Abstract
Polysialic acid (PSA) is a large carbohydrate added post-translationally to the extracellular domain of the Neural Cell Adhesion Molecule (NCAM) that influences its adhesive and other functional properties. PSA-NCAM is widely distributed in the developing nervous system where it promotes dynamic cell interactions, like those responsible for axonal growth, terminal sprouting and target innervation. Its expression becomes restricted in the adult nervous system where it is thought to contribute to various forms of neuronal and glial plasticity. We here review evidence, obtained mainly from hypothalamic neuroendocrine centers and the olfactory system, that it intervenes in structural synaptic plasticity and accompanying neuronal-glial transformations, making possible the formation and elimination of synapses that occur under particular physiological conditions.
Introduction
Although stable neurochemical and functional specificities are fundamental features of neuronal connectivity in the adult mammalian central nervous system (CNS), another important attribute is the ability to undergo function-related structural changes. Structural transformations can affect the overall morphology and therefore, interrelationships of CNS cells, both neurons and glia. They can also affect synapses, the fundamental elements of neuronal connectivity. While the concept of ‘synaptic plasticity’ is usually associated with functional modifications in preexisting synapses (like long-term potentiation or LTP), it connotes structural changes as well, including formation and elimination of synapses that can take place during normal conditions of changing neuronal activity (reviewed in refs. Citation1–Citation3). Moreover, to the classic view of synapses composed of neuronal pre- and post-synaptic elements, we must now include a third player, glia (reviewed in ref. Citation4). CNS synapses are generally encapsulated by fine distal processes of astrocytes that actively contribute to the regulation of synaptic function.Citation4 This intimate neuronal-glial association is not stable but is modifiable in several brain regions, which adds further complexity to neurotransmission and neuronal communication (reviewed in ref. Citation5). Such morphological transformations are induced by physiological conditions of heightened neuronal activity and highlight the nervous system's remarkable capacity for restructuring to meet particular functional conditions.
In the CNS, architectural specificity is attained during development and maintained in the adult through the action of a cohort of membrane-bound and extracellular matrix molecules, particularly adhesion-type glycoproteins with permissive and/or instructive functions. They include cell adhesion molecules like the cadherins, ephrins, neurexins and members of the immunoglobulin superfamily, such as the Neural Cell Adhesion Molecule (NCAM), as well as their receptors. Complex glycoproteins secreted from astrocytes into the extracellular space, like the chondroitin sulphate proteoglycans and tenascins, also contribute.Citation6 These molecules play critical roles in the establishment, maintenance and functional modulation of neuronal circuitries. During synapse formation, they intervene in axonal growth, pathfinding and target recognition and in the differentiation of pre- and post-synaptic specializations; they play a role in the regulation of synaptic size, stability, strength and plasticity as well. It is not unexpected then that such molecules play significant roles in the capacity of synapses for restructuring. The topic has gained increasing attention and there are several excellent reviews summarising recent findings.Citation7–Citation10
We here focus on NCAM, and in particular, its highly sialylated isoforms, in activity-dependent changes of synaptic connections in the adult. As we shall discuss, we now have considerable evidence showing that polysialic acid (PSA), a large, highly complex sugar on the extracellular domain of NCAM, intervenes actively in the dynamic cell interactions responsible for synaptic remodeling. While mechanisms of action are still a matter of speculation, numerous observations from different neuronal systems make it obvious that PSA-NCAM is a necessary molecular participant in activity-dependent structural synaptic plasticity in the adult CNS.
PSA-NCAM, its Structure and Possible Mode of Action
Structure.
Polysialic acid (PSA) is a linear homopolymer of α 2-8-N acetylneuraminic acid whose major carrier in vertebrates is NCAM (reviewed in refs. Citation11–Citation14). NCAM is a member of the immunoglobulin superfamily of adhesion molecules coded by a single copy gene from which at least 20–30 distinct forms can be generated by alternative splicing and post-translational modifications (reviewed in ref. Citation15). There are three major classes of polypeptides, differing in their size (NCAM 180, 140 and 120), cytoplasmic domains (NCAM 180 and 140) or in their means of attachment to the cell membrane (NCAM 120).
Polysialic acids are polymers of derivatives of nine carbon neuraminic acid which can be found in a wide range of biological components, from the coat of certain bacteria to the membranes of vertebrate neural cells. PSA is a relatively simple and large molecule, the number of monomers ranging from about 8 to over 100 (reviewed in refs. Citation11–Citation13) Polysialylation can account for up to 30% of relative molecular mass. The carbohydrate is added posttranslationally through two Golgi-associated polysialyltransferases: ST8SiaIV (PST) and ST8SiaII (STX) (reviewed in ref. Citation16).
Most of the interest linked to NCAM was prompted by the discovery that its functional properties are strongly influenced by polysialylation (reviewed in ref. Citation14). Indeed, PSA can be added to all the three forms of NCAM by post-translational modification, thus introducing new possibilities of modulating adhesion.
Mode of action.
PSA serves as a potent negative regulator of cell interactions via its unusual biophysical properties (reviewed in refs. Citation12 and Citation14), although how it intervenes to permit restructuring in the CNS remains basically undetermined.
The classical view of PSA action refers to its ability to decrease NCAM-mediated membrane-membrane adhesion through steric properties resulting from a high density of negative charges which contribute to the hydrated volume of NCAM.Citation17 The complex mechanism(s) allowing PSA to modulate NCAM-mediated cis, trans, homophilic and heterophilic interactions involve the charge and hydration of the polymer, its influence on the width of the extracellular space as well as its influence on receptors located on the same cell, other cells and/or the extracellular substrate. That there can be different degrees of polysialylation on different NCAM isoforms varying in location and time renders identification of specific mechanisms of action particularly difficult (reviewed in refs. Citation18–Citation20).
Many effects involving the attenuation of cell-cell interactions are not directly mediated through the adhesion properties of NCAM, but rather, are linked to a global effect due to steric inhibition of membrane-membrane apposition based solely on PSA. The importance of such an effect resides in the simple fact that cell-cell interactions are more influenced by the rate of receptor encounter (e.g., IgCAM, L1, cadherins, integrins) than by the intrinsic affinities of the receptors themselves.Citation17,Citation21,Citation22 In other words, PSA decreases the rate of binding among receptors on apposing cells, without affecting the intrinsic binding properties of those receptors. Alternatively, PSA can increase the magnitude of repulsive pressure between membranes sufficient to abrogate the attractive forces exerted by proteins like the cadherins, integrins and NCAM itself.Citation22
The expression of PSA on NCAM is regulated by a combination of developmental or physiological programs that control its synthesis, delivery and degradation.Citation22 The amount of PSA on the cell surface appears to depend on two major regulatory mechanisms: (1) the synthesis of PSA-NCAM on the basis of transcription and/or activity of the sialyltransferases, and (2) the turnover of the molecule at the cell surface (reviewed in refs. Citation12, Citation14 and Citation20). In many systems, biosynthesis of PSA-NCAM is regulated by neuronal activation,Citation23 including electrical activity in axons, which can also influence its expression in the target cell,Citation24 or the loss of input from the periphery.Citation25 Nevertheless, there is also strong evidence that in some cells, including glia and neuroendocrine neurons, PSA-NCAM arrives at the surface in a rapid, constitutive fashion that is not dependent on calcium entry or cell activation.Citation26–Citation29
PSA-NCAM is generally viewed as a regulator of the extracellular space. The hydrated volume of NCAM together with PSA has been estimated to be about three times that of the polypeptide, thus directly increasing the width of the extracellular space and the closeness of membrane-membrane contact.Citation30 The extracellular space is comparatively wide in embryonic tissues which are highly hydrated, whereas it becomes progressively reduced with postnatal maturation.Citation31 The fact that there are high amounts of PSA-NCAM in sites of persistent plasticity in the adult CNS with a specific time-course during postnatal brain maturationCitation32 suggests that polysialylation may provide a suitable extracellular environment in the highly static, mature nervous tissue (reviewed in ref. Citation33).
