Abstract
Synaptojanin 2 (SYNJ2) is a phosphatidylinositol (PI) phosphatase that controls two distinct functions, clathrin-mediated endocytosis and tumor cell invadopodia formation and invasion. Here, we identify a number of novel SYNJ2 binding partners, several of which have previously been shown to be necessary for invadopodia formation or clathrin-mediated endocytosis. We focus on Src family kinases. We found that Src phosphorylates SYNJ2 on Tyr490, thereby stimulating SYNJ2 5′-phosphatase activity in vitro. We also provide evidence that Src-mediated phosphorylation of SYNJ2 contributes to invadopodia formation.
Introduction
Synaptojanin (SYNJ) phosphatidylinositol (PI) phosphatases contain two distinct phosphatase domains, a Sac1 homology domain that acts on PI(3)P, PI(4)P and PI(3,5)P2 and an inositol 5′-phosphatase domain that acts on PI(4,5)P2 and PI(3,4,5)P3.Citation1 There are two synaptojanins in the human genome. The major splice isoform of SYNJ1 is brain-specific,Citation2 whereas SYNJ2 is expressed in a wide range of tissues.Citation3 The C-terminal domains of SYNJ 1 and 2 diverge in sequence. However, both contain proline-rich domains (PRDs) that mediate binding to SH3 domain-containing proteins. The C-terminal domain of SYNJ2, but not SYNJ1, also contains a Rac1-binding domain.Citation4
SYNJ1 and 2 play diverse roles in clathrin-mediated endocytosis. The primary role of SYNJ1 appears to be the uncoating of clathrin-coated vesicles,Citation5 although it also contributes to vesicle internalization.Citation6,Citation7 In contrast, synaptojanin 2 plays a critical role at an early step of clathrin-mediated endocytosis,Citation8 although it is possible that SYNJ2 also functions in vesicle decoating.
We also have shown that SYNJ2 is necessary for the invasive behavior of a number of different tumor cell types.Citation9 The role of SYNJ2 in tumor cell invasion is independent from its role in clathrin-mediated endocytosis and is likely to be mediated by its function in the formation of invadopodia.Citation9 SYNJ2 also is strongly enriched in invadopodia.Citation9 Tumor cell invadopodia are specialized membrane structures at which extracellular matrix degradation is concentrated.Citation10-Citation12 We note that invadopodia are similar in composition and structure to podosomes.Citation10 The term podosome is usually reserved for such structures found in normal cells and in Src-transformed fibroblasts, whereas the term invadopodium is restricted to cancer cells. How SYNJ2 regulates invadopodia formation remains to be elucidated. A number of proteins that are regulated by PI metabolism play key roles in invadopodia. These include Tks5 and cortactin.Citation13,Citation14 Thus, likely functions of SYNJ2 in invadopodia include regulating the localization and/or activity of these proteins.
SYNJ2 binds to a number of SH3 domain-containing proteins, including endophilin, amphiphysin and CIN85, all of which play key roles in the regulation of clathrin-mediated endocytosis.Citation15-Citation18 In this paper, we identify a number of novel SYNJ2 binding partners, several of which have been shown to be necessary for invadopodia formation. We focus on Src family kinases, as Src is a central regulator of invadopodia formation.Citation10,Citation12
Results
In order to identify novel SYNJ2 binding partners, we screened a library of purified SH3 domains immobilized on a membrane (TranSignal SH3 Domain Arrays, Panomics) using a His-tagged C-terminal domain of SYNJ2 that contains all five PRDs (). The list of positives () included two SH3 domain-containing protein that are known to interact with both SYNJ2 and SYNJ1, endophilin and Grb2, and several that previously have been shown to bind to SYNJ1, intersectin, Nck1 and SNX9.Citation15,Citation19-Citation22 Remarkably, five members of the Src family of kinases (SFK) were among the positives, Fyn, Hck, Lck, Lyn and Src. Src is known to be a key regulator of invadopodia.Citation10,Citation12 Cortactin, another protein that is necessary for invadopodia formationCitation14 also was among the positives.
Figure 1. Chemoluminescence detection of SYNJ2-interacting SH3 domains. TransSignal SH3 Domain Array membranes were incubated with a His-tagged C-terminal domain of SYNJ2 (GST-SYNJ2-C-His) and developed as described in Materials and Methods. Shown is SH3 Domain Array-I out of four arrays examined.
