Abstract
Central nervous system (CNS) development starts from neural stem cells (NSCs) which ultimately give rise to the three major cell types (neurons, oligodendrocytes and astrocytes) of the CNS. NSCs are specified in space- and time-related fashions, becoming spatially heterogeneous and generating a progressively restricted repertoire of cell types. Mammalian NSCs produce different cell types at different time points during development under the influence of multiple signaling pathways. These pathways act in a dynamic web mode to determine the fate of NSCs via modulating the expression and activity of distinct set of transcription factors which in turn trigger the transcription of neural fate-associated genes. This review thus introduces the major signal pathways、transcription factors and their cross-talks and coordinative interactions in NSC fate determination.
Introduction
The development of CNS begins as a sheet of cells made up of primary progenitors known as neuroepithelial cells. By the onset of CNS formation, neuroepithelial cells were thought be replaced by different neural stem cells (NSCs).Citation1 The current evidence suggests that NSCs presence in both embryo and adult. It gradually transform from neuroepithelial cells to radial glial cells and then to astrocyte-like cells.Citation2–Citation4
Mammalian NSCs produce different cell types at different points in development. During this course, NSCs change their morphology and produce different progeny, but also change their gene expression profiles. Different models have been proposed for the genesis of cells within CNS (). First model was built by in vitro work, which showed that NSCs could generate lineage-restricted progenitors, neural restricted progenitors and glial restricted progenitors, respectively.Citation5 This model proposes a homogeneous NSC population within the CNS first generates lineage-restricted progenitors, NRPs and GRPs, which then generate neuronal and glial cell types, respectively. Model 2 has proposed that neurons and glia cell arise from bipotential progenitors in spatially distinct domains, which generate combinations of one neuronal and one glial cell type, rather than from lineage-restricted progenitors.Citation6 Model 3 suggests that NSCs in the motor neuron progenitor (pMN) domain could sequentially generate separate olig-positive motor neurons, oligodendrocyte and probably olig-negative astrocyte precursors.Citation7 Temporally specific genes may coordinate an intrinsic developmental program, since progenitors grown in culture are found to produce certain progeny at given times, much like what progenitors do in vivo.Citation1
The mechanisms of mammalian NSC commitment are largely unknown, which may include extrinsic and intrinsic mechanisms, such as the changes of signal environment, transcription factor expression patterns and epigenetic modifications.Citation8 This review will focus on the signaling pathways and transcriptional factors which are thought to be involved in NSC fate determination. The composition and features of major signaling pathways for cell fate determination in CNS, the signals and transcription factors which initiate and promote neurogenesis, astrogenesis and oligogenesis and the mechanisms that control the timing of neural specification will be discussed sequentially.
Major Signaling Pathways Involved in CNS Development
Despite the difference in themes, each of the development stages involves induction, proliferation, locomotion, cell type or subtype determination and, meanwhile, prevention of inappropriate cell formation. To reach this goal, a precise mechanism is established, by which different signaling pathways play their roles exactly and in coordinative manners according to the time, space and intensity. So far, a panel of pathways has been proved to be involved in CNS development in stage-related patterns (). The panorama of each of the signaling pathways becomes increasingly clear in terms of extracellular stimulators, the components of cytoplasmic signaling cascade, the molecular targets and the consequences on gene transcription.
Wnt pathway.
In canonical Wnt pathway, coordinative interactions of Wnt proteins with other relevant components, β-catenin, adenomateous polyposis coli (APC), glycogen synthase kinase-3β (GSK-3β), Axin and Conductin, regulate cell proliferation and differentiation in the processes of embryogenesis.Citation9 Given lack of Wnt expression, the excess β-catenin in cytoplasm is phosphorylated by functional interactions of GSK-3β, Axin, Conductin and APC, and subjected to degradation by the ubiquitin proteasome system. Activation of Wnt signaling pathway leads to inhibition of GSK-3β activity, resulting in accumulation of cytoplasmic β-catenin that then enter to nucleus. Wnt signaling can be accomplished when β-catenin molecules translocalize into the nucleus and complex with T-cell factors (TCFs) and/or lymphocyte enhance factors (LEFs) to activate transcription of multiple target genes.Citation10
Wnt signaling is implicated in the control of cell growth and differentiation during CNS development.Citation11 Many studies show that this signaling induces neuronal and astroglial differentiation but suppresses oligodendroglial differentiation.Citation11–Citation13 Wnt signaling promotes proliferation of early NPCs, while plays positive roles in neurogenic phase,Citation14 presumably due to the difference of epigenetic statuses in the early and late NPCs.
