Abstract
Plant pathogenic fungi utilize a series of complex infection structures, in particular the appressorium, to gain entry to and colonize plant tissue. As a consequence of the accumulation of huge quantities of glycerol in the cell the appressorium generates immense intracellular turgor pressure allowing the penetration peg of the appressorium to penetrate the leaf cuticle. Autophagic processes are ubiquitous in eukaryotic cells and facilitate the bulk degradation of macromolecules and organelles. The study of autophagic processes has been extended from the model yeast Saccharomyces cerevisiae to pathogenic fungi such as the rice blast fungus Magnaporthe oryzae. Significantly, null mutants for the expression of M. oryzae autophagy gene homologs lose their pathogenicity for infection of host plants. Clarification of the functions and network of interactions between the proteins expressed by M. oryzae autophagy genes will lead to a better understanding of the role of autophagy in fungal pathogenesis and help in the development of new strategies for disease control.
Introduction
The life span of cellular macromolecules is much shorter than that of the cell itself. Ideally cells should have mechanisms to “recycle” any reusable materials in order to reduce their dependence on the extracellular environment. The vacuole in fungi is a major site where recycling is executed. The materials catabolized in this acidic compartment may be of endogenous or exogenous origin having been delivered by autophagic processes or endocytosis, respectively. Thus, autophagic processes result in the transport of cellular contents to vacuoles for degradation, thereby contributing to the homeostatic balance between synthesis and degradation within cells. It is the action of the resident complement of acid hydrolases within vacuoles that degrades the material delivered into the vacuolar lumen. The molecular “building blocks” so recovered are reused in biosynthetic pathways, providing a mechanism by which cells can respond to nutrient stress effectively by reallocating resources for use in the most essential cellular processes.Citation1,Citation2
The yeast Saccharomyces cerevisiae, itself a member of the fungi, is arguably the best-studied model system by which to investigate the molecular mechanisms of autophagy.Citation2-Citation4 With respect to mechanism, there are several distinct autophagic processes which occur in yeast cells: macroautophagy, microautophagy, cytoplasm-to-vacuole targeting (Cvt)Citation5 and vacuolar import and degradation (Vid).Citation6,Citation7 The latter two processes are specific to yeast, not occurring in mammalian cells. By contrast chaperone-mediated autophagy (CMA), which occurs in mammals, does not occur in yeast.Citation8,Citation9
An improved understanding of the molecular mechanisms of plant fungal pathogenesis and their interface with autophagic processes will ultimately lead to better control of plant fungal diseases. Diverse plant organs produce different obstacles to infection by potential fungal pathogens, and therefore successful pathogens have evolved specific strategies, especially infection structures, that are able to break through host plant roots, stems, leaves, flowers or other special tissues. The appressorium is an example of such a special infection structure, which has been well studied in Magnaporthe oryzae and Colletotrichum spp.Citation10-Citation12 Appressorium development is initiated by a fungal spore landing on, and attaching to, the plant cuticle surface. From the attached spore a germ tube is produced that grows until it recognizes a suitable site and receives environmental stimuli to activate the intracellular molecular machinery for appressorium formation and penetration. Substantial mechanical force is generated in a mature appressorium and used to deliver a penetration peg through the plant surface. Fungal infection-related development is a starvation-induced process. Nitrogen limitation has been proposed as a key signal for activating the expression of virulence genes in plant pathogens.Citation13 The noteworthy discovery of the role of macroautophagy (hereafter autophagy), following its induction by nutrient starvation, in the development of turgor in the appressorium represents a milestone emphasizing the importance of, and functional connections between, autophagy and the formation of infection structures in plant pathogenic fungi.Citation14-Citation16 It appears likely that autophagy is a pivotal process in determining the outcome of the penetration stages by plant pathogenic fungi.Citation17 Here we will review current knowledge of autophagy in plant pathogenic fungi and the functional links between autophagy and plant fungal pathogenesis.
Infection Structures of Pathogenic Fungi
Plant organs have evolved various hurdles to frustrate infection by potential pathogenic fungi, and in response pathogens have therefore developed some powerful strategies to gain entry to the underlying tissues of prospective host plants. Unlike plant bacterial pathogens, which usually overcome the barriers presented by plant surfaces through the utilization of natural openings, such as stomata, or existing wounds, many plant fungal pathogens are able to actively initiate breaching of the plant cuticle. It should be noted that some plant fungal pathogens, for example, rust fungi, can enter host cells via the natural opening of stomata. Thus, during evolution fungal pathogens have evolved specialized hyphal structures that are active during host invasion, including the infection cushion, the appressorium, and the haustorium.Citation18-Citation21 The formation and development of such structures are of vital importance to the success of the fungal infection and the eventual development of disease symptoms by the host plant. These processes are controlled by several genes and influenced by environmental factors such as temperature, pH, etc.Citation12,Citation22-Citation25 In addition, various cell wall depolymerases at the infection site are also needed by some pathogenic fungi.Citation26 In particular, appressoria and associated melanized cell wall structures are critical for penetration of host cells.
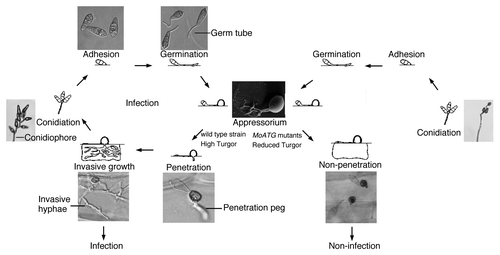
The development of an appressorium occurs as follows: A conidium of a plant fungal pathogen, for example M. oryzae or Colletotrium spp, will land on and attach to the leaf surface of its host plant, then germinate under suitable environmental conditions. The germ tube may sense a broad variety of signals derived from topographic information on leaf surfaces, physical stimuli and chemical substances produced by its host during growth.Citation20 These environmental signals will be accepted by membrane receptors of the germ tube or hyphal tip, transmitted into the fungal cell to activate appressorium development and morphogenesis through the action of certain molecular machinery, such as the adjustment of different components of the cytoskeleton, and anionic and/or electric changes mediated by mechanosensitive ion channels.Citation20 A typical life cycle of M. oryzae, which activates appressorium development during infection, is shown in .
Figure 1. Life cycle of Magnaporthe oryzae. Conidia germinate and develop a specialized infection structure, the appressorium. The structure produces a penetration peg, which will lead to invasive growth in and between host cells. This infection cycle is destroyed in the autophagy-blocked mutants.
Morphology and Physiology of the Appresorium
The defining feature of the appressorium is that it can generate huge turgor and directly rupture the plant cuticle, or even an artificial membrane that is not able to be degraded by biological enzymes, suggesting that a significant mechanical force is required for fungal infection.Citation10,Citation27-Citation29 Appressoria are usually clearly visible as dark differentiated structures at the tips of the germ tubes,Citation21 although there are instances in which they are difficult to distinguish morphologically. Under some circumstances, they may form from the hyphae. When an appressorium is separated from the germ tube by a septum, as in many Colletotrichum spp, M. oryzae, and Uromyces spp, the germ tube and spore frequently undergo autophagy, and nuclear degradation, and are devoid of cytoplasm.Citation15,Citation16,Citation30 The nucleus undergoes at least one round of mitosis and migrates with the cytoplasm into the appressorium.Citation16,Citation30 Verification of the role of autophagy is provided by analysis of the mgatg8∆ mutant, which is blocked in autophagy and forms nonfunctional appressoria.Citation16 Transfer of conidial cytoplasm into the developing appressoria is delayed in the autophagic null mutants.Citation15
For many fungi that directly penetrate the plant cuticle, the cell wall of the appressorium undergoes extensive modifications.Citation10,Citation31 In Colletotrichum spp and M. oryzae, the cell wall of the appressorium becomes thicker, multilayered, and highly melanized, but at the contact surface with the plant, the cell wall is usually less modified and thinner.Citation31 The dome-shaped appressorium with highly differentiated cell wall structures is rich in chitin and contains a melanin layer on the inner side of the cell wall. The melanin layer prevents efflux of cellular solutes and ensures the appressorium is able to accumulate substantial turgor for fungal penetration at a later time.Citation31-Citation33 The importance of melanin in infection structures is emphasized by the observation that knockout of genes in the melanin biosynthesis pathway leads to loss of the function of appressorium-mediated penetration and pathogenicity in M. oryzae.Citation31,Citation34,Citation35 Turgor pressure in the appressoria of melanin-deficient mutants is about 30–70% of that measured in fully melanized appressoria.Citation10
The force exerted during initial penetration of a single appressorium is substantial. The average penetration force of the C. graminicola appressorium reaches 17 μN, ranging from 8 to 25 μN. The pressure within the appressorium has been calculated to be 5.35 MPa, ~50 times atmospheric pressure. By comparison, the pressure of the M. oryzae appressorium is estimated to reach only a more modest 8 MPa corresponding to a force of 8 μN.Citation36 In C. graminicola, exertion of force typically starts 100 min after appressorium formation and reaches a steady level after 300 min. This level of force does not change for several hours.Citation37 These forces should be enough to breach the cuticle and epidermal cell wall of most monocotylous or dicotylous plants.