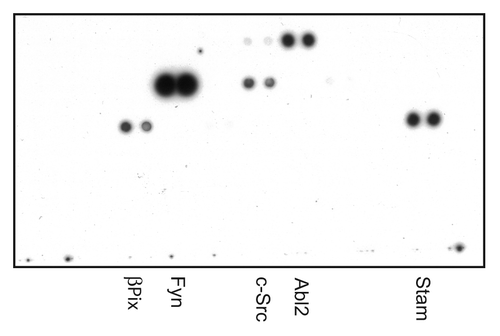
Table 1. Candidate SYNJ2 binding partners identified using protein arrays
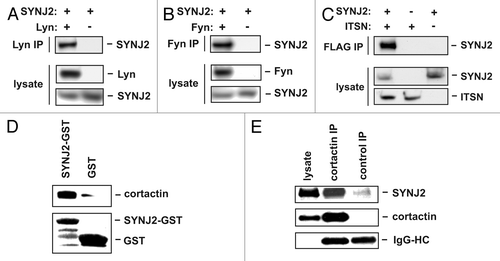
To confirm the interaction of SYNJ2 with Src family kinases in a cellular setting, we performed co-expression of full length SYNJ2 together with full length Lyn or Fyn in HEK293 cells. Immunoprecipitation of Lyn or Fyn from lysates of co-expressing cells, but not from cells in which SYNJ2 was overexpressed on its own, showed specific interaction between SYNJ2 and these kinases (). We obtained similar results for the interaction of SYNJ2 with intersectin (). We also confirmed the interaction between SYNJ2 and cortactin, using two different approaches. First, endogenous cortactin could be precipitated from SNB19 glioblastoma cell lysates with an immobilized SYNJ2 C-terminal domain GST-fusion protein, but not GST itself (). Second, SYNJ2 co-immunoprecipitated with cortactin from HEK293 cells co-expressing full length versions of both SYNJ2 and cortactin (). We also identified intersectin and cortactin using mass spectrometry, as endogenous proteins associated with SYNJ2 that was overexpressed in HEK293 cells (data not shown).
Figure 2. Confirmation of SYNJ2 interaction with select binding partners. (A) SYNJ2 interacts with Lyn. HEK293 cells were transiently co-transfected with expression plasmids for Lyn or SYNJ2 or transfected with a SYNJ2 plasmid alone. Cells were lysed 24 h post-transfection. Cell lysates were incubated with an anti-Lyn antibody, followed by protein A-agarose. Expression of Lyn and SYNJ2 in lysates and immunoprecipitated SYNJ2 were visualized by western blotting. (B) SYNJ2 interacts with Fyn. The same methodology was used as in (A), except that Lyn was replaced by Fyn. (C) SYNJ2 interacts with intersectin. HEK293 cells were transiently transfected with the indicated combinations of expression plasmids for FLAG-tagged intersectin-1 (ITSN) or SYNJ2. Cells were lysed 24 h post-transfection and lysates were incubated with an anti-FLAG antibody, followed by protein G-agarose. Expression of intersectin and SYNJ2 in lysates and immunoprecipitated SYNJ2 were visualized by western blotting. (D and E) SYNJ2 interacts with cortactin. (D) Co-immunoprecipitation experiment. HEK293 cells were transiently co-transfected with cortactin and SYNJ2 expression plasmids. Cells were lysed 24 h post-transfection and lysates were incubated either with an anti-cortactin antibody followed by protein G-sepharose or with control IgG followed by protein G-sepharose. Expression of cortactin and SYNJ2 in lysates and immunoprecipitated SYNJ2 were visualized by western blotting. Immunoglobulin heavy chain (IgG-HC) was visualized by anti-mouse HRP. (E) GST-pulldown experiment. SNB19 cell lysates were incubated with immobilized GST or GST-SJ2 (GST-SYNJ2-C). Precipitated cortactin was visualized by western blotting using an anti-cortactin antibody (upper panel) and bound GST proteins were visualized by Ponceau staining (lower panel). Data shown are representative of at least 2 experiments.
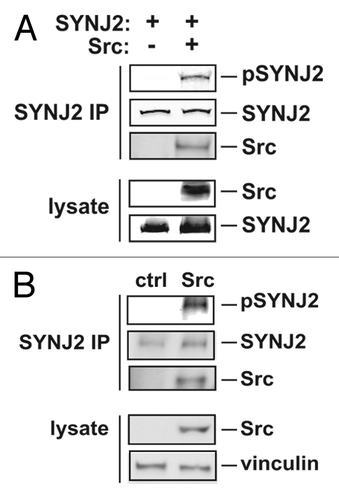
The interaction of SYNJ2 with Src family kinases suggested to us that Src may be phosphorylating SYNJ2 and thereby modulating its activity. To examine whether Src phosphorylates SYNJ2, we co-expressed full length SYNJ2 together with constitutively active Src in 293T cells, immunoprecipitated SYNJ2 with an anti-Myc antibody and determined tyrosine phosphorylation of SYNJ2 using an anti-phospho-tyrosine antibody. We observed robust Src-stimulated phosphorylation of SYNJ2 in these conditions (). We confirmed these findings by examining tyrosine phosphorylation of endogenous SYNJ2 by immunoprecipitating SYNJ2 from Src-transformed or control SNB19 glioblastoma cells ().
Figure 3. Src phosphorylates SYNJ2. (A) Tyrosine phosphorylation of SYNJ2 in Src-SYNJ2 co-expression conditions. Myc-tagged SYNJ2 was co-expressed with or without constitutively active Src (SrcY527F) in 293T cells and immunoprecipitated with an anti-Myc antibody. Tyrosine phosphorylated SYNJ2 in the immunoprecipitate was visualized by western blotting using an anti-phospho-tyrosine antibody (pSJ2). Total SYNJ2 and Src were visualized with their respective antibodies. (B) Tyrosine phosphorylation of SYNJ2 in Src-transformed cells. Endogenous SYNJ2 was immunoprecipitated from Src-transformed SNB19 glioblastoma cells or parental cells using an anti-SJ2 polyclonal antibody. Samples from cell lysates and immunoprecipitate were immunoblotted with the indicated antibodies. Data shown are representative of at least two experiments.
