphocytes |
| EP | = | electroporation |
| SCT | = | single-chain trimer |
| Trp2 | = | tyrosinase related protein 2 |
| hTERT | = | human telomerase reverse transcriptase |
| phTERT | = | optimized full-length hTERT |
| NHP | = | non-human primate |
| TT | = | tetanus toxin |
| DOM | = | fragment c domain |
| GITR | = | glucocorticoid-induced tumor necrosis factor receptor family-related genes |
| PMED | = | particle mediated epidermal delivery |
| HER2 | = | Her2/neu |
| Mam-A | = | Mammaglobin-A |
| PAP | = | Prostatic acid phosphatase |
| PSMA | = | prostate-specific membrane antigen |
| CIN | = | cervical intraepithelial neoplasia |
| HSP70 | = | heat shock protein 70 |
Introduction
Cancer remains one of the leading causes of death in the modern world. Finding effective ways to combat cancer has been one of the main goals of scientists worldwide for decades and still poses tremendous challenges. The standard treatments currently practiced in the clinic, including surgery, radiation, and chemotherapy, have shown limited success. These therapies are usually only effective against early stage localized tumors and rarely against later staged, metastatic malignancies, leading to frequent relapse. Furthermore, various agents used in radiation and chemotherapy are damaging to normal tissues, which may lead to prominent side effects. Thus, there is an urgent need for new therapeutic strategies that can specifically eliminate cancer cells and induce long lasting protection.
The immune system is the natural defense mechanism that the human body uses to combat diseases. It has been observed that intratumoral pathogenic infections can lead to spontaneous tumor regression.Citation1 This observation has demonstrated the potential antitumor properties of the immune system and has inspired the development of cancer immunotherapies including therapeutic cancer vaccines. Traditionally, vaccines have been mainly used as a preventive measure against infectious diseases, triggering the immune system to produce neutralizing antibodies against specific antigens. More recently, vaccines have also been applied as therapeutic strategies, inducing the immune system to activate cytotoxic T cells against infected cells and cancers. It has been shown that mammalian cells are capable of expressing genes encoded on plasmid DNA after transfection.Citation2 Furthermore, it was also demonstrated that intramuscular injection of plasmid DNA can lead to long term gene expression and elicit both humoral and cellular immune responses against the encoded antigen.Citation2,3 These studies have sparked the development of DNA vaccines against various diseases including influenza and HIV-1 and have demonstrated protective immunity.Citation4,5 These findings along with the recent discovery and identification of cancer antigens have propelled the investigation and development of DNA vaccines against cancer.Citation6
DNA vaccines emerge as a practical and attractive approach with great potential to translate to the clinics. Practically, they are more cost effective compared to other vaccines, such as recombinant protein, tumor cells, or viral vectors. Recent advancements in molecular biology and recombinant technologies along with the increasing identification of tumor antigens provide the tools for plasmid gene manipulation. Genes in DNA vaccines can be designed to encode different antigens as well as various other immunomodulatory molecules to manipulate the resulting immune responses. In addition, DNA vaccines allow for multiple administrations, and their safety profile have been well established in multiple studies.Citation7-9
Despite all the advantages, DNA vaccines have had limited success in producing therapeutic effects against most cancers due to poor immunogenicity. DNA vaccines have been most successfully used on cancer models where the etiological oncogenic agents are of foreign viral origin, for example, human papillomavirus-associated malignancies. However, most tumors arise from normal body tissues and express endogenous antigens that are either not recognized by or are only weakly reactive to the immune system. This is the body's natural mechanism to prevent autoimmunity in which the immune system reacts and attack the host's tissues. Furthermore, CD4+ CD25+ Foxp3 regulatory T cells inhibit the immune functions of lymphocytes capable of recognizing endogenous antigens. In addition, cytotoxic lymphocytes can be rendered anergic when the T cell receptors engage the MHC molecule: peptide complex on antigen presenting cells in the absence of costimulatory molecules. These mechanisms of central and peripheral immune tolerance limit the efficacy of DNA vaccines, which aim to exploit the host's immune system. Moreover, tumors may induce mutation or loss in the immunodominant epitopes capable in triggering the strongest T cell activation, which further hinders the therapeutic effects of DNA vaccines.
Various strategies have been investigated to enhance the potency of DNA vaccines. Plasmids encoding antigens have been designed to promote antigen expression and presentation. New vaccine delivery techniques have been explored. Immunomodulatory molecules have been used in tandem with DNA vaccines either to stimulate the immune system or reduce immunosuppression. This review focuses on the most recent developments in strategies to optimize plasmid design to enhance immunogenicity and circumvent central immune tolerance, as well as provides an update on the progress of DNA vaccines in human clinical trials.
Cancer Antigens
The advancement of genetic technologies, including genome sequencing and gene profiling, has helped with the rapid identification of tumor antigens, which has propelled the development of antigen-specific cancer immunotherapies. Cancer immunotherapies seek to use the host's immune system to eliminate cancer cells making cancer antigens critically important, as they are responsible for triggering a specific immune response. Although large numbers of cancer antigens have been discovered and are being studied in both labs and clinics, their difference in origin has resulted in great variation in immunogenicity.
Ideally, antigens for cancer vaccines should be highly immunogenic to induce strong immune responses, and only be expressed by malignant cells for specific tumor killing. However, the majority of neoplasm developments are caused by loss of growth control in normal tissues. Therefore, they present self-antigens and do not possess phenotypes that are capable of stimulating immune cells due to negative selection. During lymphocyte development, lymphocytes with high affinity toward the host's self-antigens are eliminated to prevent autoimmunity.Citation10,11 In general, the central tolerance of the host's immune system creates one of the greatest obstacles faced by scientists developing effective cancer vaccines. Investigators have been searching for cancer antigens with profiles that are capable of inducing potent immune responses. A general guideline to determine the most appropriate cancer antigens for vaccines has been created in a NCI pilot prioritization project.Citation12
Scientists have been working on identifying tumor antigens that are recognized by the immune system, and various tumor-associated antigens (TAAs) have been characterized.Citation13 The first cancer antigen reported that could be recognized by T cells, MAGE-A1, represents a class of TAAs known as the cancer-testis (CT) antigens.Citation14,15 CT antigens are good candidates for cancer vaccines as they are only expressed on particular tumor cells and immune privileged germ line tissues and not on normal adult cells.Citation16 Other CT antigens include NY-ESO-1 and SSX.Citation17,18 This class of TAAs is expressed on a number of tumors, and vaccines against NY-ESO-1 are currently in clinical trials.Citation19 New CT antigens are constantly identified and studied. Several investigations have used MAGE-C1 or MAGE-C2 as targets for multiple myeloma treatments.Citation20,21 The gene MAPE is also being assessed as a biomarker for various solid tumors.Citation22 Other CT antigens, such as HORMAD1, CXorf61, and ACTL8, as potential therapeutic targets are being evaluated.Citation23
Another class of TAAs that can serve as targets for cancer vaccines are overexpressed self-proteins.Citation24 An example of this class of TAAs is HER-2/neu, an oncoprotein most commonly associated with breast cancer.Citation25 Expression of these proteins is significantly upregulated on neoplastic cells in a wide range of tissue types on such a level that can potentially surpass the recognition threshold of T lymphocytes and trigger an antitumor immune response.
Differentiation antigens are cell type specific and shared between tumors and the normal tissue of origin.Citation26 For example, both melanoma and normal melanocytes express GP100, Tyrosinase, and Melan-A/MART-1.Citation27-29 Other differentiation antigens include PSA,Citation30 Mammoglobin-A,Citation31 and carcinoembryonic antigen (CEA) overexpressed in prostate cancer, breast carcinoma, and colon cancer respectively.Citation32 Thus, differentiation antigens may serve as good targets for cancer vaccines?
Among different tumor self-antigens, unique tumor-specific TAAs are the most immunogenic. These antigens are expressed only on tumor cells and not on any normal tissueCitation33 as a result of somatic point mutations. Antigens from this class can be generated from aberrant gene expression, such as transcription of alternative open reading frame through alternate start codons and introns, incomplete splicing, and posttranslational modifications.Citation34-37 It has been reported that proteasomes can also be used to produce unique antigenic peptides by splicing precursor proteins.Citation38 These mutated proteins often play a critical role in the oncogenic process and therefore survive immune selection in order to maintain tumor growth and proliferation. Due to their unique nature of being expressed only on tumors and not on any other normal tissues, these tumor-specific TAAs can be recognized as non-self and not be subjected to central immune tolerance, leading to generation of the highest antitumor effect when incorporated into cancer vaccines. However, certain complications prevent vaccines incorporating unique tumor-specific antigens from being readily translated into the clinics. The same tumor type can be affected by different point mutations making the identification of a widely applicable antigen for a vaccine difficult.Citation39 Furthermore, a single tumor can contain multiple mutations, whose roles in tumor proliferation would have to be characterized before the ideal vaccine antigen can be determined.Citation40 Recently, more and more unique tumor antigens have been slowly identified, such as CDK-4 and β-catenin.Citation24 Another class of unique tumor antigen is the immunoglobulin idiotype (Id) displayed on most malignant B cells. The Ids on B cell receptors are clone specific, making them unique to every B cell. This antigen represents another ideal target for cancer vaccines, in which multiple studies have been done on idiotypic vaccination.Citation41
Despite extensive studies on various self TAAs showing promise in eliciting antitumor effects in animal models, these successes have yet to be replicated in humans, held back by their inherent low immunogenicity. In fact, the most ideal antigens for cancer vaccine come from oncogenic viruses, such as the human papillomavirus, the etiological cause of cervical cancer.Citation42,43 The integration of viral genes into the host's genome after infection leads to expression of viral oncoproteins, which are foreign proteins readily recognized by lymphocytes and subjected to effector mechanisms. Since most cancer antigens come from self-tissue, cancer vaccines have to be innovatively engineered to overcome inherent immune tolerance.
