Abstract
Adhesion events mediated by cadherin and integrin adhesion receptors have fundamental roles in the maintenance of the physiological balance of epithelial tissues, and it is well established that perturbations in their normal functional activity and/or changes in their expression are associated with tumorigenesis. Over the last decades, increasing evidence of a dynamic collaborative interaction between these complexes through their shared interactions with cytoskeletal proteins and common signaling pathways has emerged not only as an important regulator of several aspects of epithelial cell behavior, but also as a coordinated adhesion module that senses and transmits signals from and to the epithelia surrounding microenvironment. The tight regulation of their crosstalk is particularly important during epithelial remodeling events that normally take place during morphogenesis and tissue repair, and when defective it leads to cell transformation and aggravated responses of the tumor microenvironment that contribute to tumorigenesis. In this review we highlight some of the interactions that regulate their crosstalk and how this could be implicated in regulating signals across epithelial tissues to sustain homeostasis.
Introduction
Epithelial tissues are highly specialized structures that shelter and separate body compartments. Their essential roles as protective barriers extend to additional fundamental functions including the absorption of solutes and nutrients, secretion and containment of body fluids. Epithelial sheets attach to their underlying basement membrane constituted by a supporting network of extracellular matrix proteins (ECM).Citation1 Epithelia are generally non-vascularized and receive their nutrients and oxygen from the underlying connective tissue or stroma, which is constituted by different cells types including resident immune cells, fat cells, blood vessels, fibroblasts and their secreted ECM. Under physiological conditions, epithelial homeostasis is maintained via precise control of cellular proliferation, in which the interactions between epithelial cells, and their communication with the underlying stroma plays a fundamental role. Defects in the functional interactions between cells and in their crosstalk with the surrounding tissue microenvironment are associated with increased epithelial proliferation and cancer.Citation2
Epithelial cells connect to their epithelial neighbors by an array of intercellular adhesion complexes; these physically sustain the maintenance of the epithelial barriers, but also participate in a wide spectrum of signaling pathways that regulate cell behavior. These complexes include adherens junctions (AJs), tight junctions (TJs) and desmosomes.Citation3 In addition, epithelial sheets attach to the underlying basement membrane through focal adhesion (FAs) and hemidesmosomes,Citation4-Citation6 which also provide signaling cues for the regulation of cell behavior, including cell polarity, proliferation and migration ().Citation1
Figure 1. Complexes implicated in cell-cell and cell-matrix adhesion. Cell-cell adhesion complexes: in blue, tight junctions (TJs) composed by the transmembrane proteins occludins, claudins (1–24), junctional adhesion molecule (JAM A–C), and the cytoplasmic proteins zonula occludens (ZO 1–3), the cell polarity protein partitioning defective-3 (PAR3), MUPP1 and MAG1. In green, adherens junctions (AJs) composed by Cadherin and Nectin complexes. The transmembrane protein E-cadherin binds to the cytoplasmic proteins p120 catenin, α-catenin and β-catenin, whereas Nectin binds to afadin. In orange, desmosomes are constituted by the transmembrane proteins desmoglein (DSG 1–4) and desmocollin (DSC 1–3), and the cytoplasmic proteins plakoglobin (JUP), plakophilins (PKP 1–3) and desmoplakin (DSP) that anchor to intermediate filaments. Cell-matrix adhesion complexes: in red, the hemidesmosomes (HD), formed by the transmembrane α- and β-integrins and the cytoplasmic proteins BP180, BO230 and plectin. In purple, the focal adhesion complex (FAs), composed by the transmembrane α- and β-integrins and cytoplasmic scaffolding proteins that associate to the actin cytoskeleton.
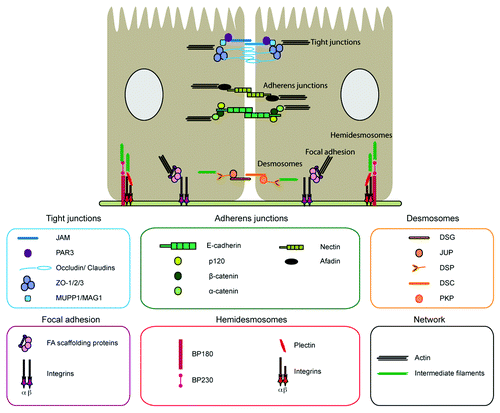
All these specialized adhesion structures present a paradigmatic organization; they are constituted by transmembrane adhesive glycoproteins that physically link epithelial cells to the extracellular matrix or to neighboring cells. Adhesion receptors, through their cytoplasmic domain, interact with scaffolding proteins that anchor these complexes to different cytoskeletal structures, including actin filaments, microtubules and keratin intermediate filaments, thus preserving architectural integrity across epithelial sheets ().
Without diminishing the key roles of other junctions in sustaining epithelial architecture and homeostasis, FAs and AJs are of particular relevance. FAs are unique in promoting the attachment of epithelial sheets to the underlying basement membrane, and AJs are crucial to control the specificity of cell adhesion that coordinates cell sorting within tissues. In addition, AJs are the initiators of cell-cell contact formation and cohesive interactions, promoting the formation and maintenance of desmosomes and TJs.Citation7 FAs and AJs maintain a dynamic biochemical and mechanosensory crosstalk toward the maintenance of epithelial architecture and function. FAs-mediated signals promote AJs-mediated cell cohesion by regulating the cytoskeleton and tension of epithelial cells.Citation8-Citation10 Reciprocally, AJs are also implicated in the regulation of the adhesive function of FAs in epithelia.Citation11,Citation12 In addition, their functions extend outside of the realm of epithelial cell adhesion and cytoskeletal organization, being implicated in an array of cellular processes including epithelial cell proliferation, migration, differentiation and apoptosis. The loss or failures in the regulation of FAs and AJs prevailing functional activity are causative of enhanced epithelial cell proliferation, loss of differentiation and the acquisition of tumorigenic and invasive characteristics.Citation13,Citation14
Taken together, the crosstalk between AJs and FAs regulates the precision of dynamic cell changes and rearrangements of epithelial tissues during homeostasis. Importantly, these adhesion receptors sense signals arising from the microenvironment and coordinate transient increases in epithelial cell proliferation, regulated cell migration and local epithelial remodeling and differentiation to repair severed tissues in response to injury.Citation12 When AJs and FAs are defective, these responses become unchecked, increasing the likelihood of epithelial diseases, including cancer.
However, our understanding of the contributions of FAs and AJs in sustaining epithelial homeostasis keeps broadening over the years. Several evidences indicate that the functions of some FAs and AJs proteins can no longer be strictly implicated only in the regulation of epithelial architecture and epithelial cell behavior. In this review, we highlight findings about the collaborative role of FAs and AJs associated proteins in integrating signals in epithelial cells and how they can participate in the communication with the surrounding tissue microenvironment. These signals regulate epithelial behavior and stromal cells responses, such as acute inflammation and angiogenesis, to fulfill a balanced epithelial proliferation and differentiation, migration and tissue repair. Alterations in FAs and AJs may result in a chronic “activation” of the stroma. This involves changes in the expression of ECM proteins, ECM degrading enzymes, growth factors and cytokines, that promote angiogenesis, inflammatory recruitment and the differentiation of fibroblasts into myofibroblasts, which in turn sustain growth-promoting signals involved in tumorigenesis.Citation2 Thus, when the signals mediated by some FAs and AJs proteins are defective, alterations in epithelial adhesion and homeostasis take place, and the “activation” of stromal cells cannot resolve, promoting uncontrolled epithelial proliferation, genetic mutations, evasion of senescence and apoptosis that lead to disease states, including carcinoma formation and metastatic spread. These findings are of clinical interest and are providing novel insights about the molecular mechanisms regulated by FAs and AJs that not only influence epithelial behavior but also the tissue microenvironment, and their implications in epithelial physiology and disease.
Molecular Composition of Focal Adhesions and Adherens Junctions
Focal adhesions
Epithelial sheets anchor to the underlying basement membrane that approach them to the stroma, via dynamic complexes formed by integrin receptors (). Epithelial integrin receptors form two different types of junctions, FAs linked to the actomyosin cytoskeleton, and hemidesmosomes that are linked to intermediate filaments.Citation6,Citation15,Citation16 FAs are present in most epithelial tissues, whereas hemidesmosomes are mainly found in skin.Citation15-Citation17 While the roles of FAs are better defined, the mechanical and signaling properties of hemidesmosomes are still poorly understood.