Regulation.
The fact that PSA can be rapidly expressed at the cell surface,Citation23,Citation27,Citation29 along with the demonstration of relatively fast changes in the morphology and contacts of cells expressing high amounts of PSA-NCAM,Citation34 raises the possibility that polysialylation may be involved in rapid modulation of cell surface interactions. Rapid conformational changes can affect neurons and glia, either when both cell types express PSA, like in the hypothalamic magnocellular nuclei,Citation26,Citation27 or when it is expressed only by the neuronal compartment, such as migrating neuroblasts in the process of adult neurogenesis.Citation35–Citation37
Apart from specific molecular mechanisms, “the coupling of changes in PSA expression with the relevant biological process is the key to the creation of permissive conditions at the appropriate time and place.”Citation18 This concept is supported by observations from PSA-negative but NCAM-positive mice obtained by simultaneous deletion of the polysialyltransferases PST and STX,Citation38,Citation39 observations that highlight the interplay between NCAM and PSA in regulating NCAM interactions and neuronal differentiation. These animals display a severe phenotype characterized by postnatal growth and brain defects beyond features described in NCAM-null mice. Since these defects are rescued by additional deletion of NCAM, it is possible that they originate from a gain of NCAM functions due to the deficiency in PSA. Accordingly, differentiation can be induced by enabling heterophilic NCAM signals at cell-cell contacts that are otherwise prevented by polysialylationCitation38 and dynamic changes in PSA can modulate the cell response by affecting the integration of homophilic and heterophilic NCAM interactions.Citation40
Through these different mechanisms, PSA-NCAM can play a prominent role in various forms of physiologically-occurring structural plasticity in the adult CNS, which include changes in the shape of neurons and associated glia and therefore in their interrelationships, in the length and arborization patters of axons and dendrites, in the formation and elimination of synapses, in activity-induced plasticity, and in adult neurogenesis.
PSA-NCAM in Synaptic Plasticity
Synaptic plasticity and its modulation through adhesion molecules appeared very early in evolution and both processes are functionally present in simple nervous systems. For example, a group of cell surface proteins designated Aplysia cell adhesion molecules (apCAM's) and fasciclin II, which are homologues of vertebrate NCAM in the marine mollusk Aplysia and in Drosophila, respectively, are required for morphological rearrangement of synapses.Citation41,Citation42 Polysialylation, from the evolutionary point of view appears more important in vertebrates since invertebrate NCAM homologues are not polysialylated and the attenuation of cell interactions can occur through downregulation of adhesion receptor distribution on the cell surfaceCitation43 or by expression of other anti-adhesive proteins.Citation44 The appearence of PSA in complex nervous systems may thus represent a mechanism allowing more global effects on cell interactions with respect to an increase in complexity due to the regulation of many different genes (reviewed in ref. Citation18).
During maturation of neuronal systems throughout the postnatal period, the expression of PSA-NCAM becomes progressively confined to pre- and post-synaptic sites.Citation45–Citation47 Indeed, as shown by many observations from different regions, polysialylation is critical for establishing proper synaptic contacts. For instance, experimental PSA removal either by genetic manipulation or enzymatic digestion can induce ectopic synapse formation (as in the hippocampusCitation48) or may alter the very formation of synapses (as between chick oculomotor axons and ciliary neuronsCitation49). More recently, it was seen that developmental and activity-dependent regulation of PSA plays a major role in the maturation of certain GABAergic innervations in the visual cortex since premature removal of PSA resulted in precocious innervation and enhanced inhibitory synaptic transmission.Citation50 By the third postnatal week in rodents, most synapses lose PSA-NCAM, a change that is paralleled by important modifications in PSA expression related to other types of plasticity, such as persistent neurogenesis (reviewed in ref. Citation33 and ).
There is now a considerable literature showing that polysialylation plays an essential role in functional synaptic plasticity in the adult. The most studied and perhaps most complex model of this kind of neuronal plasticity is the hippocampus, where it is clear that long-lasting changes at excitatory synapses like long-term potentiation (LTP) and long-term depression (LTD) are dependent on PSA expression.Citation51–Citation54 Selective PSA removal prevents induction of both LTP and LTD in cultured hippocampal neurons, and in slices prepared from NCAM knock-out mice.Citation52 A more recent study, in which the role of PSA and NCAM at synapses formed by entorhinal cortex axons in the dentate gyrus were analysed in vivo in NCAM-, STX- and PST-deficient mice,Citation55 revealed further complexity, suggesting that PSA may be important for basal synaptic transmission in newly generated granule cells, whereas LTP in the dentate gyrus depends on NCAM glycoprotein alone.
The phenomenon of structural synaptic plasticity in the hippocampus appears more complex since there is an ongoing remodelingCitation56–Citation58 together with that linked to neurogenesis as a result of proliferative/differentiative activity of stem/progenitor cells.Citation59,Citation60 Since this issue has been widely addressed in several recent reviews (reviewed in refs. Citation14 , Citation33 and Citation61) it shall not be discussed further here. Rather, in the following sections we will address the issue of PSA and synaptic remodeling in two other contexts in which they are highly correlated (1) in certain hypothalamic neuroendocrine centers, where PSA expression, synaptic rewiring and concomitant astrocytic changes are functionally linked to specific physiological states and (2) in the olfactory system and olfactory pathways, where polysialylation is associated with a wide range of structural plastic events, among which synaptic plasticity and indirectly, learning and memory.