Notch pathway
The Notch proteins are 300-kDa single-pass transmembrane receptor, which function in CNS development by guiding cell fate determination.Citation15 So far, four Notch receptors (Notch1–Notch4) and five structurally similar Notch ligands, Delta 1, Delta 3, Delta 4, Jagged 1 and Jagged 2, have been identified in mammals. The extracellular domain of the two single-pass transmembrane ligands, delta and serrate (jagged), interacts with the extracellular domain of Notch receptor on the adjacent cell. This ligand binding triggers a series of proteolytic events that results in the cleavage of Notch to generate Notch intracellular domain (NICD). Since NICD contains nuclear import signals, it has the chance to translocate into the nucleusCitation16 and then associates with the constitutive DNA binding protein CSL (CBF1, Suppressor of hairless, Lag-1). By this way, the CSL complex is converted from a transcriptional repressor to a transcriptional activator.Citation17 In the nervous system, Notch signals activate expressions of HES (hairy and enhancer of split paralogues) family genes such as Hes1 and Hes5 as well as the HES-related genes Hesr1 and Hesr2 which belong to basic helix-loop-helix (bHLH) transcriptional repressor.Citation18 Recently, the gene encoding brain lipid-binding protein (BLBP) was identified as a Notch target.Citation19 This gene has RBPjκ (recombining binding protein suppressor of hairless) binding sites which respond to activated Notch. Additionally, both N-Myc and c-Myc are targeted by Notch, which play manifold roles in regulating cell maintenance, renewal and apoptosis. These evidences reflect the widely ranged importance of Notch signaling in the developmental processes such as stem cell self-renewal, cell fate determination and instruction of differentiation and apoptosis.Citation15 During mammalian CNS development, Notch signaling plays active roles in maintaining neuronal progenitor renewal, inhibiting neuronal commitment, promoting astrocyte-oriented glial fates, preventing oligodendrocyte formation as well as brain morphogenesis and neuronal migration.Citation20
BMP pathway
BMPs (Bone Morphogenetic Proteins) are a large subclass (more than 20 members) of the transforming growth factor-beta (TGFβ) super family, which are expressed in many tissues under physiologic conditions, and are regulated through reversible interactions with extracellular antagonists, including noggin, chordin, follistatin and DAN.Citation21 BMPs are classified into several subgroups such as BMP2 and BMP4 on the basis of sequence similarities and homology.Citation22 The action of BMP is mediated by heterotetrameric serine/threonine kinase receptors. There are two specific BMP receptor subunits, BMPRI (BMP Receptor Type-I) and BMPRII, which are required for high-affinity binding and signaling.Citation23 The binding of BMP to its receptor complex results in BMPRI activation and then SMAD1, SMAD5 and SMAD8 phosphorylations.Citation24 The phosphorylated BMP-specific SMADs form a complex with the co-SMAD or SMAD4 and then translocate into the nucleus to activate transcription of specific genes by binding with the GCCG or CAGA motif in the promoter regions of many BMP-responsive genes.Citation25 They are also recruited to the promoters of BMP-responsive genes by high affinity cofactors such as OAZ, which binds to the promoter of STAT3, leading to the transactivation of the glial fibrillary acidic protein gene, a marker for astroglial differentiation.Citation23,Citation24,Citation26 BMPs are involved in many developmental processes, including cell proliferation, differentiation, apoptosis and intercellular interactions during morphogenesis in a gene dosage dependent manner. In the ectoderm, BMPs activate two biochemical pathways, one mediated by SMADsCitation27 and the other mediated by the p38/MAPK (mitogen-activated protein kinase) pathway downstream of TAK1 (TGFβ activated kinase-1).Citation28
In addition to promoting bone formation, BMPs, especially BMP2 and BMP4, exhibit diverse activities in brain development.Citation29 They act at different stages of neural development and in different regions of the CNS to regulate proliferation and determine the cell fate and the manner of differentiation. BMP signaling also inhibits the transformation from primitive ectoderm into neural ectoderm, and once the neural tissue is established, BMP signaling has a positive influence in the regulation of dorsal neural cell type formation.Citation30 Moreover, BMP signaling cross-talks with other key developmental pathways such as Wnt and Sonic Hedgehog (Shh) to coordinate cell proliferation and patterning, allowing the formation of the appropriate numbers and types of differentiated neurons.Citation31 BMPs induce astrocyte specification rather than proliferation of pre-existing astrocyte precursors and inhibit oligodendrocyte specification.Citation32
STAT3 pathway.