At the molecular level, integrin receptors are non-covalently linked heterodimers of α- and β-integrin subunits (). In FAs, the extracellular domain of these glycoproteins physically links cells to the ECM, and the cytoplasmic domain binds to scaffolding proteins that strengthen cell adhesion through their associations with the actin cytoskeleton, such as talin, kindlin, vinculin, α-actinin and paxillin.Citation18-Citation21 The heterogeneous combination of the different α- and β-subunits identified to date determines the integrin receptor type and the binding specificity of epithelial cells to diverse ECM ligands, including fibronectin, laminins and collagens.Citation22 This binding is not only involved in the mere physical attachment of cells to their substrates, but it also stimulates the signaling activity of the integrin receptors (affinity), which upon binding undergo conformational changes and integrin clustering (avidity).Citation12 Indeed, it is well established that cell morphology and function can be controlled by the stiffness of the extracellular matrix.Citation23,Citation24 Integrin receptors present both “outside-in” and “inside-out” signaling properties. “Outside-in” signaling occurs when the integrin receptor binds to its ligand and the signal is transmitted from the integrin receptor into the cell. “Inside-out” signaling refers to the shift of integrins between low- and high-affinity conformations that modulate their ligand binding.Citation21 The “inside-out” properties of integrins are regulated by external stimuli that are transduced intracellularly. These signals modulate the direct binding of regulatory proteins, including talin and kindlins, to the short cytoplasmic domain of integrins, and induce a shift in the activation state of integrins.Citation21 Several signaling transduction proteins are localized in FAs, including focal adhesion kinase (FAK), sarcoma kinases (Src), Abelson kinases (Abl), integrin-linked kinase (ILK) and small GTPases of both Ras and Rho families, including Rap1, Rac1, Cdc42 and Rho GTPases.Citation25,Citation26 Thus, integrin receptors act as sensors allowing cells to respond to different cues arising from the surrounding environment, instructing epithelial cells to change their behavior, including cell proliferation, cell survival and migration, in order to sustain tissue homeostasis.Citation27-Citation33
Adherens junctions
AJs localize at the boundary of epithelial cells, and they are highly dynamic structures constituted by two different types of junctions, Cadherin-catenins complexes anchored to cortical actin and microtubulesCitation34 and Nectin-afadin complexes anchored to the actin cytoskeleton ().Citation35 Classical cadherins are transmembrane glycoproteins that promote homophilic interactions between cells via their extracellular domain, constituted by five cadherin repeats (ECs), in a Ca2+-dependent manner.Citation36 E-cadherin is predominantly expressed in epithelial cells, and through its highly conserved cytoplasmic domain binds to the armadillo-repeat proteins β-cateninCitation37 and p120-catenin.Citation38 γ-catenin/plakoglobin, a β-catenin homologous protein, primarily associated with desmosomes, also associates to classical cadherins, and it can functionally substitute β-catenin when β-catenin is limited or modified.Citation39,Citation40 In turn, monomeric α-catenin binds to the complex via its interaction with β-catenin and indirectly anchors AJs complexes to the actin cytoskeleton,Citation41,Citation42 through its connections with actin-binding proteins such as Eplin, vinculin and α-actinin.Citation43-Citation45 While further work is needed to understand the complexity of the actin organization machinery that dynamically integrates AJs to the actin-myosin cytoskeleton, the recruitment of several actin polymerizing proteins at adhesion sites is regulated by α-catenin,Citation42,Citation46,Citation47 such as formin and vasodilator-stimulated protein (Vasp)/Enabled (Ena).Citation48,Citation49 In addition, other actin nucleating and remodeling proteins are recruited and activated at cell protrusion sites including Actin-related-proteins-2/3 (Arp2/3) and Cortactin proteins,Citation47,Citation50 as well as the family of small Rho GTPases (Rac1, Cdc42 and RhoA), which participates in the regulation of AJs and actin organization. This promotes AJs clustering, extension in the interphase of adhesion, and contact stabilization and strength, to achieve the formation of polarized epithelia, which involves dynamic interactions between different molecular players that have been documented in excellent reviews published elsewhere.Citation51
The other component of AJs is the nectin-afadin complex, constituted by the family of transmembrane proteins referred as to nectins, which are immunoglobulin-like cell adhesion molecules that promote homophilic and heterophilic interactions between epithelial cells in a Ca2+-independent way.Citation52 Nectins anchor to the actin cytoskeleton via ALL1-fused gene from chromosome 6 protein (AF6/afadin), an actin binding protein.Citation53-Citation55 These proteins are effectors of Rap1, a Ras-like small GTPase with an essential function in the formation of AJs.Citation56 AF6/Afadin also binds to α-catenin, providing a physical link between nectins and cadherins,Citation57 which promotes the collaboration of these complexes during the formation and stabilization of AJs. In addition, AF6/Afadin binds to various cell junction proteins, including ZO-1 in tight junctions,Citation58 thus, promoting the establishment of the intercellular junctional complex and polarized epithelia.Citation56,Citation59
Adhesive Crosstalk between Epithelial FAs and AJs
Over the past decades it has been recognized that adhesion receptors cooperate to sustain epithelial architecture acting as sensors of signals arising from their epithelial neighbors or from the surrounding tissue microenvironment. These can be biochemical or mechanical inputs that are translated into intracellular signals to regulate different aspects of cell behavior including proliferation, differentiation, cell survival, cell polarity and migration. These tightly coordinated events allow morphogenesis, tissue remodeling and sustain epithelial homeostasis. But how these signals are coordinated through the cooperation of integrin and cadherin adhesions is not completely understood. Several molecular and biochemical analyses have identified a number of common dynamic connections with the cytoskeleton and with signaling effectors.Citation60-Citation62 Thus, despite different molecular structures, integrin and cadherin-mediated adhesions operate with common biophysical characteristics to transmit and respond to biochemical signals and mechanical forces across epithelia.Citation63,Citation64
The interdependence of FAs and AJs has been observed in different systems. For example, mouse kidney epithelial cells deficient in α3β1 integrin fail to form the subcortical actin organization, and present a reduced AJs binding to α-actinin and anchoring to the actin cytoskeleton.Citation65 In skin keratinocytes, blocking antibodies to β1- and α3-integrins delay the formation of cadherin mediated adhesion and epithelial sheet formation, but keratinocytes terminal differentiation is not impaired,Citation66 consistent with the maintenance of cell-cell contacts in β1-integrin deficient mouse epidermis.Citation67,Citation68 These and other results overall suggest that integrins reinforce E-cadherin-dependent cell-cell adhesion. Conversely, reduction or loss of the functional activity of AJs proteins results in increased integrin expression and ECM adhesion. Some examples include the observations in cultured keratinocytes, in which blocking antibodies for P- and E-cadherin in differentiating keratinocytes, prevents the downregulation of integrin expression in differentiated cells.Citation69,Citation70 Furthermore, in mice with epidermal deficiency of both P- and E-cadherin, signs of actin disorganization are also evident, accompanied by an atypical expression of β4-integrin in the differentiated layers of the epidermis.Citation71 These and other results imply the existence of an inverse relationship between the assembly/disassembly of cadherin- and integrin-dependent adhesive structures. Overall, the results mentioned above highlight the relevance of a tight coordinated control of cadherin-integrin crosstalk to maintain epithelial homeostasis and prevent tumor cells to progress to an invasive state.
Adhesive crosstalk between epithelial FAs and AJs: mechanisms and players
Among the events that are initiated by FA- and AJ-mediated adhesion, the reorganization of cytoskeletal networks is one of the major outcomes that takes place. This involves changes in actin filaments, microtubules and intermediate filaments.Citation62 Although the molecular mechanisms that are involved in this response are not completely understood, a number of molecules have been implicated in this process. Despite the overwhelming diversity of players that have been documented to date, the ones that participate in the reorganization of the actin cytoskeleton are so far the best understood. Here we mention some of the actin cytoskeleton connections and signaling effectors that participate in FAs and AJs convergent signaling; these can be proteins that physically interact or modulate the actin cytoskeleton and link adhesion receptors to the peripheral cytoskeleton belt, including vinculin, α-actinin, Arp2/3 and cortactin, or a myriad of signaling proteins that are activated upon FA- and AJ-mediated adhesion including Src, FAK and small GTPases of both Ras and Rho families.Citation25,Citation26
One of the first perceptions that indicate that FAs and AJs intersect in an integrated network is that they connect to the actin cytoskeleton through several common molecules, including vinculin and α-actinin. However, vinculin and α-actinin interactions at FAs seem to be more stable than at AJs.Citation72 This may be explained by the fact that the dynamic recruitment of these common proteins to FAs and AJs seems to occur through different domains; indeed, vinculin binds to talin in FAs and is recruited through a different domain to cadherin-mediated adhesions in a β-catenin-dependent way, and activated by α-catenin during AJs formation.Citation73,Citation74
Several other common actin regulators are recruited to FAs and AJs, such as the important actin nucleator complex Arp2/3. In FAs, Arp2/3 is recruited by vinculin,Citation75 and at AJs it physically associates to E-cadherin upon direct E-cadherin ligation.Citation76 At these sites, Arp2/3 regulates actin assembly and its disruption compromises membrane protrusions and cell attachment.Citation77 These activities at FAs and AJs are coordinated by a key actin-binding scaffolding protein referred to as cortactin, which integrates all these dynamic actin networks in many cellular processes, including cell migration, endocytosis and cell adhesion.Citation50
Cortactin is also a key player that coordinates cell-signaling events and integrates cellular responses upon FAs and AJs formation. The activity of cortactin is modulated by phosphorylation of several kinases, but perhaps the best characterized is through the tyrosine kinase Src. This kinase also localizes to both integrin and cadherin adhesions, and it is transiently activated upon their engagement.Citation78,Citation79 Src is also implicated in coordinating signals between FAs and AJs, since overactivation of Src signaling can induce downregulation of E-cadherin by an integrin-dependent mechanism.Citation80-Citation83 However, much remains to be learnt about how Src phosphorylation regulates the crosstalk between FAs and AJs. Interestingly, it has been shown that cortactin binds to the AJs protein p120-catenin,Citation84 which like cortactin, was originally identified as a Src kinase substrate.Citation85 In addition, it has been observed that reductions of p120-catenin expression, which destabilize cadherin-catenin complexes and promote cadherin internalization along the endocytic pathway,Citation86 lead to a decrease in FA formation and impair the recruitment of cortactin to adhesion sites.Citation84 This may position cortactin along with p120-catenin in the cascade of events regulated by Src that coordinate the crosstalk between integrin-cadherin complexes.
The recruitment of Src to FAs can be promoted by FAK,Citation87 which physically interacts with the cytoplasmic tail of β1-integrin.Citation88,Citation89 Of note, downregulation or suppression of FAK signaling has been also implicated in the crosstalk of integrin-cadherin adhesion, since its loss perturbs the assembly of cadherins complexes in HeLa cells.Citation9 However, this has not been observed in epithelial tissues such as in FAK-deficient mouse epidermisCitation90,Citation91 and FAK-deficient mammary epithelia;Citation92 although its loss confers resistance to tumorigenesis in both tissues.Citation92,Citation93 This is likely due to the expression of its homolog proline rich tyrosine kinase 2 (PYK2) that may compensate for the loss of FAK.Citation91,Citation94
Src/FAK signaling has the ability to regulate membrane dynamics by modulating the activity of the small Rho-GTPases,Citation12,Citation25,Citation95 which are so far the best understood mode of integration of FAs and AJs mediated signals.Citation96,Citation97 Small GTPases are subjected to a GTP/GDP exchange cycle in which specific guanine nucleotide exchange factors (GEFs) promote the exchange of GDP to GTP, thereby converting small GTPases into an active signaling competent form.Citation98,Citation99 In many cell types, Rac negatively regulates the activity of Rho,Citation100 and the balance between these antagonistic activities is critical for the coordination of cell adhesion. Rac1 and Cdc42 are primarily involved in the formation of filopodia and membrane ruffles respectively, while RhoA generates myosin-based contractility and stabilizes membrane dynamics through the activation of several downstream effectors, including ROCK. These events are activated upon both integrin and cadherin ligation.Citation25,Citation87,Citation101-Citation104 For example, at initial stages of E-cadherin ligation both Rac and Cdc42 activities are transiently increased, while RhoA activity is initially decreased over few hours.Citation105-Citation109 Rac inhibition significantly reduces the ability of cadherin ligation to induce actin assembly.Citation109 When RhoA is inhibited, cells exhibit impaired spreading on fibronectin and decreased cell-cell adhesion. In the case of Cdc42, it has been shown that it limits Rho activation to prevent an excess of actin-mediated tension that otherwise will lead to loss of cell-cell junctions.Citation110
Several components of both FAs and AJs are implicated in mediating Rho GTPases signaling in an integrated crosstalk. For example exogenous expression of β1-integrin can activate RhoA and Rac1, leading to the disruption of cadherin-dependent cell-cell adhesion.Citation111 Additionally, the AJs protein p120-catenin has been directly implicated in the regulation of Rho GTPases. When overexpressed, it decreases Rho activity while increasing Rac and Cdc42 activity.Citation112-Citation116 p120-catenin has been also implicated in the crosstalk between Rac and Rho through the recruitment of p190RhoGAP, a Rho inhibitor.Citation117 This event may take place at AJs, coupling Rac activation to localized inhibition of Rho, which is essential for AJs formation. Interestingly, loss of p120-catenin leads to a constitutive activation of RhoA GTPase in fibroblasts and epithelial cells,Citation116,Citation117 which leads to an increase in FAs formation. This increase in FAs correlated with alterations in cell spreading, and cell motility in fibroblastsCitation117 and in keratinocytes (Epifano and Perez-Moreno, unpublished data).
An additional key player in coordinating FAs and AJs communication is ILK, which is a serine/threonine kinase recruited to FAs via direct interactions with β1- and β3-integrin cytoplasmic tails. ILK activation is related to the stabilization of β-catenin, which has dual roles in AJs and the canonical Wnt signaling pathway.Citation118,Citation119 Upon ILK activation, β-catenin stabilization and nuclear accumulation can take place, along with the formation of the transcriptional β-catenin/Tcf complexes and the regulation of Wnt signaling effectors.
Finally, Rap1, the closest homolog of the small GTPase Ras, is also a key player in the dynamic integrin-cadherin crosstalk.Citation59,Citation120 Rap1 regulates the “inside-out” activation of integrin, and E-cadherin plays a major role in the “outside-in” regulation of Rap1. In these lines, the disruption of E-cadherin mediated contacts and its internalization by endocytosis leads to the activation of Rap1, which is associated with an increased Src kinase activity. Interestingly, the E-cadherin internalized vesicles do not contain p120-catenin, and of note, p120-catenin in the cytoplasm has been reported to modulate the activity of Rho GTPases.Citation112 Upon cadherin internalization, the activated Rap1 mediates the formation of mature integrin-mediated adhesions.Citation121 This has been related to the capability of Rap1 to regulate both recycling, avidity and affinity of integrins.Citation59,Citation122-Citation124 These findings also suggest a major role of Rap1 in transmitting information from cadherin-based to integrin-based adhesive structures to couple cadherin inhibition to integrin activation during the remodeling of epithelial tissues.