PSA-NCAM is Necessary for Axonal and Synaptic Remodeling in Neuroendocrine Systems
The strongest evidence showing that PSA is essential for axon and synapse remodeling has been obtained from hypothalamic neuroendocrine structures, like the supraoptic (SON) and paraventricular (PVN) nuclei of the hypothalamo-neurohypophysial system (HNS), the arcuate nuclei and the median eminence of several species (reviewed in refs. Citation3 , Citation5 , Citation62 and Citation63). It is now clear that in these particular hypothalamic nuclei, certain circuits controlling the activity of the neuroendocrine neurons remain relatively plastic throughout life, a plasticity coincident with restructuring of adjacent astrocytes and dependent on neuronal activity. In neurohemal projection sites of these neurons, like the neurohypophysis and the external area of the median eminence, there are concomitant changes in the number and morphology of neurosecretory axons and terminals as well as remodeling of associated glia. All these brain areas express high levels of PSA.Citation26,Citation64–Citation68
The SON and magnocellular portions of the PVN are composed of magnocellular neurons secreting the vital neurohormones, oxytocin and vasopressin. In these nuclei, there is a striking synapse turnover in relation to specific physiological conditions leading to enhanced oxytocin secretion, like parturition, lactation and chronic dehydration (reviewed in refs. Citation3 and Citation62). An obvious manifestation is the appearance of boutons making synaptic contact onto more than one postsynaptic elements simultaneously (‘multiple’ synapses) but careful synaptic density analyses reveal that synapse formation affects boutons making single synaptic contacts as well. Analyses of ultrathin sections in which pre- and post-synaptic elements were immunoidentified allowed to show that the circuits undergoing plasticity are inhibitory (GABAergic) and excitatory (glutamatergic and noradrenergic) impinging on oxytocin neurons. The most important increases affect GABAergic inputs so that in the SON of lactating rats, about 50% of all axo-somatic and axo-dendritic synapses are GABAergic, compared to 35% in the SON of virgin rats under basal conditions. Once stimulation is over, synaptic densities revert to control values. These synaptic transformations are clearly associated with astrocytic changes since an enhanced number of synapses is invariably associated with a reduction in astrocytic coverage of oxytocinergic somata and dendrites; the return to control synaptic levels occurs together with a reestablishment of glial coverage. As seen in more recent work, synaptic remodeling in this system can be reproduced in vitro, in acute slices from adult hypothalamus that include the SON.Citation34,Citation69 Electron microscopy and patch clamp electrophysiology of these preparations allowed to show unequivocably that synapse formation is very rapid, occurring within an hour, is reversible within hours as well and results in the formation of functional, inhibitory synapses.
Another hypothalamic center in which there are synaptic changes clearly linked to varying conditions of physiological conditions is the arcuate nucleus. In adult females, there is a natural phasic synaptic turnover that occurs in relation to the ovarian cycle.Citation63 Thus, the number of axo-somatic GABAergic synapses on arcuate neurons falls significantly between the morning and afternoon of proestrus, remains low throughout estrus, and rises to baseline levels on metestrus, to fall again at proestrus. Concomitantly, like in the magnocellular nuclei,Citation3 there is increased and decreased astrocytic ensheathment of neuronal profiles.Citation70 This negative correlation between the level of glial ensheathment and number of synaptic inputs in relation to fluctuations in steroid levels has been detected in the arcuate nucleus of rodents and primates. In sheep, seasonal variations in glial ensheathment and synaptic inputs occur in the preoptic area. Thus, the number of synapses decreases while astrocytic coverage of neurons increases between the breeding and non-breeding seasons in conjunction with the diminished pulsatile GnRH secretion that leads to anestrous.Citation64
In the neurohemal structures containing the axons of these neurons, glial transformations give rise to a reduced glial ensheathment of neurosecretory terminals which, in perivascular areas, leads to a significantly enlarged neurovascular contact zone (reviewed in ref. Citation5). For example, in the rat neurohypophysis under basal conditions of neurosecretion, about 40% of the perivascular basal lamina is contacted by neurosecretory terminals and about 60% by processes of astrocytic-like pituicytes. During all conditions that stimulate neurosecretion, there is retraction of pituicyte processes from the perivascular zone as well as proliferation of neurosecretory terminals and these proportions are reversed.Citation71 At the end of stimulation, glial coverage of perivascular areas returns to control levels. The axons of arcuate GnRH neurons project to the external layer of the median eminence where fluctuating steroid levels lead to axonal-glial changes similar to those characterising the neurohypophysis. They result in retraction of end feet of tanycytes (modified ependymoglial cells) from the perivascular zone and exposure of GnRH terminals to fenestrated portal capillaries.Citation72
As noted earlier (Section 2), neurons and glia in these structures constitutively express high levels of PSA-NCAM (), an expression that is not markedly affected by changing conditions of neurosecretion.Citation26,Citation27,Citation66,Citation68 This does not mean that PSA is of no consequence to their ability to undergo activity-dependent axonal and synaptic remodeling. On the contrary, it is a prerequisite for all dynamic phases of such plasticity. Thus, specific enzymatic removal of PSA from NCAM in the SON in situ inhibited glial and neuronal remodeling associated with lactation and chronic dehydration; there was no effect if PSA was removed once morphological changes had already taken place.Citation26 Likewise, enzymatic removal of PSA from cell surfaces in the neurohypophysis prevented stimulation-related induction and reversal of axon and glial changes but had no effect once remodeling had occurred.Citation71 In the arcuate nucleus,Citation73 enzymatic removal of PSA inhibited the onset of synaptic changes associated with reproductive status. It is noteworthy that such morphofunctional changes, at least in the magnocellular nuclei, modify the activity of both polysialyltransferases responsible for polysialylation.Citation74
A molecule like NCAM can be considered permissive for morphological plasticity in the neuroendocrine centers since its expression is not tightly linked to neuronal activity yet it appears obligatory for their morphological plasticity. As discussed above, this was clearly evident from enzyme perturbation experiments in which PSA was punctually removed by endosialidase application at particular stages of physiological function. Nonetheless, there can be compensatory replacement by other molecules since remodeling is visible in lactating and salt loaded transgenic mice genetically deficient in NCAM.Citation75 It is highly likely that these observations reveal a redundancy in the molecular systems permitting morphological plasticity. Similar phenomena have been described in other neuronal systems in which NCAM is highly expressed and which continue to undergo plasticity in NCAM-/- mutants.Citation19 In a sense, the specific identity of molecules permissive for remodeling may not be important provided that they activate the same intracellular mechanism. The response of neurons and astrocytes implicated in remodeling would be invariant and independent of the identity of the factor, provided the proper stimulus intervenes.
PSA-NCAM in the Olfactory System and Related Pathways: An Indirect Involvement in Synapse Remodeling
Formation/elimination/remodeling of synapses are decisive for memory storage and for learning-induced changes in the wiring diagram of many neuronal circuitries (reviewed in refs. Citation2, Citation57 and Citation76 ). It has been recognised that PSA-NCAM plays a prominent role in this context, a role which nonetheless remains poorly understood.
The olfactory pathway, in which immunocytochemical evidence for polysialylation was first obtained,Citation77 is a striking illustration of several aspects of structural neuronal plasticity, including synaptic remodeling at different levels of the olfactory pathway, from olfactory receptors located in the nose to cortical circuitries, axonal/dendritic outgrowth towards and within the olfactory bulb, and adult neurogenesis both at peripheral and central levels (reviewed in ref. Citation78). PSA-NCAM is associated with most anatomical/cellular elements of this system, including the olfactory sheet and olfactory nerve,Citation77 the main and accessory olfactory bulb,Citation66,Citation77,Citation79,Citation80 the rostral migratory stream originating from the forebrain subventricular zone (SVZ),Citation35,Citation36,Citation81 and cortical areas receiving and processing olfactory inputsCitation82–Citation84 ().
PSA-NCAM and plasticity in the olfactory bulb.