The enormous turgor pressure that develops in the appressorium can be ascribed to the intracellular accumulation of soluble carbohydrates such as glycerol, erythritol, arabinitol, mannitol and glucose.Citation38 In M. oryzae, it has been shown that turgor in the appressorium is a consequence of the accumulation of huge quantities of glycerol in a central vacuole of the cell.Citation21 Formation of a central vacuole filled with glycerol is not observed in M. oryzae mutants deleted for one of the MoATG2, MoATG4, MoATG5, MoATG9 or MoATG18 genes. Thus, it was confirmed that autophagy is essential for turgor generation in the appressoria of M. oryzae ().Citation14,Citation15,Citation39
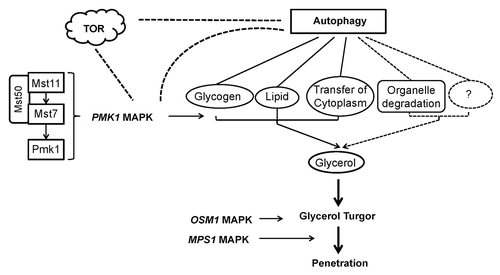
Figure 2. Model for the relationships between autophagy, appressorium turgor and the MAPK pathway in M. oryzae. In this model, the PMK1 MAPK pathway controls the mobilization of glycogen and lipid reserves to the developing appressorium. Glycerol accumulation is regulated by autophagy, transfer of cytoplasm, and degradation of glycogen, lipid, organelles, or other materials. Appressorium turgor is generated and hydrostatic turgor is translated into the force required for cuticle penetration. The OSM1 and MPS1 MAPK pathways may regulate development of turgor pressure and application of turgor pressure to penetration, respectively. Dashed lines indicate the conjectural relationships.
More generally, the deletion of ATG genes leads to morphological or physiological changes in the fungal life cycle, accompanied by reduced pathogenicity for the host. Though different fungal species may have similar lifestyles and pathogenic strategies, the control of basic cellular processes during fungal infection can be substantially different among species.Citation40 Thus far, the influence of individual ATG genes has been studied in relatively few pathogenic fungi, foremost among these is M. oryzae. In a moatg1∆ mutant, the appressorial turgor pressure is much lower than that in the wild type; only ~2.8% of the appressoria form long penetration pegs 24 h after inoculation, compared with ~49.7% in the wild type.Citation14 In a moatg4∆ mutant, both the conidial germination and appressorium formation rates are reduced. However, as time elapses the differences become slight, such that 24 h after inoculation, there is no significant difference in the rate of appressorium formation between the mutant and wild type.Citation40 Nevertheless, the mutant is more osmotically sensitive when exposed to 2 M glycerol for 10 min.Citation40 Transfer of conidial cytoplasm into the developing appressorium is delayed, but not blocked in the moatg5∆ mutant, which loses its pathogenicity.Citation15
Signaling Mechanisms Contributing to the Infection Structure
Mitogen-activated protein kinases (MAPKs) regulate many cellular processes in eukaryotic cells in response to extracellular stimuli.Citation41-Citation43 To date, five MAPK signaling pathways have been studied in S. cerevisiae.Citation44 Two MAPKs, Slt2 and Hog1, are required for mitophagy in S. cerevisiae.Citation45 The yeast Slt2 and several upstream components of its signal transduction pathway are necessary for pexophagy, but not for pexophagosome formation or other nonselective and selective forms of autophagy.Citation46 Three MAPK genes have been identified in M. oryzae: PMK1 (pathogenicity MAP kinase), MPS1 (MAP kinase for penetration and sporulation) and OSM1 (osmoregulation MAP kinase), that are homologous to S. cerevisiae FUS3 and KSS1, SLT2 and HOG1, respectively.Citation47-Citation49 Targeted gene disruption of M. oryzae PMK1 has revealed that the cognate MAPK signaling pathway is important for regulation of formation of the infection structure. Thus pmk1∆ mutants lose their capability for forming appressoria on some surfaces and are unable to rupture the rice cuticle to cause rice blast lesions.Citation47 Two upstream regulatory components, of this pathway, a MAPK kinase (MAPKK) Mst7, and a MAPKK kinase (MAPKKK) Mst11, have been identified in M. oryzae.Citation50 Functional analyses confirmed that both are indispensable for appressorium formation with Mst11 acting upstream of Mst7. Mst7 is responsible for Pmk1 phosphorylation and interacts physically with Pmk1 during formation of the appressorium by means of a conserved MAPK-docking site on Mst7; deletion of this site abolishes appressorium formation.Citation51 There is a sterile α-motif (SAM) domain found in the Mst11 MAPKKK, facilitating interaction with the SAM-containing protein Mst50, which acts as the adaptor or scaffold protein linking Mst11 with Mst7. Deletion of MST50 leads to inability to form appressoria and thus loss of pathogenicity.Citation52 While it is clear that this signaling pathway contributes to the development of the infection structure, as yet there are no data available that specifically link it with autophagy in plant pathogenic fungi ().
Molecular Mechanisms of Autophagy in Pathogenic Fungi
Progress in understanding the mechanistic basis of autophagy has been greatly facilitated by the discovery of the ATG genes and characterization of the encoded proteins. Studies in S. cervisiae have identified 36 ATG genes required for autophagic processes.Citation3 Most of these genes are also found in the filamentous fungi. The homologs of proteins involved in autophagic processes in several plant pathogenic fungi, including Phaeosphaeria nodorum (Pn), Sclerotinia sclerotiorum (Ss), Puccinia graminis (Pg), Botrytis cinerea (Bc), M. oryzae (Mo), and U. maydis (Um), are given in . These proteins are ubiquitous in plant pathogenic fungi. However, only a limited number of functional studies concerning autophagic processes have been performed in pathogenic fungi.
Table 1. Homologs of proteins involved in autophagy in plant pathogenic fungi
1. Magnaporthe oryzae
Autophagy contributes to development and differentiation in M. oryzae.Citation14,Citation15 Evidence for the involvement of 24 autophagy-related genes in the pathogenicity of M. oryzae has been provided by genome-wide functional analysis.Citation53 Nonselective autophagy plays a key role in the pathogenicity of M. oryzae. Deletion of any of the genes necessary for nonselective autophagy (e.g MoATG1 to MoATG10, MoATG12, MoATG15, MoATG16, and MoATG18) renders the mutant fungi unable to cause rice blast. In contrast, deletion of any of the genes necessary for selective autophagy does not affect pathogenicity. Thus, the moatg11∆, moatg24∆, moatg26∆, moatg27∆, moatg28∆, and moatg29∆ mutants are not severely impaired in their ability to cause disease.
We now consider studies concerning autophagic processes in M. oryzae in the context of established findings in the yeast S. cerevisiae.
A. Nitrogen and TOR signaling
A Ser/Thr protein kinase, TOR (target of rapamycin), acts as a central regulator of autophagy and mediates the response of cells to nutrient starvation. The function of TOR is mediated by two distinct multiprotein complexes, TOR complex (TORC) 1 and 2. It is TORC1 which is thought to couple growth signals to cellular metabolism.Citation54 This complex is particularly inhibited by rapamycin, and therefore rapamycin is an activator of macroautophagy that is commonly used in yeast (and other species since the structure and function of TORC1 is conserved in evolution). In yeast, TORC1 functions as a switch between macroautophagy and the Cvt pathway by altering phosphorylation of Atg13.Citation55 Under normal growth conditions TORC1 directly phosphorylates Atg13, which results in reduced Atg1 kinase activity. Inhibition of TOR by rapamycin causes a nutrient stress response, including inhibition of translation initiation, arrest in the G1 phase of the cell cycle, glycogen accumulation, downregulation of glycolysis, and autophagy.Citation56 Nitrogen is a particularly important nutrient in TOR signaling. In M. oryzae, MPG1 encoding a hydrophobin required for pathogenicity is strongly upregulated under conditions of nitrogen limitation.Citation57 Invasive growth is affected by nitrogen sources. Many genes identified in screens for nitrogen starvation-induced transcripts are also upregulated during plant infection.Citation58 Thus, nitrogen source seems to act as a metabolic switch to trigger expression of infection-related genes in the rice blast fungus. However, the role of TOR in nutrient regulation of fungal virulence on plants has not been examined so far.