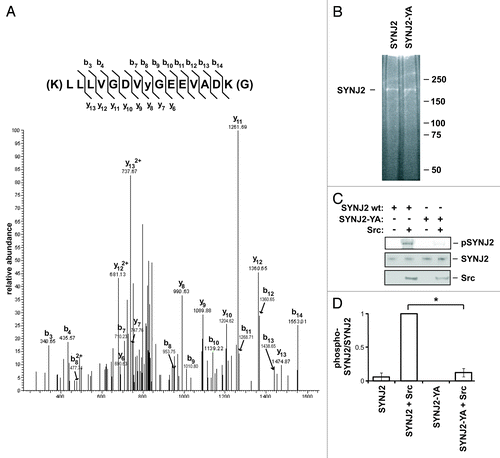
To identify SYNJ2 tyrosine residues that are phosphorylated by Src, we performed an in vitro phosphorylation assay using Myc-tagged SYNJ2 purified from 293T cells and recombinant His-tagged Src purified from Sf21 insect cells. Phosphorylated SYNJ2 peptides resulting from a trypsin digest were analyzed by tandem mass spectrometry coupled to liquid chromatography (LC-MS/MS). The only tyrosine-phosphorylated residue detected was Tyr490 () located between the Sac1 and 5′-phosphatase domains. To confirm that Tyr490 is indeed a substrate for Src phosphorylation, we mutated this residue to Ala using site-directed mutagenesis. We observed that the Src-stimulated in vitro tyrosine phosphorylation of the Y490A mutant was strongly inhibited in comparison to that of wild-type SYNJ2 ().
Figure 4. Src phosphorylates SYNJ2 on Tyr490. (A) Tandem mass spectrum of tryptic peptide containing Tyr490 where the phosphorylated tyrosine is denoted as “y.” SYNJ2 phosphorylated by Src in vitro was analyzed by LC-MS/MS as described in Materials and Methods. (B) Sypro Ruby-stained SDS-PAGE gel illustrating the purity of recombinant SYNJ2 and SYNJ2-Y490A purified from 293T cells. (C) Phosphorylation of wild-type SYNJ2 and SYNJ2-Y490A by Src in vitro. (D) Quantification of western blot data. Bars represent the average (± SEM) of three independent experiments (* = p < 0.005, two-tailed t-test).
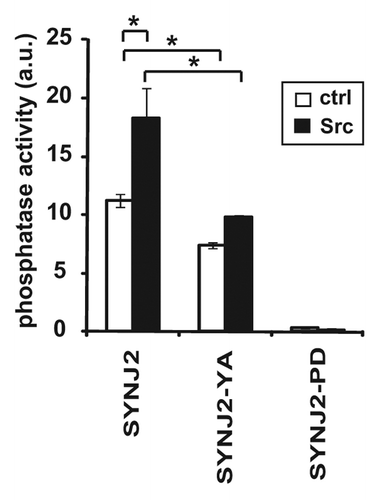
To examine the functional consequences of Src-mediated phosphorylation of SYNJ2, we determined whether Src stimulates SYNJ2 5′-phosphatase activity using a fluorescence polarization-based, competitive assay that quantifies the production of PI(3,4)P2 from PtdInd(3,4,5)P3. We found that incubation with Src significantly enhanced SYNJ2 5′-phosphatase activity (). In addition, basal 5′-phosphatase activity of SYNJ2-Y490A was lower than that of wild-type SYNJ2 and Src only marginally stimulated this activity. These results demonstrate that Src-stimulated phosphorylation of Tyr490 enhances SYNJ2 5′-phosphatase activity.
Figure 5. Src stimulates SYNJ2 5′-phosphatase activity. Wild-type SYNJ2 and SYNJ2-Y490A were phosphorylated in vitro by Src. Subsequently, the 5′-phosphatase activities of wild-type and mutant SYNJ2 proteins were determined using a fluorescence polarization assay as described in Materials and Methods. A 5′-phosphatase-deficient mutant of SYNJ2 (SJ2-PD) was used as negative control. Data shown are the average (± SEM) of triplicate wells and are representative of at least three independent experiments (*, p < 0.05, two-tailed t-test).
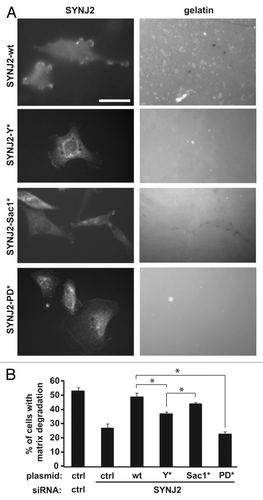
To further examine whether Src-mediated phosphorylation of SYNJ2 contributes to the formation of invadopodia, we simultaneously depleted endogenous SYNJ2 from SNB19 cells and reconstituted with wild-type or mutant versions of SYNJ2. SiRNA-mediated depletion of SYNJ2 was accomplished using a 3′ UTR-targeting oligo that effectively depletes endogenous SYNJ2 (by approximately 90%), but does not target the respective reconstituted proteins. We observed that wild-type SYNJ2 largely reversed the inhibitory effect of depleting endogenous SYNJ2, whereas the Y490A mutant was much less effective (). In contrast, the 5′-phosphatase-deficient mutant was completely ineffective in reconstituting invadopodia formation. Reconstitution by the Sac1-deficient mutant was not significantly different from that of wild-type SYNJ2. These results strongly suggest that Src-mediated phosphorylation of Tyr490 contributes to invadopodia formation by stimulating SYNJ2 5′-phosphatase activity.