A multipotent stem cell generates new olfactory receptor neurons in the olfactory sheet throughout adult life.Citation85 These sensory neurons are physiologically replaced with an average life-span of 30 days.Citation86 Their axons growing towards the olfactory bulb strongly express PSA-NCAM,Citation77 an expression presumably essential for their extension and path-finding.Citation87 The olfactory axons are wrapped by a specialized type of glia, the ensheathing glia, which enable them to enter the mature CNSCitation88 and they also express PSA-NCAM.Citation77,Citation89
How glial cells and PSA are related to continuous synaptogenesis and synaptic plasticity in the olfactory bulb remains an open field of investigation. In addition to continuous axon outgrowth from the olfactory nerve, this region is enriched throughout life by new, migrating cells generated in the forebrain SVZ.Citation33,Citation36,Citation59 Another issue that remains obscure is the role of PSA in synapse formation in the context of adult neurogenesis. Such a process cannot be viewed as a faithful recapitulation of embryonic events, since there are substantial differences in the sites of production and integration of new cells at different stages of CNS maturation.Citation33,Citation90 Newly generated neuroblasts reaching the olfactory bulb cease to express PSA-NCAM two-three weeks after their genesis, as they differentiate into granule and periglomerular interneurons (reviewed in ref. Citation33). Yet, the newly-added interneurons establish functional synapses only after they mature and integrate into circuits of the olfactory bulb.Citation90 Besides a well established role in migration, polysialylation seems to be important in maintaining newly generated cells in an undifferentiated state until they reach their final destination.Citation33,Citation39,Citation40,Citation61 In cells isolated from the SVZ, removal of PSA specifically promotes neurite induction and maturation.Citation91 All these observations indicate that, like for ocular dominance plasticity,Citation50 PSA is essential to the maturation of interneuron synaptic innervation. In the olfactory bulb, downregulation and loss of PSA on the membrane of newly generated cells could be a signal for their differentiation and integration.
A peculiar extracellular environment, provided by the diffuse expression of molecules like PSA-NCAM and extracellular matrix tenascins,Citation92 could render the region more permissive and would not require specific polysialylation of the newlyborn cells themselves during the phase of their integration. In this context, it is noteworthy that a strong diffuse expression of PSA-NCAM is associated with synaptic remodeling in the hypothalamic neuroendocrine centersCitation26,Citation64,Citation68,Citation73 and in the superficial laminae of the spinal cord.Citation93 In other words, the role of PSA in brain regions maintaining a ‘juvenile’ environment for adult neurogenesis could be different from that of cells expressing PSA in more ‘static’ areas (see below).
Like in the hippocampus, because of the overlapping of functiona/structural synaptic plasticity and persistent neurogenesis, the role of adhesion molecules and synaptic plasticity in the olfactory bulb continues to be a widely open field of investigation.
Is PSA-NCAM important for learning/memory in the olfactory pathway?
Experimental manipulations affecting the expression or the function of PSA-NCAM alter the ability of animals to learn.Citation19,Citation50,Citation83,Citation94 Conversely, learning alters the expression of the molecule (reviewed in refs. Citation95 and Citation96). By examining PSA-NCAM distribution (), it becomes obvious that polysialylation characterizes complex, widely interconnected olfactory neural circuits and pathways: (1) the olfactory bulb and the primary olfactory cortex, consisting of three-layered cortical structures (allocortical areas and transitional allo/iso-cortical areas) and (2) the medial temporal lobe (including the hippocampal formation, the perirhinal and parahippocampal—postrhinal—cortices), a bidirectional pathway between cortical regions and the hippocampal formation.Citation97,Citation98 Most of these neuronal circuits represent multiple levels of the same pathway (primary sensory, projection and associative) and are involved in memory storage and learning.Citation78,Citation99 Their distribution provides correlative evidence for an involvement of polysialylation in synaptic remodeling.Citation100
The hippocampus can be considered an extension of the olfactory pathway, both structurally, due to its connections with the primary olfactory cortex and functionally, since selective types of olfactory memory depend on the hippocampal formation (reviewed in ref. Citation78). Polysialylation is important at this level as well as in the olfactory system proper. For example, changes in NCAM and PSA-NCAM expression occur in the piriform cortex and hippocampus in relation to learning of olfactory discrimination tasks,Citation100 and transient polysialylation in the dentate gyrus is linked to memory for odor-reward association.Citation101
Structures of the medial temporal lobe are involved in memory consolidation, a process in which memories are initially dependent on the medial temporal lobe but gradually become established as long-term memories in other brain areas.Citation102 Also, spatial and contextual memory/learning depend on corticohippocampal circuitry.Citation102,Citation103 In most cases, there is PSA-mediated plasticity. In the hippocampus, transient modulations of polysialylation follow spatial learning and memoryCitation50,Citation104 while enzymatic removal of PSA affects spatial learning.Citation50 A transient increase in the frequency of polysialylated cells in the adult hippocampus,Citation95 septo-hippocampal pathwayCitation105 and entorhinal cortexCitation83 is detectable during memory consolidation. These changes occur early (between 6 and 12 hours) and return to basal levels within 72 hours following periods of consolidation related to the selective retention and/or elimination of spines and polysialylation (reviewed in ref. Citation106). Also consolidation of passive avoidance learning is associated with transient increases of polysialylated neurons in layer II of the entorhinal/perirhinal cortex.Citation83,Citation84 Finally, recent studies provided evidence that PSA regulates synaptic plasticity at synapses formed by entorhinal cortex axons in the dentate gyrus in vivo,Citation55 thereby extending previous in vitro results concerning the role of NCAM in LTP and presumably learning and memory.Citation51,Citation107,Citation108
In conclusion, there is now much evidence to support the involvement of PSA in several aspects of brain cognitive functions, exerting its effects at different levels of widely interconnected neuronal circuitries. Although suggestive of synaptic remodeling, the role of PSA-NCAM in isolated (non-newly generated) cortical neurons intercalated on the olfactory pathways remains a matter of debate. Hence, if at some levels of the olfactory/hippocampal systems PSA-NCAM is unequivocally related to neurogenesis, at other levels, it could be independent, as it is probably the case in most of the temporal lobe cortices.Citation83,Citation84,Citation95,Citation103
PSA-NCAM on immature neurons in the mature brain: a potential reservoir for new synapses?
It is generally thought that in areas of the hard-wired adult brain like the cerebral cortex, the morphology of most neuronal cells and their interconnections become fixed after specific time points or critical periods (reviewed in ref. Citation109). Nevertheless, an increasing number of observations indicate that there are cortical neurons that continue to express molecules involved in structural remodelling, like PSA-NCAM.Citation66,Citation82,Citation83,Citation110 Thus, as expected, newly born cells generated in the context of adult neurogenesis express high levels of the glycoprotein.Citation33,Citation59 However, there are also non-newly generated cells expressing PSA-NCAM, and this could be illustrating their capacity for global morphological remodelling throughout life.Citation111,Citation112 In addition, they typically display a ‘punctate’ immunolabeling of their surface membranesCitation33,Citation110 () and this could represent ongoing synaptic remodeling of their inputs. These neurons receive only sparse synaptic contacts.Citation111 The observations that they are covered by astrocytic lamellae along with swellings of the extracellular spaces suggest that such polysialylated cortical neurons may be reservoirs of cells that have not completed their differentiation programme and are still available for new synaptic contacts.Citation111,Citation112
Examples such as the above provide a model in which polysialylation and dynamic processes of astrocytes may provide a space suitable for new synapse formation on a group of neurons within the static environment of the mature nervous tissue (reviewed in ref. Citation5). As we have described in this review, high levels of PSA are expressed throughout life by particular groups of neurons and their associated astrocytes (as in the hypothalamic magnocellular nuclei) as well as by isolated neuronal cells with some participation of glia (as in the olfactory bulb and cortex). Ongoing studies aim to unravel the function of the latter cells as well as the mechanisms that allow their persistent potential for plasticity.