Overall, these are some compelling evidences observed in epithelial cell lines, normal epithelial tissues and primary cells that indicate the existence of collaborative molecular networks between cell-cell and cell-matrix adhesions that sustain epithelial cell adhesion. The tight spatiotemporal regulation of the dynamic coordination of these connections is therefore fundamental for tissue architecture. FAs and AJs collaborative pathways also share the ability to influence different aspects of cell behavior, which are important for morphogenesis, the physiological balance of adult epithelial tissues, and with inherent implications in cancer.
FAs and AJs as Integrated Modules that Sense and Respond to Signals Arising from Epithelia and Stroma
The integrated mechanistic crosstalk of integrins and cadherins allow epithelial cells to respond to biochemical or mechanical inputs that are translated into intracellular signals to regulate the physiological balance of epithelial cells.Citation23,Citation24,Citation62,Citation125,Citation126 The direct involvement of several FAs or AJs proteins in regulating several aspects of epithelial behavior, beyond the mere mechanical interconnection of cells, has been extensively documented in seminal publications that have been published over the past decades. Several FAs and AJs are bona fide tumor suppressors and oncogenes, and overall their loss of expression or normal functional activity correlate to different epithelial alterations depending on the epithelial system, ranging from loss of cell polarity, reduced or increased cell migration, increased or decreased epithelial proliferation, cell survival or apoptosis or circumvention of differentiation and senescence. These result in defects that can perturb epithelial morphogenesis, loss of tissue function and have very important implications in cancer. In this regard, one of the most profound alterations that take place by defective FAs or AJs adhesive properties, expression or signaling is the occurrence of epithelial-to-mesenchymal transition (EMT), which is a complex molecular program involved in morphogenesis and cancer that results in a gain of migratory and invasive characteristics. The readers are referred to some of several excellent reviews published elsewhere covering different aspects of cell behavior regulated by FAs or AJs during epithelial homeostasis and cancer.Citation127-Citation133 In this section, we will highlight some examples of how the integrated crosstalk of FAs and AJs in epithelial cells can sense and respond to signals that arise from the underlying stroma, and boost both epithelial and stromal responses to sustain homeostasis, and when defective may lead to cancer ().
Figure 2. A reciprocal and coordinated regulation of FAs and AJs in epithelial cells is involved in the crosstalk between epithelia and stroma. FAs and AJs share downstream signaling molecules, including Rho GTPases and Src, and interactions with actomyosin cytoskeleton, which contribute to the coordination of their adhesive network. Whereas FAs assembly is related to a strengthening of AJ-mediated adhesion, AJs disassembly is related to an increase of FAs-mediated adhesion. During tumorigenesis the coordinated crosstalk between FAs and AJs in epithelial cells is impaired, and may result in the chronic activation of the stroma (e.g., secretion of soluble factors, cytokines, chemokines, MMPs, and changes in ECM composition). These can generate perpetuating signaling loops without a clear endpoint that, if unresolved, may lead to further epithelial transformation with gain of migratory and invasive characteristics. In this figure, some specific signaling adhesive modules are shown using color-coded annotations as well as their points of convergence.
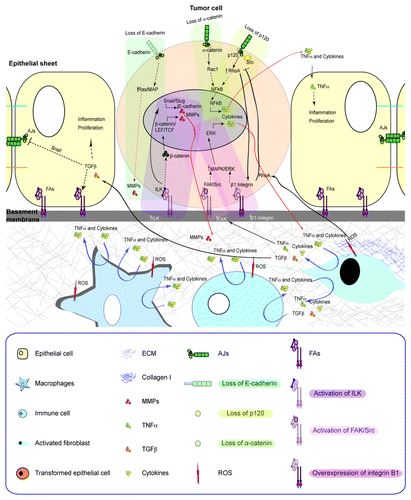
Epithelial cells are able to present dynamic spatiotemporal changes in phenotype and function referred as to “plasticity.” They also display “reciprocity” characteristics, which is the ability to process signals from the environment.Citation14,Citation134-Citation137 These events are fundamental during tissue remodeling processes such as morphogenesis and wound healing, which are tightly controlled and with well-defined phases. During these events, transient increases of cell proliferation, collective cell migration and epithelial remodeling are observed to reestablish a functional epithelium,Citation138 and are unrestrained in cancer cells.Citation136 This has led to the establishment of parallels between tumors and a defective perpetuating process of wound repair.Citation139
Upon injury, cellular junctions are severed, and AJs undergo a dynamic assembly and disassembly state that needs to be coordinated to integrin activation and adhesion to the ECM. This process requires the remodeling of the actin cytoskeleton and microtubule networks to allow epithelial spreading and polarized collective cell migration on a temporary secreted ECM toward the maintenance of tissue architecture. Interestingly, in Xenopus laevis it has been recently observed that keratin intermediate filaments can associate to plakoglobin at AJs and regulate collective cell migration.Citation140 The contributions of this binding to the regulation of the affinity of integrins, perhaps through scaffolding proteins including plectins, is an important line of future investigation.
If the integrated crosstalk between FAs and AJs is perturbed, it can lead to important changes in cell behavior (). Indeed, the reduced strength of both cadherins, and switch in the expression of integrins and FAs attachment to ECMs,Citation141-Citation143 can result in increased individual cell migration, which can be mesenchymal or even switch to an ameboid state that is FAs independent.Citation144,Citation145 Epithelial repair upon injury also includes an acute inflammatory response and recruitment of immune cells and changes in the underlying connective tissue, including the formation of blood vessels, activation of fibroblasts and changes in the ECM and secreted growth factors that actively participate in promoting wound healing. These responses also need to be tightly controlled, since their chronic activation contributes to cancer ().Citation2 Different signals arise form the epithelial cells and the tissue microenvironment including members of growth factors (TGF-β, TNF-α, KGF and HGF), chemokines, interleukins, prostaglandins, matrix metalloproteinases, changes in ECM composition and the generation of reactive oxygen species,Citation136,Citation146-Citation161 that can affect the activation state of cadherin and integrin adhesion receptors ().
FAs and AJs in epithelia and stroma communication: adhesion and signaling
The coordination of the FAs and AJs crosstalk modulated by Rho GTPases has been observed in several tissue remodeling events. For example, AJs breakage upon injury may induce the accumulation of p120-catenin in the cytoplasm, which in turn activates Rac and Cdc42, and cell migration.Citation162 This is consistent with the observed roles of p120-catenin in cell migration, which could take place in a cadherin independent manner.Citation163 Its causal involvement in this response has been observed in some transformed cell systems, including ovarian cancer cells, in which the loss of p120-catenin can block their migration.Citation164 Interestingly, conditional loss of function studies in mice have identified another role for p120-catenin, which consist in regulating signals from epithelial cells that emanate to the stroma and prevent chronic inflammation in epithelial tissues, including skin,Citation116 intestine,Citation165 esophagus and stomach.Citation166 This involves the Rho-dependent activation of the inflammatory mediator NFκB, chronic inflammation and cancer ().Citation116,Citation166,Citation167 Conversely, upstream of p120-catenin, both epithelial growth factor (EGF) and hepatocyte growth factor (HGF) have been shown to induce cell scattering and relocalization of p120-catenin from the membrane to the cytoplasm activating Rho GTPase activity and changes in FAs. Interestingly, increases in RhoA have also been related to the activation of β-catenin mediated signals in keratinocytes, which results in tissue tension that induces epidermal hyperplasia and tumor growth.Citation168
Loss of α-catenin in the epidermis has also been implicated in NFκB activation, inflammation and cancer, but mediated in part by Rac1 ().Citation116,Citation169 In addition, α-catenin null epidermal cells show enhanced migratory behavior, increased sensitivity to insulin growth factor stimulation (IGF) and elevated Ras and MAPK activity.Citation170
β-catenin, through its function in Wnt signaling, has a key function in integrating signals arising from FAs to respond to changes in the surrounding epithelia environment. In this line, as we mentioned in a previous section, β-catenin can be stabilized and translocated to the nucleus upon activation of ILK.Citation171 Interestingly, moderate increases in the normal ILK activity are sufficient to trigger Wnt signaling, downregulate E-cadherin expression in intestinal and mammary epithelial cells, and induce tumorigenic characteristics,Citation172,Citation173 that are triggered by the expression of the EMT transcription factors Snail and Slug ().Citation174 Furthermore, transgenic mice expressing ILK in mammary epithelia exhibits an accelerated tumor progression and an expansion of breast luminal progenitor cells.Citation175 Conversely, its loss leads to reductions in the appearance of inflammation-related cancers in a tumor carcinogenesis protocol, which is related to decreased expression of MCP1 and the chemokine CCL2, limited ECM changes along with a scarce inflammatory response.Citation176
β-catenin can also functionally associate to FAs through its interactions with the laminin receptor α3β1-integrin. Primary α3β1-integrin null cells fail to activate β-catenin upon TGFβ1 mediated signals, since in the absence of α3β1-integrin active-pSmad is unable to associate to β-catenin. Thus, this reduces the capability of the cells to respond to TGFβ1 and undergo EMT events.Citation177
Non-canonical Wnt signals are also at the nexus of FAs and AJs mediated signals, and promote a coordination of signals along with the surrounding stroma. Wnt5, a prototype Wnt for β-catenin-independent signaling has been implicated in proliferation and inflammation. In Xenopus laevis, Wnt5 is able to stimulate fibronectin and AJs assembly in a Rac dependent way,Citation178 thus regulating morphogenetic movements, whereas in mammalian cells, Wnt5 cooperates with integrin signaling to stimulate FAs turnover in migrating cells.Citation179
FAs and AJs in epithelia and stroma communication: growth factors and ROS
Cadherin and integrin complexes interact, sense and respond to the activation of several growth factors, however, their coordinated modulation through TGFβ is so far the best understood. TGFβ not only activates integrins (), but also regulates the expression of numerous integrins and integrin-associated proteins.Citation180,Citation181 Both FAs and AJs also integrate the signals mediated by this factor. Some examples include the observations in keratinocyte cell lines, where it induces loss of cadherin mediated adhesion, elevated Ras and MAPK activity and matrix metalloproteinase 9 (MMP9) expression.Citation182 Interestingly these effects are mediated by the activation of Rac1 and are accompanied by increase in cell migration and invasion, and the expression of the E-cadherin repressor Snail1 leading to EMT.Citation183 Conversely, it has been shown that loss of TGFβ receptor II in epidermal cells leads to the development of squamous carcinomas of rectal and genital epithelia, which are normally proliferative and subjected to stress, but not in the normal epidermis. However, its loss promoted accelerated tumor formation in epidermis in a mutated Ras background. Interestingly, TGFβRII-null keratinocytes also display increased integrin-FAK-Src signaling and migration. These data are consistent with the dual roles of TGFβ attributed during tumorigenesis: it acts as a tumor promoter during cancer progression, and as a tumor suppressor at early stages of tumorigenesis.Citation184 Of note, it is known that TGFβ also activates other signaling pathways including p38 MAPK, JNK and PI3K-AktCitation185,Citation186 that have been involved in the secretion of cytokines and chemokines influencing the inflammatory tumor microenvironment,Citation187 inducing angiogeniesisCitation188 and overall boosting the invasive properties of tumor cells.Citation189
In addition, HGF has been involved in promoting cell scattering and key aspects of EMT.Citation129,Citation190 It has been observed that HGF induced-cell scattering is enhanced by collagen and fibronectin and increases integrin mediated adhesion. Interestingly, it has been suggested that the loss of cadherin-mediated adhesion observed upon HGF treatment, is not due to decreases in the functional E-cadherin adhesive properties, but rather to increased integrin mediated adhesion and actomyosin traction forces that may disrupt cell-cell adhesion.Citation191
Another growth factor that can activate signals that are coordinated by both FAs and AJs is tumor necrosis factor α (TNFα, ), which is one of the major mediators of inflammation and inflammation-associated cancers.Citation160,Citation192,Citation193 TNFα has the ability to induce the phosphorylation of FAK, and regulate FAK-mediated signaling and coordinately enhance tumor invasion.Citation194 TNFα is expressed in both epithelia and stroma, along with other inflammatory mediators,Citation195,Citation196 thus increasing the responses toward tissue repair or cancer. Its activation can be mediated by the NFκB-Snail signaling pathway,Citation160,Citation192,Citation197 JNKCitation198 and/or Wnt/β-catenin.Citation199,Citation200
Finally, emerging evidence indicates that reactive oxygen species (ROS) may regulate the coordinated crosstalk of FAs and AJs.Citation161 These are a number of reactive molecules and free radicals derived from molecular oxygen, such as hydrogen peroxide;Citation201 which are produced during the mitochondrial aerobic respiration or by oxidoreductase enzymes and catalyzed oxidation of metals, and they are strong oxidants with potential deleterious effects. Their levels are highly increased in events such as fibrosis and inflammation, and in turn, they act as cellular messengers that participate in cell signaling and in the regulation of gene expression.Citation201 In these lines, it has been proposed that ROS influence the “inside-out” signaling of integrins by inducing their conformational change and activation ().Citation202 Of note, signaling proteins such as Rho GTPases and phosphatases are also target proteins of ROS. In addition, ROS can also modulate the stability of cadherin-mediated adhesions.Citation161
Concluding Remarks
The crosstalk between FAs and AJs is involved in the regulation of dynamic cell changes and rearrangements of epithelial cells within tissues, and several evidences indicate that they embark in a “tug of war” to sustain epithelial architecture and homeostasis.Citation12,Citation23,Citation62 However, their functions extend beyond the realm of cell adhesion to act as important sensors of the tissue environment. This is of particular relevance, since epithelial tissues are non-vascularized and receive their nutrients and oxygen from the stroma. In this regard, FAs and AJs may act as integrated adhesive units to sense and respond to signals arising from the tissue microenvironment. Future research will provide more insights about how their integrated functions promote epithelia and stroma communication; however, evidence has emerged in the past years implicating several FAs and AJs proteins in the regulation of signaling pathways involved in this event. Through them, they can promote a dynamic crosstalk between epithelial cells and stroma to coordinate transient increases in cell proliferation, regulated cell migration, inflammation, epithelial remodeling and differentiation to sustain the physiological balance of epithelial tissues. We propose that when the coordinated crosstalk between AJs and FAs is defective, all these above responses become unchecked, increasing the likelihood of epithelial diseases, including cancer (). Interestingly, recent approaches to study the role of some of these proteins in the stroma, such as β-catenin or FAK, have also underscore their relevance in sustaining stromal function.Citation203,Citation204 This also has to be tightly modulated since chronic activation of the stroma including fibroblasts, endothelial cells, inflammatory cells and changes in the ECM responses and generation of ROS are related to cancer and metastasis dissemination. Thus, FAs and AJs may regulate the overall tissue context, and it will be interesting to learn how they coordinate different aspects in epithelial physiology and disease.