Immune Activation by DNA Vaccines
The principal concept of cancer immunotherapy is to introduce various tumor antigens into the host to facilitate immune system-mediated clearance of tumor cells. Thus, the ability of a particular therapy to induce robust immune responses has a direct and significant impact on its effectiveness. Various immunotherapies including antibody therapies, cytokine therapies, adoptive T cell therapies, and cancer vaccines have been studied both pre-clinically and clinically. Among the various forms of cancer vaccines, DNA vaccines represent a promising strategy to induce such potent immune responses. A plasmid DNA that encodes antigen and other genes of interest under the control of a mammalian promoter is delivered into the host's tissues, and subsequently transfected into the cells allowing for in vivo production and expressionCitation44 by the host's protein expression machineries. DNA vaccines are shown to be able to trigger innate immune responses, and depending on their designs and sites of delivery, DNA vaccines can also elicit antigen specific humoral and cellular immune responses.Citation45
The plasmids utilized in DNA vaccines are of bacterial origin and have been shown to stimulate innate immune responses.Citation45 The bacterial DNA appears to serve as a ligand that stimulates Toll-like receptors (TLRs), a class of membrane-spanning proteins on dendritic cells that plays an important role in the innate immune system by recognizing pathogen-associated molecular patterns. Specifically, the hypomethylated CpG dinucleotides motif that is common in bacterial DNA, but rare in mammalian DNA, interacts with TLR9.Citation46 TLR9 is expressed in a number of immune cells, such as dendritic cells, B cells, and natural killer cells, which get stimulated as the introduced DNA is picked up either by direct transfection or phagocytosis.Citation47 Activation of TLR9 leads to a cascade of pro-inflammatory responses and results in the production of various cytokines. The local inflammation and increased production of cytokines from the innate immune responses can attract and activate additional immune cells, such as lymphocytes, and enhance subsequent specific immune responses.Citation48 Particularly, activation of TLR9, through the signaling of MyD88, leads to activation of interferon regulatory factor (IRF) 7, resulting in expression of Type I interferons (IFNs) (). Furthermore, the adjuvant effect of plasmid DNA was also found to be mediated by Tank-binding kinase 1 (TBK1). The presence of intracellular DNA plasmids in the cytosol can be sensed by DNA sensors, such as DAI, H2B, IFI16, DDX41, LRRFIP1, and cGAS, which activate TBK1 and stimulator of interferon genes (STING), leading to activation of IRF3 and production of Type I IFNs.Citation49 These signaling pathways were shown to be essential in the activation of antigen specific T cells and B cells.Citation50,51
Figure 1. Immune activation following DNA Vaccination. Intramuscular administration of DNA vaccine leads to transfection of DNA plasmids encoding antigens mainly in myocytes with some transfection in dendritic cells. DNA sensors such as DAI, H2B, IFI16, DDX41, LRRFIP1, and cGAS are able to detect the presence of dsDNA in the cytosol and induce the activation of STING-TBK1 signaling cascade leading to activation of IRF3 and resulting in expression of Type I IFNs. TLR9 can recognize the unmethylated CpG DNA, which through the signaling of MyD88 activates IRF7 also resulting in expression of Type I IFNs. Dendritic cells can pick up the myocyte-expressed antigens through phagocytosis as they get secreted or released following apoptosis. The antigens are then processed and presented through MHC class I to CD8+ T cells in cross-presentation. Interestingly, this process is promoted by Type I IFNs. Alternatively, dendritic cells can be directly transfected and express the antigens, which then can be presented through MHC class I to CD8+ T cells.
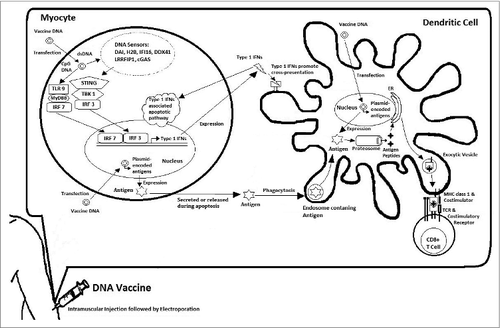
DNA vaccines can be delivered intradermally with devices like Gene Gun, leading to transfection of epidermal keratinocytes and Langerhans cells.Citation52-54 Langerhans cells are immature dendritic cells residing in the skin that actively participate in the capture and processing of antigens. Transfection of Langerhans cells with DNA leads to the expression and processing of antigens, which directly enter the presentation pathway. Langerhans cells, which serve as professional antigen presenting cells (APCs), then migrate to the lymph nodes and present antigens to naïve T-cells.Citation55,56 These endogenously produced antigens are presented by major histocompatibility complex (MHC) class I molecules on Langerhans cells to activate CD8+ cytotoxic lymphocytes (CTLs).Citation53
Intramuscular delivery of DNA vaccines results in the transfection of myocytes. Although myocytes are efficient in expressing the transfected antigens, they are incapable of activating strong specific immune responses since they are not professional APCs.Citation57 Instead, APCs, like dendritic cells, are attracted to the site of transfection, where inflammation and cytokines are generated by vaccination. APCs then capture antigens produced by transfected cells through means like phagocytosis.Citation58 These exogenous antigens are then presented by dendritic cells through MHC class II molecules and interact with CD4+ helper T cells, resulting in the activation of a humoral response.Citation59 Alternatively, the captured exogenous antigens can be presented in the context of MHC class I molecules through cross presentation to CD8+ cytotoxic T cells, leading to activation of a cellular immune response.Citation60 Importantly, Type I IFNs promote this process.Citation61 Direct transfection of DNA may also occur in APCs where direct antigen presentation through MHC class I can prime CD8+ T cells.Citation47,56
The activation of cellular immune responses is important in eliciting antitumor immunity. Specifically, a potent CD8+ cytotoxic T cell response has been shown to correlate strongly with a positive prognosis of tumor control and clearance. DNA vaccines are particularly suited to induce CD8+ T cell responses, as they can generate antigens intracellularly, which triggers the MHC class I antigen presentation pathway.Citation62 Though in principle DNA vaccines are capable of inducing therapeutic antitumor immune responses, some limitations have prevented easy and direct applications of DNA vaccines in the clinics. The designs of DNA vaccines have to be strategically optimized to achieve the desired translational efficacy.
Advantages and Disadvantages of DNA Vaccine
The use of DNA plasmids to generate antigens in vivo for cancer immunotherapy offers several advantages and practical benefits (). Molecular recombinant DNA technology allows for flexible design of DNA vectors to encode a wide range of antigens and immunomodulatory molecules.Citation63 The DNA plasmid also possesses intrinsic abilities to induce innate immune responses.Citation45 By utilizing different routes of administration and manipulating the antigen processing pathways, DNA vaccines can preferentially trigger the activation of either Th1 Helper T cells or Th2 Helper T cells and polarize the resulting immune response into being either humoral or cellular based. DNA vaccines also have the advantage of eliciting CD8+ CTL-mediated immune responses more important for tumor killing.Citation62 Since the antigens are expressed intracellularly by the plasmid-transfected host's cells, they can be presented by MHC class I and prime antigen-specific CD8+ T cells. In addition to the antigens, the vector can also encode various immunomodulatory molecules like immunostimulatory cytokinesCitation64, or agents that can target the antigens to specific processing and presentation pathways, leading to selected antibody or T-cell-mediated effects.Citation65 DNA plasmid transfection into host cells also allows for steady expression and supply of antigens.
Table 1. Advantages of DNA Vaccine
DNA vaccines represent a platform for cancer immunotherapy with the potential for mass application. They can be easily engineered and produced rapidly in large quantities. DNA vectors are very stable and can be easily stored and transported. Their safety has also been well demonstrated in both animal models and human clinical trials.Citation66-68 Unlike live attenuated bacterial or viral vaccines, there is no risk of pathogenic infection. Furthermore, the DNA vectors do not elicit anti-vector neutralizing antibody production, so multiple doses of the vaccine can be administered.Citation7 Various methods of DNA vaccine delivery have resulted in only minor, tolerable discomfort with no significant adverse effects. Practical features such as safety, ease of manufacture, and low cost make DNA vaccines an appealing option.
Despite all the positive characteristics, DNA cancer vaccines still suffer from relatively low immunogenicity, which hampers desired clinical success. Naked DNA does not easily spread from cell to cell in vivo. APCs do not readily take up expressed antigens and activate satisfactory immune responses. Thus, effective strategies that help enhance DNA vaccine potency need to be developed.
DNA Vaccine Delivery for Optimal Priming
Delivery of the encoded DNA vector is the first step of the immune activating cascade of DNA vaccines. Due to the inherently low immunogenic nature of DNA vaccines, it is imperative to adopt the route of administration that can trigger the strongest immune priming.