Conclusions
As seen from the numerous observations described in this review, the presence of large amounts of PSA on the extracellular surface of neurons and astrocytes of the adult CNS appears critical for the morphological transformations linked to synapse formation and elimination occuring during diverse physiological conditions. The complex carbohydrate may provide particular extracellular space conditions favorable for such remodeling, for example, by greatly reducing adhesivity between adjacent cells and their synapses and/or the cells and their immediate extracellular matrix. In addition, PSA and/or its carrier NCAM may intervene more actively to allow interaction with other molecules on the cell surface of neurons and/or astrocytes, including other cell adhesion molecules, growth factors and receptors.Citation20,Citation52,Citation113–Citation115 These interactions may be critical for synapse remodeling since they will then trigger intracellular signaling mechanisms that ultimately result in gene activation and new protein synthesis underlying the morphological transformations. Analysis of mechanisms of PSA action as well as the identification of molecular factors with which it may intervene to permit structural synaptic plasticity is therefore of utmost relevance and remains a challenging task for future research.
Figures and Tables
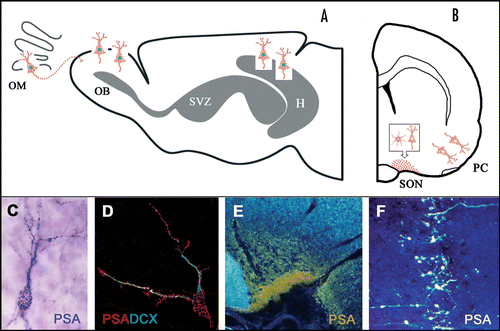
Figure 1 Highly sialylated Neural Cell Adhesion Molecule (PSA-NCAM) in adult brain cell populations involved in different types of synaptic plasticity. Top: diagrammatic representation of sagittal (A) and coronal (B) views of the rat brain showing areas containing cells enriched in PSA-NCAM. In grey, neurogenic sites. H, hippocampus; OB, olfactory bulb; OM, olfactory mucosa; PC, piriform cortex; SON, supraoptic nucleus; SVZ, subventricular zone. Bottom: Immunoreactions for polysialic acid (PSA) reveal highly immunopositive neurons (C, D and F) in restricted areas known to undergo plasticity, like the olfactory bulb (C), hippocampus (D) and the piriform cortex (F). In neuroendocrine centers like the SON (E), a strong reaction fills the neuropile and is due to immunoreaction on neuronal and glial (astrocytic) processes. Note that newly generated (top, green nuclei) and non-newly generated cells (top, unstained nuclei) display a typical punctate reaction on their cell membrane (see also C and D). DCX, doublecortin.
Acknowledgements
This work was partially supported by grants from the Compagnia di San Paolo (Progetto NEUROTRANSPLANT 2004.2019 and 1553 IT/CV), Regione Piemonte (CIPE 2004-A14), M.U.R.S.T. (PRIN), University of Turin (to L.B.) and Inserm (to D.T.T.).
References
- Geinisman Y. Structural synaptic modifications associated with hippocampal LTP and behavioral learning. Cereb Cortex 2000; 10:952 - 962
- Chklovskii DB, Mel BW, Svoboda K. Cortical rewiring and information storage. Nature 2004; 431:782 - 788
- Theodosis DT, Trailin A, Poulain DA. Remodeling of astrocytes, a prerequisite for synapse turnover in the adult brain? Insights from the oxytocin system of the hypothalamus. Am J Physiol Regul Integr Comp Physiol 2006; 290:1175 - 1182
- Araque A, Parpura V, Sanzgiri RP, Haydon PG. Tripartite synapses: glia, the unacknowledged partner. Tr Neurosci 1999; 22:208 - 215
- Theodosis DT, Poulain DA, Oliet SHR. Activity-dependent structural and functional plasticity of astrocyte-neuron interactions. Physiol Rev 2008; 88:983 - 1008
- Dityatev A, Schachner M. The extracellular matrix and synapses. Cell Tissue Res 2006; 326:647 - 654
- Yamagata M, Sanes JR, Weiner JA. Synaptic adhesion molecules. Curr Opin Cell Biol 2003; 15:621 - 632
- Kleene R, Schachner M. Glycans and neural cell interactions. Nature Rev Neurosci 2004; 5:195 - 208
- Dalva MB, McClelland AC, Kayser MS. Cell adhesion molecules: signalling functions at the synapse. Nat Rev Neurosci 2007; 8:206 - 220
- McAllister AK. Dynamic aspects of CNS synapse formation. Annu Rev Neurosci 2007; 30:425 - 450
- Rougon G. Structure, metabolism and cell biology of polysialic acid. Eur J Cell Biol 1993; 61:197 - 207
- Rougon G, Hobert O. New insights into the diversity and function of neuronal immunoglobulin superfamily molecules. Annu Rev Neurosci 2003; 26:207 - 238
- Muhlenhoff M, Eckhardt M, Gerardy-Schahn R. Polysialic acid: three-dimensional structure, biosynthesis and function. Curr Opin Struct Biol 1998; 8:558 - 564
- Rutishauser U. Polysialic acid in the plasticity of the developing and adult vertebrate nervous system. Nat Rev Neurosci 2008; 9:26 - 35
- Edelman GM, Crossin KL. Cell adhesion molecules: implications for a molecular histology. Annu Rev Biochem 1991; 60:155 - 190
- Angata K, Fukuda M. Polysialyltransferases: major players in polysialic acid synthesis on the neural cell adhesion molecule. Biochimie 2003; 85:195 - 206
- Yang P, Major D, Rutishauser U. Role of charge and hydration in effects of polysialic acid on molecular interactions on and between cell membranes. J Biol Chem 1994; 269:23039 - 23044
- Bruses JL, Rutishauser U. Roles, regulation and mechanism of polysialic acid function during neural development. Biochimie 2001; 83:635 - 643
- Durbec P, Cremer H. Revisiting the function of PSA-NCAM in the nervous system. Molecular Neurobiology 2001; 24:53 - 64
- Kiss JZ, Muller D. Contribution of neural cell adhesion molecule to neuronal and synaptic plasticity. Rev Neurosci 2001; 12:297 - 310
- Fujimoto I, Bruses JL, Rutishauser U. Regulation of cell adhesion by polysialic acid. Effects on cadherin, immunoglobulin cell adhesion molecule, and integrin function and independence from neural cell adhesion molecule binding or signaling activity. J Biol Chem 2001; 276:31745 - 31751
- Johnson CP, Fujimoto I, Rutishauser U, Leckband DE. Direct evidence that neural cell adhesion molecule (NCAM) polysialylation increases intermembrane repulsion and abrogates adhesion. J Biol Chem 2005; 280:137 - 145
- Kiss JZ, Wang C, Olive S, Rougon G, Lang J, Baetens D, et al. Activity-dependent mobilization of the adhesion molecule polysialic NCAM to the cell surface of neurons and endocrine cells. EMBO J 1994; 13:5284 - 5292