Acknowledgments
We thank Marta Shahbazi, Ljiljana Dukanovic, Juan Guinea-Viniegra and Helia Schönthaler, members of the BBVAF-Cancer Cell Biology Program, for insightful discussions and for helpful comments on the manuscript. C.E. was a recipient of the Spanish Juan de la Cierva fellowship. M.P.-M.'s group is supported by grants from the Spanish Ministry of Science and Innovation (BFU2009-11885), the Association for International Cancer Research AICR-UK (10-0746) and from the CNIO (BC1102).
References
- Rozario T, DeSimone DW. The extracellular matrix in development and morphogenesis: a dynamic view. Dev Biol 2010; 341:126 - 40; http://dx.doi.org/10.1016/j.ydbio.2009.10.026; PMID: 19854168
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell 2011; 144:646 - 74; http://dx.doi.org/10.1016/j.cell.2011.02.013; PMID: 21376230
- Perez-Moreno M, Jamora C, Fuchs E. Sticky business: orchestrating cellular signals at adherens junctions. Cell 2003; 112:535 - 48; http://dx.doi.org/10.1016/S0092-8674(03)00108-9; PMID: 12600316
- Green KJ, Gaudry CA. Are desmosomes more than tethers for intermediate filaments?. Nat Rev Mol Cell Biol 2000; 1:208 - 16; http://dx.doi.org/10.1038/35043032; PMID: 11252896
- Danen EHJ, Sonnenberg A. Integrins in regulation of tissue development and function. J Pathol 2003; 201:632 - 41; http://dx.doi.org/10.1002/path.1472; PMID: 14648669
- Tsuruta D, Hashimoto T, Hamill KJ, Jones JCR. Hemidesmosomes and focal contact proteins: functions and cross-talk in keratinocytes, bullous diseases and wound healing. J Dermatol Sci 2011; 62:1 - 7; PMID: 21376539
- Gumbiner BM. Cell adhesion: the molecular basis of tissue architecture and morphogenesis. Cell 1996; 84:345 - 57; http://dx.doi.org/10.1016/S0092-8674(00)81279-9; PMID: 8608588
- Marsden M, DeSimone DW. Integrin-ECM interactions regulate cadherin-dependent cell adhesion and are required for convergent extension in Xenopus. Curr Biol 2003; 13:1182 - 91; http://dx.doi.org/10.1016/S0960-9822(03)00433-0; PMID: 12867028
- Yano H, Mazaki Y, Kurokawa K, Hanks SK, Matsuda M, Sabe H. Roles played by a subset of integrin signaling molecules in cadherin-based cell-cell adhesion. J Cell Biol 2004; 166:283 - 95; http://dx.doi.org/10.1083/jcb.200312013; PMID: 15263022
- Martinez-Rico C, Pincet F, Thiery J-P, Dufour S. Integrins stimulate E-cadherin-mediated intercellular adhesion by regulating Src-kinase activation and actomyosin contractility. J Cell Sci 2010; 123:712 - 22; http://dx.doi.org/10.1242/jcs.047878; PMID: 20144995
- Balzac F, Avolio M, Degani S, Kaverina I, Torti M, Silengo L, et al. E-cadherin endocytosis regulates the activity of Rap1: a traffic light GTPase at the crossroads between cadherin and integrin function. J Cell Sci 2005; 118:4765 - 83; http://dx.doi.org/10.1242/jcs.02584; PMID: 16219685
- Papusheva E, Heisenberg C-P. Spatial organization of adhesion: force-dependent regulation and function in tissue morphogenesis. EMBO J 2010; 29:2753 - 68; http://dx.doi.org/10.1038/emboj.2010.182; PMID: 20717145
- Cavallaro U, Christofori G. Multitasking in tumor progression: signaling functions of cell adhesion molecules. Ann N Y Acad Sci 2004; 1014:58 - 66; http://dx.doi.org/10.1196/annals.1294.006; PMID: 15153420
- Desgrosellier JS, Cheresh DA. Integrins in cancer: biological implications and therapeutic opportunities. Nat Rev Cancer 2010; 10:9 - 22; http://dx.doi.org/10.1038/nrc2748; PMID: 20029421
- Fuchs E, Raghavan S. Getting under the skin of epidermal morphogenesis. Nat Rev Genet 2002; 3:199 - 209; http://dx.doi.org/10.1038/nrg758; PMID: 11972157
- Simpson CL, Patel DM, Green KJ. Deconstructing the skin: cytoarchitectural determinants of epidermal morphogenesis. Nat Rev Mol Cell Biol 2011; 12:565 - 80; http://dx.doi.org/10.1038/nrm3175; PMID: 21860392
- Borradori L, Sonnenberg A. Structure and function of hemidesmosomes: more than simple adhesion complexes. J Invest Dermatol 1999; 112:411 - 8; http://dx.doi.org/10.1046/j.1523-1747.1999.00546.x; PMID: 10201522
- Čabrijan L, Lipozenčić J. Adhesion molecules in keratinocytes. Clin Dermatol 2011; 29:427 - 31; http://dx.doi.org/10.1016/j.clindermatol.2011.01.012; PMID: 21679870
- Plow EF, Haas TA, Zhang L, Loftus J, Smith JW. Ligand binding to integrins. J Biol Chem 2000; 275:21785 - 8; http://dx.doi.org/10.1074/jbc.R000003200; PMID: 10801897
- Calderwood DA, Shattil SJ, Ginsberg MH. Integrins and actin filaments: reciprocal regulation of cell adhesion and signaling. J Biol Chem 2000; 275:22607 - 10; http://dx.doi.org/10.1074/jbc.R900037199; PMID: 10801899
- Moser M, Legate KR, Zent R, Fässler R. The tail of integrins, talin, and kindlins. Science 2009; 324:895 - 9; http://dx.doi.org/10.1126/science.1163865; PMID: 19443776
- Gilcrease MZ. Integrin signaling in epithelial cells. Cancer Lett 2007; 247:1 - 25; http://dx.doi.org/10.1016/j.canlet.2006.03.031; PMID: 16725254
- Schwartz MA, DeSimone DW. Cell adhesion receptors in mechanotransduction. Curr Opin Cell Biol 2008; 20:551 - 6; http://dx.doi.org/10.1016/j.ceb.2008.05.005; PMID: 18583124
- Mammoto T, Ingber DE. Mechanical control of tissue and organ development. Development 2010; 137:1407 - 20; http://dx.doi.org/10.1242/dev.024166; PMID: 20388652
- Schwartz MA, Shattil SJ. Signaling networks linking integrins and rho family GTPases. Trends Biochem Sci 2000; 25:388 - 91; http://dx.doi.org/10.1016/S0968-0004(00)01605-4; PMID: 10916159
- Kim C, Ye F, Ginsberg MH. Regulation of integrin activation. Annu Rev Cell Dev Biol 2011; 27:321 - 45; http://dx.doi.org/10.1146/annurev-cellbio-100109-104104; PMID: 21663444
- Hynes RO. Integrins: bidirectional, allosteric signaling machines. Cell 2002; 110:673 - 87; http://dx.doi.org/10.1016/S0092-8674(02)00971-6; PMID: 12297042
- Schwartz MA, Ginsberg MH. Networks and crosstalk: integrin signalling spreads. Nat Cell Biol 2002; 4:E65 - 8; http://dx.doi.org/10.1038/ncb0402-e65; PMID: 11944032
- Giancotti FG, Ruoslahti E. Integrin signaling. Science 1999; 285:1028 - 32; http://dx.doi.org/10.1126/science.285.5430.1028; PMID: 10446041
- Liddington RC, Ginsberg MH. Integrin activation takes shape. J Cell Biol 2002; 158:833 - 9; http://dx.doi.org/10.1083/jcb.200206011; PMID: 12213832
- Leahy DJ. Implications of atomic-resolution structures for cell adhesion. Annu Rev Cell Dev Biol 1997; 13:363 - 93; http://dx.doi.org/10.1146/annurev.cellbio.13.1.363; PMID: 9442878
- Cohen M, Joester D, Geiger B, Addadi L. Spatial and temporal sequence of events in cell adhesion: from molecular recognition to focal adhesion assembly. Chembiochem 2004; 5:1393 - 9; http://dx.doi.org/10.1002/cbic.200400162; PMID: 15457530
- Geiger B, Spatz JP, Bershadsky AD. Environmental sensing through focal adhesions. Nat Rev Mol Cell Biol 2009; 10:21 - 33; http://dx.doi.org/10.1038/nrm2593; PMID: 19197329
- Meng W, Takeichi M. Adherens junction: molecular architecture and regulation. Cold Spring Harb Perspect Biol 2009; 1:a002899; http://dx.doi.org/10.1101/cshperspect.a002899; PMID: 20457565
- Nakanishi H, Takai Y. Roles of nectins in cell adhesion, migration and polarization. Biol Chem 2004; 385:885 - 92; http://dx.doi.org/10.1515/BC.2004.116; PMID: 15551862
- Gumbiner BM. Regulation of cadherin-mediated adhesion in morphogenesis. Nat Rev Mol Cell Biol 2005; 6:622 - 34; http://dx.doi.org/10.1038/nrm1699; PMID: 16025097
- Knudsen KA, Wheelock MJ. Plakoglobin, or an 83-kD homologue distinct from beta-catenin, interacts with E-cadherin and N-cadherin. J Cell Biol 1992; 118:671 - 9; http://dx.doi.org/10.1083/jcb.118.3.671; PMID: 1639850
- Reynolds AB, Herbert L, Cleveland JL, Berg ST, Gaut JR. p120, a novel substrate of protein tyrosine kinase receptors and of p60v-src, is related to cadherin-binding factors beta-catenin, plakoglobin and armadillo. Oncogene 1992; 7:2439 - 45; PMID: 1334250
- Lewis JE, Wahl JK 3rd, Sass KM, Jensen PJ, Johnson KR, Wheelock MJ. Cross-talk between adherens junctions and desmosomes depends on plakoglobin. J Cell Biol 1997; 136:919 - 34; http://dx.doi.org/10.1083/jcb.136.4.919; PMID: 9049256
- Miravet S, Piedra J, Castaño J, Raurell I, Francí C, Duñach M, et al. Tyrosine phosphorylation of plakoglobin causes contrary effects on its association with desmosomes and adherens junction components and modulates beta-catenin-mediated transcription. Mol Cell Biol 2003; 23:7391 - 402; http://dx.doi.org/10.1128/MCB.23.20.7391-7402.2003; PMID: 14517306
- Yamada S, Pokutta S, Drees F, Weis WI, Nelson WJ. Deconstructing the cadherin-catenin-actin complex. Cell 2005; 123:889 - 901; http://dx.doi.org/10.1016/j.cell.2005.09.020; PMID: 16325582
- Drees F, Pokutta S, Yamada S, Nelson WJ, Weis WI. Alpha-catenin is a molecular switch that binds E-cadherin-beta-catenin and regulates actin-filament assembly. Cell 2005; 123:903 - 15; http://dx.doi.org/10.1016/j.cell.2005.09.021; PMID: 16325583