The Snf1 kinase complex belongs to a highly conserved family of serine/threonine protein kinases and is involved in glycogen biosynthesis, lipid biosynthesis, regulation of general stress responses and autophagy in yeast.Citation59 Snf1 and Pho85 act as positive and negative regulators of autophagy, respectively, via Atg1 and Atg13.Citation60 Homologs of each of the Snf1 kinase complex subunits (Snf1, Snf4, Sip1, Sip2, and Gal83) have been found in M. oryzae. MoSNF1, a homolog of yeast SNF1, contributes to growth of the colony, shape and germination of the conidia, formation of the appressoria and pathogenicity of the rice blast fungus.Citation61 However, there are no reports about the relationship between the Snf1 kinase complex and autophagy.
B. The Atg1-Atg13 kinase complex
The yeast proteins Atg1, Atg13 and Atg17 form an Atg1 protein kinase complex.Citation62 In M. oryzae, the MoATG1 gene encodes a Ser/Thr protein kinase, which is highly conserved among eukaryotes. Disruption of the MoATG1 gene significantly affects fungal ability to survive under starvation conditions, with the mutant showing reduced conidiation, conidial germination, lipid turnover, and appressorium turgor. As a result, the moatg1∆ mutant loses its penetration ability and pathogenicity for rice and barley.Citation14 Notably, in each of the moatg13∆ and moatg17∆ mutants, autophagy was not completely blocked and pathogenicity was not affected (unpublished data). Furthermore, MoAtg9 localized to multiple punctate structures under control of the MoAtg1 protein, and these multiple dots tended to pool into one central structure within/near the vacuoles of the moatg1∆ mutant. However, such a central structure is not detected in proximity of the vacuoles of moatg13∆ and moatg17∆ mutants.Citation63 These findings are not consistent with observations in yeast.Citation64,Citation65 It can be speculated that MoAtg13 and/or MoAtg17 has lost its function in autophagy in M. oryzae and therefore the Atg1-Atg13 complex might not be preserved across all fungi during evolution. Alternatively there may be functional homologs of ATG13 and ATG17 encoded in the M. oryzae genome, which are yet to be identified as such. Moreover, a gene encoding Atg31, which is also a constituent of the Atg1 kinase complex in yeast,Citation66 has not been identified in M. oryzae. Therefore, it is possible that novel specific molecular mechanisms, distinct from those in yeast, exist for the induction of autophagy in M. oryzae.
C. Ubiquitin-like conjugation systems
The yeast autophagy pathway requires two ubiquitin-like conjugation systems, Atg8 and Atg12–Atg5-Atg16, for autophagosome maturation.Citation67 It was reported that moatg8∆ mutants are unable to undergo conidial cell collapse and have lost their pathogenicity to the plant.Citation16 MoAtg8 is also involved in the regulation of glycogen metabolism during conidiogenesis.Citation53,Citation68 Analysis of the cellular localization pattern and flux of Atg8 is a reliable marker for autophagy in a variety of organisms.Citation69 Expression of an EGFP-MoAtg8 fusion is sufficient to complement the moatg8∆ mutant phenotypes, providing evidence that it is functional within M. oryzae, and therefore a reliable marker for analysis of the cellular pattern of autophagy.Citation53 Additionally, RFP-MoAtg8-PE has been established as a reliable marker for autophagosomes and autophagic vacuoles in aerial hyphae and conidiophores.Citation68
Atg8 undergoes C-terminal cleavage by the cysteine protease activity of Atg4 to expose a glycine residue in yeast.Citation70 Yeast complementation revealed that MoAtg4 can functionally complement the defects of a yeast ATG4 deletion mutant. The direct interaction between MoAtg4 and MoAtg8 is detected in both yeast two-hybrid and bimolecular fluorescence complementation (BiFC) assays. BiFC data indicated also that the MoAtg4-MoAtg8 interaction is enhanced only during nitrogen starvation. Recombinant MoAtg4 harboring a substitution of Cys to Ser at position 206 (MoAtg4C206S) is unable to cleave MoAtg8 in vitro, either in the absence or presence of DTT. These data suggest that Cys206 is part of the active site of MoAtg4Citation40 and show that MoAtg4 is a conserved cysteine protease required for autophagy.
In yeast the Atg12–Atg5 complex interacts with Atg16 to form a complex of 2:2:2 stoichiometry based on homodimerization of Atg16,Citation71 which localizes to the phagophore assembly site (PAS).Citation72,Citation73 The Atg12–Atg5-Atg16 complex is not associated with the completed autophagosome,Citation74 dissociating just prior to, or after, completion of the fusion event that finalizes autophagosome formation.Citation3 The MoATG5 gene encodes a conserved domain that is essential for autophagosome formation in M. oryzae. Deletion of the MoATG5 gene shows defective autophagy, shortened life span of aerial hyphae, reduced conidiation and perithecia formation, delayed germination of conidia and slower transfer of conidial cytoplasm, and reduction in appressorial turgor. As a result, the moatg5∆ mutant loses the ability to penetrate and infect rice and barley.Citation15 Laser excitation epifluorescence microscopy showed that the GFP-MoAtg5 protein is distributed evenly throughout cells accompanied by the presence of some sharply defined puncta in the cytoplasm of conidia, mycelia and appressoria. MoAtg8 localizes to the PAS as reported previously.Citation63 When a DsRed2-MoAtg8 fusion protein is expressed in an moatg5∆ mutant also expressing GFP-MoAtg5, then the DsRed2-MoAtg8 protein colocalizes with GFP-MoAtg5 puncta. The GFP-MoAtg5 and DsRed2-MoAtg8 fusion proteins are colocalized in the perivacuolar region in M. oryzae (unpublished data) consistent with at least transient localization to the PAS.
D. Atg9 cycling system
An Atg9 complex that includes Atg2 and Atg18 also seems to be involved in the autophagy of pathogenic fungi. In yeast, Atg9 is a transmembrane protein localized not only at the PAS, but also at cytosolic punctate compartments whose identity remains uncertain,Citation75 but may be tubulovesicular clusters.Citation76 The recycling of Atg9 from the PAS to its storage sites was demonstrated to be blocked in an Atg1-deficient mutant, and loss of Atg2, Atg13, Atg14 or Atg18 appears to produce a similar phenotype with regard to their effect on Atg9 retrograde movement in yeast.Citation64 When GFP-MoAtg9 is expressed in wild-type conidial and appressoria cells of M. oryzae, multiple green puncta are detected. The distribution pattern of EGFP-MgAtg9 is changed in these cells in moatg1∆, moatg2∆ or moatg18∆ mutants with prominent large green puncta that colocalize with DsRed2-MoAtg8 being detected. By contrast, the expression pattern of the same fusion proteins is not changed in a moatg13∆ mutant.Citation63 It was concluded that cycling of MgAtg9 through multiple sites (as indicated by colocalization studies) to its storage pools in the cytosol of M. oryzae depends on MoATG1, MoATG2, and MoATG18, but not MoATG13. Loss of MoATG1, MoATG2 and MoATG18 might prevent MoATG9 from leaving the PAS-like structures in M. oryzae. Deletion of MoATG2 or MoATG18 also affects conidiation, the production of appressorium turgor, pathogenicity for rice and sexual reproductive ability.Citation14
E. PtdIns3K complex I
The PtdIns3K complex I, which consists of Vps30/Atg6, Atg14, Vps34 (the only PtdIns3K in yeast) and Vps15, plays a pivotal role at the PAS in yeast.Citation77,Citation78 Most of the core proteins in this complex are fully conserved in plant pathogenic fungi (). Intriguingly, the gene for one component of the complex, ATG14, remains to be definitively identified. A sequence having very weak similarity to yeast ATG14 has been identified in M. oryzae by performing a re-analysis of NCBI protein databases. This putative homolog of ATG14 contains a conserved Cys-rich motif at its N terminus.Citation79 Disruption of the putative ATG14 in M. oryzae produced a highly similar phenotype to the moatg1∆ mutant (unpublished data).