Figure 6. Src-dependent stimulation of SYNJ2 5′-phosphatase activity regulates invadopodia formation. (A) Representative micrographs showing reconstitution of depleted endogenous SYNJ2 from SNB19 cells with either wild-type or mutant versions of SYNJ2 (left panels) and cell-associated matrix degradation (right panels). SNB19 cells were first transfected with an siRNA that targets the SYNJ2 3′ UTR and the next day transfected with the indicated plasmids. Control cells were similarly transfected with control siRNA (targeting luciferase) and a pcDNA-Myc plasmid. Two days after transfection, cells were plated on FITC-gelatin-coated coverslips and fixed 21 h later to assay invadopodia formation, as described in Materials and Methods. (B) Quantification of invadopodia formation. For each experiment and condition, the means were determined for at least 10 fields comprising a total of at least 25 cells. Data shown are the average values (± SEM) obtained from at least three independent experiments (*, p < 0.05, two-tailed t-test).
Discussion
In this paper, we identify a number of novel binding partners of the SYNJ2 PI phosphatase and focus on Src family kinases. We show that Src phosphorylates SYNJ2, thereby enhancing its 5′-phosphatase activity. We also provide evidence that Src-mediated stimulation of SYNJ2 activity contributes to invadopodia formation.
Src is a central regulator of invadopodia and a number of Src substrates have been shown to be essential for invadopodia formation, including cortactin and Tks5/FishCitation10-Citation12 Cortactin is an F-actin-binding protein and activates the Arp2/3 actin-nucleation complex.Citation23 Cortactin also is thought to stimulate protease secretion by regulating vesicular trafficking.Citation24 We note that cortactin also plays a role in clathrin-mediated endocytosis.Citation25 Tks5 is a scaffold protein that contains five SH3 domains and is targeted to invadopodia via its PX domain that binds to PtdIns(3)P and PtdIns(3,4)P2.Citation13 Tks5 binds to members of the ADAM family of transmembrane proteases and to proteins involved in the regulation of actin dynamics, including cortactin and N-WASP.Citation13,Citation26 The data presented in this paper add SYNJ2 to the array of Src substrates that control invadopodia formation.
Interestingly, kinetic analysis of Src-induced podosome formation in NIH-3T3 cells has indicated that clustering of PtdIns(3,4)P2 is an early and necessary event in podosome formation.Citation26 In addition, both PI3K and SYNJ2 have been shown to be necessary for the formation of podosomes and invadopodia.Citation26,Citation27 Importantly, we also showed that it is the 5′-phosphatase, but not Sac1 enzymatic activity of SYNJ2 that regulates invadopodia formation. Thus, these findings, together with our observation that Src stimulates SYNJ2 5′-phosphatase activity, strongly suggest that SYNJ2 stimulates invadopodia/podosome formation by producing PtdIns(3,4)P2 through hydrolysis of PtdIns(3,4,5)P3 and subsequent recruitment of Tks5. It is also possible however that SYNJ2 contributes to invadopodia formation by hydrolyzing PtdIns(4,5)P2. Indeed, type-I phosphatidylinositol 4-phosphate 5-kinase α (PIP5KI α) has been shown to be essential for invadopodia formationCitation28 and a number of proteins that have been implicated in invadopodia formation are regulated by PtdIns(4,5)P2, including the actin filament severing protein cofilin and N-WASP, a protein that regulates actin nucleation via Arp2/3.Citation29,Citation30
We also note that both Src and SYNJ2 play critical roles in clathrin-mediated endocytosis.Citation8,Citation31,Citation32 Thus, it is likely that Src-stimulated SYNJ2 5′-phosphatase activity also contributes to this function.
In addition to Src family members and cortactin, we have identified two other potential binding partners of SYNJ2 that have been implicated in the formation of invadopodia and/or podosomes, including Abl2/Arg and PSTPIP1. The tyrosine kinase Arg has been shown to function downstream of Src in the functional maturation of invadopodia, at least in part by phosphorylating cortactin and stimulating actin nucleation.Citation33 PSTPIP1 is an adaptor protein that is primarily expressed in hematopoietic cells and also controls the actin cytoskeleton, in part via its interaction with the protein tyrosine phosphatase PTP-PEST and the actin nucleating protein WASP.Citation34 Mutations in PSTPIP1 that disrupt binding to PTP-PEST are responsible for pyogenic sterile arthritis, pyoderma gangrenosum and acne (PAPA) syndrome, an auto-inflammatory disorder.Citation34 Recently, PSTPIP1 has been shown to function in podosome formation and macrophage migration.Citation35
Our screen also identified intersectin and SNX9, two proteins that have been implicated in the regulation of clathrin-mediated endocytosis, as novel binding partners of SYNJ2. Intersectin is a multi-domain adaptor protein that is thought to stabilize an endocytic multi-protein complex.Citation36,Citation37 SNX9 is a member of the sorting nexin family and plays multiple roles in clathrin-mediated endocytosis, including regulation of dynamin acitivity and membrane curvature.Citation38 Whether intersectin and SNX9 also function in the formation of podosomes and invadopodia remains to be established.