- Abe K, Takeichi M. EPLIN mediates linkage of the cadherin catenin complex to F-actin and stabilizes the circumferential actin belt. Proc Natl Acad Sci U S A 2008; 105:13 - 9; http://dx.doi.org/10.1073/pnas.0710504105; PMID: 18093941
- Watabe-Uchida M, Uchida N, Imamura Y, Nagafuchi A, Fujimoto K, Uemura T, et al. alpha-Catenin-vinculin interaction functions to organize the apical junctional complex in epithelial cells. J Cell Biol 1998; 142:847 - 57; http://dx.doi.org/10.1083/jcb.142.3.847; PMID: 9700171
- Knudsen KA, Soler AP, Johnson KR, Wheelock MJ. Interaction of alpha-actinin with the cadherin/catenin cell-cell adhesion complex via alpha-catenin. J Cell Biol 1995; 130:67 - 77; http://dx.doi.org/10.1083/jcb.130.1.67; PMID: 7790378
- Scott JA, Yap AS. Cinderella no longer: alpha-catenin steps out of cadherin’s shadow. J Cell Sci 2006; 119:4599 - 605; http://dx.doi.org/10.1242/jcs.03267; PMID: 17093264
- Lecuit T. alpha-catenin mechanosensing for adherens junctions. Nat Cell Biol 2010; 12:522 - 4; http://dx.doi.org/10.1038/ncb2066; PMID: 20453846
- Vasioukhin V, Fuchs E. Actin dynamics and cell-cell adhesion in epithelia. Curr Opin Cell Biol 2001; 13:76 - 84; http://dx.doi.org/10.1016/S0955-0674(00)00177-0; PMID: 11163137
- Kobielak A, Pasolli HA, Fuchs E. Mammalian formin-1 participates in adherens junctions and polymerization of linear actin cables. Nat Cell Biol 2004; 6:21 - 30; http://dx.doi.org/10.1038/ncb1075; PMID: 14647292
- Ren G, Crampton MS, Yap AS. Cortactin: Coordinating adhesion and the actin cytoskeleton at cellular protrusions. Cell Motil Cytoskeleton 2009; 66:865 - 73; http://dx.doi.org/10.1002/cm.20380; PMID: 19437513
- Maiden SL, Hardin J. The secret life of α-catenin: moonlighting in morphogenesis. J Cell Biol 2011; 195:543 - 52; http://dx.doi.org/10.1083/jcb.201103106; PMID: 22084304
- Brakeman PR, Liu KD, Shimizu K, Takai Y, Mostov KE. Nectin proteins are expressed at early stages of nephrogenesis and play a role in renal epithelial cell morphogenesis. Am J Physiol Renal Physiol 2009; 296:F564 - 74; http://dx.doi.org/10.1152/ajprenal.90328.2008; PMID: 19116242
- Takai Y, Miyoshi J, Ikeda W, Ogita H. Nectins and nectin-like molecules: roles in contact inhibition of cell movement and proliferation. Nat Rev Mol Cell Biol 2008; 9:603 - 15; http://dx.doi.org/10.1038/nrm2457; PMID: 18648374
- Harris TJC, Tepass U. Adherens junctions: from molecules to morphogenesis. Nat Rev Mol Cell Biol 2010; 11:502 - 14; http://dx.doi.org/10.1038/nrm2927; PMID: 20571587
- Niessen CM. Tight junctions/adherens junctions: basic structure and function. J Invest Dermatol 2007; 127:2525 - 32; http://dx.doi.org/10.1038/sj.jid.5700865; PMID: 17934504
- Kooistra MRH, Dubé N, Bos JL. Rap1: a key regulator in cell-cell junction formation. J Cell Sci 2007; 120:17 - 22; http://dx.doi.org/10.1242/jcs.03306; PMID: 17182900
- Hoshino T, Sakisaka T, Baba T, Yamada T, Kimura T, Takai Y. Regulation of E-cadherin endocytosis by nectin through afadin, Rap1, and p120ctn. J Biol Chem 2005; 280:24095 - 103; http://dx.doi.org/10.1074/jbc.M414447200; PMID: 15857834
- Yamada A, Irie K, Fukuhara A, Ooshio T, Takai Y. Requirement of the actin cytoskeleton for the association of nectins with other cell adhesion molecules at adherens and tight junctions in MDCK cells. Genes Cells 2004; 9:843 - 55; http://dx.doi.org/10.1111/j.1365-2443.2004.00768.x; PMID: 15330861
- Bos JL. Linking Rap to cell adhesion. Curr Opin Cell Biol 2005; 17:123 - 8; http://dx.doi.org/10.1016/j.ceb.2005.02.009; PMID: 15780587
- Brunton VG, MacPherson IRJ, Frame MC. Cell adhesion receptors, tyrosine kinases and actin modulators: a complex three-way circuitry. Biochim Biophys Acta 2004; 1692:121 - 44; http://dx.doi.org/10.1016/j.bbamcr.2004.04.010; PMID: 15246683
- Mège R-M, Gavard J, Lambert M. Regulation of cell-cell junctions by the cytoskeleton. Curr Opin Cell Biol 2006; 18:541 - 8; http://dx.doi.org/10.1016/j.ceb.2006.08.004; PMID: 16905303
- Weber GF, Bjerke MA, DeSimone DW. Integrins and cadherins join forces to form adhesive networks. J Cell Sci 2011; 124:1183 - 93; http://dx.doi.org/10.1242/jcs.064618; PMID: 21444749
- Chen X, Gumbiner BM. Crosstalk between different adhesion molecules. Curr Opin Cell Biol 2006; 18:572 - 8; http://dx.doi.org/10.1016/j.ceb.2006.07.002; PMID: 16859906
- Maruthamuthu V, Aratyn-Schaus Y, Gardel ML. Conserved F-actin dynamics and force transmission at cell adhesions. Curr Opin Cell Biol 2010; 22:583 - 8; http://dx.doi.org/10.1016/j.ceb.2010.07.010; PMID: 20728328
- Wang Z, Symons JM, Goldstein SL, McDonald A, Miner JH, Kreidberg JA. (Alpha)3(beta)1 integrin regulates epithelial cytoskeletal organization. J Cell Sci 1999; 112:2925 - 35; PMID: 10444387
- Jensen PJ, Wheelock MJ. Beta 1 integrins do not have a major role in keratinocyte intercellular adhesion. Exp Cell Res 1995; 219:322 - 31; http://dx.doi.org/10.1006/excr.1995.1235; PMID: 7543849
- Grose R, Hutter C, Bloch W, Thorey I, Watt FM, Fässler R, et al. A crucial role of beta 1 integrins for keratinocyte migration in vitro and during cutaneous wound repair. Development 2002; 129:2303 - 15; PMID: 11959837
- Raghavan S, Vaezi A, Fuchs E. A role for alphabeta1 integrins in focal adhesion function and polarized cytoskeletal dynamics. Dev Cell 2003; 5:415 - 27; http://dx.doi.org/10.1016/S1534-5807(03)00261-2; PMID: 12967561
- Hodivala KJ, Watt FM. Evidence that cadherins play a role in the downregulation of integrin expression that occurs during keratinocyte terminal differentiation. J Cell Biol 1994; 124:589 - 600; http://dx.doi.org/10.1083/jcb.124.4.589; PMID: 8106556
- Zhu AJ, Watt FM. Expression of a dominant negative cadherin mutant inhibits proliferation and stimulates terminal differentiation of human epidermal keratinocytes. J Cell Sci 1996; 109:3013 - 23; PMID: 9004036
- Tinkle CL, Pasolli HA, Stokes N, Fuchs E. New insights into cadherin function in epidermal sheet formation and maintenance of tissue integrity. Proc Natl Acad Sci U S A 2008; 105:15405 - 10; http://dx.doi.org/10.1073/pnas.0807374105; PMID: 18809908
- Palovuori R, Eskelinen S. Role of vinculin in the maintenance of cell-cell contacts in kidney epithelial MDBK cells. Eur J Cell Biol 2000; 79:961 - 74; http://dx.doi.org/10.1078/0171-9335-00120; PMID: 11152287
- Peng X, Cuff LE, Lawton CD, DeMali KA. Vinculin regulates cell-surface E-cadherin expression by binding to beta-catenin. J Cell Sci 2010; 123:567 - 77; http://dx.doi.org/10.1242/jcs.056432; PMID: 20086044
- Peng X, Maiers JL, Choudhury D, Craig SW, Demali KA. Alpha catenin uses a novel mechanism to activate vinculin. J Biol Chem 2012; 287:7728 - 37; http://dx.doi.org/10.1074/jbc.M111.297481; PMID: 22235119
- DeMali KA, Barlow CA, Burridge K. Recruitment of the Arp2/3 complex to vinculin: coupling membrane protrusion to matrix adhesion. J Cell Biol 2002; 159:881 - 91; http://dx.doi.org/10.1083/jcb.200206043; PMID: 12473693
- Kovacs EM, Yap AS. The web and the rock: cell adhesion and the ARP2/3 complex. Dev Cell 2002; 3:760 - 1; http://dx.doi.org/10.1016/S1534-5807(02)00374-X; PMID: 12479800
- Verma S, Shewan AM, Scott JA, Helwani FM, den Elzen NR, Miki H, et al. Arp2/3 activity is necessary for efficient formation of E-cadherin adhesive contacts. J Biol Chem 2004; 279:34062 - 70; http://dx.doi.org/10.1074/jbc.M404814200; PMID: 15159390
- Frame MC, Fincham VJ, Carragher NO, Wyke JA. v-Src’s hold over actin and cell adhesions. Nat Rev Mol Cell Biol 2002; 3:233 - 45; http://dx.doi.org/10.1038/nrm779; PMID: 11994743
- McLachlan RW, Kraemer A, Helwani FM, Kovacs EM, Yap AS. E-cadherin adhesion activates c-Src signaling at cell-cell contacts. Mol Biol Cell 2007; 18:3214 - 23; http://dx.doi.org/10.1091/mbc.E06-12-1154; PMID: 17553930
- Avizienyte E, Frame MC. Src and FAK signalling controls adhesion fate and the epithelial-to-mesenchymal transition. Curr Opin Cell Biol 2005; 17:542 - 7; http://dx.doi.org/10.1016/j.ceb.2005.08.007; PMID: 16099634
- Frame MC. Src in cancer: deregulation and consequences for cell behaviour. Biochim Biophys Acta 2002; 1602:114 - 30; PMID: 12020799
- Avizienyte E, Wyke AW, Jones RJ, McLean GW, Westhoff MA, Brunton VG, et al. Src-induced de-regulation of E-cadherin in colon cancer cells requires integrin signalling. Nat Cell Biol 2002; 4:632 - 8; PMID: 12134161
- Avizienyte E, Fincham VJ, Brunton VG, Frame MC. Src SH3/2 domain-mediated peripheral accumulation of Src and phospho-myosin is linked to deregulation of E-cadherin and the epithelial-mesenchymal transition. Mol Biol Cell 2004; 15:2794 - 803; http://dx.doi.org/10.1091/mbc.E03-12-0879; PMID: 15075377
- Boguslavsky S, Grosheva I, Landau E, Shtutman M, Cohen M, Arnold K, et al. p120 catenin regulates lamellipodial dynamics and cell adhesion in cooperation with cortactin. Proc Natl Acad Sci U S A 2007; 104:10882 - 7; http://dx.doi.org/10.1073/pnas.0702731104; PMID: 17576929