F. SNAREs and HOPs
In yeast, docking and fusion of autophagosomes with the vacuolar membrane requires the SNARE proteins Vam3 and Vti1 (found on the vacuolar membrane), Vam7 and Ykt6 (found on the autophagosome), NSF, SNAP, Sec17, Sec18 and Sec19, the Rab protein Ypt7, and members of the class C Vps/HOPS complex.Citation80 Components of this machinery also mediate other intracellular vesicle fusion events, an essential cellular process of eukaryotic cells.Citation81 MoSec22 and MoVam7, the orthologs of yeast SNARE proteins Sec22 and Vam7, can also be demonstrated to have functions in vacuole assembly, which underlie the growth, conidiation, appressorium formation, and pathogenicity of M. oryzae.Citation82,Citation83 Similarly, MoMon1 and MoYpt7 are the orthologs of the yeast proteins Mon1 and Ypt7. The phenotypes of momon1∆ and moypt7∆ mutants share common features with the mosec22∆ mutant; they could not develop appressoria and had lost the ability to infect plants (unpublished data). Clearly, components of docking and fusion play key roles in vacuole formation and membrane trafficking that is linked to fungal pathogenicity.
2. Colletotrichum spp
In the cucumber anthracnose fungus C. orbiculare (syn. C. lagenarium) peroxisome degradation (pexophagy) occurs when it infects host plants. A random insertional mutagenesis screening was performed to identify genes involved in the pathogenesis of this fungus. A homolog of P. pastoris ATG26, which encodes a sterol glucosyltransferase that enhances pexophagy in this methylotrophic yeast, was isolated. The coatg26∆ mutant develops appressoria, but exhibits a specific defect in the subsequent host invasion step. Analysis using a GFP-tagged fusion protein suggests that CoAtg26 is localized at putative PAS sites. It has been proposed that CoAtg26 is involved in the regulation of the efficiency of pexophagy in the mature appressoria.Citation84 These data show that selective autophagy might be required during plant infection by C. orbiculare.
Unlike the coatg26∆ mutant, which is able to form appressoria, the coatg8∆ mutant is defective in the entire autophagic pathway and cannot form normal appressoria in the earlier steps of morphogenesis. By contrast, the moatg8∆ mutant of M. oryzae retains the ability to develop appressoria. This shows the diversity of function of some autophagy proteins and of the autophagy process as a whole in different plant pathogenic fungi.
Clk1, a homolog of yeast ATG1, was identified by a random insertional mutagenesis screen in C. lindemuthianum. For the clk1∆ mutant, very few lesions are produced on the intact leaves. However, a marked increase in the number of lesions is clearly visible 7 d after inoculation on wounded leaves, compared with intact leaves. However, even where the number of lesions on veins is much higher on wounded leaves, their extension is still limited and no maceration is observed. Clk1 transcripts are present both in pure cultures of the fungus and during the time-course of host infection, indicating the gene may be constitutively expressed. Furthermore, it was suggested that the signal transduction pathway involving the Clk1 protein kinase may be regulated either at a post-transcriptional level or through its substrates.Citation85
3. Fusarium graminearum
Autophagy plays critical roles in the pathogenicity of the Fusarium head blight fungus Fusarium graminearum. The FgATG15 gene encodes a lipolytic enzyme that plays an important role in the development of the fungus. Deletion of FgATG15 leads to defects in conidiogenesis, conidial shapes, germination, growth rate, and aerial hyphae formation. Under nutrient starvation conditions, the wild type degrades stored lipid droplets while the mutant loses this ability. Wheat head blight is severely attenuated in the fgatg15∆ mutant. Disease severity in the mutant is 9%, compared with 92% and 88% in the wild type and ectopic strains, respectively. The concentration of the mycotoxin deoxynivalenol (DON, a type B trichothecene, an epoxy-sesquiterpenoid) produced by the fgatg15∆ mutant is 55% less than that of the wild-type strain. These results imply that FgATG15 is involved in numerous developmental processes and could be exploited as an antifungal target.Citation86
Recently, a fgatg8∆ mutant was generated by gene replacement and found to be unable to form autophagic compartments. The fgatg8∆ mutant shows no formation of fruiting bodies (perithecia), reduced conidia production and collapse of its aerial mycelium after a few days in culture. The mutant contains lipid droplets indicative of nitrogen starvation and/or an inability to use storage lipids, suggesting autophagy-dependent lipid utilization (lipophagy) in this fungus. The capacity to reallocate nutrients and support nonassimilating fungal structures is reduced in the mutant. Although the ability to infect barley and wheat remains normal, the mutant is unable to spread from spikelet to spikelet in wheat. Collectively the data were interpreted to mean that autophagy provides a mechanism for supplying nutrients to nonassimilating structures necessary for growth and is important for plant colonization.Citation87
4. Ustilago maydis
Autophagy genes are important for the development and virulence of the corn smut fungus U. maydis. In U. maydis, umatg8∆ or umatg1∆ deletion mutants prevent the vacuolar accumulation of autophagic bodies and dramatically reduce survival under carbon-starvation conditions. Deletion of UmATG8 affects the budding of haploid sporidia, gall formation and teliospore production in ears of mature maize.Citation88 The umatg1∆ deletion mutants have phenotypic similarity to umatg8∆ deletion mutants, but with lower severity, such that umatg1∆ mutants are only slightly less pathogenic than the wild type, and teliospore (the thick-walled resting spore of U. maydis) production is not affected.Citation88 Moreover, in the double-deletion mutant, plant gall induction is completely suppressed.Citation88
5. Other filamentous fungi
Autophagy has been monitored in the filamentous fungus Podospora anserina. In this organism, autophagy can be induced by starvation or rapamycin treatment, or by heterokaryon incompatibility genes.Citation89,Citation90 Analysis of paatg1∆, paatg8∆, and pspA∆ mutants revealed that autophagy is essential for formation of aerial hyphae and for female organ differentiation, and is involved in spore germination.Citation91,Citation92
In the opportunistic mold pathogen, Aspergillus fumigatus, starvation-associated foraging has been studied. Hyphal plugs were transferred from rich medium onto starvation medium, thereby forcing the organism to use autophagy to fuel any further growth. When conidia from a strain expressing GFP-AfAtg8 are incubated in Aspergillus minimal medium the accumulation of autophagic bodies in vacuoles is observed by laser confocal microscopy.Citation93