In conclusion, our findings shed new light on the molecular mechanisms that regulate SYNJ2. In addition, the identification of a large number of SYNJ2 binding partners strongly suggests that, in addition to regulating PI metabolism, SYNJ2 may function as a scaffold to coordinate signaling events that regulate clathrin mediated endocytosis and the formation of podosomes and invadopodia.
Materials and Methods
Reagents
Plasmids
A Myc-tagged wild-type SYNJ2 plasmid (pcDNA-Myc-SYNJ2) was constructed by inserting a PCR product containing the full length SYNJ2 gene into the ClaI/EcoRV sites of the pCAN-Myc1 vector. The following mutant versions of SYNJ2 were introduced into pcDNA-Myc-SYNJ2 using the QuickChange Site-Directed Mutagenesis Kit (Stratagene): Src-phosphorylation mutant (SYNJ2-Y490A), Sac1 mutant (SYNJ2-C386A) and 5′-phosphatase mutant (SYNJ2-R796A/R803F).
GST-SYNJ2-C, containing most of the SYNJ2 C-terminal domain (aa 1,023–1,497), was subcloned from pEXV-2H28Citation4 into the EcoRI site of pFASTBac (Invitrogen) and subsequentially into the BamHI/NotI sites of pET-42 (Novagen). GST-SYNJ2-C-His, a shorter version of the SYNJ2 C-terminal domain that still contains all five PRDs (aa 1,051–1,381), was generated by PCR and subcloned into pET-42b. pRK5-intersectin-1 was a gift from Nathalie Lamarche-Vane.Citation39 The cortactin-expression plasmid pEMS1 was a gift from Ed Schuuring. pRK5-Fyn was purchased from Adgene. pDONR223-Lyn was purchased from Adgene and subcloned into pRN and subsequently into pCDNA.
Small interfering RNA
An siRNA targeting the SYNJ2 3′ UTR (target sequence: 5′-AAUCACCAGGCCAGUGUUUCU) was designed using the Dharmacon web tool and obtained from Dharmacon. An siRNA directed against GL2 luciferase with target sequence 5′- AACGTACGCGGAATACTTCGATT was purchased from Ambion.
Proteins
Recombinant His-tagged Src protein was obtained from Millipore (14-326). Myc-SYNJ2 and Myc-SYNJ2-Y490A were expressed in 293T cells and purified using the c-Myc tagged Protein Mild Purification Kit (MBL) following the protocol provided by the manufacturer.
Antibodies
Polyclonal antibodies against SYNJ2 are as previously described.Citation4 Anti-cortactin (4F11, 05-180) and anti-Src (GD11, 05-184) antibodies were obtained from Millipore. Anti-Fyn (Sc-16) and anti-Lyn (Sc-15) antibodies were obtained from Santa Cruz Biotechnology. Anti-HA (1 666 606) and anti-His6-Peroxidase (1 965 085) antibodies were from Roche. The anti-FLAG M2 antibody (200472) was from Stratagene. The anti-Myc antibody (1 667 203) used for the immunofluorescence experiments was from Roche. To visualize phospho-tyrosine we used a mixture of antibodies, 4G10 (05-321, Millipore) and PY20 (610000, BD Biosciences).
Cell culture
SNB19 cells were grown in Dulbecco’s modified Eagle’s medium (DMEM), supplemented with 10% fetal bovine serum and penicillin/streptomycin at 37°C in a humidified incubator containing 5% CO2. SNB19 cells that stably express constitutively active chicken Src were established by co-transfection of the pSGT-SrcY527F plasmid (gift from Sara Courtneidge) and pRL-puro (Clontech) for puromycin selection. Clonal populations were selected in 0.75 μg/ml of puromycin. Isolated clones were characterized by Src immunoblotting.
SH3 domain array screening
TransSignal SH3 Domain Array membranes (Panomics) were incubated with bacterial lysates containing GST-SYNJ2-C-His protein. The membranes were washed and developed according to the protocol provided by the manufacturer.
In vitro SYNJ2 phosphorylation assay
Two micrograms of Myc-SYNJ2 or Myc-SYNJ2-Y490A protein were incubated with 20 ng of Src in kinase buffer (60 mM HEPES pH 7.5, 5 mM MgCl2, 5 mM MnCl2 and 3 μM Na3VO4) with 1.25 mM DTT and 20 μM of ATP for 30 min at room temperature. The reaction was stopped by addition of Laemmli sample buffer (Bio-Rad Laboratory). Subsequently, proteins were visualized using 1D SDS-PAGE, followed by western blotting or Sypro Ruby staining (for mass spectrometric analysis).
Mass spectrometry
The Sypro Ruby-stained band corresponding to SYNJ2 was digested in trypsin (10 μg/mL) at 37°C overnight and tandem mass spectrometry coupled to liquid chromatography (LC-MS/MS) analysis was performed as described previously.Citation40 Following positive protein identification of the SYNJ2 2B2 isoform by protein database searching as described previously,Citation40 the tandem mass spectra were also searched in a biased way using the primary sequence of SYNJ2 2B2, considering additional variable modifications (addition of +79.999 on serine, threonine and tyrosine residues). The tandem mass spectra of modified peptides from this search were visually inspected for good quality. In addition, their appropriate data files were further re-searched in an unbiased way using the ipihuman v 3.65 protein database, considering the same additional variable modifications, in order to ensure that these spectra did not correlate better to other human proteins.