- Fredette B, Rutishauser U, Landmesser L. Regulation and activity-dependence of N-cadherin, NCAM isoforms and polysialic acid on chick myotubes during development. J Cell Biol 1993; 123:1867 - 1888
- Bonfanti L, Merighi A, Theodosis DT. Dorsal rhizotomy induces transient expression of the highly sialylated isoform of the neural cell adhesion molecule in neurons and astrocytes of the adult spinal cord. Neuroscience 1996; 74:619 - 623
- Theodosis DT, Bonhomme R, Vitiello S, Rougon G, Poulain DA. Cell surface expression of polysialic acid on NCAM is a prerequisite for activity-dependent morphological neuronal and glial plasticity. J Neurosci 1999; 19:10228 - 10236
- Pierre K, Bonhomme R, Dupouy B, Poulain DA, Theodosis DT. The polysialylated Neural Cell Adhesion Molecule (PSA-NCAM) reaches cell surfaces of hypothalamic neurons and astrocytes via the constitutive pathway. Neuroscience 2001; 103:133 - 142
- Alcaraz G, Goridis C. Biosynthesis and processing of polysialylated NCAM by AtT-20 cells. Eur J Cell Biol 1991; 55:165 - 173
- Scheidegger P, Papay J, Zuber C, Lackie PM, Roth J. Cellular site of synthesis and dynamics of cell surface re-expression of polysialic acid of the neural cell adhesion molecule. Eur J Biochem 1994; 225:1097 - 1103
- Yang P, Yin X, Rutishauser U. Intercellular space is affected by the polysialic acid content of N-CAM. J Cell Biol 1992; 116:1487 - 1496
- Bondareff W, Narotzky R. Age changes in the neuronal microenvironment. Science 1972; 176:1135 - 1136
- Peretto P, Giachino C, Aimar P, Fasolo A, Bonfanti L. Chain formation and glial tube assembly in the shift from neonatal to adult subventricular zone of the rodent forebrain. J Comp Neurol 2005; 487:407 - 427
- Bonfanti L. PSA-NCAM in mammalian structural plasticity and neurogenesis. Prog Neurobiol 2006; 80:129 - 164
- Langle SL, Poulain DA, Theodosis DT. Induction of rapid, activity-dependent neuronalglial remodeling in the adult hypothalamus in vitro. Eur J Neurosci 2003; 18:206 - 214
- Bonfanti L, Theodosis DT. Expression of polysialylated neural cell adhesion molecule by proliferating cells in the subependymal layer of the adult rat, in its rostral extension and in the olfactory bulb. Neuroscience 1994; 62:291 - 305
- Rousselot P, Lois C, Alvarez-Buylla A. Embryonic (psa) N-CAM reveals chains of migrating neuroblasts between the lateral ventricle and the olfactory bulb of adult mice. J Comp Neurol 1995; 351:51 - 61
- Doetsch F, Garcia-Verdugo JM, Alvarez-Buylla A. Cellular composition and three-dimensional organization of the subventricular germinal zone in the adult mammalian brain. J Neurosci 1997; 17:5041 - 5046
- Weinhold B, Seidenfaden R, Rockle I, Muhlenhoff M, Schertzinger F, Conzelmann S, et al. Genetic ablation of polysialic acid causes severe developmental defects rescued by deletion of the neural cell adhesion molecule. J Biol Chem 2005; 280:42971 - 42977
- Seidenfaden R, Krauter A, Schertzinger F, Gerardy-Schahn R, Hildebrandt H. Polysialic acid directs tumor cell growth by controlling heterophilic neural cell adhesion molecule interactions. Mol Cell Biol 2003; 23:5908 - 5918
- Seidenfaden R, Desoeuvre A, Bosio A, Virard I, Cremer H. Glial conversion of SVZ-derived committed neuronal precursors after ectopic grafting into the adult brain. Mol Cell Neurosci 2006; 32:187 - 198
- Mayford M, Barzilai A, Keller F, Schacher S, Kandel ER. Modulation of an NCAM-related adhesion molecule with long-term synaptic plasticity in Aplysia. Science 1992; 256:638 - 644
- Schuster CM, Davis GW, Fetter RD, Goodman CS. Genetic dissection of structural and functional components of synaptic plasticity I. Fasciclin II controls synaptic stabilization and growth. Neuron 1996; 17:641 - 654
- Peter N, Aronoff B, Wu F, Schacher S. Decrease in growth cone-neurite fasciculation by sensory or motor cells in vitro accompanies downregulation of Aplysia cell adhesion molecules by neurotransmitters. J Neurosci 1994; 14:1413 - 1421
- Fambrough D, Goodman CS. The Drosophila beaten path gene encodes a novel secreted protein that regulates defasciculation at motor axon choice points. Cell 1996; 87:1049 - 1058
- Szele FG, Dowling JJ, Gonzales C, Theveniau M, Rougon G, Chesselet M-F. Pattern of expression of highly polysialylated neural cell adhesion molecule in the developing and adult rat striatum. Neuroscience 1994; 60:133 - 144
- Uryu K, Butler AK, Chesselet MF. Synaptogenesis and ultrastructural localization of the polysialylated neural cell adhesion molecule in the developing striatum. J Comp Neurol 1999; 405:216 - 232
- Bouzioukh F, Tell F, Rougon G, Jean A. Dual effects of NMDA receptor activation on polysialylated neural cell adhesion molecule expression during brainstem postnatal development. Eur J Neurosci 2001; 14:1194 - 1202
- Seki T, Rutishauser U. Removal of polysialic acid-neural cell adhesion molecule induces aberrant mossy fiber innervation and ectopic synaptogenesis in the hippocampus. J Neurosci 1998; 18:3757 - 3766
- Bruses JL, Chauvet N, Rubio ME, Rutishauser U. Polysialic acid and the formation of oculomotor synapses on chick ciliary neurons. J Comp Neurol 2002; 446:244 - 256
- Di Cristo G, Chattopadhyaya B, Kuhlman SJ, Fu Y, Bélanger MC, Wu CZ, et al. Activity-dependent PSA expression regulates inhibitory maturation and onset of critical period plasticity. Nat Neurosci 2007; 10:1569 - 1577
- Becker CG, Artola A, Gerardy-Schahn R, Becker T, Welzl H, Schachner M. The polysialic acid modification of the neural cell adhesion molecule is involved in spatial learning and hippocampal long-term potentiation. J Neurosci Res 1996; 45:143 - 152
- Muller D, Wang C, Skibo G, Toni M, Cremer H, Calaora V, Rougon G, Kiss JZ. PSA-NCAM is required for activity-induced synaptic plasticity. Neuron 1996; 17:413 - 422
- Dityatev A, Dityateva G, Sytnyk V, Delling M, Toni N, Nikonenko I, et al. Polysialylated neural cell adhesion molecule promotes remodeling and formation of hippocampal synapses. J Neurosci 2004; 24:9372 - 9382