In Aspergillus oryzae, a aoatg8∆ mutant was constructed and autophagy monitored by the expression of DsRed2-AoAtg8 and EGFP-AoAtg8 fusion proteins.Citation94 Under normal growth conditions the fusion proteins are localized in dot PAS-like structures, whereas starvation or rapamycin treatment cause their accumulation in vacuoles. Cytosolically expressed DsRed2 (not fused to AoAtg8) is also taken up into vacuoles under starvation conditions, or during the differentiation of conidiophores and conidial germination. The aoatg8∆ mutant does not form aerial hyphae and conidia, and DsRed2 is not localized in vacuoles under starvation conditions.Citation94 Subsequently aoatg4∆, aoatg13∆ and aoatg15∆ mutants were examined by following EGFP-AoAtg8 fluorescence.Citation95 In the aoatg13∆ mutant only limited accumulation of EGFP-AoAtg8 at the PAS, or in autophagosomes and vacuoles was observed. In the aoatg4∆ mutant no vacuolar uptake is detected and only PAS-like structures are detected, whereas in the aoatg15∆ mutant autophagic bodies accumulate in vacuoles. Conidiation in the aoatg4∆ and aoatg15∆ mutants is not detected, and it is decreased in the aoatg13∆ mutant compared with the wild-type strain. The aoatg15∆ mutant also displays a marked reduction in development of aerial hyphae. Thus, it is suggested that autophagy functions in both the differentiation of aerial hyphae and conidial germination in A. oryzae.Citation94
Autophagy plays key roles in fruiting-body development of the homothallic ascomycete Sordaria macrospora.Citation96 SmATG7 is markedly upregulated under amino acid starvation conditions and during late stages of sexual development. SmATG7 is essential for fungal viability, and autophagy is disturbed in Smatg7-RNAi mutants of S. macrospore, which produce aberrant fruiting bodies. These data provide a starting point for probing the diverse functions of autophagy in a sophisticated microbial genetic system.Citation96
Tricherdoma reesei (teleomorph Hypocrea jecorina), a soil-borne, green-spored ascomycete fungus has the ability to secrete large amounts of cellulolytic enzymes (cellulases and hemicellulases). The functions of TrAtg5, the homolog of yeast Atg5, were studied in a tratg5∆ mutant generated using targeted gene disruption. The mutant is blocked in autophagy and produces very few hyphal branches, with abnormal conidiophores and no more than two conidia on single abnormal conidiophores. By contrast, conidia aggregate into fascicles and pustules with plump conidiophores in the wild-type fungus. In addition, the tratg5∆ mutant shows reduced conidiation and sensitivity to nutrient materials.Citation97
Penicillium chrysogenum, is used for industrial production of penicillin (PEN) a β-lactam antibiotic. The penicillin biosynthesis pathway takes place in the cytosol and in peroxisomes in this fungus. Deletion of the P. chrysogenum ATG1 gene impairs autophagy and causes sporulation defects as in other filamentous fungi. Extensive autophagy-related degradation of cytosolic components and peroxisomes are observed to occur in vacuolated, late hyphal elements of P. chrysogenum. Cell degeneration is delayed and the levels of PEN biosynthesis enzymes are increased in the Pcatg1 null mutant. Collectively the data demonstrate that impairment of autophagy in P. chrysogenum leads to significantly increased production levels of PEN.Citation98
Candida albicans is considered a commensal organism of humans, colonizing the oral cavity, gastrointestinal, and reproductive tracts. However, when host defenses are compromised, C. albicans can transform into a tissue invasive pathogen. The role played by autophagy in facilitating asymptomatic host colonization, persistence, and transition of C. albicans into its pathogenic form has not been fully explored. Resistance to nitrogen starvation has been studied in two mutants, caatg9∆ and cavps11∆. While the former shows defects in autophagosome formation, the latter is blocked in vacuolar fusion. The caatg9∆ mutant survives within and kills a mouse macrophage-like cell line as efficiently as control strains, which suggests that autophagy plays little or no role in C. albicans differentiation, or during its interaction with mammalian host cells.Citation99
Cryptococcus neoformans is a yeast-like fungus that causes a lethal meningoencephalitis in a broad spectrum of immunocompromised patients. Survival of the fungus within the hostile and nutrient-deprived environments of the host has recently been shown to depend on the induction of autophagy.Citation100 PtdIns3K signaling of autophagy is required for starvation tolerance and virulence of this fungus. A cnatg8∆ mutant demonstrates markedly attenuated virulence in a mouse model of infection.Citation101
Concluding Remarks
Much has been learned about the morphological, physiological and biochemical characteristics of appressorium development and function during recent years. Autophagy is necessary for the formation of conidia and appressoria and for normal development and pathogenicity of Magnaporthe or Colletotrichum spp. It is likely that many factors influence the control of autophagy during pathogenesis. A problem in identifying ATG genes and their related functions in filamentous fungi is that functionally redundant genes exist in many fungal genomes. The significance of multiple gene copies in these fungi remains to be established.
The importance of autophagy for plant pathogenic fungi has a fundamental difference with its importance for yeasts because of the special penetration structure utilized in plant fungal infection. There are still many questions that need to be answered even concerning the autophagy machinery itself in pathogenic fungi. How might autophagy contribute to the development of turgor in the appressorium? Is the degradation of lipids and glycogen important for this process? What signaling pathways control autophagy in plant pathogenic fungi? Does the selective autophagy of organelles such as mitochondria, peroxisomes and endoplasmic reticulum contribute to the infectivity of plant pathogenic fungi?
Acknowledgment
This study was supported by grants (No. 30925029 and 31000077) from the National Natural Science Foundation of China to Fu-Cheng Lin and Xiao-Hong Liu, and by a grant (No. LQ12C14003) from the Zhejiang Provincial Natural Science Foundation of China to Fei Xu.
References
- Nakatogawa H, Suzuki K, Kamada Y, Ohsumi Y. Dynamics and diversity in autophagy mechanisms: lessons from yeast. Nat Rev Mol Cell Biol 2009; 10:458 - 67; http://dx.doi.org/10.1038/nrm2708; PMID: 19491929
- Wang C-W, Klionsky DJ. The molecular mechanism of autophagy. Mol Med 2003; 9:65 - 76; PMID: 12865942
- Mizushima N, Yoshimori T, Ohsumi Y. The role of Atg proteins in autophagosome formation. Annu Rev Cell Dev Biol 2011; 27:107 - 32; http://dx.doi.org/10.1146/annurev-cellbio-092910-154005; PMID: 21801009
- Inoue Y, Klionsky DJ. Regulation of macroautophagy in Saccharomyces cerevisiae.. Semin Cell Dev Biol 2010; 21:664 - 70; http://dx.doi.org/10.1016/j.semcdb.2010.03.009; PMID: 20359542
- Shintani T, Huang W-P, Stromhaug PE, Klionsky DJ. Mechanism of cargo selection in the cytoplasm to vacuole targeting pathway. Dev Cell 2002; 3:825 - 37; http://dx.doi.org/10.1016/S1534-5807(02)00373-8; PMID: 12479808
- Lynch-Day MA, Klionsky DJ. The Cvt pathway as a model for selective autophagy. FEBS Lett 2010; 584:1359 - 66; http://dx.doi.org/10.1016/j.febslet.2010.02.013; PMID: 20146925