DNA plasmids are usually introduced intradermally or intramuscularly with the majority of the vectors ending up in the extracellular space. However, most cells at the injection site are inefficient in uptaking the injected DNA vectors resulting in low transfection efficiency. Large amounts of DNA would need to be administered for sufficient DNA uptake, reducing the cost efficiency of DNA vaccines. Strategies that directly target DNA into APCs have been explored. Within a DNA vaccine delivery device called Gene Gun, plasmid DNA is coated onto heavy metal nanoparticles, usually made of gold, and bombarded into the keratinocytes with the help of compressed helium as an acceleration force. This results in direct introduction of the antigen into the immature dendritic cells, which can process the antigen and migrate to lymph nodes where they present the antigens and activate lymphocytes. This system is able to induce substantial CD8+ T cell responses and is shown to be very efficient, requiring only an amount of DNA in the nanogram range.Citation53,69 The ease of administration also makes Gene Gun an attractive approach.
The intramuscular injection route has been another focus of DNA vaccine delivery. It has been shown that skeletal muscles are capable of generating long-term expression of transfected plasmid DNA and triggering strong immune responses in mouse models. However, scaling up from mice to human subjects, the low transfection efficiency means that a large amount of DNA would need to be applied, making such an approach impractical.Citation58 A potential solution to this problem is electroporation (EP). EP greatly increases the uptake of plasmid DNA in muscle cells by applying brief electric pulses that transiently permeabilize the cell membrane.Citation70,71 The result is a one thousand fold increase in antigen delivery compared to naked DNA injection alone.Citation72 EP itself also serves as a form of adjuvant by damaging the application site, leading to inflammation and cytokine release and ultimately recruiting APCs, such as dendritic cells and macrophages.Citation73,74 Furthermore, EP greatly reduces the amount of DNA required and greatly enhances both cellular and humoral immune responses.Citation75-77 In addition, EP does not induce an immune response against the delivery mechanism, allowing for repeated administration, and is also very well tolerated by patients showing no apparent long-term adverse effects. It appears that EP technology may be the key for successful DNA vaccine translation and is being tested in several clinical trials.Citation78,79
DNA Vaccine Design: Circumventing Immune Tolerance
Numerous strategies to improve the immunogenicity of DNA vaccines have been developed and studied, including plasmid designs that produce enhanced epitopes, which more effectively activate lymphocytes, coadministration of DNA encoding immunomodulatory and immunostimulatory molecules, the use of prime boost strategies, and approaches to break the immunosuppressive networks in the tumor microenvironment (reviewed in Citation63). The problem of immune tolerance still remains a major hurdle for DNA vaccination, as most tumor antigens are self-antigens that cannot trigger potent immune responses. Attempts have been made to engineer DNA vaccines that can break immune tolerance. Previously, a study showed that DNA vaccine encoding alphavirus replicon can activate the innate immune pathway and elicit antitumor immune responses against self-TAA tyrosinase related protein 1, effectively overcoming immune tolerance.Citation80
Recently, more innovative DNA vaccine designs aiming to circumvent central tolerance have been investigated in which antigen-specific antitumor immune responses were observed. One approach to circumvent central tolerance against self antigens is by creating a DNA construct encoding a secreted chimeric protein consisting of a single-chain trimer (SCT) of MHC class I heavy chain, β2-microglobulin, and peptide antigen linked to immunoglobulin G.Citation81 This chimeric protein is shown to form a dimer that can strongly bind to antigen-specific CD8+ T cells with high efficiency and directly induce T cell activation and proliferation. The SCT region of the construct possesses the property of directly and stably displaying an antigenic peptide to CD8+ cytotoxic T cells with high affinity. This effect provides an advantage of bypassing the antigen-processing pathway that may result in suboptimal efficiency of antigen presentation. Furthermore, the IgG domain of the construct causes the chimeric protein to form a dimer and bind to the Fc receptors on APCs, such as dendritic cells, which migrate to the lymph nodes where the antigen on the chimeric protein is displayed to prime antigen-specific T cells. Importantly, intradermal vaccination of DNA encoding the SCT chimeric protein linked to a melanoma antigen tyrosinase related protein 2 (Trp2), a self-antigen also expressed in tumors, is able to elicit strong Trp2-specific CD8+ T cell mediated immune responses and demonstrates therapeutic antitumor effects in B16 melanoma tumor models in mice.Citation81 This innovative DNA construct demonstrates translational value by inducing strong immune responses against a self-antigen. Further studies need to be performed to examine the effect of DNA vaccination using this approach against other self-antigens in tumors.
The potential of using xenogeneic versions of antigens in DNA vaccines to bypass central immune tolerance has been investigated, mostly in melanoma. Interestingly, a recent study reported the use xenogeneic p53 in colon cancer. The tumor suppressor gene p53 is mutated and overexpressed in various cancers. P53 represents another self-antigen in which only low affinity CD8+ T cells against p53 may be generated resulting in weak antigen-specific immune responses. A study shows that a xenogeneic version of the p53 gene is able to induce potent p53-specific immune responses.Citation82 Interestingly, intramuscular vaccination of DNA encoding the human p53 gene followed by electroporation elicits a strong CD8+ T cell response against mouse p53 in mice. Furthermore, these effects are shown to induce both prophylactic and therapeutic antitumor effects against murine colon cancer MC38 expressing mouse p53.Citation82 It is likely that the xenogeneic version of p53 is recognized by the immune system as foreign due to its origin from a different species. It is important to note that this particular strategy utilizes DNA sequences encoding genes homologous between 2 species. Therefore, the expressed antigen has to be similar to be recognized as the same host antigen, yet different to bypass tolerance against the self-antigen. Future studies are needed to evaluate the effectiveness of the xenogeneic p53 DNA vaccine on other p53-expressing cancers.
The DNA vaccine strategy of encoding a xenogeneic version of an antigen was examined in another pre-clinical study. Human telomerase reverse transcriptase (hTERT) is another self-antigen highly expressed in a wide range of human cancers. A DNA vaccine encoding a synthetic, highly optimized full-length hTERT (phTERT) has been engineered. Intramuscular injection of the DNA followed by electroporation has elicited a strong CD8+ T cell response against hTERT and has generated activated T cells expressing CD107a, IFNγ, and TNFα in mice.Citation83 More importantly, vaccination with phTERT is able to induce robust CD8+ T cell-mediated antitumor immune responses in a non-human primate (NHP) model. Immune studies in NHP models carry significant translational potential as the immune systems of NHPs closely match that of humans. In addition, in this particular DNA vaccine study, sequence homology analysis reveals that hTERT shares 96% identity with NHP TERT. Vaccination with phTERT in NHPs leads to improved tumor control and survival in both prophylactic and therapeutic studies in HPV-16 associated tumor models.Citation83 A cytotoxicity assay also showed that phTERT-induced T cells can eliminate hTERT target cells. phTERT is shown to be able to break immune tolerance in the human-related NHP model, and warrants further investigation.
A heterologous prime boost vaccine system using DNA and an adenoviral vector has been reported to induce antigen-specific immune responses and antitumor effects against self-TAAs. Evidence has shown that DNA plasmids are most efficient in priming the immune system, while boosting with a viral vector leads to superior immune responses compared to boosting with additional DNA plasmids. Intramuscular injection of 2 DNA plasmids encoding TAA constructs consisting of CEA fused to the B subunit of Escherichia coli heat-labile toxin and the extracellular and transmembrane domain Her2 followed by electroporation, followed by boosting with an adenoviral subtype 6 dicistronic vector carrying the same genes was able to generate both T cell-mediated and antibody responses against both TAAs in mice.Citation84 Furthermore, the vaccine system was able to induce therapeutic antitumor immunity in both Her2+ mammary tumors and CEA+ colon tumors. The heterologous prime boost vaccine demonstrated immune activation against immune tolerant self-TAAs, and its safety and immunogenicity in humans are currently being evaluated in a phase I clinical trial.Citation79
A DNA vaccine encoding wild type c-Myb cDNA flanked by 2 helper T cell epitopes of tetanus toxin(TT) has been shown to trigger an immune response against the self- antigen c-Myb by utilizing associative recognition. The epitopes of tetanus toxin are highly immunogenic to helper T cells, which help induce CD8+ cytotoxic T cell and CD4+ helper T cell responses against the weakly immunogenic c-Myb.Citation85 Interestingly, the DNA vaccine is administered intravenously as opposed to the common intramuscular or intradermal delivery routes. Nevertheless, the vaccine is able to generate prophylactic antitumor immunity mediated by CD8+ and CD4+ T cells against murine colon cancer model MC83 in the absence of adjuvant or immunostimulatory molecules.Citation85 The use of TT derived T cell epitopes in DNA vaccines have been further investigated in recent studies. A fusion DNA vaccine coupling fragment C domain (DOM) of TT with PASD1, a CT antigen, was able to induce CTL response against human multiple myeloma in HLA transgenic mice.Citation86 In another study, DNA encoding the TT domain fused with S9C, an immunodomant HLA-A2-binding peptide in NY-ESO-1, generated T cells capable of killing tumor cells expressing endogenous NY-ESO-1.Citation87 These studies demonstrate the potential of DNA vaccines incorporating TT epitopes in enhancing immunogenicity. The effect of utilizing potent tetanus toxin epitopes to help elicit immune responses against weak self-antigens should be further studied using other TAAs.