Signal transducer and activators of transcription 3 (STAT3) is an important family member of cytoplasmic proteins with SH2 (Src Homology-2) domains which act as signal messengers and transcription factors. STAT3 is activated via the tyrosine phosphorylation cascade after ligand binding with the cytokine receptor-kinase complex and growth factor-receptor complex such as IL-6 (Interleukin-6), OSM (Oncostatin-M), leukemia inhibitory factor (LIF), and ciliary neurotrophic factor (CNTF).Citation33 The phosphorylated STAT proteins dimerize and move into the nucleus, recruiting p300 and binding to specific sequences in target gene promoter. STAT3-mediated signaling is one of the main mechanisms for promoting astrocyte differentiation by inhibiting neuronal differentiation in the embryonic cortex.Citation34 It has been known that STAT3 can promote NSC proliferation, while in the presence of activated BMP and Notch signaling, it induces astrogenesis.Citation35,Citation36 Because of the above molecular effects in different stages of embryonic CNS development, any alteration of this signaling may lead to the developmental defectsCitation37 or primitive neuroectodermal tumors such as medulloblastoma due to mal-differentiation.Citation38
Shh pathway.
Sonic hedgehog (Shh) is a member of the hedgehog family. Shh was initially identified as a morphogen that is directly responsible for dorso-ventral patterning of the CNS.Citation39 As a secreted protein, Shh undergoes autocatalytic processing and modification for gaining activity prior to secretion. The Shh precursor protein is cleaved to yield a ∼19 kDa N-terminal domain (signaling domain; N-Shh) and a ∼25 kDa C-terminal domain (catalytic domain). Cholesterol modification is important for secretion and long-range activity of the Shh protein.Citation40 The canonical Hedgehog signaling is mediated by two transmembrane proteins, a twelve-pass membrane protein binds Hedgehog ligand Patched (Ptc) and a seven-pass membrane protein as a signal transducer Smoothened (Smo).Citation41 When Shh is absent, Ptc acts as a transcriptional repressor and constitutively interacts with and inhibits Smo activity. When Shh binds Ptc, the inhibitory effects of Ptc's on Smo are released, leading to the nuclear translocation of Ci/Gli protein as a transcriptional activator. Gli is the key transcription factor for Shh signaling. There are three Gli genes with distinct transcriptional functions in vertebrates. The downstream genes of Shh signaling include proliferation-related genes N-Myc, cyclin D1, E2f1 and E2f2 and those involved in de novo vascularization during embryonic development such as all three VEGF-1 (vascular endothelial growth factor-1) isoforms and angiopoietins-1 and -2.Citation42,Citation43
Shh is involved in the determination of cell fate and embryonic patterning during early vertebrate development in time and space-associated patterns. Shh is able to promote proliferation of both neural and non-neural tissues. It confers on neural stem cells proliferating competence both temporally and spatially by regulating the expression of N-Myc, cyclin D1, E2f1 and E2f2.Citation42,Citation43 In addition to the roles in proliferation and differentiation, Shh signaling seems to control cell death,Citation44 because some of its downstream genes such as N-Myc and cyclin D1 are deeply involved in the regulation of cell survival. Shh signaling has been implicated in the specification and subsequent development of oligodendrocytes. A possible molecular link between Shh signaling and oligodendrocyte specification was revealed with cloning a subclass of neural basic helix-loop-helix (bHLH) transcription factors, Olig1 and Olig2.Citation44 Again, alterations of Shh signaling are closely associated with neuroectodermal tumor formation.Citation45
PDGF/EGF pathway.
FGF/PDGF/EGF belongs to receptor tyrosine kinase superfamily, which exerts their stimulatory effects by binding to both of isoforms of these tyrosine kinase receptors. Ligand binding induces receptor dimerization and autophosphorylation, allowing binding and activation of cytoplasmic SH2-domain (Src homology) containing signaling molecules.Citation46 Activation of their intrinsic tyrosine-specific protein kinases results in increased turnover of phosphatidylinositols and the sequential release of inositol triphosphates. These events coincide with the activation of other tyrosine kinase pathways, because the most important downstream protein tyrosine kinases (Tyk) activated in this cascade belong to the Janus kinase (JAK) that leads to activation of JAK-STAT signaling and initiation of astrogenesis. Additionally, FGF/PDGF/EGF plays its roles together with other signaling pathways. For example, it functions coordinatively with Notch and beta1-integrin Pathways in neural stem cells.Citation47 In neural precursors, the PDGF/EGF-activated JAK-STAT pathway involves two tyrosine kinases, JAK-1 and Tyk-2, resulting in nuclear translocation of two latent cytoplasmic transcription factors, STAT-1 and STAT-2. Both PDGF and EGF promote neuronal differentiation or oligodendrocyte in committed NSCs, while promote proliferation during the early phase of NSC development.Citation48
Regulatory Factors in Neuronal Fate Determination
Main signaling pathways for neuronal fate determination.