- Brown CR, Dunton D, Chiang HL. The vacuole import and degradation pathway utilizes early steps of endocytosis and actin polymerization to deliver cargo proteins to the vacuole for degradation. J Biol Chem 2010; 285:1516 - 28; http://dx.doi.org/10.1074/jbc.M109.028241; PMID: 19892709
- Li W, Yang Q, Mao Z. Chaperone-mediated autophagy: machinery, regulation and biological consequences. Cell Mol Life Sci 2011; 68:749 - 63; http://dx.doi.org/10.1007/s00018-010-0565-6; PMID: 20976518
- Arias E, Cuervo AM. Chaperone-mediated autophagy in protein quality control. Curr Opin Cell Biol 2011; 23:184 - 9; http://dx.doi.org/10.1016/j.ceb.2010.10.009; PMID: 21094035
- Dean RA. Signal pathways and appressorium morphogenesis. Annu Rev Phytopathol 1997; 35:211 - 34; http://dx.doi.org/10.1146/annurev.phyto.35.1.211; PMID: 15012522
- Deising HB, Werner S, Wernitz M. The role of fungal appressoria in plant infection. Microbes Infect 2000; 2:1631 - 41; http://dx.doi.org/10.1016/S1286-4579(00)01319-8; PMID: 11113382
- Talbot NJ. On the trail of a cereal killer: Exploring the biology of Magnaporthe grisea.. Annu Rev Microbiol 2003; 57:177 - 202; http://dx.doi.org/10.1146/annurev.micro.57.030502.090957; PMID: 14527276
- Snoeijers SS, Perez-Garcia A, Joosten MHAJ, De Wit PJGM. The effect of nitrogen on disease development and gene expression in bacterial and fungal plant pathogens. Eur J Plant Pathol 2000; 106:493 - 506; http://dx.doi.org/10.1023/A:1008720704105
- Liu XH, Lu JP, Zhang L, Dong B, Min H, Lin FC. Involvement of a Magnaporthe grisea serine/threonine kinase gene, MgATG1, in appressorium turgor and pathogenesis. Eukaryot Cell 2007; 6:997 - 1005; http://dx.doi.org/10.1128/EC.00011-07; PMID: 17416896
- Lu JP, Liu XH, Feng XX, Min H, Lin FC. An autophagy gene, MgATG5, is required for cell differentiation and pathogenesis in Magnaporthe oryzae.. Curr Genet 2009; 55:461 - 73; http://dx.doi.org/10.1007/s00294-009-0259-5; PMID: 19629489
- Veneault-Fourrey C, Barooah M, Egan M, Wakley G, Talbot NJ. Autophagic fungal cell death is necessary for infection by the rice blast fungus. Science 2006; 312:580 - 3; http://dx.doi.org/10.1126/science.1124550; PMID: 16645096
- Talbot NJ, Kershaw MJ. The emerging role of autophagy in plant pathogen attack and host defence. Curr Opin Plant Biol 2009; 12:444 - 50; http://dx.doi.org/10.1016/j.pbi.2009.05.008; PMID: 19625208
- Staples RC. Nutrients for a rust fungus: the role of haustoria. Trends Plant Sci 2001; 6:496 - 8; http://dx.doi.org/10.1016/S1360-1385(01)02126-4; PMID: 11701352
- Szabo LJ, Bushnell WR. Hidden robbers: the role of fungal haustoria in parasitism of plants. Proc Natl Acad Sci U S A 2001; 98:7654 - 5; http://dx.doi.org/10.1073/pnas.151262398; PMID: 11438718
- Tucker SL, Talbot NJ. Surface attachment and pre-penetration stage development by plant pathogenic fungi. Annu Rev Phytopathol 2001; 39:385 - 417; http://dx.doi.org/10.1146/annurev.phyto.39.1.385; PMID: 11701871
- Howard RJ, Valent B. Breaking and entering: host penetration by the fungal rice blast pathogen Magnaporthe grisea.. Annu Rev Microbiol 1996; 50:491 - 512; http://dx.doi.org/10.1146/annurev.micro.50.1.491; PMID: 8905089
- Tucker SL, Talbot NJ. Surface attachment and pre-penetration stage development by plant pathogenic fungi. Annu Rev Phytopathol 2001; 39:385 - 417; http://dx.doi.org/10.1146/annurev.phyto.39.1.385; PMID: 11701871
- Tani M, Ishida N, Furusawa I. Effects of temperature and antibiotics on appressorium formation in spores of Colletotrichum lagenarium.. Can J Microbiol 1977; 23:626 - 9; http://dx.doi.org/10.1139/m77-091; PMID: 871968
- Leandro LF, Gleason ML, Nutter FW, Wegulo SN, Dixon PM. Influence of Temperature and Wetness Duration on Conidia and Appressoria of Colletotrichum acutatum on Symptomless Strawberry Leaves. Phytopathology 2003; 93:513 - 20; http://dx.doi.org/10.1094/PHYTO.2003.93.4.513; PMID: 18944367
- Miyara I, Shafran H, Davidzon M, Sherman A, Prusky D. pH Regulation of ammonia secretion by Colletotrichum gloeosporioides and its effect on appressorium formation and pathogenicity. Mol Plant Microbe Interact 2010; 23:304 - 16; http://dx.doi.org/10.1094/MPMI-23-3-0304; PMID: 20121452
- Kikot GE, Hours RA, Alconada TM. Contribution of cell wall degrading enzymes to pathogenesis of Fusarium graminearum: a review. J Basic Microbiol 2009; 49:231 - 41; http://dx.doi.org/10.1002/jobm.200800231; PMID: 19025875
- Greenberg JT. Programmed Cell Death in Plant-Pathogen Interactions. Annu Rev Plant Physiol Plant Mol Biol 1997; 48:525 - 45; http://dx.doi.org/10.1146/annurev.arplant.48.1.525; PMID: 15012273
- Wilson RA, Talbot NJ. Under pressure: investigating the biology of plant infection by Magnaporthe oryzae.. Nat Rev Microbiol 2009; 7:185 - 95; http://dx.doi.org/10.1038/nrmicro2032; PMID: 19219052
- de Jong JC, McCormack BJ, Smirnoff N, Talbot NJ. Glycerol generates turgor in rice blast. Nature 1997; 389:244; http://dx.doi.org/10.1038/38418
- Nesher I, Barhoom S, Sharon A. Cell cycle and cell death are not necessary for appressorium formation and plant infection in the fungal plant pathogen Colletotrichum gloeosporioides.. BMC Biol 2008; 6:9; http://dx.doi.org/10.1186/1741-7007-6-9; PMID: 18275611
- Howard RJ, Valent B. Breaking and entering: host penetration by the fungal rice blast pathogen Magnaporthe grisea.. Annu Rev Microbiol 1996; 50:491 - 512; http://dx.doi.org/10.1146/annurev.micro.50.1.491; PMID: 8905089
- Takano Y, Kubo Y, Kuroda I, Furusawa I. Temporal Transcriptional Pattern of Three Melanin Biosynthesis Genes, PKS1, SCD1, and THR1, in Appressorium-Differentiating and Nondifferentiating Conidia of Colletotrichum lagenarium.. Appl Environ Microbiol 1997; 63:351 - 4; PMID: 16535499
- Fernández-Alvarez A, Elías-Villalobos A, Ibeas JI. The O-mannosyltransferase PMT4 is essential for normal appressorium formation and penetration in Ustilago maydis.. Plant Cell 2009; 21:3397 - 412; http://dx.doi.org/10.1105/tpc.109.065839; PMID: 19880800
- Vidal-Cros A, Viviani F, Labesse G, Boccara M, Gaudry M. Polyhydroxynaphthalene reductase involved in melanin biosynthesis in Magnaporthe grisea. Purification, cDNA cloning and sequencing. Eur J Biochem 1994; 219:985 - 92; http://dx.doi.org/10.1111/j.1432-1033.1994.tb18581.x; PMID: 8112349
- Thompson JE, Fahnestock S, Farrall L, Liao DI, Valent B, Jordan DB. The second naphthol reductase of fungal melanin biosynthesis in Magnaporthe grisea: tetrahydroxynaphthalene reductase. J Biol Chem 2000; 275:34867 - 72; http://dx.doi.org/10.1074/jbc.M006659200; PMID: 10956664
- Money NP. Turgor pressure and the mechanics of fungal penetration. Can J Bot 1995; 73:96 - 102; http://dx.doi.org/10.1139/b95-231
- Bechinger C, Giebel KF, Schnell M, Leiderer P, Deising HB, Bastmeyer M. Optical measurements of invasive forces exerted by appressoria of a plant pathogenic fungus. Science 1999; 285:1896 - 9; http://dx.doi.org/10.1126/science.285.5435.1896; PMID: 10489364
- Beever RE, Laracy EP. Osmotic adjustment in the filamentous fungus Aspergillus nidulans.. J Bacteriol 1986; 168:1358 - 65; PMID: 3536874