It has been shown that administration of molecules to modulate immune cells can enhance the potency of DNA vaccines against self-antigens. A previous study showed that blockade of CTLA-4 with anti-CTLA-4 antibody after vaccination with DNA encoding melanoma differentiation antigens Trp2 and gp100 resulted in enhanced B16 tumor rejection.Citation88 In another study, an anti-glucocorticoid-induced tumor necrosis factor receptor family-related gene (GITR) monoclonal antibody was also shown to enhance CD8+ T cell immune responses against the same melanoma antigens.Citation89 These earlier studies demonstrate the adjuvant effect of immunostimulatory agents. In a recent study, plasmid DNA encoding p53 or gp100 along with DNA encoding CD40L, a costimulatory molecule that activates APCs, were injected intramuscularly and followed by electroporation. Significant increase in antigen specific CTL cellular immune responses was observed in both p53 and gp100 tumor models.Citation90 Furthermore, the immune responses were shown to be CD8+ T cell mediated and result in potent antitumor response. The results show that immunostimulatory molecules can be incorporated into DNA vaccines to enhance the vaccines’ potency. Future DNA vaccine strategies should aim to incorporate immunomodulating molecules either as adjuvants or encoded in DNA plasmids to further enhance the immunogenicity against self-antigens.
Importantly, these DNA vaccine strategies demonstrating effectiveness in inducing antigen-specific immune responses against self-antigens have not been shown to trigger damaging autoimmune attacks against normal tissues. The first DNA vaccine that has broken immune tolerance and has been translated into the market was actually implemented in dogs. A DNA vaccine encoding xenogeneic tyrosinase has been shown to elicit antitumor response against oral melanoma in dogs and demonstrated safety and therapeutic efficacy in a phase I trial.Citation91,92 These promising results have led to the USDA licensure of the DNA vaccine, Oncept, as an immunotherapy for oral melanoma in dogs.Citation93 Although no DNA vaccines for humans have been standardized for cancer treatment, numerous clinical trials are currently underway to assess their translational potential.
Human Clinical Trials
Clinical trials provide opportunities to evaluate whether DNA vaccines will be able to fulfill their ultimate goal of demonstrating efficacy in treating humans. Since the first DNA vaccine clinical trial on HIV-1, numerous clinical trials have demonstrated the safety and tolerability of various DNA vaccine platforms. Injection of DNA plasmids is well tolerated by patients with minimal to no systemic toxicities being reported. Integration of DNA vectors into the host genomes is not a concern, and no detectable increase in antibodies against DNA is observed.Citation94 The widely accepted safety profile of DNA vaccines has led to relaxed requirements for FDA approval and the frequent combination of first and second phase trials into one trial. Since the safety of DNA vaccines is well established, the main interest in clinical trials has become demonstrating efficacy. Here, we report a brief summary of results from a number of recent clinical trials on DNA vaccines against cancer ().
Table 2. Human Clinical Trials
Melanoma
Malignant melanoma expresses a number of TAAs that can be used as targets for DNA vaccination. Intranodal delivery of DNA plasmids encoding Melan-A (MART-1) and tyrosinase have been shown to elicit both cellular and humoral-mediated immune responses in stage IV melanoma patients in previous trials.Citation95 A phase I trial has been conducted using DNA encoding xenogeneic mouse gp100 or human gp100.Citation66 Mouse or human gp100 plasmid DNA were injected with 3 dosages (100, 500, or 1500 ug) intramuscularly every 3 weeks, and then with the gp100 of the other species 3 times. Only mild toxicity was observed at the injection site in 12 out of 19 enrolled patients. Furthermore, CD8+ T cells binding gp100 HLA-A2 restricted tetramers were elicited in 5 patients while one patient showed an increase in IFNγ+ CD8+ T cells. However, no difference in progression-free survival was found between patients with or without immune responses.
Another pilot clinical trial was conducted to compare the immunological responses of intramuscular delivery and particle mediated epidermal delivery (PMED) of the xenogeneic gp100 DNA.Citation96 27 stage IIB-IV melanoma patients received 8 vaccinations with either PMED or IM over 4 months with either 4ug or 2000ug of mouse gp100 DNA respectively. The safety profile of PMED was found to be comparable to that of the intramuscular administration route. Furthermore, PMED seemed to induce higher IFNγ+ CD8+ T cell production while requiring a significantly lower dose of DNA. Although 30% of the vaccinated patients displayed immune responses, no significant clinical outcomes were observed.
Breast cancer
Her2/neu (HER2) is an oncoprotein overexpressed in breast cancer, and is used as a target antigen for DNA vaccines in clinical trials. In a pilot clinical trial, a vaccine using DNA plasmid encoding full-length signaling-deficient version of HER2 was administered along with low doses of IL-2 and GM-CSF in metastatic HER2-expressing breast cancer patients. The vaccine was well tolerated with no clinical toxicity or autoimmunity observed. Although no improved T cell responses were observed, the vaccine was able to generate long-term antibody responses, and 2 out of the 6 patients who completed all 3 cycles of vaccination survived for more than 4 years after the vaccinations.Citation97 Another vaccine targeting dendritic cells with DNA encoding chimeric rat/human HER2 was tested in 28 HER2 breast and 16 HER2 pancreatic cancer patients.Citation98 The chimeric plasmid was able to induce T cell responses and significantly hinder HER2+ tumor growth, with the ability to circumvent suppressor effects of regulatory T cells, IL10, and TGF-β. No therapeutic outcome was reported. Thus, further testing should be conducted.
Mammaglobin-A (Mam-A) is another protein overexpressed in breast cancer. Preclinical studies have shown that a vaccine encoding Mam-A cDNA can generate Mam-A-specific CD8+ T cell immune responses. This led to a phase I trial of the Mam-A DNA vaccine in stage IV metastatic breast cancer patients. Patients were vaccinated with Mam-A cDNA on days 0, 28, and 56. Interestingly, an increase in Th1 CD4+ T cells was observed. Furthermore, the activated CD4+ helper T cells shifted from expressing IL-10 to expressing IFNγ and induced preferential lysis of Mam-A-expressing breast cancer cells.Citation99 This result shows that Mam-A cDNA vaccine is able to elicit antitumor immunity against breast cancer. Further studies should be conducted to evaluate long-term therapeutic outcomes.
Colorectal cancer
Application of a DNA vaccine against colorectal cancer has also been studied in a phase I clinical trial. The vaccine plasmid encodes a modified version of the human carcinoembryonic antigen (CEA) gene fused to a promiscuous T helper epitope of the tetanus toxoid, and has been shown to be immunogenic in mice. In the trial, 10 patients were treated with cyclophosphamide intravenously before the first vaccination. CEA66 DNA plasmids were injected either intradermally (2mg) or intramuscularly (8mg) on week 0, 2, and 6, along with subcutaneous injection of GM-CSF(150ug). Only minor adverse effects, such as fatigue, headache, arthralgia, chest tightness, and myalgia, were observed at the vaccination site. During the follow-up period, one patient had a recurrence, and interestingly, another patient was diagnosed with urinary bladder cancer, which was unrelated to the DNA vaccination treatment.Citation67
Prostate cancer
Preclinical studies have demonstrated that a DNA vaccine encoding Prostatic acid phosphatase (PAP), a prostate associated antigen, can elicit PAP-specific CD8+ T cell immune responses. A phase I/II trial was conducted using a DNA vaccine encoding human PAP to treat 22 stage D0 prostate cancer patients. In this dose escalation study with 100 mg, 500 mg, or 1500 mg of plasmids, DNA was injected intradermally along with the adjuvant GM-CSF (200mg) 6 times at 14-day intervals. During the one-year observation after treatment, 3 of 22 patients developed PAP-specific IFNγ+ CD8+ T cells and 9 of 22 patients showed proliferation of PAP-specific CD4+ and CD8+ T cells.Citation100 Although no PSA values in patients declined by more than 50%, several patients were reported with a decrease in the rate of serum PSA rise after treatment. A subsequent study was conducted on this vaccine and found that multiple boosting can enhance the elicited immune responses.Citation101 Future trials evaluating the vaccine's clinical therapeutic effect are warranted.
In a phase I/II trial, a DNA vaccine encoding a tumor derived epitope from prostate-specific membrane antigen (PSMA) fused to a domain of fragment C of tetanus toxin was administered either intramuscularly alone or intramuscularly followed by electroporation. The tetanus toxin fragment C domain was shown to induce CD4+ T cell help and elicit antibody responses. Furthermore, intramuscular injection of the vaccine followed by EP generated a greater amount of antibodies. The results showed that intramuscular injection followed by EP is safe and can generate strong humoral responses. Moreover, PSMA-specific CD8+ T cells were detected in patients after administration of the DNA vaccine.Citation102 However, no effect of DNA dose on outcome was detected. Nevertheless, a reduction on the rate of disease progression was observed.