5′-phosphatase assay
SYNJ2 5′-phosphatase activity was measured using the 5′-PI(3,4,5)P3 Phosphatase Fluorescence Polarization Activity Assay kit (Echelon) according to the manufacturer’s protocol. The assay is a competitive assay for the product of the phosphatase reaction PI(3,4)P2. Briefly, 50 ng of purified wild-type SYNJ2 or mutant SYNJ2-Y490A, was first in vitro phosphorylated (or not) by Src (7 ng Src protein in kinase buffer + 1 mM DTT, 100 μM ATP) for 30 min at room temperature. Control reaction was performed in kinase buffer without Src protein. Next, samples were diluted into the kit-provided reaction buffer and subsequently mixed with PIP3 substrate for 30–40 min at room temperature. The 5′-phosphatase reaction was stopped by heating the sample at 95°C for 3 min. Subsequently, the PI(3,4)P2 detector and the fluorescent PI(3,4)P2 probe were added and incubated for 30 min. Fluorescence polarization was examined using a VictorX plate reader provided with 550 nm excitation and 580 nm polarizing emission filters.
Invadopodia formation assay
FITC-gelatin-coated coverslips were prepared as described previouslyCitation41 and modified for SNB19 cells.Citation9 Cells were cultured on FITC-gelatin-coated coverslips in DMEM with 0.25% FBS for 24 h. Subsequently, cells were fixed in 4% formaldehyde/PBS and subsequently, permeabilized with 0.1% Triton X-100/PBS. Control cells were stained with Rhodamine-conjugated phalloidin and cells that express recombinant proteins were stained with an anti-Myc antibody, followed by a Texas Red anti-mouse antibody. Coverslips were mounted in ProlongGold mounting medium (Invitrogen) and images were collected using an IX70 Olympus inverted microscope equipped with a 60× objective, an Orca II CCD camera (Hamamatsu) and ESee (Inovision) image analysis software. Invadopodia formation was determined by counting the percentage of cells that had associated matrix degradation. For the reconstitution studies, only cells that express low to medium levels of the respective SYNJ2 constructs were analyzed.
| Abbreviations: | ||
| LC-MS/MS | = | tandem mass spectrometry coupled to liquid chromatography |
| PI | = | phosphatidylinositol |
| PRD | = | proline-rich domain |
| SYNJ2 | = | synaptojanin 2 |
Acknowledgments
The authors thank Sara Courtneidge (Sanford-Burnham Medical Research Institute) for stimulating discussions and a gift of pSGT-SrcY527F, Nathalie Lamarche (McGill University) for a gift of pRK5-intersectin and Ed Schuuring (University Medical Center Groningen) for a gift of pEMS1. This work was supported by NIH grants CA87567 (M.S.), ES06694 (SWEHSC) and CA023074 (AZCC) and grants from the Virginia G. Piper Charitable Trust (K.P.) and the Flinn Foundation of Arizona (K.P.).
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Related Research Data
References
- Mitchell CA, Gurung R, Kong AM, Dyson JM, Tan A, Ooms LM. Inositol polyphosphate 5-phosphatases: lipid phosphatases with flair. IUBMB Life 2002; 53:25 - 36; http://dx.doi.org/10.1080/15216540210815; PMID: 12018404
- Ramjaun AR, McPherson PS. Tissue-specific alternative splicing generates two synaptojanin isoforms with differential membrane binding properties. J Biol Chem 1996; 271:24856 - 61; http://dx.doi.org/10.1074/jbc.271.40.24856; PMID: 8798761
- Nemoto Y, Arribas M, Haffner C, DeCamilli P. Synaptojanin 2, a novel synaptojanin isoform with a distinct targeting domain and expression pattern. J Biol Chem 1997; 272:30817 - 21; http://dx.doi.org/10.1074/jbc.272.49.30817; PMID: 9388224
- Malecz N, McCabe PC, Spaargaren C, Qiu R, Chuang Y, Symons M. Synaptojanin 2, a novel Rac1 effector that regulates clathrin-mediated endocytosis. [In Process Citation] Curr Biol 2000; 10:1383 - 6; http://dx.doi.org/10.1016/S0960-9822(00)00778-8; PMID: 11084340
- Cremona O, Di Paolo G, Wenk MR, Lüthi A, Kim WT, Takei K, et al. Essential role of phosphoinositide metabolism in synaptic vesicle recycling. Cell 1999; 99:179 - 88; http://dx.doi.org/10.1016/S0092-8674(00)81649-9; PMID: 10535736
- Dickman DK, Horne JA, Meinertzhagen IA, Schwarz TL. A slowed classical pathway rather than kiss-and-run mediates endocytosis at synapses lacking synaptojanin and endophilin. Cell 2005; 123:521 - 33; http://dx.doi.org/10.1016/j.cell.2005.09.026; PMID: 16269341