- Liu XH, Lu JP, Lin FC. Autophagy during conidiation, conidial germination and turgor generation in Magnaporthe grisea.. Autophagy 2007; 3:472 - 3; PMID: 17495517
- Liu TB, Liu XH, Lu JP, Zhang L, Min H, Lin FC. The cysteine protease MoAtg4 interacts with MoAtg8 and is required for differentiation and pathogenesis in Magnaporthe oryzae.. Autophagy 2010; 6:74 - 85; http://dx.doi.org/10.4161/auto.6.1.10438; PMID: 19923912
- Xu JR. Map kinases in fungal pathogens. Fungal Genet Biol 2000; 31:137 - 52; http://dx.doi.org/10.1006/fgbi.2000.1237; PMID: 11273677
- Zhao X, Mehrabi R, Xu JR. Mitogen-activated protein kinase pathways and fungal pathogenesis. Eukaryot Cell 2007; 6:1701 - 14; http://dx.doi.org/10.1128/EC.00216-07; PMID: 17715363
- Rispail N, Soanes DM, Ant C, Czajkowski R, Grünler A, Huguet R, et al. Comparative genomics of MAP kinase and calcium-calcineurin signalling components in plant and human pathogenic fungi. Fungal Genet Biol 2009; 46:287 - 98; http://dx.doi.org/10.1016/j.fgb.2009.01.002; PMID: 19570501
- Chen RE, Thorner J. Function and regulation in MAPK signaling pathways: lessons learned from the yeast Saccharomyces cerevisiae.. Biochim Biophys Acta 2007; 1773:1311 - 40; http://dx.doi.org/10.1016/j.bbamcr.2007.05.003; PMID: 17604854
- Mao K, Wang K, Zhao M, Xu T, Klionsky DJ. Two MAPK-signaling pathways are required for mitophagy in Saccharomyces cerevisiae.. J Cell Biol 2011; 193:755 - 67; http://dx.doi.org/10.1083/jcb.201102092; PMID: 21576396
- Manjithaya R, Jain S, Farré JC, Subramani S. A yeast MAPK cascade regulates pexophagy but not other autophagy pathways. J Cell Biol 2010; 189:303 - 10; http://dx.doi.org/10.1083/jcb.200909154; PMID: 20385774
- Xu JR, Staiger CJ, Hamer JE. Inactivation of the mitogen-activated protein kinase Mps1 from the rice blast fungus prevents penetration of host cells but allows activation of plant defense responses. Proc Natl Acad Sci U S A 1998; 95:12713 - 8; http://dx.doi.org/10.1073/pnas.95.21.12713; PMID: 9770551
- Dixon KP, Xu JR, Smirnoff N, Talbot NJ. Independent signaling pathways regulate cellular turgor during hyperosmotic stress and appressorium-mediated plant infection by Magnaporthe grisea.. Plant Cell 1999; 11:2045 - 58; PMID: 10521531
- Bruno KS, Tenjo F, Li L, Hamer JE, Xu JR. Cellular localization and role of kinase activity of PMK1 in Magnaporthe grisea.. Eukaryot Cell 2004; 3:1525 - 32; http://dx.doi.org/10.1128/EC.3.6.1525-1532.2004; PMID: 15590826
- Zhao X, Kim Y, Park G, Xu JR. A mitogen-activated protein kinase cascade regulating infection-related morphogenesis in Magnaporthe grisea.. Plant Cell 2005; 17:1317 - 29; http://dx.doi.org/10.1105/tpc.104.029116; PMID: 15749760
- Zhao X, Xu JR. A highly conserved MAPK-docking site in Mst7 is essential for Pmk1 activation in Magnaporthe grisea.. Mol Microbiol 2007; 63:881 - 94; http://dx.doi.org/10.1111/j.1365-2958.2006.05548.x; PMID: 17214742
- Park G, Xue C, Zhao X, Kim Y, Orbach M, Xu JR. Multiple upstream signals converge on the adaptor protein Mst50 in Magnaporthe grisea.. Plant Cell 2006; 18:2822 - 35; http://dx.doi.org/10.1105/tpc.105.038422; PMID: 17056708
- Kershaw MJ, Talbot NJ. Genome-wide functional analysis reveals that infection-associated fungal autophagy is necessary for rice blast disease. Proc Natl Acad Sci U S A 2009; 106:15967 - 72; http://dx.doi.org/10.1073/pnas.0901477106; PMID: 19717456
- Noda T, Ohsumi Y. Tor, a phosphatidylinositol kinase homologue, controls autophagy in yeast. J Biol Chem 1998; 273:3963 - 6; http://dx.doi.org/10.1074/jbc.273.7.3963; PMID: 9461583
- Kamada Y, Funakoshi T, Shintani T, Nagano K, Ohsumi M, Ohsumi Y. Tor-mediated induction of autophagy via an Apg1 protein kinase complex. J Cell Biol 2000; 150:1507 - 13; http://dx.doi.org/10.1083/jcb.150.6.1507; PMID: 10995454
- Crespo JL, Powers T, Fowler B, Hall MN. The TOR-controlled transcription activators GLN3, RTG1, and RTG3 are regulated in response to intracellular levels of glutamine. Proc Natl Acad Sci U S A 2002; 99:6784 - 9; http://dx.doi.org/10.1073/pnas.102687599; PMID: 11997479
- Talbot NJ, Ebbole DJ, Hamer JE. Identification and characterization of MPG1, a gene involved in pathogenicity from the rice blast fungus Magnaporthe grisea.. Plant Cell 1993; 5:1575 - 90; PMID: 8312740
- Donofrio NM, Oh Y, Lundy R, Pan H, Brown DE, Jeong JS, et al. Global gene expression during nitrogen starvation in the rice blast fungus, Magnaporthe grisea.. Fungal Genet Biol 2006; 43:605 - 17; http://dx.doi.org/10.1016/j.fgb.2006.03.005; PMID: 16731015
- Hedbacker K, Carlson M. SNF1/AMPK pathways in yeast. Front Biosci 2008; 13:2408 - 20; http://dx.doi.org/10.2741/2854; PMID: 17981722
- Wang Z, Wilson WA, Fujino MA, Roach PJ. Antagonistic controls of autophagy and glycogen accumulation by Snf1p, the yeast homolog of AMP-activated protein kinase, and the cyclin-dependent kinase Pho85p. Mol Cell Biol 2001; 21:5742 - 52; http://dx.doi.org/10.1128/MCB.21.17.5742-5752.2001; PMID: 11486014
- Yi M, Park JH, Ahn JH, Lee YH. MoSNF1 regulates sporulation and pathogenicity in the rice blast fungus Magnaporthe oryzae.. Fungal Genet Biol 2008; 45:1172 - 81; http://dx.doi.org/10.1016/j.fgb.2008.05.003; PMID: 18595748
- Kabeya Y, Kamada Y, Baba M, Takikawa H, Sasaki M, Ohsumi Y. Atg17 functions in cooperation with Atg1 and Atg13 in yeast autophagy. Mol Biol Cell 2005; 16:2544 - 53; http://dx.doi.org/10.1091/mbc.E04-08-0669; PMID: 15743910
- Dong B, Liu XH, Lu JP, Zhang FS, Gao HM, Wang HK, et al. MgAtg9 trafficking in Magnaporthe oryzae.. Autophagy 2009; 5:946 - 53; http://dx.doi.org/10.4161/auto.5.7.9161; PMID: 19556868
- Reggiori F, Tucker KA, Stromhaug PE, Klionsky DJ. The Atg1-Atg13 complex regulates Atg9 and Atg23 retrieval transport from the pre-autophagosomal structure. Dev Cell 2004; 6:79 - 90; http://dx.doi.org/10.1016/S1534-5807(03)00402-7; PMID: 14723849
- Yen W-L, Legakis JE, Nair U, Klionsky DJ. Atg27 is required for autophagy-dependent cycling of Atg9. Mol Biol Cell 2007; 18:581 - 93; http://dx.doi.org/10.1091/mbc.E06-07-0612; PMID: 17135291
- Kabeya Y, Noda NN, Fujioka Y, Suzuki K, Inagaki F, Ohsumi Y. Characterization of the Atg17-Atg29-Atg31 complex specifically required for starvation-induced autophagy in Saccharomyces cerevisiae.. Biochem Biophys Res Commun 2009; 389:612 - 5; http://dx.doi.org/10.1016/j.bbrc.2009.09.034; PMID: 19755117
- Ohsumi Y, Mizushima N. Two ubiquitin-like conjugation systems essential for autophagy. Semin Cell Dev Biol 2004; 15:231 - 6; http://dx.doi.org/10.1016/j.semcdb.2003.12.004; PMID: 15209383
- Deng YZ, Ramos-Pamplona M, Naqvi NI. Autophagy-assisted glycogen catabolism regulates asexual differentiation in Magnaporthe oryzae.. Autophagy 2009; 5:33 - 43; http://dx.doi.org/10.4161/auto.5.1.7175; PMID: 19115483
- Klionsky DJ, Abeliovich H, Agostinis P, Agrawal DK, Aliev G, Askew DS, et al. Guidelines for the use and interpretation of assays for monitoring autophagy in higher eukaryotes. Autophagy 2008; 4:151 - 75; PMID: 18188003
- Geng J, Klionsky DJ. The Atg8 and Atg12 ubiquitin-like conjugation systems in macroautophagy. ‘Protein modifications: beyond the usual suspects’ review series. EMBO Rep 2008; 9:859 - 64; http://dx.doi.org/10.1038/embor.2008.163; PMID: 18704115