- Reynolds AB. p120-catenin: Past and present. Biochim Biophys Acta 2007; 1773:2 - 7; http://dx.doi.org/10.1016/j.bbamcr.2006.09.019; PMID: 17175391
- Davis MA, Ireton RC, Reynolds AB. A core function for p120-catenin in cadherin turnover. J Cell Biol 2003; 163:525 - 34; http://dx.doi.org/10.1083/jcb.200307111; PMID: 14610055
- Playford MP, Schaller MD. The interplay between Src and integrins in normal and tumor biology. Oncogene 2004; 23:7928 - 46; http://dx.doi.org/10.1038/sj.onc.1208080; PMID: 15489911
- Schaller MD, Otey CA, Hildebrand JD, Parsons JT. Focal adhesion kinase and paxillin bind to peptides mimicking beta integrin cytoplasmic domains. J Cell Biol 1995; 130:1181 - 7; http://dx.doi.org/10.1083/jcb.130.5.1181; PMID: 7657702
- Chen LM, Bailey D, Fernandez-Valle C. Association of beta 1 integrin with focal adhesion kinase and paxillin in differentiating Schwann cells. J Neurosci 2000; 20:3776 - 84; PMID: 10804218
- Essayem S, Kovacic-Milivojevic B, Baumbusch C, McDonagh S, Dolganov G, Howerton K, et al. Hair cycle and wound healing in mice with a keratinocyte-restricted deletion of FAK. Oncogene 2006; 25:1081 - 9; http://dx.doi.org/10.1038/sj.onc.1209130; PMID: 16247468
- Schober M, Raghavan S, Nikolova M, Polak L, Pasolli HA, Beggs HE, et al. Focal adhesion kinase modulates tension signaling to control actin and focal adhesion dynamics. J Cell Biol 2007; 176:667 - 80; http://dx.doi.org/10.1083/jcb.200608010; PMID: 17325207
- Lahlou H, Sanguin-Gendreau V, Zuo D, Cardiff RD, McLean GW, Frame MC, et al. Mammary epithelial-specific disruption of the focal adhesion kinase blocks mammary tumor progression. Proc Natl Acad Sci U S A 2007; 104:20302 - 7; http://dx.doi.org/10.1073/pnas.0710091104; PMID: 18056629
- McLean GW, Komiyama NH, Serrels B, Asano H, Reynolds L, Conti F, et al. Specific deletion of focal adhesion kinase suppresses tumor formation and blocks malignant progression. Genes Dev 2004; 18:2998 - 3003; http://dx.doi.org/10.1101/gad.316304; PMID: 15601818
- Fan H, Guan J-L. Compensatory function of Pyk2 protein in the promotion of focal adhesion kinase (FAK)-null mammary cancer stem cell tumorigenicity and metastatic activity. J Biol Chem 2011; 286:18573 - 82; http://dx.doi.org/10.1074/jbc.M110.200717; PMID: 21471206
- Machacek M, Hodgson L, Welch C, Elliott H, Pertz O, Nalbant P, et al. Coordination of Rho GTPase activities during cell protrusion. Nature 2009; 461:99 - 103; http://dx.doi.org/10.1038/nature08242; PMID: 19693013
- Arthur WT, Noren NK, Burridge K. Regulation of Rho family GTPases by cell-cell and cell-matrix adhesion. Biol Res 2002; 35:239 - 46; http://dx.doi.org/10.4067/S0716-97602002000200016; PMID: 12415742
- Huveneers S, Danen EHJ. Adhesion signaling—crosstalk between integrins, Src and Rho. J Cell Sci 2009; 122:1059 - 69; http://dx.doi.org/10.1242/jcs.039446; PMID: 19339545
- Etienne-Manneville S, Hall A. Rho GTPases in cell biology. Nature 2002; 420:629 - 35; http://dx.doi.org/10.1038/nature01148; PMID: 12478284
- Heasman SJ, Ridley AJ. Mammalian Rho GTPases: new insights into their functions from in vivo studies. Nat Rev Mol Cell Biol 2008; 9:690 - 701; http://dx.doi.org/10.1038/nrm2476; PMID: 18719708
- Nimnual AS, Taylor LJ, Bar-Sagi D. Redox-dependent downregulation of Rho by Rac. Nat Cell Biol 2003; 5:236 - 41; http://dx.doi.org/10.1038/ncb938; PMID: 12598902
- Wheelock MJ, Johnson KR. Cadherin-mediated cellular signaling. Curr Opin Cell Biol 2003; 15:509 - 14; http://dx.doi.org/10.1016/S0955-0674(03)00101-7; PMID: 14519384
- Yap AS, Kovacs EM. Direct cadherin-activated cell signaling: a view from the plasma membrane. J Cell Biol 2003; 160:11 - 6; http://dx.doi.org/10.1083/jcb.200208156; PMID: 12507993
- McLachlan RW, Yap AS. Not so simple: the complexity of phosphotyrosine signaling at cadherin adhesive contacts. J Mol Med (Berl) 2007; 85:545 - 54; http://dx.doi.org/10.1007/s00109-007-0198-x; PMID: 17429596
- Huveneers S, Arslan S, van de Water B, Sonnenberg A, Danen EHJ. Integrins uncouple Src-induced morphological and oncogenic transformation. J Biol Chem 2008; 283:13243 - 51; http://dx.doi.org/10.1074/jbc.M800927200; PMID: 18326486
- Noren NK, Niessen CM, Gumbiner BM, Burridge K. Cadherin engagement regulates Rho family GTPases. J Biol Chem 2001; 276:33305 - 8; http://dx.doi.org/10.1074/jbc.C100306200; PMID: 11457821
- Kovacs EM, Ali RG, McCormack AJ, Yap AS. E-cadherin homophilic ligation directly signals through Rac and phosphatidylinositol 3-kinase to regulate adhesive contacts. J Biol Chem 2002; 277:6708 - 18; http://dx.doi.org/10.1074/jbc.M109640200; PMID: 11744701
- Goodwin M, Kovacs EM, Thoreson MA, Reynolds AB, Yap AS. Minimal mutation of the cytoplasmic tail inhibits the ability of E-cadherin to activate Rac but not phosphatidylinositol 3-kinase: direct evidence of a role for cadherin-activated Rac signaling in adhesion and contact formation. J Biol Chem 2003; 278:20533 - 9; http://dx.doi.org/10.1074/jbc.M213171200; PMID: 12672818
- Gavard J, Lambert M, Grosheva I, Marthiens V, Irinopoulou T, Riou J-F, et al. Lamellipodium extension and cadherin adhesion: two cell responses to cadherin activation relying on distinct signalling pathways. J Cell Sci 2004; 117:257 - 70; http://dx.doi.org/10.1242/jcs.00857; PMID: 14657280
- Kraemer A, Goodwin M, Verma S, Yap AS, Ali RG. Rac is a dominant regulator of cadherin-directed actin assembly that is activated by adhesive ligation independently of Tiam1. Am J Physiol Cell Physiol 2007; 292:C1061 - 9; http://dx.doi.org/10.1152/ajpcell.00073.2006; PMID: 17020937
- Warner SJ, Longmore GD. Cdc42 antagonizes Rho1 activity at adherens junctions to limit epithelial cell apical tension. J Cell Biol 2009; 187:119 - 33; http://dx.doi.org/10.1083/jcb.200906047; PMID: 19805632
- Gimond C, van Der Flier A, van Delft S, Brakebusch C, Kuikman I, Collard JG, et al. Induction of cell scattering by expression of beta1 integrins in beta1-deficient epithelial cells requires activation of members of the rho family of GTPases and downregulation of cadherin and catenin function. J Cell Biol 1999; 147:1325 - 40; http://dx.doi.org/10.1083/jcb.147.6.1325; PMID: 10601344
- Anastasiadis PZ, Moon SY, Thoreson MA, Mariner DJ, Crawford HC, Zheng Y, et al. Inhibition of RhoA by p120 catenin. Nat Cell Biol 2000; 2:637 - 44; http://dx.doi.org/10.1038/35023588; PMID: 10980705
- Anastasiadis PZ. p120-ctn: A nexus for contextual signaling via Rho GTPases. Biochim Biophys Acta 2007; 1773:34 - 46; http://dx.doi.org/10.1016/j.bbamcr.2006.08.040; PMID: 17028013
- Braga VM, Yap AS. The challenges of abundance: epithelial junctions and small GTPase signalling. Curr Opin Cell Biol 2005; 17:466 - 74; http://dx.doi.org/10.1016/j.ceb.2005.08.012; PMID: 16112561
- McCrea PD, Park J-I. Developmental functions of the P120-catenin sub-family. Biochim Biophys Acta 2007; 1773:17 - 33; http://dx.doi.org/10.1016/j.bbamcr.2006.06.009; PMID: 16942809
- Perez-Moreno M, Davis MA, Wong E, Pasolli HA, Reynolds AB, Fuchs E. p120-catenin mediates inflammatory responses in the skin. Cell 2006; 124:631 - 44; http://dx.doi.org/10.1016/j.cell.2005.11.043; PMID: 16469707
- Wildenberg GA, Dohn MR, Carnahan RH, Davis MA, Lobdell NA, Settleman J, et al. p120-catenin and p190RhoGAP regulate cell-cell adhesion by coordinating antagonism between Rac and Rho. Cell 2006; 127:1027 - 39; http://dx.doi.org/10.1016/j.cell.2006.09.046; PMID: 17129786
- Nelson WJ, Nusse R. Convergence of Wnt, beta-catenin, and cadherin pathways. Science 2004; 303:1483 - 7; http://dx.doi.org/10.1126/science.1094291; PMID: 15001769
- Clevers H. Wnt/beta-catenin signaling in development and disease. Cell 2006; 127:469 - 80; http://dx.doi.org/10.1016/j.cell.2006.10.018; PMID: 17081971
- Retta SF, Balzac F, Avolio M. Rap1: a turnabout for the crosstalk between cadherins and integrins. Eur J Cell Biol 2006; 85:283 - 93; http://dx.doi.org/10.1016/j.ejcb.2005.09.007; PMID: 16546572
- Balzac F, Avolio M, Degani S, Kaverina I, Torti M, Silengo L, et al. E-cadherin endocytosis regulates the activity of Rap1: a traffic light GTPase at the crossroads between cadherin and integrin function. J Cell Sci 2005; 118:4765 - 83; http://dx.doi.org/10.1242/jcs.02584; PMID: 16219685
- Bos JL, de Bruyn K, Enserink J, Kuiperij B, Rangarajan S, Rehmann H, et al. The role of Rap1 in integrin-mediated cell adhesion. Biochem Soc Trans 2003; 31:83 - 6; http://dx.doi.org/10.1042/BST0310083; PMID: 12546659
- Caron E. Cellular functions of the Rap1 GTP-binding protein: a pattern emerges. J Cell Sci 2003; 116:435 - 40; http://dx.doi.org/10.1242/jcs.00238; PMID: 12508104
- Dustin ML, Bivona TG, Philips MR. Membranes as messengers in T cell adhesion signaling. Nat Immunol 2004; 5:363 - 72; http://dx.doi.org/10.1038/ni1057; PMID: 15052266