- Eckhardt M, Bukalo O, Chazal G, Wang L, Goridis C, Schachner M, et al. Mice deficient in the polysialyltransferase ST8SiaIV/PST-1 allow discrimination of the roles of neural cell adhesion molecule protein and polysialic acid in neural development and synaptic plasticity. J Neurosci 2000; 20:5234 - 5244
- Stoenica L, Senkov O, Gerardy-Schahn R, Weinhold B, Schachner M, Dityatev A. In vivo synaptic plasticity in the dentate gyrus of mice deficient in the neural cell adhesion molecule NCAM or its polysialic acid. Eur J Neurosci 2006; 23:2255 - 2264
- Diano S, Farr SA, Benoit SC, McNay EW, da Silva I, Horvath B, et al. Ghrelin controls hippocampal spine synapse density and memory performance. Nat Neurosci 2006; 9:381 - 388
- Galimberti I, Gogolla N, Alberi S, Santos AF, Muller D, Caroni P. Long-term rearrangements of hippocampal mossy fiber terminal connectivity in the adult regulated by experience. Neuron 2006; 50:749 - 763
- Muller D, Mendez P, De Roo M, Klauser P, Steen S, Poglia L. Role of NCAM in spine dynamics and synaptogenesis. Neurochem Res 2008; Epub
- Gage FH. Mammalian neural stem cells. Science 2000; 287:1433 - 1438
- Ge S, Sailor KA, Ming GL, Song H. Synaptic integration and plasticity of new neurons in the adult hippocampus. J Physiol 2008; 586:3759 - 3765
- Gascon E, Vutskits L, Kiss JZ. The Role of PSA-NCAM in adult neurogenesis. Neurochem Res 2008; Epub
- Theodosis DT. Oxytocin-secreting neurons: a physiological model of morphological neuronal and glial plasticity in the adult hypothalamus. Front Neuroendocrinol 2002; 23:101 - 135
- Garcia-Segura LM, Chowen JA, Parducz A, Naftolin F. Gonadal hormones as promoters of structural synaptic plasticity: cellular mechanisms. Prog Neurobiol 1994; 44:279 - 307
- Viguié C, Jansen HT, Glass JD, Watanabe M, Billings HJ, Coolen L, et al. Potential for polysialylated form of neural cell adhesion molecule-mediated neuroplasticity within the gonadotropin-releasing hormone neurosecretory system of the ewe. Endocrinology 2001; 142:1317 - 1324
- Theodosis DT, Rougon G, Poulain DA. Retention of embryonic features by an adult neuronal system capable of plasticity: Polysialylated N-CAM in the hypothalamo-neurohypophysial system. Proc Natl Acad Sci USA 1991; 88:5494 - 5498
- Bonfanti L, Olive S, Poulain DA, Theodosis DT. Mapping of the distribution of polysialylated neural cell adhesion molecule throughout the central nervous system of the adult rat: an immunohistochemical study. Neuroscience 1992; 49:419 - 436
- Kiss JZ, Wang C, Rougon G. Nerve-dependent expression of high polysialic acid neural cell adhesion molecule in neurohypophysial astrocytes of adult rats. Neuroscience 1993; 53:213 - 221
- Perera AD, Lagenaur CF, Plant TM. Postnatal expression of polysialic acid-neural cell adhesion molecule (PSA-NCAM) in the hypothalamus of the male rhesus monkey (Macaca mulatta). Endocrinology 1994; 133:2729 - 2735
- Theodosis DT, Koksman JJ, Trailin A, Langle SL, Piet R, Lodder JC, et al. Oxytocin and estrogen promote rapid formation of functional GABA synapses in the adult supraoptic nucleus. Mol Cell Neurosci 2006; 31:785 - 794
- Garcia-Segura LM, Luquin S, Parducz A, Naftolin F. Gonadal hormone regulation of glial fibrillary acidic protein immunoreactivity and glial ultrastructure in the rat neuroendocrine hypothalamus. Glia 1994; 10:59 - 69
- Monlezun S, Ouali S, Poulain DA, Theodosis DT. Polysialic acid is required for active phases of morphological plasticity of neurosecretory axons and their glia. Mol Cell Neurosci 2005; 29:516 - 524
- de Serrano S, Estrella C, Loyens A, Cornea A, Ojeda SR, Beauvillain JC, et al. Vascular endothelial cells promote acute plasticity in ependymoglial cells of the neuroendocrine brain. J Neurosci 2004; 24:10353 - 10363
- Hoyk Z, Parducz A, Theodosis DT. The highly sialylated isoform of the neural cell adhesion molecule is required for estradiol-induced morphological synaptic plasticity in the adult arcuate nucleus. Eur J Neurosci 2001; 13:649 - 656
- Soares S, von Boxberg Y, Ravaille-Veron M, Vincent JD, Nothias F. Morphofunctional plasticity in the adult hypothalamus induces regulation of polysialic acid-neural cell adhesion molecule through changing activity and expression levels of polysialyltransferases. J Neurosci 2000; 20:2551 - 2557
- Theodosis DT, Schachner M, Neumann ID. Oxytocin neuron activation in NCAM-deficient mice: anatomical and functional consequences. Eur J Neurosci 2004; 20:3270 - 3280
- Zuo Y, Lin A, Chang P, Gan WB. Development of long-term dendritic spine stability in diverse regions of cerebral cortex. Neuron 2005; 46:181 - 189
- Miragall F, Kadmon G, Husmann M, Schachner M. Expression of cell adhesion molecules in the olfactory system of the adult mouse: presence of the embryonic form of N-CAM. Dev Biol 1988; 129:516 - 531
- Wilson DA, Best AR, Sullivan RM. Plasticity in the olfactory system: lessons from the neurobiology of memory. Neuroscientist 2004; 10:513 - 524
- Bonfanti L, Peretto P, Merighi A, Fasolo A. Newly-generated cells from the rostral migratory stream in the accessory olfactory bulb of the adult rat. Neuroscience 1997; 81:489 - 502
- Peretto P, Giachino C, Panzica GC, Fasolo A. Sexually dimorphic neurogenesis is topographically matched with the anterior accessory olfactory bulb of the adult rat. Cell Tissue Res 2001; 306:385 - 389
- Ono K, Tomasiewicz H, Magnuson T, Rutishauser U. N-CAM mutation inhibits tangential neuronal migration and is phenocopied by enzymatic removal of polysialic acid. Neuron 1994; 13:595 - 609
- Seki T, Arai Y. Expression of highly polysialylated NCAM in the neocortex and piriform cortex of the developing and the adult rat. Anat Embryol (Berl) 1991; 184:395 - 401
- O'Connell AW, Fox GB, Barry T, Murphy KJ, Fichera G, Foley AG, et al. Spatial learning activates neural cell adhesion molecule polysialylation in a corticohippocampal pathway within the medial temporal lobe. J Neurochem 1997; 68:2538 - 2546