Cervical cancer
DNA vaccines theoretically should generate the strongest immune responses against cervical cancer due its etiological factor being human papillomavirus infection. HPV E6 and E7 are foreign antigens and are only expressed in transformed cancer cells, making them ideal targets. Various DNA vaccines encoding the viral oncoproteins HPV E6 and E7 have shown to generate potent cellular and humoral immune responses in mice. Numerous fusion DNA vaccines encoding HPV E7 and other manipulative molecules have been developed. A phase I trial was conducted in patients with grade 2/3 cervical intraepithelial neoplasia (CIN). This DNA vaccine encodes a modified version of HPV E7 incapable of binding retinoblastoma protein and fused to heat shock protein 70 (HSP70) as well as a secretion signal sequence. The patients received 3 IM vaccinations (0.5, 1, or 3mg) on days 0, 28, and 56. Histologic outcomes based on resection were evaluated at week 15, and histologic regressions were observed in 33% of the patients in the highest-dose cohort.Citation68
In another phase I trial, the effect of a highly optimized DNA vaccine, VGX-3100, encoding HPV 16 and 18 E6/E7 antigens was evaluated in 18 patients with grade 2/3 CIN. The DNA vaccine was injected intramuscularly followed by electroporation 3 times in a dose escalating manner (0.3, 1, and 3 mg). The vaccine was able to induce HPV specific CD8+ T cells that loaded granzyme B and perforin.Citation103 Higher levels of interferon gamma were also observed in the 3 mg cohort compared to the other 2 doses. The immunization was also well tolerated with no dose limiting toxicity. It would be of interest to evaluate whether this treatment strategy can lead to regression or clearance of lesion.
In another study conducted on grade 2/3 CIN patients, intramuscular injection of DNA vaccine targeting HPV16 E6/E7 was shown to increase local CD8+ T cell responses in the tumor microenvironment. Particularly, histologic and molecular changes, including an average of threefold increase in intensity of CD8+ T cell infiltrates in both the stromal and epithelial compartments, were observed. Increased expression of genes associated with immune activation and effector functions were also detected in CD8+ T cell infiltrates.Citation104 Specifically, the histological changes in the stroma were characterized by increases in the expressions of immune activation gene CXCR3 and effector function genes Tbet and IFN β. The result from this study shows that HPV DNA vaccine can induce robust localized effector immune responses in cervical cancer patients.
Conclusion
DNA vaccine has demonstrated promising potential as an effective immunotherapeutic strategy against cancer. DNA plasmids can be easily designed and manipulated to induce potent cell-mediated immune responses. Furthermore, their ease of transportation and storage makes mass production and administration easily achievable. In addition, increasing numbers of clinical trials on various DNA vaccines against different cancers have shown promising results.
Although numerous strategies to enhance the immunogenicity and potency of DNA vaccines have already been developed and studied, future DNA vaccines should aim to further enhance antitumor immunity by circumventing immune tolerance, breaking the immunosuppressive networks in the tumor microenvironment, and inducing long lasting memory. DNA plasmids should be optimally designed to induce the strongest priming of immune responses. The activities of immunosuppressive agents in the tumor microenvironment, such as regulatory T cells and myeloid derived suppressor cells, need to be controlled. Furthermore, DNA vaccines can be used in conjunction with other cancer treatments to further control and eliminate tumors. As efforts to optimize DNA vaccines continue, we will come closer to alleviating the burden of cancer.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank Annie Wu for her helpful comments and critical review of the manuscript. This review is not intended to be an encyclopedic one, and the authors apologize to those not cited.
Funding
This work was supported by the National Institutes of Health/National Cancer Institute Cervical Cancer SPORE P50CA098252, Head and Neck Cancer SPORE P50 CA-DE019032, R43CA174436, 2R01CA114425–06, and R01CA142691 grants.
References
- Nauts HC, Fowler GA, Bogatko FH. A review of the influence of bacterial infection and of bacterial products (Coley's toxins) on malignant tumors in man; a critical analysis of 30 inoperable cases treated by Coley's mixed toxins, in which diagnosis was confirmed by microscopic examination selected for special study. Acta Med Scand Suppl 1953; 276:1-103; PMID:13039964
- Wolff JA, Malone RW, Williams P, Chong W, Acsadi G, Jani A, Felgner PL. Direct gene transfer into mouse muscle in vivo. Science 1990; 247:1465-8; PMID:1690918; http://dx.doi.org/10.1126/science.1690918
- Raz E, Carson DA, Parker SE, Parr TB, Abai AM, Aichinger G, Gromkowski SH, Singh M, Lew D, Yankauckas MA, et al. Intradermal gene immunization: the possible role of DNA uptake in the induction of cellular immunity to viruses. Proc Natl Acad Sci U S A 1994; 91:9519-23.
- Ulmer JB, Donnelly JJ, Parker SE, Rhodes GH, Felgner PL, Dwarki VJ, Gromkowski SH, Deck RR, DeWitt CM, Friedman A, et al. Heterologous protection against influenza by injection of DNA encoding a viral protein. Science 1993; 259:1745-9; PMID:8456302; http://dx.doi.org/10.1126/science.8456302
- Wang B, Ugen KE, Srikantan V, Agadjanyan MG, Dang K, Refaeli Y, Sato AI, Boyer J, Williams WV, Weiner DB. Gene inoculation generates immune responses against human immunodeficiency virus type 1. Proc Natl Acad Sci U S A 1993; 90:4156-60.
- Wang RF, Rosenberg SA. Human tumor antigens for cancer vaccine development. Immunol Rev 1999; 170:85-100; PMID:10566144; http://dx.doi.org/10.1111/j.1600-065X.1999.tb01331.x
- Smith HA. Regulation and review of DNA vaccine products. Dev Biol 2000; 104:57-62; PMID:11713825
- Rinaldi M, Signori E, Rosati P, Cannelli G, Parrella P, Iannace E, Monego G, Ciafre SA, Farace MG, Iurescia S, et al. Feasibilty of in utero DNA vaccination following naked gene transfer into pig fetal muscle: transgene expression, immunity and safety. Vaccine 2006; 24:4586-91; PMID:16154671; http://dx.doi.org/10.1016/j.vaccine.2005.08.030
- MacGregor RR, Boyer JD, Ugen KE, Lacy KE, Gluckman SJ, Bagarazzi ML, Chattergoon MA, Baine Y, Higgins TJ, Ciccarelli RB, et al. First human trial of a DNA-based vaccine for treatment of human immunodeficiency virus type 1 infection: safety and host response. J Infect Dis 1998; 178:92-100; PMID:9652427; http://dx.doi.org/10.1086/515613
- Gallegos AM, Bevan MJ. Central tolerance: good but imperfect. Immunol Rev 2006; 209:290-6; PMID:16448550; http://dx.doi.org/10.1111/j.0105-2896.2006.00348.x
- Kreuwel HT, Sherman LA. The T-cell repertoire available for recognition of self-antigens. Curr Opin Immunol 2001; 13:639-43; PMID:11677083; http://dx.doi.org/10.1016/S0952-7915(01)00272-2
- Cheever MA, Allison JP, Ferris AS, Finn OJ, Hastings BM, Hecht TT, Mellman I, Prindiville SA, Viner JL, Weiner LM, et al. The prioritization of cancer antigens: a national cancer institute pilot project for the acceleration of translational research. Clin Cancer Res 2009; 15:5323-37; PMID:19723653; http://dx.doi.org/10.1158/1078-0432.CCR-09-0737
- Novellino L, Castelli C, Parmiani G. A listing of human tumor antigens recognized by T cells: March 2004 update. Cancer Immunol Immunother 2005; 54:187-207; PMID:15309328; http://dx.doi.org/10.1007/s00262-004-0560-6
- van der Bruggen P, Traversari C, Chomez P, Lurquin C, De Plaen E, Van den Eynde B, Knuth A, Boon T. A gene encoding an antigen recognized by cytolytic T lymphocytes on a human melanoma. Science 1991; 254:1643-7; PMID:1840703; http://dx.doi.org/10.1126/science.1840703
- Caballero OL, Chen YT. Cancer/testis (CT) antigens: potential targets for immunotherapy. Cancer Sci 2009; 100:2014-21; PMID:19719775; http://dx.doi.org/10.1111/j.1349-7006.2009.01303.x
- Van Der Bruggen P, Zhang Y, Chaux P, Stroobant V, Panichelli C, Schultz ES, Chapiro J, Van Den Eynde BJ, Brasseur F, Boon T. Tumor-specific shared antigenic peptides recognized by human T cells. Immunol Rev 2002; 188:51-64; PMID:12445281; http://dx.doi.org/10.1034/j.1600-065X.2002.18806.x
- Jager E, Chen YT, Drijfhout JW, Karbach J, Ringhoffer M, Jager D, Arand M, Wada H, Noguchi Y, Stockert E, et al. Simultaneous humoral and cellular immune response against cancer-testis antigen NY-ESO-1: definition of human histocompatibility leukocyte antigen (HLA)-A2-binding peptide epitopes. J Exp Med 1998; 187:265-70; PMID:9432985; http://dx.doi.org/10.1084/jem.187.2.265
- Garcia Leme J, Morato M, Souza MZ. Anti-inflammatory action of glucagon in rats. Br J Pharmacol 1975; 55:65-8; PMID:1182350; http://dx.doi.org/10.1111/j.1476-5381.1975.tb07611.x
- Odunsi K, Matsuzaki J, Karbach J, Neumann A, Mhawech-Fauceglia P, Miller A, Beck A, Morrison CD, Ritter G, Godoy H, et al. Efficacy of vaccination with recombinant vaccinia and fowlpox vectors expressing NY-ESO-1 antigen in ovarian cancer and melanoma patients. Proc Natl Acad Sci U S A 2012; 109:5797-802.