- le Duc Q, Shi Q, Blonk I, Sonnenberg A, Wang N, Leckband D, et al. Vinculin potentiates E-cadherin mechanosensing and is recruited to actin-anchored sites within adherens junctions in a myosin II-dependent manner. J Cell Biol 2010; 189:1107 - 15; http://dx.doi.org/10.1083/jcb.201001149; PMID: 20584916
- Liu Z, Tan JL, Cohen DM, Yang MT, Sniadecki NJ, Ruiz SA, et al. Mechanical tugging force regulates the size of cell-cell junctions. Proc Natl Acad Sci U S A 2010; 107:9944 - 9; http://dx.doi.org/10.1073/pnas.0914547107; PMID: 20463286
- Birchmeier W, Behrens J. Cadherin expression in carcinomas: role in the formation of cell junctions and the prevention of invasiveness. Biochim Biophys Acta 1994; 1198:11 - 26; PMID: 8199193
- Janes SM, Watt FM. New roles for integrins in squamous-cell carcinoma. Nat Rev Cancer 2006; 6:175 - 83; http://dx.doi.org/10.1038/nrc1817; PMID: 16498442
- Thiery JP, Acloque H, Huang RYJ, Nieto MA. Epithelial-mesenchymal transitions in development and disease. Cell 2009; 139:871 - 90; http://dx.doi.org/10.1016/j.cell.2009.11.007; PMID: 19945376
- Conacci-Sorrell M, Zhurinsky J, Ben-Ze’ev A. The cadherin-catenin adhesion system in signaling and cancer. J Clin Invest 2002; 109:987 - 91; PMID: 11956233
- Hajra KM, Fearon ER. Cadherin and catenin alterations in human cancer. Genes Chromosomes Cancer 2002; 34:255 - 68; http://dx.doi.org/10.1002/gcc.10083; PMID: 12007186
- Guo W, Giancotti FG. Integrin signalling during tumour progression. Nat Rev Mol Cell Biol 2004; 5:816 - 26; http://dx.doi.org/10.1038/nrm1490; PMID: 15459662
- Thiery JP. Epithelial-mesenchymal transitions in tumour progression. Nat Rev Cancer 2002; 2:442 - 54; http://dx.doi.org/10.1038/nrc822; PMID: 12189386
- Petersen OW, Rønnov-Jessen L, Weaver VM, Bissell MJ. Differentiation and cancer in the mammary gland: shedding light on an old dichotomy. Adv Cancer Res 1998; 75:135 - 61; http://dx.doi.org/10.1016/S0065-230X(08)60741-1; PMID: 9709809
- Nelson CM, Khauv D, Bissell MJ, Radisky DC. Change in cell shape is required for matrix metalloproteinase-induced epithelial-mesenchymal transition of mammary epithelial cells. J Cell Biochem 2008; 105:25 - 33; http://dx.doi.org/10.1002/jcb.21821; PMID: 18506791
- Friedl P, Alexander S. Cancer invasion and the microenvironment: plasticity and reciprocity. Cell 2011; 147:992 - 1009; http://dx.doi.org/10.1016/j.cell.2011.11.016; PMID: 22118458
- Xu R, Boudreau A, Bissell MJ. Tissue architecture and function: dynamic reciprocity via extra- and intra-cellular matrices. Cancer Metastasis Rev 2009; 28:167 - 76; http://dx.doi.org/10.1007/s10555-008-9178-z; PMID: 19160017
- Razzell W, Wood W, Martin P. Swatting flies: modelling wound healing and inflammation in Drosophila. Dis Model Mech 2011; 4:569 - 74; http://dx.doi.org/10.1242/dmm.006825; PMID: 21810906
- Dvorak HF. Tumors: wounds that do not heal. Similarities between tumor stroma generation and wound healing. N Engl J Med 1986; 315:1650 - 9; PMID: 3537791
- Weber GF, Bjerke MA, DeSimone DW. A mechanoresponsive cadherin-keratin complex directs polarized protrusive behavior and collective cell migration. Dev Cell 2012; 22:104 - 15; http://dx.doi.org/10.1016/j.devcel.2011.10.013; PMID: 22169071
- Mercurio AM, Rabinovitz I. Towards a mechanistic understanding of tumor invasion--lessons from the alpha6beta 4 integrin. Semin Cancer Biol 2001; 11:129 - 41; http://dx.doi.org/10.1006/scbi.2000.0364; PMID: 11322832
- Breuss JM, Gallo J, DeLisser HM, Klimanskaya IV, Folkesson HG, Pittet JF, et al. Expression of the beta 6 integrin subunit in development, neoplasia and tissue repair suggests a role in epithelial remodeling. J Cell Sci 1995; 108:2241 - 51; PMID: 7673344
- Zutter MM, Santoro SA, Staatz WD, Tsung YL. Re-expression of the alpha 2 beta 1 integrin abrogates the malignant phenotype of breast carcinoma cells. Proc Natl Acad Sci U S A 1995; 92:7411 - 5; http://dx.doi.org/10.1073/pnas.92.16.7411; PMID: 7638207
- Huttenlocher A, Horwitz AR. Integrins in cell migration. Cold Spring Harb Perspect Biol 2011; 3:a005074; http://dx.doi.org/10.1101/cshperspect.a005074; PMID: 21885598
- Calvo F, Sahai E. Cell communication networks in cancer invasion. Curr Opin Cell Biol 2011; 23:621 - 9; http://dx.doi.org/10.1016/j.ceb.2011.04.010; PMID: 21570276
- Furue M. Epithelial tumor, invasion and stroma. Ann Dermatol 2011; 23:125 - 31; http://dx.doi.org/10.5021/ad.2011.23.2.125; PMID: 21747608
- Ungefroren H, Sebens S, Seidl D, Lehnert H, Hass R. Interaction of tumor cells with the microenvironment. Cell Commun Signal 2011; 9:18; http://dx.doi.org/10.1186/1478-811X-9-18; PMID: 21914164
- Mantovani A, Allavena P, Sica A, Balkwill F. Cancer-related inflammation. Nature 2008; 454:436 - 44; http://dx.doi.org/10.1038/nature07205; PMID: 18650914
- Morgan MR, Humphries MJ, Bass MD. Synergistic control of cell adhesion by integrins and syndecans. Nat Rev Mol Cell Biol 2007; 8:957 - 69; http://dx.doi.org/10.1038/nrm2289; PMID: 17971838
- Mueller MM, Fusenig NE. Tumor-stroma interactions directing phenotype and progression of epithelial skin tumor cells. Differentiation 2002; 70:486 - 97; http://dx.doi.org/10.1046/j.1432-0436.2002.700903.x; PMID: 12492491
- Erez N, Truitt M, Olson P, Arron ST, Hanahan D. Cancer-Associated Fibroblasts Are Activated in Incipient Neoplasia to Orchestrate Tumor-Promoting Inflammation in an NF-kappaB-Dependent Manner. Cancer Cell 2010; 17:135 - 47; http://dx.doi.org/10.1016/j.ccr.2009.12.041; PMID: 20138012
- Bhowmick NA, Chytil A, Plieth D, Gorska AE, Dumont N, Shappell S, et al. TGF-beta signaling in fibroblasts modulates the oncogenic potential of adjacent epithelia. Science 2004; 303:848 - 51; http://dx.doi.org/10.1126/science.1090922; PMID: 14764882
- Kalluri R, Zeisberg M. Fibroblasts in cancer. Nat Rev Cancer 2006; 6:392 - 401; http://dx.doi.org/10.1038/nrc1877; PMID: 16572188
- Hu M, Polyak K. Molecular characterisation of the tumour microenvironment in breast cancer. Eur J Cancer 2008; 44:2760 - 5; http://dx.doi.org/10.1016/j.ejca.2008.09.038; PMID: 19026532
- Olumi AF, Grossfeld GD, Hayward SW, Carroll PR, Tlsty TD, Cunha GR. Carcinoma-associated fibroblasts direct tumor progression of initiated human prostatic epithelium. Cancer Res 1999; 59:5002 - 11; PMID: 10519415
- Orimo A, Gupta PB, Sgroi DC, Arenzana-Seisdedos F, Delaunay T, Naeem R, et al. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell 2005; 121:335 - 48; http://dx.doi.org/10.1016/j.cell.2005.02.034; PMID: 15882617
- Pietras K, Pahler J, Bergers G, Hanahan D. Functions of paracrine PDGF signaling in the proangiogenic tumor stroma revealed by pharmacological targeting. PLoS Med 2008; 5:e19; http://dx.doi.org/10.1371/journal.pmed.0050019; PMID: 18232728
- Camps JL, Chang SM, Hsu TC, Freeman MR, Hong SJ, Zhau HE, et al. Fibroblast-mediated acceleration of human epithelial tumor growth in vivo. Proc Natl Acad Sci U S A 1990; 87:75 - 9; http://dx.doi.org/10.1073/pnas.87.1.75; PMID: 2296606
- Gribben J, Rosenwald A, Gascoyne R, Lenz G. Targeting the microenvironment. Leuk Lymphoma 2010; 51:Suppl 1 34 - 40; http://dx.doi.org/10.3109/10428194.2010.500072; PMID: 20815761
- Wu Y, Zhou BP. TNF-alpha/NF-kappaB/Snail pathway in cancer cell migration and invasion. Br J Cancer 2010; 102:639 - 44; http://dx.doi.org/10.1038/sj.bjc.6605530; PMID: 20087353
- Goitre L, Pergolizzi B, Ferro E, Trabalzini L, Retta SF. Molecular Crosstalk between Integrins and Cadherins: Do Reactive Oxygen Species Set the Talk?. J Signal Transduct 2012; 2012:807682; http://dx.doi.org/10.1155/2012/807682; PMID: 22203898
- Anastasiadis PZ, Reynolds AB. Regulation of Rho GTPases by p120-catenin. Curr Opin Cell Biol 2001; 13:604 - 10; http://dx.doi.org/10.1016/S0955-0674(00)00258-1; PMID: 11544030
- Ichii T, Takeichi M. p120-catenin regulates microtubule dynamics and cell migration in a cadherin-independent manner. Genes Cells 2007; 12:827 - 39; http://dx.doi.org/10.1111/j.1365-2443.2007.01095.x; PMID: 17584295
- Cheung LWT, Leung PCK, Wong AST. Cadherin switching and activation of p120 catenin signaling are mediators of gonadotropin-releasing hormone to promote tumor cell migration and invasion in ovarian cancer. Oncogene 2010; 29:2427 - 40; http://dx.doi.org/10.1038/onc.2009.523; PMID: 20118984
- Smalley-Freed WG, Efimov A, Short SP, Jia P, Zhao Z, Washington MK, et al. Adenoma formation following limited ablation of p120-catenin in the mouse intestine. PLoS One 2011; 6:e19880; http://dx.doi.org/10.1371/journal.pone.0019880; PMID: 21611205
- Stairs DB, Bayne LJ, Rhoades B, Vega ME, Waldron TJ, Kalabis J, et al. Deletion of p120-catenin results in a tumor microenvironment with inflammation and cancer that establishes it as a tumor suppressor gene. Cancer Cell 2011; 19:470 - 83; http://dx.doi.org/10.1016/j.ccr.2011.02.007; PMID: 21481789