- Mani M, Lee SY, Lucast L, Cremona O, Di Paolo G, De Camilli P, et al. The dual phosphatase activity of synaptojanin1 is required for both efficient synaptic vesicle endocytosis and reavailability at nerve terminals. Neuron 2007; 56:1004 - 18; http://dx.doi.org/10.1016/j.neuron.2007.10.032; PMID: 18093523
- Rusk N, Le PU, Mariggio S, Guay G, Lurisci C, Nabi IR, et al. Synaptojanin 2 functions at an early step of clathrin-mediated endocytosis. Curr Biol 2003; 13:659 - 63; http://dx.doi.org/10.1016/S0960-9822(03)00241-0; PMID: 12699622
- Chuang YY, Tran NL, Rusk N, Nakada M, Berens ME, Symons M. Role of synaptojanin 2 in glioma cell migration and invasion. Cancer Res 2004; 64:8271 - 5; http://dx.doi.org/10.1158/0008-5472.CAN-04-2097; PMID: 15548694
- Gimona M, Buccione R, Courtneidge SA, Linder S. Assembly and biological role of podosomes and invadopodia. Curr Opin Cell Biol 2008; 20:235 - 41; http://dx.doi.org/10.1016/j.ceb.2008.01.005; PMID: 18337078
- Stylli SS, Kaye AH, Lock P. Invadopodia: at the cutting edge of tumour invasion. J Clin Neurosci 2008; 15:725 - 37; http://dx.doi.org/10.1016/j.jocn.2008.03.003; PMID: 18468901
- Destaing O, Block MR, Planus E, Albiges-Rizo C. Invadosome regulation by adhesion signaling. Curr Opin Cell Biol 2011; 23:597 - 606; http://dx.doi.org/10.1016/j.ceb.2011.04.002; PMID: 21550788
- Abram CL, Seals DF, Pass I, Salinsky D, Maurer L, Roth TM, et al. The adaptor protein fish associates with members of the ADAMs family and localizes to podosomes of Src-transformed cells. J Biol Chem 2003; 278:16844 - 51; http://dx.doi.org/10.1074/jbc.M300267200; PMID: 12615925
- Artym VV, Zhang Y, Seillier-Moiseiwitsch F, Yamada KM, Mueller SC. Dynamic interactions of cortactin and membrane type 1 matrix metalloproteinase at invadopodia: defining the stages of invadopodia formation and function. Cancer Res 2006; 66:3034 - 43; http://dx.doi.org/10.1158/0008-5472.CAN-05-2177; PMID: 16540652
- Nemoto Y, Wenk MR, Watanabe M, Daniell L, Murakami T, Ringstad N, et al. Identification and characterization of a synaptojanin 2 splice isoform predominantly expressed in nerve terminals. J Biol Chem 2001; 276:41133 - 42; http://dx.doi.org/10.1074/jbc.M106404200; PMID: 11498538
- Song W, Zinsmaier KE. Endophilin and synaptojanin hook up to promote synaptic vesicle endocytosis. Neuron 2003; 40:665 - 7; http://dx.doi.org/10.1016/S0896-6273(03)00726-8; PMID: 14622570
- Gallop JL, McMahon HT. BAR domains and membrane curvature: bringing your curves to the BAR. Biochem Soc Symp 2005; 223 - 31; PMID: 15649145
- Kowanetz K, Husnjak K, Höller D, Kowanetz M, Soubeyran P, Hirsch D, et al. CIN85 associates with multiple effectors controlling intracellular trafficking of epidermal growth factor receptors. Mol Biol Cell 2004; 15:3155 - 66; http://dx.doi.org/10.1091/mbc.E03-09-0683; PMID: 15090612
- Adayev T, Chen-Hwang MC, Murakami N, Wang R, Hwang YW. MNB/DYRK1A phosphorylation regulates the interactions of synaptojanin 1 with endocytic accessory proteins. Biochem Biophys Res Commun 2006; 351:1060 - 5; http://dx.doi.org/10.1016/j.bbrc.2006.10.169; PMID: 17097615
- Zhao ZS, Manser E, Lim L. Interaction between PAK and nck: a template for Nck targets and role of PAK autophosphorylation. Mol Cell Biol 2000; 20:3906 - 17; http://dx.doi.org/10.1128/MCB.20.11.3906-3917.2000; PMID: 10805734
- So CW, Sham MH, Chew SL, Cheung N, So CK, Chung SK, et al. Expression and protein-binding studies of the EEN gene family, new interacting partners for dynamin, synaptojanin and huntingtin proteins. Biochem J 2000; 348:447 - 58; http://dx.doi.org/10.1042/0264-6021:3480447; PMID: 10816441
- Yeow-Fong L, Lim L, Manser E. SNX9 as an adaptor for linking synaptojanin-1 to the Cdc42 effector ACK1. FEBS Lett 2005; 579:5040 - 8; http://dx.doi.org/10.1016/j.febslet.2005.07.093; PMID: 16137687
- Weaver AM, Karginov AV, Kinley AW, Weed SA, Li Y, Parsons JT, et al. Cortactin promotes and stabilizes Arp2/3-induced actin filament network formation. Curr Biol 2001; 11:370 - 4; http://dx.doi.org/10.1016/S0960-9822(01)00098-7; PMID: 11267876