- Fox GB, Fichera G, Barry T, O'Connell AW, Gallagher HC, Murphy KJ, et al. Consolidation of passive avoidance learning is associated with transient increases of polysialylated neurons in layer II of the rat medial temporal cortex. J Neurobiol 2000; 45:135 - 141
- Mackay-Sim A. Bonfanti L. Neurogenesis in the olfactory mucosa: accessible neural stem cells in adult humans. Postnatal and Adult Neurogenesis 2008; Trivandrum, India Research Signpost In Press
- Graziadei PPC, Monti-Graziadei GA. Neurogenesis and neuron regeneration in the olfactory system of mammals I. Morphological aspects of differentiation and structural organization of the olfactory neurons. J Neurocytol 1979; 8:1 - 18
- Aoki K, Nakahara Y, Yamada S, Eto K. Role of polysialic acid on outgrowth of rat olfactory receptor neurons. Mech Dev 1999; 85:103 - 110
- Ramon-Cueto A, Valverde F. Olfactory bulb ensheathing glia: a unique cell type with axonal growth-promoting properties. Glia 1995; 14:163 - 173
- Miragall F, Dermietzel R. Immunocytochemical localization of cell adhesion molecules in the developing and mature olfactory system. Microsc Res Tech 1992; 23:157 - 172
- Grubb MS, Nissant A, Murray K, Lledo PM. Functional maturation of the first synapse in olfaction: development and adult neurogenesis. J Neurosci 2008; 28:2919 - 2932
- Rockle I, Seidenfaden R, Weinhold B, Muhlenhoff M, Gerardy-Schahn R, Hildebrandt H. Polysialic acid controls NCAM-induced differentiation of neuronal precursors into calretinin-positive olfactory bulb interneurons. Dev Neurobiol 2008; 68:1170 - 1184
- Saghatelyan A, de Chevigny A, Schachner M, Lledo PM. Tenascin-R mediates activity-dependent recruitment of neuroblasts in the adult mouse forebrain. Nat Neurosci 2004; 7:347 - 356
- El Maarouf A, Kolesnikov Y, Pasternak G, Rutishauser U. Polysialic acid-induced plasticity reduces neuropathic insult to the central nervous system. Proc Natl Acad Sci USA 2005; 102:11516 - 11520
- Cremer H, Lange R, Christoph A, Plomann M, Vopper G, Roes J, et al. Inactivation of the N-CAM gene in mice results in size reduction of the olfactory bulb and deficits in spatial learning. Nature 1994; 367:455 - 459
- Regan CM, Fox GB. Polysialylation as a regulator of neural plasticity in rodent learning and aging. Neurochem Res 1995; 20:593 - 598
- Murphy KJ, O'Connell AW, Regan CM. Repetitive and transient increases in hippocampal neural cell adhesion molecule polysialylation state following multitrial spatial training. J Neurochem 1996; 67:1268 - 1274
- Burwell RD, Witter MP, Amaral DG. Perirhinal and postrhinal cortices of the rat: a review of the neuroanatomical literature and comparison with findings from the monkey brain. Hippocampus 1995; 5:390 - 408
- Alvarez P, Squire LR. Memory consolidation and the medial temporal lobe: a simple network model. Proc Natl Acad Sci USA 1994; 91:7041 - 7045
- Brennan PA, Keverne EB. Neural mechanisms of mammalian olfactory learning. Prog Neurobiol 1997; 51:457 - 481
- Knafo S, Barkai E, Herrero AI, Libersat F, Sandi C, Venero C. Olfactory learning-related NCAM expression is state, time and location specific and is correlated with individual learning capabilities. Hippocampus 2005; 15:316 - 325
- Foley AG, Hedigan K, Roullet P, Moricard Y, Murphy KJ, Sara SJ, et al. Consolidation of memory for odour-reward association requires transient polysialylation of the neural cell adhesion molecule in the rat hippocampal dentate gyrus. J Neurosci Res 2003; 74:570 - 576
- Burwell RD, Saddoris MP, Bucci DJ, Wiig KA. Corticohippocampal contributions to spatial and contextual learning. J Neurosci 2004; 24:3826 - 3836
- Burwell RD, Bucci DJ, Sanborn MR, Jutras MJ. Perirhinal and postrhinal contributions to remote memory for context. J Neurosci 2004; 24:11023 - 11028
- Venero C, Herrero AI, Touyarot K, Cambon K, Lopez-Fernandez MA, Berezin V, et al. Hippocampal upregulation of NCAM expression and polysialylation plays a key role on spatial memory. Eur J Neurosci 2006; 23:1585 - 1595
- Foley AG, Ronn LC, Murphy KJ, Regan CM. Distribution of polysialylated neural cell adhesion molecule in rat septal nuclei and septohippocampal pathway: transient increase of polysialylated interneurons in the subtriangular septal zone during memory consolidation. J Neurosci Res 2003; 74:807 - 817
- Murphy KJ, Regan CM. Contributions of cell adhesion molecules to altered synaptic weightings during memory consolidation. Neurobiol Learn Mem 1998; 70:73 - 81
- Dityatev A, Dityateva G, Schachner M. Synaptic strength as a function of post-versus presynaptic expression of the neural cell adhesion molecule NCAM. Neuron 2000; 26:207 - 217
- Bukalo O, Fentrop N, Lee AY, Salmen B, Law JW, Wotjak CT, et al. Conditional ablation of the neural cell adhesion molecule reduces precision of spatial learning, long-term potentiation, and depression in the CA1 subfield of mouse hippocampus. J Neurosci 2004; 24:1565 - 1577
- Hensch TK. Critical period plasticity in local cortical circuits. Nat Rev Neurosci 2005; 6:877 - 888
- Nacher J, Alonso-Llosa G, Rosell D, McEwen B. PSA-NCAM expression in the piriform cortex of the adult rat. Modulation by NMDA receptor antagonist administration. Brain Res 2002; 927:111 - 121
- Gomez-Climent MA, Castillo-Gomez E, Varea E, Guirado R, Blasco-Ibanez JM, Crespo C, et al. A population of prenatally generated cells in the rat paleocortex maintains an immature neuronal phenotype into adulthood. Cereb Cortex 2008; 18:2229 - 2240
- Luzzati F, Bonfanti L, Fasolo A, Peretto P. DCX and PSA-NCAM expression identify a cell population preferentially distributed in associative areas of different pallial derivatives and vertebrate species. Cereb Cortex 2008; http://dx.doi.org/10.1093/cercor/bhn145
- Kiss JZ. A role of adhesion molecules in neuroglial plasticity. Mol Cell Endocrinol 1998; 140:89 - 94
- Vutskits L, Djebbara-Hannas Z, Zhang H, Paccaud JP, Durbec P, Rougon G, et al. PSA-NCAM modulates BDNF-dependent survival and differentiation of cortical neurons. Eur J Neurosci 2001; 13:1391 - 1402
- Povlsen GK. The neural cell adhesion molecule and epidermal growth factor receptor: signaling crosstalk. Neurochem Res 2008; http://dx.doi.org/10.1007/s11064-008-9651-7