- Perez-Moreno M, Song W, Pasolli HA, Williams SE, Fuchs E. Loss of p120 catenin and links to mitotic alterations, inflammation, and skin cancer. Proc Natl Acad Sci U S A 2008; 105:15399 - 404; http://dx.doi.org/10.1073/pnas.0807301105; PMID: 18809907
- Samuel MS, Lopez JI, McGhee EJ, Croft DR, Strachan D, Timpson P, et al. Actomyosin-mediated cellular tension drives increased tissue stiffness and β-catenin activation to induce epidermal hyperplasia and tumor growth. Cancer Cell 2011; 19:776 - 91; http://dx.doi.org/10.1016/j.ccr.2011.05.008; PMID: 21665151
- Kobielak A, Fuchs E. Links between α-catenin, NF-kappaB, and squamous cell carcinoma in skin. Proc Natl Acad Sci U S A 2006; 103:2322 - 7; http://dx.doi.org/10.1073/pnas.0510422103; PMID: 16452166
- Vasioukhin V, Bauer C, Degenstein L, Wise B, Fuchs E. Hyperproliferation and defects in epithelial polarity upon conditional ablation of alpha-catenin in skin. Cell 2001; 104:605 - 17; http://dx.doi.org/10.1016/S0092-8674(01)00246-X; PMID: 11239416
- Oloumi A, Syam S, Dedhar S. Modulation of Wnt3a-mediated nuclear beta-catenin accumulation and activation by integrin-linked kinase in mammalian cells. Oncogene 2006; 25:7747 - 57; http://dx.doi.org/10.1038/sj.onc.1209752; PMID: 16799642
- Novak A, Hsu SC, Leung-Hagesteijn C, Radeva G, Papkoff J, Montesano R, et al. Cell adhesion and the integrin-linked kinase regulate the LEF-1 and beta-catenin signaling pathways. Proc Natl Acad Sci U S A 1998; 95:4374 - 9; http://dx.doi.org/10.1073/pnas.95.8.4374; PMID: 9539744
- Oloumi A, McPhee T, Dedhar S. Regulation of E-cadherin expression and beta-catenin/Tcf transcriptional activity by the integrin-linked kinase. Biochim Biophys Acta 2004; 1691:1 - 15; http://dx.doi.org/10.1016/j.bbamcr.2003.12.002; PMID: 15053919
- Serrano I, McDonald PC, Lock FE, Dedhar S. Role of the integrin-linked kinase (ILK)/Rictor complex in TGFβ-1-induced epithelial-mesenchymal transition (EMT). Oncogene 2012; 31:1 - 12; http://dx.doi.org/10.1038/onc.2012.30; PMID: 21643006
- Oloumi A, Maidan M, Lock FE, Tearle H, McKinney S, Muller WJ, et al. Cooperative signaling between Wnt1 and integrin-linked kinase induces accelerated breast tumor development. Breast Cancer Res 2010; 12:R38; http://dx.doi.org/10.1186/bcr2592; PMID: 20565980
- Assi K, Patterson S, Dedhar S, Owen D, Levings M, Salh B. Role of epithelial integrin-linked kinase in promoting intestinal inflammation: effects on CCL2, fibronectin and the T cell repertoire. BMC Immunol 2011; 12:42; http://dx.doi.org/10.1186/1471-2172-12-42; PMID: 21806815
- Kim Y, Kugler MC, Wei Y, Kim KK, Li X, Brumwell AN, et al. Integrin alpha3beta1-dependent beta-catenin phosphorylation links epithelial Smad signaling to cell contacts. J Cell Biol 2009; 184:309 - 22; http://dx.doi.org/10.1083/jcb.200806067; PMID: 19171760
- Dzamba BJ, Jakab KR, Marsden M, Schwartz MA, DeSimone DW. Cadherin adhesion, tissue tension, and noncanonical Wnt signaling regulate fibronectin matrix organization. Dev Cell 2009; 16:421 - 32; http://dx.doi.org/10.1016/j.devcel.2009.01.008; PMID: 19289087
- Matsumoto S, Kikuchi A. Regulation of focal adhesion dynamics by Wnt5a signaling. Methods Mol Biol 2012; 839:215 - 27; http://dx.doi.org/10.1007/978-1-61779-510-7_17; PMID: 22218904
- Pechkovsky DV, Scaffidi AK, Hackett TL, Ballard J, Shaheen F, Thompson PJ, et al. Transforming growth factor beta1 induces alphavbeta3 integrin expression in human lung fibroblasts via a beta3 integrin-, c-Src-, and p38 MAPK-dependent pathway. J Biol Chem 2008; 283:12898 - 908; http://dx.doi.org/10.1074/jbc.M708226200; PMID: 18353785
- Margadant C, Sonnenberg A. Integrin-TGF-β crosstalk in fibrosis, cancer and wound healing. EMBO Rep 2010; 11:97 - 105; http://dx.doi.org/10.1038/embor.2009.276; PMID: 20075988
- Santibáñez JF, Olivares D, Guerrero J, Martínez J. Cyclic AMP inhibits TGFbeta1-induced cell-scattering and invasiveness in murine-transformed keratinocytes. Int J Cancer 2003; 107:715 - 20; http://dx.doi.org/10.1002/ijc.11457; PMID: 14566820
- Santibanez JF, Pérez-Gómez E, Fernandez-L A, Garrido-Martin EM, Carnero A, Malumbres M, et al. The TGF-beta co-receptor endoglin modulates the expression and transforming potential of H-Ras. Carcinogenesis 2010; 31:2145 - 54; http://dx.doi.org/10.1093/carcin/bgq199; PMID: 20884686
- Bierie B, Moses HL. Tumour microenvironment: TGFbeta: the molecular Jekyll and Hyde of cancer. Nat Rev Cancer 2006; 6:506 - 20; http://dx.doi.org/10.1038/nrc1926; PMID: 16794634
- Moustakas A, Heldin C-H. Non-Smad TGF-beta signals. J Cell Sci 2005; 118:3573 - 84; http://dx.doi.org/10.1242/jcs.02554; PMID: 16105881
- Zhang YE. Non-Smad pathways in TGF-beta signaling. Cell Res 2009; 19:128 - 39; http://dx.doi.org/10.1038/cr.2008.328; PMID: 19114990
- Ikushima H, Miyazono K. TGFbeta signalling: a complex web in cancer progression. Nat Rev Cancer 2010; 10:415 - 24; http://dx.doi.org/10.1038/nrc2853; PMID: 20495575
- Hasegawa Y, Takanashi S, Kanehira Y, Tsushima T, Imai T, Okumura K. Transforming growth factor-beta1 level correlates with angiogenesis, tumor progression, and prognosis in patients with nonsmall cell lung carcinoma. Cancer 2001; 91:964 - 71; http://dx.doi.org/10.1002/1097-0142(20010301)91:5<964::AID-CNCR1086>3.0.CO;2-O; PMID: 11251948
- Derynck R, Akhurst RJ, Balmain A. TGF-beta signaling in tumor suppression and cancer progression. Nat Genet 2001; 29:117 - 29; http://dx.doi.org/10.1038/ng1001-117; PMID: 11586292
- Birchmeier W, Brinkmann V, Niemann C, Meiners S, DiCesare S, Naundorf H, et al. Role of HGF/SF and c-Met in morphogenesis and metastasis of epithelial cells. Ciba Found Symp 1997; 212:230 - 40, discussion 240-6; PMID: 9524774
- de Rooij J, Kerstens A, Danuser G, Schwartz MA, Waterman-Storer CM. Integrin-dependent actomyosin contraction regulates epithelial cell scattering. J Cell Biol 2005; 171:153 - 64; http://dx.doi.org/10.1083/jcb.200506152; PMID: 16216928
- Wu K, Bonavida B. The activated NF-kappaB-Snail-RKIP circuitry in cancer regulates both the metastatic cascade and resistance to apoptosis by cytotoxic drugs. Crit Rev Immunol 2009; 29:241 - 54; PMID: 19538137
- Wu Y, Deng J, Rychahou PG, Qiu S, Evers BM, Zhou BP. Stabilization of snail by NF-kappaB is required for inflammation-induced cell migration and invasion. Cancer Cell 2009; 15:416 - 28; http://dx.doi.org/10.1016/j.ccr.2009.03.016; PMID: 19411070
- Mon NN, Ito S, Senga T, Hamaguchi M. FAK signaling in neoplastic disorders: a linkage between inflammation and cancer. Ann N Y Acad Sci 2006; 1086:199 - 212; http://dx.doi.org/10.1196/annals.1377.019; PMID: 17185517
- Sato H, Seiki M. Regulatory mechanism of 92 kDa type IV collagenase gene expression which is associated with invasiveness of tumor cells. Oncogene 1993; 8:395 - 405; PMID: 8426746
- Wielockx B, Lannoy K, Shapiro SD, Itoh T, Itohara S, Vandekerckhove J, et al. Inhibition of matrix metalloproteinases blocks lethal hepatitis and apoptosis induced by tumor necrosis factor and allows safe antitumor therapy. Nat Med 2001; 7:1202 - 8; http://dx.doi.org/10.1038/nm1101-1202; PMID: 11689884
- Bates RC, Mercurio AM. Tumor necrosis factor-alpha stimulates the epithelial-to-mesenchymal transition of human colonic organoids. Mol Biol Cell 2003; 14:1790 - 800; http://dx.doi.org/10.1091/mbc.E02-09-0583; PMID: 12802055
- Hagemann T, Wilson J, Kulbe H, Li NF, Leinster DA, Charles K, et al. Macrophages induce invasiveness of epithelial cancer cells via NF-kappa B and JNK. J Immunol 2005; 175:1197 - 205; PMID: 16002723
- Oguma K, Oshima H, Aoki M, Uchio R, Naka K, Nakamura S, et al. Activated macrophages promote Wnt signalling through tumour necrosis factor-alpha in gastric tumour cells. EMBO J 2008; 27:1671 - 81; http://dx.doi.org/10.1038/emboj.2008.105; PMID: 18511911
- Stemmer V, de Craene B, Berx G, Behrens J. Snail promotes Wnt target gene expression and interacts with beta-catenin. Oncogene 2008; 27:5075 - 80; http://dx.doi.org/10.1038/onc.2008.140; PMID: 18469861
- Hancock JT, Desikan R, Neill SJ. Hydrogen peroxide and nitric oxide in plant defence: revealing potential targets for oxidative stress tolerance?. Biofactors 2001; 15:99 - 101; http://dx.doi.org/10.1002/biof.5520150210; PMID: 12016335
- Gregg D, de Carvalho DD, Kovacic H. Integrins and coagulation: a role for ROS/redox signaling?. Antioxid Redox Signal 2004; 6:757 - 64; http://dx.doi.org/10.1089/1523086041361604; PMID: 15242557
- Bielefeld KA, Amini-Nik S, Whetstone H, Poon R, Youn A, Wang J, et al. Fibronectin and beta-catenin act in a regulatory loop in dermal fibroblasts to modulate cutaneous healing. J Biol Chem 2011; 286:27687 - 97; http://dx.doi.org/10.1074/jbc.M111.261677; PMID: 21652705
- Wong VW, Rustad KC, Akaishi S, Sorkin M, Glotzbach JP, Januszyk M, et al. Focal adhesion kinase links mechanical force to skin fibrosis via inflammatory signaling. Nat Med 2012; 18:148 - 52; http://dx.doi.org/10.1038/nm.2574; PMID: 22157678