- de Carvalho F, Vettore AL, Colleoni GW. Cancer/Testis Antigen MAGE-C1/CT7: new target for multiple myeloma therapy. Clin Dev Immunol 2012; 2012:257695; PMID:22481966
- Reinhard H, Yousef S, Luetkens T, Fehse B, Berdien B, Kroger N, Atanackovic D. Cancer-testis antigen MAGE-C2/CT10 induces spontaneous CD4+ and CD8+ T-cell responses in multiple myeloma patients. Blood Cancer J 2014; 4:e212; PMID:24813080; http://dx.doi.org/10.1038/bcj.2014.31
- Goodison S, Urquidi V. The cancer testis antigen PRAME as a biomarker for solid tumor cancer management. Biomark Med 2012; 6:629-32; PMID:23075240; http://dx.doi.org/10.2217/bmm.12.65
- Yao J, Caballero OL, Yung WK, Weinstein JN, Riggins GJ, Strausberg RL, Zhao Q. Tumor subtype-specific cancer-testis antigens as potential biomarkers and immunotherapeutic targets for cancers. Cancer Immunol Res 2014; 2:371-9; PMID:24764584; http://dx.doi.org/10.1158/2326-6066.CIR-13-0088
- Lowe DB, Shearer MH, Jumper CA, Kennedy RC. Towards progress on DNA vaccines for cancer. Cell Mol Life Sci 2007; 64:2391-403; PMID:17565444; http://dx.doi.org/10.1007/s00018-007-7165-0
- Ross JS, Fletcher JA. The HER-2/neu oncogene in breast cancer: prognostic factor, predictive factor, and target for therapy. Stem Cells 1998; 16:413-28; PMID:9831867; http://dx.doi.org/10.1002/stem.160413
- Buonaguro L, Petrizzo A, Tornesello ML, Buonaguro FM. Translating tumor antigens into cancer vaccines. Clin Vaccine Immunol 2011; 18:23-34; PMID:21048000; http://dx.doi.org/10.1128/CVI.00286-10
- Bakker AB, Schreurs MW, de Boer AJ, Kawakami Y, Rosenberg SA, Adema GJ, Figdor CG. Melanocyte lineage-specific antigen gp100 is recognized by melanoma-derived tumor-infiltrating lymphocytes. J Exp Med 1994; 179:1005-9; PMID:8113668; http://dx.doi.org/10.1084/jem.179.3.1005
- Wang RF, Appella E, Kawakami Y, Kang X, Rosenberg SA. Identification of TRP-2 as a human tumor antigen recognized by cytotoxic T lymphocytes. J Exp Med 1996; 184:2207-16; PMID:8976176; http://dx.doi.org/10.1084/jem.184.6.2207
- Kawakami Y, Eliyahu S, Sakaguchi K, Robbins PF, Rivoltini L, Yannelli JR, Appella E, Rosenberg SA. Identification of the immunodominant peptides of the MART-1 human melanoma antigen recognized by the majority of HLA-A2-restricted tumor infiltrating lymphocytes. J Exp Med 1994; 180:347-52; PMID:7516411; http://dx.doi.org/10.1084/jem.180.1.347
- Corman JM, Sercarz EE, Nanda NK. Recognition of prostate-specific antigenic peptide determinants by human CD4 and CD8 T cells. Clin Exp Immunol 1998; 114:166-72; PMID:9822272; http://dx.doi.org/10.1046/j.1365-2249.1998.00678.x
- Jaramillo A, Majumder K, Manna PP, Fleming TP, Doherty G, Dipersio JF, Mohanakumar T. Identification of HLA-A3-restricted CD8+ T cell epitopes derived from mammaglobin-A, a tumor-associated antigen of human breast cancer. Int J Cancer 2002; 102:499-506; PMID:12432553; http://dx.doi.org/10.1002/ijc.10736
- Berinstein NL. Carcinoembryonic antigen as a target for therapeutic anticancer vaccines: a review. J Clin Oncol 2002; 20:2197-207; PMID:11956282; http://dx.doi.org/10.1200/JCO.2002.08.017
- Parmiani G, De Filippo A, Novellino L, Castelli C. Unique human tumor antigens: immunobiology and use in clinical trials. J Immunol 2007; 178:1975-9; PMID:17277099; http://dx.doi.org/10.4049/jimmunol.178.4.1975
- Coulie PG, Lehmann F, Lethe B, Herman J, Lurquin C, Andrawiss M, Boon T. A mutated intron sequence codes for an antigenic peptide recognized by cytolytic T lymphocytes on a human melanoma. Proc Natl Acad Sci U S A 1995; 92:7976-80.
- Guilloux Y, Lucas S, Brichard VG, Van Pel A, Viret C, De Plaen E, Brasseur F, Lethe B, Jotereau F, Boon T. A peptide recognized by human cytolytic T lymphocytes on HLA-A2 melanomas is encoded by an intron sequence of the N-acetylglucosaminyltransferase V gene. J Exp Med 1996; 183:1173-83; PMID:8642259; http://dx.doi.org/10.1084/jem.183.3.1173
- Wang RF, Parkhurst MR, Kawakami Y, Robbins PF, Rosenberg SA. Utilization of an alternative open reading frame of a normal gene in generating a novel human cancer antigen. J Exp Med 1996; 183:1131-40; PMID:8642255; http://dx.doi.org/10.1084/jem.183.3.1131
- Skipper JC, Hendrickson RC, Gulden PH, Brichard V, Van Pel A, Chen Y, Shabanowitz J, Wolfel T, Slingluff CL, Jr., Boon T, et al. An HLA-A2-restricted tyrosinase antigen on melanoma cells results from posttranslational modification and suggests a novel pathway for processing of membrane proteins. J Exp Med 1996; 183:527-34; PMID:8627164; http://dx.doi.org/10.1084/jem.183.2.527
- Vigneron N, Stroobant V, Chapiro J, Ooms A, Degiovanni G, Morel S, van der Bruggen P, Boon T, Van den Eynde BJ. An antigenic peptide produced by peptide splicing in the proteasome. Science 2004; 304:587-90; PMID:15001714; http://dx.doi.org/10.1126/science.1095522
- Wortzel RD, Philipps C, Schreiber H. Multiple tumour-specific antigens expressed on a single tumour cell. Nature 1983; 304:165-7; PMID:6602947; http://dx.doi.org/10.1038/304165a0
- Fox EJ, Salk JJ, Loeb LA. Cancer genome sequencing–an interim analysis. Cancer Res 2009; 69:4948-50; PMID:19509220; http://dx.doi.org/10.1158/0008-5472.CAN-09-1231
- Bendandi M. Idiotype vaccines for lymphoma: proof-of-principles and clinical trial failures. Nat Rev Cancer 2009; 9:675-81; PMID:19701243; http://dx.doi.org/10.1038/nrc2717
- Damania B. DNA tumor viruses and human cancer. Trends Microbiol 2007; 15:38-44; PMID:17113775; http://dx.doi.org/10.1016/j.tim.2006.11.002
- Roden R, Wu TC. How will HPV vaccines affect cervical cancer? Nat Rev Cancer 2006; 6:753-63; PMID:16990853; http://dx.doi.org/10.1038/nrc1973
- Webster RG, Robinson HL. DNA vaccines: a review of developments. BioDrugs 1997; 8:273-92; PMID:18020519
- Rice J, Ottensmeier CH, Stevenson FK. DNA vaccines: precision tools for activating effective immunity against cancer. Nat Rev Cancer 2008; 8:108-20; PMID:18219306; http://dx.doi.org/10.1038/nrc2326
- Klinman DM. Adjuvant activity of CpG oligodeoxynucleotides. Int Rev Immunol 2006; 25:135-54; PMID:16818369; http://dx.doi.org/10.1080/08830180600743057
- Ulmer JB, Otten GR. Priming of CTL responses by DNA vaccines: direct transfection of antigen presenting cells versus cross-priming. Dev Biol 2000; 104:9-14; PMID:11713829
- Hoebe K, Jiang Z, Tabeta K, Du X, Georgel P, Crozat K, Beutler B. Genetic analysis of innate immunity. Adv Immunol 2006; 91:175-226; PMID:16938541; http://dx.doi.org/10.1016/S0065-2776(06)91005-0
- Kouji Kobiyama, Nao Jounai, Taiki Aoshi, Miyuki Tozuka, Fumihiko Takeshita, Cevayir Coban, and Ken J. Ishii. Innate Immune Signaling by, and Genetic Adjuvants for DNA Vaccination. Vaccines 2013; 1:278-92.