- Fujioka Y, Noda NN, Nakatogawa H, Ohsumi Y, Inagaki F. Dimeric coiled-coil structure of Saccharomyces cerevisiae Atg16 and its functional significance in autophagy. J Biol Chem 2010; 285:1508 - 15; http://dx.doi.org/10.1074/jbc.M109.053520; PMID: 19889643
- Matsushita M, Suzuki NN, Obara K, Fujioka Y, Ohsumi Y, Inagaki F. Structure of Atg5.Atg16, a complex essential for autophagy. J Biol Chem 2007; 282:6763 - 72; http://dx.doi.org/10.1074/jbc.M609876200; PMID: 17192262
- Suzuki K, Kubota Y, Sekito T, Ohsumi Y. Hierarchy of Atg proteins in pre-autophagosomal structure organization. Genes Cells 2007; 12:209 - 18; http://dx.doi.org/10.1111/j.1365-2443.2007.01050.x; PMID: 17295840
- Suzuki K, Kirisako T, Kamada Y, Mizushima N, Noda T, Ohsumi Y. The pre-autophagosomal structure organized by concerted functions of APG genes is essential for autophagosome formation. EMBO J 2001; 20:5971 - 81; http://dx.doi.org/10.1093/emboj/20.21.5971; PMID: 11689437
- Mari M, Griffith J, Rieter E, Krishnappa L, Klionsky DJ, Reggiori F. An Atg9-containing compartment that functions in the early steps of autophagosome biogenesis. J Cell Biol 2010; 190:1005 - 22; http://dx.doi.org/10.1083/jcb.200912089; PMID: 20855505
- Nair U, Jotwani A, Geng J, Gammoh N, Richerson D, Yen W-L, et al. SNARE proteins are required for macroautophagy. Cell 2011; 146:290 - 302; http://dx.doi.org/10.1016/j.cell.2011.06.022; PMID: 21784249
- Strahl T, Thorner J. Synthesis and function of membrane phosphoinositides in budding yeast, Saccharomyces cerevisiae.. Biochim Biophys Acta 2007; 1771:353 - 404; PMID: 17382260
- Kihara A, Noda T, Ishihara N, Ohsumi Y. Two distinct Vps34 phosphatidylinositol 3-kinase complexes function in autophagy and carboxypeptidase Y sorting in Saccharomyces cerevisiae.. J Cell Biol 2001; 152:519 - 30; http://dx.doi.org/10.1083/jcb.152.3.519; PMID: 11157979
- Bartoszewska M, Kiel JAKW. The role of macroautophagy in development of filamentous fungi. Antioxid Redox Signal 2011; 14:2271 - 87; http://dx.doi.org/10.1089/ars.2010.3528; PMID: 20712412
- Yorimitsu T, Klionsky DJ. Autophagy: molecular machinery for self-eating. Cell Death Differ 2005; 12:Suppl 2 1542 - 52; http://dx.doi.org/10.1038/sj.cdd.4401765; PMID: 16247502
- Ostrowicz CW, Meiringer CT, Ungermann C. Yeast vacuole fusion: a model system for eukaryotic endomembrane dynamics. Autophagy 2008; 4:5 - 19; PMID: 17932463
- Dou X, Wang Q, Qi Z, Song W, Wang W, Guo M, et al. MoVam7, a conserved SNARE involved in vacuole assembly, is required for growth, endocytosis, ROS accumulation, and pathogenesis of Magnaporthe oryzae.. PLoS One 2011; 6:e16439; http://dx.doi.org/10.1371/journal.pone.0016439; PMID: 21283626
- Song W, Dou X, Qi Z, Wang Q, Zhang X, Zhang H, et al. R-SNARE homolog MoSec22 is required for conidiogenesis, cell wall integrity, and pathogenesis of Magnaporthe oryzae.. PLoS One 2010; 5:e13193; http://dx.doi.org/10.1371/journal.pone.0013193; PMID: 20949084
- Asakura M, Ninomiya S, Sugimoto M, Oku M, Yamashita S, Okuno T, et al. Atg26-mediated pexophagy is required for host invasion by the plant pathogenic fungus Colletotrichum orbiculare.. Plant Cell 2009; 21:1291 - 304; http://dx.doi.org/10.1105/tpc.108.060996; PMID: 19363139
- Dufresne M, Bailey JA, Dron M, Langin T. clk1, a serine/threonine protein kinase-encoding gene, is involved in pathogenicity of Colletotrichum lindemuthianum on common bean. Mol Plant Microbe Interact 1998; 11:99 - 108; http://dx.doi.org/10.1094/MPMI.1998.11.2.99; PMID: 9450334
- Nguyen LN, Bormann J, Le GT, Stärkel C, Olsson S, Nosanchuk JD, et al. Autophagy-related lipase FgATG15 of Fusarium graminearum is important for lipid turnover and plant infection. Fungal Genet Biol 2011; 48:217 - 24; http://dx.doi.org/10.1016/j.fgb.2010.11.004; PMID: 21094265
- Josefsen L, Droce A, Sondergaard TE, Sørensen JL, Bormann J, Schäfer W, et al. Autophagy provides nutrients for nonassimilating fungal structures and is necessary for plant colonization but not for infection in the necrotrophic plant pathogen Fusarium graminearum.. Autophagy 2012; 8:326 - 37; http://dx.doi.org/10.4161/auto.18705; PMID: 22240663
- Nadal M, Gold SE. The autophagy genes ATG8 and ATG1 affect morphogenesis and pathogenicity in Ustilago maydis.. Mol Plant Pathol 2010; 11:463 - 78; http://dx.doi.org/10.1111/j.1364-3703.2010.00620.x; PMID: 20618705
- Dementhon K, Paoletti M, Pinan-Lucarré B, Loubradou-Bourges N, Sabourin M, Saupe SJ, et al. Rapamycin mimics the incompatibility reaction in the fungus Podospora anserina.. Eukaryot Cell 2003; 2:238 - 46; http://dx.doi.org/10.1128/EC.2.2.238-246.2003; PMID: 12684373
- Pinan-Lucarré B, Iraqui I, Clavé C. Podospora anserina target of rapamycin. Curr Genet 2006; 50:23 - 31; http://dx.doi.org/10.1007/s00294-006-0064-3; PMID: 16614869
- Pinan-Lucarré B, Balguerie A, Clavé C. Accelerated cell death in Podospora autophagy mutants. Eukaryot Cell 2005; 4:1765 - 74; http://dx.doi.org/10.1128/EC.4.11.1765-1774.2005; PMID: 16278443
- Pinan-Lucarré B, Clavé C. Monitoring autophagy in the filamentous fungus Podospora anserina. Methods Enzymol 2008; 451:251 - 70; http://dx.doi.org/10.1016/S0076-6879(08)03218-7; PMID: 19185726
- Richie DL, Askew DS. Autophagy in the filamentous fungus Aspergillus fumigatus.. Methods Enzymol 2008; 451:241 - 50; http://dx.doi.org/10.1016/S0076-6879(08)03217-5; PMID: 19185725
- Kikuma T, Ohneda M, Arioka M, Kitamoto K. Functional analysis of the ATG8 homologue Aoatg8 and role of autophagy in differentiation and germination in Aspergillus oryzae.. Eukaryot Cell 2006; 5:1328 - 36; http://dx.doi.org/10.1128/EC.00024-06; PMID: 16896216
- Kikuma T, Kitamoto K. Analysis of autophagy in Aspergillus oryzae by disruption of Aoatg13, Aoatg4, and Aoatg15 genes. FEMS Microbiol Lett 2011; 316:61 - 9; http://dx.doi.org/10.1111/j.1574-6968.2010.02192.x; PMID: 21204928
- Nolting N, Bernhards Y, Pöggeler S. SmATG7 is required for viability in the homothallic ascomycete Sordaria macrospora.. Fungal Genet Biol 2009; 46:531 - 42; http://dx.doi.org/10.1016/j.fgb.2009.03.008; PMID: 19351563
- Liu XH, Yang J, He RL, Lu JP, Zhang CL, Lu SL, et al. An autophagy gene, TrATG5, affects conidiospore differentiation in Trichoderma reesei.. Res Microbiol 2011; 162:756 - 63; http://dx.doi.org/10.1016/j.resmic.2011.06.011; PMID: 21740968
- Bartoszewska M, Kiel JA, Bovenberg RA, Veenhuis M, van der Klei IJ. Autophagy deficiency promotes beta-lactam production in Penicillium chrysogenum.. Appl Environ Microbiol 2011; 77:1413 - 22; http://dx.doi.org/10.1128/AEM.01531-10; PMID: 21169429
- Palmer GE, Kelly MN, Sturtevant JE. Autophagy in the pathogen Candida albicans.. Microbiology 2007; 153:51 - 8; http://dx.doi.org/10.1099/mic.0.2006/001610-0; PMID: 17185534
- Hu G, Gibbons J, Williamson PR. Analysis of autophagy during infections of Cryptococcus neoformans.. Methods Enzymol 2008; 451:323 - 42; http://dx.doi.org/10.1016/S0076-6879(08)03222-9; PMID: 19185730
- Hu G, Hacham M, Waterman SR, Panepinto J, Shin S, Liu X, et al. PI3K signaling of autophagy is required for starvation tolerance and virulenceof Cryptococcus neoformans.. J Clin Invest 2008; 118:1186 - 97; http://dx.doi.org/10.1172/JCI32053; PMID: 18259613