- Clark ES, Weaver AM. A new role for cortactin in invadopodia: regulation of protease secretion. Eur J Cell Biol 2008; 87:581 - 90; http://dx.doi.org/10.1016/j.ejcb.2008.01.008; PMID: 18342393
- Zhu J, Yu D, Zeng XC, Zhou K, Zhan X. Receptor-mediated endocytosis involves tyrosine phosphorylation of cortactin. J Biol Chem 2007; 282:16086 - 94; http://dx.doi.org/10.1074/jbc.M701997200; PMID: 17420251
- Oikawa T, Itoh T, Takenawa T. Sequential signals toward podosome formation in NIH-src cells. J Cell Biol 2008; 182:157 - 69; http://dx.doi.org/10.1083/jcb.200801042; PMID: 18606851
- Yamaguchi H, Yoshida S, Muroi E, Yoshida N, Kawamura M, Kouchi Z, et al. Phosphoinositide 3-kinase signaling pathway mediated by p110α regulates invadopodia formation. J Cell Biol 2011; 193:1275 - 88; http://dx.doi.org/10.1083/jcb.201009126; PMID: 21708979
- Yamaguchi H, Yoshida S, Muroi E, Kawamura M, Kouchi Z, Nakamura Y, et al. Phosphatidylinositol 4,5-bisphosphate and PIP5-kinase Ialpha are required for invadopodia formation in human breast cancer cells. Cancer Sci 2010; 101:1632 - 8; http://dx.doi.org/10.1111/j.1349-7006.2010.01574.x; PMID: 20426790
- Yonezawa N, Nishida E, Iida K, Yahara I, Sakai H. Inhibition of the interactions of cofilin, destrin, and deoxyribonuclease I with actin by phosphoinositides. J Biol Chem 1990; 265:8382 - 6; PMID: 2160454
- Miki H, Miura K, Takenawa T. N-WASP, a novel actin-depolymerizing protein, regulates the cortical cytoskeletal rearrangement in a PIP2-dependent manner downstream of tyrosine kinases. EMBO J 1996; 15:5326 - 35; PMID: 8895577
- Ahn S, Kim J, Lucaveche CL, Reedy MC, Luttrell LM, Lefkowitz RJ, et al. Src-dependent tyrosine phosphorylation regulates dynamin self-assembly and ligand-induced endocytosis of the epidermal growth factor receptor. J Biol Chem 2002; 277:26642 - 51; http://dx.doi.org/10.1074/jbc.M201499200; PMID: 12011079
- Cao H, Chen J, Krueger EW, McNiven MA. SRC-mediated phosphorylation of dynamin and cortactin regulates the “constitutive” endocytosis of transferrin. Mol Cell Biol 2010; 30:781 - 92; http://dx.doi.org/10.1128/MCB.00330-09; PMID: 19995918
- Mader CC, Oser M, Magalhaes MA, Bravo-Cordero JJ, Condeelis J, Koleske AJ, et al. An EGFR-Src-Arg-cortactin pathway mediates functional maturation of invadopodia and breast cancer cell invasion. Cancer Res 2011; 71:1730 - 41; http://dx.doi.org/10.1158/0008-5472.CAN-10-1432; PMID: 21257711
- Smith EJ, Allantaz F, Bennett L, Zhang D, Gao X, Wood G, et al. Clinical, Molecular, and Genetic Characteristics of PAPA Syndrome: A Review. Curr Genomics 2010; 11:519 - 27; http://dx.doi.org/10.2174/138920210793175921; PMID: 21532836
- Cortesio CL, Wernimont SA, Kastner DL, Cooper KM, Huttenlocher A. Impaired podosome formation and invasive migration of macrophages from patients with a PSTPIP1 mutation and PAPA syndrome. Arthritis Rheum 2010; 62:2556 - 8; http://dx.doi.org/10.1002/art.27521; PMID: 20506269
- Marie B, Sweeney ST, Poskanzer KE, Roos J, Kelly RB, Davis GW. Dap160/intersectin scaffolds the periactive zone to achieve high-fidelity endocytosis and normal synaptic growth. Neuron 2004; 43:207 - 19; http://dx.doi.org/10.1016/j.neuron.2004.07.001; PMID: 15260957
- Koh TW, Verstreken P, Bellen HJ. Dap160/intersectin acts as a stabilizing scaffold required for synaptic development and vesicle endocytosis. Neuron 2004; 43:193 - 205; http://dx.doi.org/10.1016/j.neuron.2004.06.029; PMID: 15260956
- Lundmark R, Carlsson SR. SNX9 - a prelude to vesicle release. J Cell Sci 2009; 122:5 - 11; http://dx.doi.org/10.1242/jcs.037135; PMID: 19092055
- Hussain NK, Jenna S, Glogauer M, Quinn CC, Wasiak S, Guipponi M, et al. Endocytic protein intersectin-l regulates actin assembly via Cdc42 and N-WASP. Nat Cell Biol 2001; 3:927 - 32; http://dx.doi.org/10.1038/ncb1001-927; PMID: 11584276
- Zhang J, Dinh TN, Kappeler K, Tsaprailis G, Chen QM. La autoantigen mediates oxidant induced de novo Nrf2 protein translation. Mol Cell Proteomics 2012; 11:M111 - , 015032; http://dx.doi.org/10.1074/mcp.M111.015032; PMID: 22207702
- Bowden ET, Coopman PJ, Mueller SC. Invadopodia: unique methods for measurement of extracellular matrix degradation in vitro. Methods Cell Biol 2001; 63:613 - 27; http://dx.doi.org/10.1016/S0091-679X(01)63033-4; PMID: 11060862