- Ishii KJ, Kawagoe T, Koyama S, Matsui K, Kumar H, Kawai T, Uematsu S, Takeuchi O, Takeshita F, Coban C, et al. TANK-binding kinase-1 delineates innate and adaptive immune responses to DNA vaccines. Nature 2008; 451:725-9; PMID:18256672; http://dx.doi.org/10.1038/nature06537
- Ishikawa H, Ma Z, Barber GN. STING regulates intracellular DNA-mediated, type I interferon-dependent innate immunity. Nature 2009; 461:788-92; PMID:19776740; http://dx.doi.org/10.1038/nature08476
- Fuller DH, Loudon P, Schmaljohn C. Preclinical and clinical progress of particle-mediated DNA vaccines for infectious diseases. Methods 2006; 40:86-97; PMID:16997717; http://dx.doi.org/10.1016/j.ymeth.2006.05.022
- Trimble C, Lin CT, Hung CF, Pai S, Juang J, He L, Gillison M, Pardoll D, Wu L, Wu TC. Comparison of the CD8+ T cell responses and antitumor effects generated by DNA vaccine administered through gene gun, biojector, and syringe. Vaccine 2003; 21:4036-42; PMID:12922140; http://dx.doi.org/10.1016/S0264-410X(03)00275-5
- Lauterbach H, Gruber A, Ried C, Cheminay C, Brocker T. Insufficient APC capacities of dendritic cells in gene gun-mediated DNA vaccination. J Immunol 2006; 176:4600-7; PMID:16585550; http://dx.doi.org/10.4049/jimmunol.176.8.4600
- Porgador A, Irvine KR, Iwasaki A, Barber BH, Restifo NP, Germain RN. Predominant role for directly transfected dendritic cells in antigen presentation to CD8+ T cells after gene gun immunization. J Exp Med 1998; 188:1075-82; PMID:9743526; http://dx.doi.org/10.1084/jem.188.6.1075
- Akbari O, Panjwani N, Garcia S, Tascon R, Lowrie D, Stockinger B. DNA vaccination: transfection and activation of dendritic cells as key events for immunity. J Exp Med 1999; 189:169-78; PMID:9874573; http://dx.doi.org/10.1084/jem.189.1.169
- Fu TM, Ulmer JB, Caulfield MJ, Deck RR, Friedman A, Wang S, Liu X, Donnelly JJ, Liu MA. Priming of cytotoxic T lymphocytes by DNA vaccines: requirement for professional antigen presenting cells and evidence for antigen transfer from myocytes. Mol Med 1997; 3:362-71; PMID:9234241
- Dupuis M, Denis-Mize K, Woo C, Goldbeck C, Selby MJ, Chen M, Otten GR, Ulmer JB, Donnelly JJ, Ott G, et al. Distribution of DNA vaccines determines their immunogenicity after intramuscular injection in mice. J Immunol 2000; 165:2850-8; PMID:10946318; http://dx.doi.org/10.4049/jimmunol.165.5.2850
- Casares S, Inaba K, Brumeanu TD, Steinman RM, Bona CA. Antigen presentation by dendritic cells after immunization with DNA encoding a major histocompatibility complex class II-restricted viral epitope. J Exp Med 1997; 186:1481-6; PMID:9348305; http://dx.doi.org/10.1084/jem.186.9.1481
- Chattergoon MA, Robinson TM, Boyer JD, Weiner DB. Specific immune induction following DNA-based immunization through in vivo transfection and activation of macrophages/antigen-presenting cells. J Immunol 1998; 160:5707-18; PMID:9637479
- Desmet CJ, Ishii KJ. Nucleic acid sensing at the interface between innate and adaptive immunity in vaccination. Nat Rev Immunol 2012; 12:479-91; PMID:22728526; http://dx.doi.org/10.1038/nri3247
- Anderson RJ, Schneider J. Plasmid DNA and viral vector-based vaccines for the treatment of cancer. Vaccine 2007; 25(Suppl 2):B24-34; PMID:17698262; http://dx.doi.org/10.1016/j.vaccine.2007.05.030
- Fioretti D, Iurescia S, Fazio VM, Rinaldi M. DNA vaccines: developing new strategies against cancer. J Biomed Biotechnol 2010; 2010:174378; PMID:20368780; http://dx.doi.org/10.1155/2010/174378
- Ercole CJ, Oliveira Filho EK. Microsurgical vasovasostomy under local anesthesia: an outpatient setting. Semin Urol 1992; 10:180-3; PMID:1523372
- Zhu D, Stevenson FK. DNA gene fusion vaccines against cancer. Curr Opin Mol Ther 2002; 4:41-8; PMID:11883694
- Yuan J, Ku GY, Gallardo HF, Orlandi F, Manukian G, Rasalan TS, Xu Y, Li H, Vyas S, Mu Z, et al. Safety and immunogenicity of a human and mouse gp100 DNA vaccine in a phase I trial of patients with melanoma. Cancer Immunity 2009; 9:5; PMID:19496531
- Staff C, Mozaffari F, Haller BK, Wahren B, Liljefors M. A Phase I safety study of plasmid DNA immunization targeting carcinoembryonic antigen in colorectal cancer patients. Vaccine 2011; 29:6817-22; PMID:21195077; http://dx.doi.org/10.1016/j.vaccine.2010.12.063
- Trimble CL, Peng S, Kos F, Gravitt P, Viscidi R, Sugar E, Pardoll D, Wu TC. A phase I trial of a human papillomavirus DNA vaccine for HPV16+ cervical intraepithelial neoplasia 2/3. Clin Cancer Res 2009; 15:361-7; PMID:19118066; http://dx.doi.org/10.1158/1078-0432.CCR-08-1725
- Fynan EF, Webster RG, Fuller DH, Haynes JR, Santoro JC, Robinson HL. DNA vaccines: protective immunizations by parenteral, mucosal, and gene-gun inoculations. Proc Natl Acad Sci U S A 1993; 90:11478-82
- Aihara H, Miyazaki J. Gene transfer into muscle by electroporation in vivo. Nat Biotechnol 1998; 16:867-70; PMID:9743122; http://dx.doi.org/10.1038/nbt0998-867
- Mathiesen I. Electropermeabilization of skeletal muscle enhances gene transfer in vivo. Gene Ther 1999; 6:508-14; PMID:10476210; http://dx.doi.org/10.1038/sj.gt.3300847
- Sardesai NY, Weiner DB. Electroporation delivery of DNA vaccines: prospects for success. Curr Opin Immunol 2011; 23:421-9; PMID:21530212; http://dx.doi.org/10.1016/j.coi.2011.03.008
- Ahlen G, Soderholm J, Tjelle T, Kjeken R, Frelin L, Hoglund U, Blomberg P, Fons M, Mathiesen I, Sallberg M. In vivo electroporation enhances the immunogenicity of hepatitis C virus nonstructural 3/4A DNA by increased local DNA uptake, protein expression, inflammation, and infiltration of CD3+ T cells. J Immunol 2007; 179:4741-53; PMID:17878373; http://dx.doi.org/10.4049/jimmunol.179.7.4741
- Liu J, Kjeken R, Mathiesen I, Barouch DH. Recruitment of antigen-presenting cells to the site of inoculation and augmentation of human immunodeficiency virus type 1 DNA vaccine immunogenicity by in vivo electroporation. J Virol 2008; 82:5643-9; PMID:18353952; http://dx.doi.org/10.1128/JVI.02564-07
- Widera G, Austin M, Rabussay D, Goldbeck C, Barnett SW, Chen M, Leung L, Otten GR, Thudium K, Selby MJ, et al. Increased DNA vaccine delivery and immunogenicity by electroporation in vivo. J Immunol 2000; 164:4635-40; PMID:10779767; http://dx.doi.org/10.4049/jimmunol.164.9.4635
- Babiuk S, Baca-Estrada ME, Foldvari M, Storms M, Rabussay D, Widera G, Babiuk LA. Electroporation improves the efficacy of DNA vaccines in large animals. Vaccine 2002; 20:3399-408; PMID:12213410; http://dx.doi.org/10.1016/S0264-410X(02)00269-4
- Tollefsen S, Tjelle T, Schneider J, Harboe M, Wiker H, Hewinson G, Huygen K, Mathiesen I. Improved cellular and humoral immune responses against Mycobacterium tuberculosis antigens after intramuscular DNA immunisation combined with muscle electroporation. Vaccine 2002; 20:3370-8; PMID:12213407; http://dx.doi.org/10.1016/S0264-410X(02)00289-X
- Low L, Mander A, McCann K, Dearnaley D, Tjelle T, Mathiesen I, Stevenson F, Ottensmeier CH. DNA vaccination with electroporation induces increased antibody responses in patients with prostate cancer. Hum Gene Ther 2009; 20:1269-78; PMID:19619001; http://dx.doi.org/10.1089/hum.2009.067
- Diaz CM, Chiappori A, Aurisicchio L, Bagchi A, Clark J, Dubey S, Fridman A, Fabregas JC, Marshall J, Scarselli E, et al. Phase 1 studies of the safety and immunogenicity of electroporated HER2/CEA DNA vaccine followed by adenoviral boost immunization in patients with solid tumors. J Translat Med 2013; 11:62; PMID:23497415; http://dx.doi.org/10.1186/1479-5876-11-62
- Leitner WW, Hwang LN, deVeer MJ, Zhou A, Silverman RH, Williams BR, Dubensky TW, Ying H, Restifo NP. Alphavirus-based DNA vaccine breaks immunological tolerance by activating innate antiviral pathways. Nat Med 2003; 9:33-9; PMID:12496961; http://dx.doi.org/10.1038/nm813
- Kang TH, Mao CP, La V, Chen A, Hung CF, Wu TC. Innovative DNA vaccine to break immune tolerance against tumor self-antigen. Hum Gene Ther 2013; 24:181-8; PMID:23268564; http://dx.doi.org/10.1089/hum.2012.141
- Soong RS, Trieu J, Lee SY, He L, Tsai YC, Wu TC, Hung CF. Xenogeneic human p53 DNA vaccination by electroporation breaks immune tolerance to control murine tumors expressing mouse p53. PloS One 2013; 8:e56912; PMID:23457640; http://dx.doi.org/10.1371/journal.pone.0056912
- Yan J, Pankhong P, Shin TH, Obeng-Adjei N, Morrow MP, Walters JN, Khan AS, Sardesai NY, Weiner DB. Highly optimized DNA vaccine targeting human telomerase reverse transcriptase stimulates potent antitumor immunity. Cancer Immunol Res 2013; 1:179-89; PMID:24777680; http://dx.doi.org/10.1158/2326-6066.CIR-13-0001
- Aurisicchio L, Peruzzi D, Koo G, Wei WZ, La Monica N, Ciliberto G. Immunogenicity and therapeutic efficacy of a dual-component genetic cancer vaccine cotargeting carcinoembryonic antigen and HER2/neu in preclinical models. Human Gene Therapy 2014; 25:121-31; PMID:24195644; http://dx.doi.org/10.1089/hum.2013.103
- Williams BB, Wall M, Miao RY, Williams B, Bertoncello I, Kershaw MH, Mantamadiotis T, Haber M, Norris MD, Gautam A, et al. Induction of T cell-mediated immunity using a c-Myb DNA vaccine in a mouse model of colon cancer. Cancer Immunol Immunother 2008; 57:1635-45; PMID:18386000; http://dx.doi.org/10.1007/s00262-008-0497-2
- Joseph-Pietras D, Gao Y, Zojer N, Ait-Tahar K, Banham AH, Pulford K, Rice J, Savelyeva N, Sahota SS. DNA vaccines to target the cancer testis antigen PASD1 in human multiple myeloma. Leukemia 2010; 24:1951-9; PMID:20861911; http://dx.doi.org/10.1038/leu.2010.196
- Campos-Perez J, Rice J, Escors D, Collins M, Paterson A, Savelyeva N, Stevenson FK. DNA fusion vaccine designs to induce tumor-lytic CD8+ T-cell attack via the immunodominant cysteine-containing epitope of NY-ESO 1. Int J Cancer 2013; 133:1400-7; PMID:23494538; http://dx.doi.org/10.1002/ijc.28156
- Gregor PD, Wolchok JD, Ferrone CR, Buchinshky H, Guevara-Patino JA, Perales MA, Mortazavi F, Bacich D, Heston W, Latouche JB, et al. CTLA-4 blockade in combination with xenogeneic DNA vaccines enhances T-cell responses, tumor immunity and autoimmunity to self antigens in animal and cellular model systems. Vaccine 2004; 22:1700-8; PMID:15068853; http://dx.doi.org/10.1016/j.vaccine.2003.10.048
- Cohen AD, Diab A, Perales MA, Wolchok JD, Rizzuto G, Merghoub T, Huggins D, Liu C, Turk MJ, Restifo NP, et al. Agonist anti-GITR antibody enhances vaccine-induced CD8(+) T-cell responses and tumor immunity. Cancer Res 2006; 66:4904-12; PMID:16651447; http://dx.doi.org/10.1158/0008-5472.CAN-05-2813
- Soong RS, Song L, Trieu J, Lee SY, He L, Tsai YC, Wu TC, Hung CF. Direct T cell activation via CD40 ligand generates high avidity CD8+ T cells capable of breaking immunological tolerance for the control of tumors. PloS One 2014; 9:e93162; PMID:24664420; http://dx.doi.org/10.1371/journal.pone.0093162
- Bergman PJ, McKnight J, Novosad A, Charney S, Farrelly J, Craft D, Wulderk M, Jeffers Y, Sadelain M, Hohenhaus AE, et al. Long-term survival of dogs with advanced malignant melanoma after DNA vaccination with xenogeneic human tyrosinase: a phase I trial. Clin Cancer Res 2003; 9:1284-90; PMID:12684396
- Bergman PJ. Canine oral melanoma. Clin Tech Small Anim Pract 2007; 22:55-60; PMID:17591290; http://dx.doi.org/10.1053/j.ctsap.2007.03.004
- USDA licenses DNA vaccine for treatment of melanoma in dogs. J Am Vet Med Assoc 2010; 236:495; PMID:20187810; http://dx.doi.org/10.2460/javma.236.5.488
- Ledwith BJ, Manam S, Troilo PJ, Barnum AB, Pauley CJ, Griffiths TG, 2nd, Harper LB, Beare CM, Bagdon WJ, Nichols WW. Plasmid DNA vaccines: investigation of integration into host cellular DNA following intramuscular injection in mice. Intervirology 2000; 43:258-72; PMID:11251381; http://dx.doi.org/10.1159/000053993
- Weber J, Boswell W, Smith J, Hersh E, Snively J, Diaz M, Miles S, Liu X, Obrocea M, Qiu Z, et al. Phase 1 trial of intranodal injection of a Melan-A/MART-1 DNA plasmid vaccine in patients with stage IV melanoma. J Immunother 2008; 31:215-23; PMID:18481391; http://dx.doi.org/10.1097/CJI.0b013e3181611420
- Ginsberg BA, Gallardo HF, Rasalan TS, Adamow M, Mu Z, Tandon S, Bewkes BB, Roman RA, Chapman PB, Schwartz GK, et al. Immunologic response to xenogeneic gp100 DNA in melanoma patients: comparison of particle-mediated epidermal delivery with intramuscular injection. Clin Cancer Res 2010; 16:4057-65; PMID:20647477; http://dx.doi.org/10.1158/1078-0432.CCR-10-1093
- Norell H, Poschke I, Charo J, Wei WZ, Erskine C, Piechocki MP, Knutson KL, Bergh J, Lidbrink E, Kiessling R. Vaccination with a plasmid DNA encoding HER-2/neu together with low doses of GM-CSF and IL-2 in patients with metastatic breast carcinoma: a pilot clinical trial. J Translat Med 2010; 8:53; PMID:20529245; http://dx.doi.org/10.1186/1479-5876-8-53
- Occhipinti S, Sponton L, Rolla S, Caorsi C, Novarino A, Donadio M, Bustreo S, Satolli MA, Pecchioni C, Marchini C, et al. Chimeric rat/human HER2 efficiently circumvents HER2 tolerance in cancer patients. Clin Cancer Res 2014; 20:2910-21; PMID:24668647; http://dx.doi.org/10.1158/1078-0432.CCR-13-2663
- Tiriveedhi V, Fleming TP, Goedegebuure PS, Naughton M, Ma C, Lockhart C, Gao F, Gillanders WE, Mohanakumar T. Mammaglobin-A cDNA vaccination of breast cancer patients induces antigen-specific cytotoxic CD4+ICOShi T cells. Breast Cancer Res Treat 2013; 138:109-18; PMID:22678162; http://dx.doi.org/10.1007/s10549-012-2110-9
- McNeel DG, Dunphy EJ, Davies JG, Frye TP, Johnson LE, Staab MJ, Horvath DL, Straus J, Alberti D, Marnocha R, et al. Safety and immunological efficacy of a DNA vaccine encoding prostatic acid phosphatase in patients with stage D0 prostate cancer. J Clin Oncol 2009; 27:4047-54; PMID:19636017; http://dx.doi.org/10.1200/JCO.2008.19.9968
- Becker JT, Olson BM, Johnson LE, Davies JG, Dunphy EJ, McNeel DG. DNA vaccine encoding prostatic acid phosphatase (PAP) elicits long-term T-cell responses in patients with recurrent prostate cancer. J Immunother 2010; 33:639-47; PMID:20551832; http://dx.doi.org/10.1097/CJI.0b013e3181dda23e
- Chudley L, McCann K, Mander A, Tjelle T, Campos-Perez J, Godeseth R, Creak A, Dobbyn J, Johnson B, Bass P, et al. DNA fusion-gene vaccination in patients with prostate cancer induces high-frequency CD8(+) T-cell responses and increases PSA doubling time. Cancer Immunol Immunother 2012; 61:2161-70; PMID:22729556; http://dx.doi.org/10.1007/s00262-012-1270-0
- Bagarazzi ML, Yan J, Morrow MP, Shen X, Parker RL, Lee JC, Giffear M, Pankhong P, Khan AS, Broderick KE, et al. Immunotherapy against HPV16/18 generates potent TH1 and cytotoxic cellular immune responses. Sci Transl Med 2012; 4:155ra38; PMID:23052295; http://dx.doi.org/10.1126/scitranslmed.3004414
- Maldonado L, Teague JE, Morrow MP, Jotova I, Wu TC, Wang C, Desmarais C, Boyer JD, Tycko B, Robins HS, et al. Intramuscular therapeutic vaccination targeting HPV16 induces T cell responses that localize in mucosal lesions. Sci Transl Med 2014; 6:221ra13; PMID:24477000; http://dx.doi.org/10.1126/scitranslmed.3007323
- Occhipinti S, Sponton L, Rolla S, Caorsi C, Novarino A, Donadio M, Bustreo S, Satolli MA, Pecchioni C, Marchini C, et al. Chimeric rat/human HER2 efficiently circumvents HER2 tolerance in cancer patients. Clin Cancer Res 2014; PMID:24668647; http://dx.doi.org/10.1158/1078-0432.CCR-13-2663