Wnt signaling. Wnt ligands and receptors are expressed and play their roles in both the expansion and neurogenic phases.Citation49 It seems that β-catenin, a centrol player of Wnt signaling, exerts effects on proliferation and differentiation or both, depending on the context of other signaling cascades.Citation12 The effects of Wnt signaling in cultured neural progenitor cells are modified from pro-differentiation to maintenance of the proliferating state by FGF2.Citation12,Citation49,Citation50 Ectopic expression of stabilized β-catenin in early NPCs (E8) results in over proliferation of NPCs and an enlarged neocortex.Citation49,Citation51 Wnt exhibits its neurogenic effects on late NPCs (E13.5), because it causes cell cycle arrest and neuronal differentiation in developing neocortex.Citation49,Citation52,Citation53 The neurogenic effect of Wnt signaling is powerful in terms of induction of neuronal differentiation because nuclear accumulation of β-catenin alone is sufficient to induce neuronal lineage commitment. Canonical Wnt pathway increases the expression of neurogenin1 (Ngn1) and Ngn2, a sort of proneural basic helix-loop-helix (bHLH) proteins, through direct activation of their promoters by the β-catenin/TCF complex.Citation49,Citation52 The critical role of β-catenin in Ngn1 and Ngn2 expression was also evidenced by conditional knockout of β-catenin gene in the cerebral cortex and neural crest stem cells.Citation49,Citation53,Citation54 All these results indicate the instructive role of Wnt pathway in neurogenesis. Interestingly, the Wnt/ β-catenin pathway promotes proliferation via upregulation of cyclin D1, cyclin D2, c-Myc expression in other systems, while it appears to promote neuronal differentiation via upregulating Ngn1 and Ngn2 expression in the neocortex.Citation49,Citation52
Notch signaling. NSCs may be manipulated and enriched via regulating Notch activation. Ectopic expression of the Notch target Hes1 in the developing forebrain of mouse embryos inhibited neuronal differentiation, keeping the HES1-positive cells at the embryonic ventricular or the adult ependymal zone.Citation20 Altered expression of other Notch targets such as HES5, HESR1 and HESR2 may result in similar consequences. Therefore, Notch maintains pluripotent neural stem cell (NSC) renewal by inhibition of their differentiation until the correct differentiation cue is available.
For a normal CNS development, it is necessary to keep neurons in an appropriate proportion and to initiate gliogenesis in time. Therefore, the expression of proneural genes should be restrained as neurons have reached to suitable number at individual positions. A mechanism called ‘lateral inhibition’ is thus developed, which is defined as the commitment to a neural fate by one cell had the consequence of inhibiting its neighbors to follow the same fate.Citation55 Notch ligand in the progenitor cells activates the Notch signaling cascade in neighboring cells, resulting in the expression of Hes/Her/Esr genes which, in turn, directly downregulate proneural gene expression. Through this approach, the expression of proneural genes is restricted to single cells which enter a neural-differentiation pathway.Citation55 During vertebrate neurogenesis, proneural genes such as Mash1 and Ngns trigger transcriptional activation of the Notch receptor ligand Delta. Therefore, the commitment to a neural fate by one cell had the consequence of inhibiting its neighbors to follow the same fate. The embryos lacking Notch function may exhibit neurogenic phenotype because the disruption of Notch-dependent cellular communications causes all of the neighboring cells to develop into neuronal progenitors. When the mouse embryo harbors the mutant forms of those Notch effectors, virtually all the neural stem cells differentiate prematurely into neurons, resulting in severe disorganized neural tube and absence of normal brain structures, suggesting the significance of Notch signaling in controlling appropriate cellular composition of CNS by preventing surplus neuron formation and promoting glial progenitor-to-astrocyte transition.Citation20
It has been found that Notch 1 and Notch 2 present in the nuclei of almost all cortical post-mitotic neurons and that many terminally differentiated neurons expressed the compositional elements of Notch pathway.Citation56 These evidences suggest that Notch signaling is paved in mature neurons but unable to reverse the differentiation process. In other words, Notch and its signaling components may be regarded as one of the terminal differentiation symbols of neurons. So far, the biological significance of those Notch proteins in mature neurons remains to be figured out.
BMP signaling. BMPs promote proliferation of early NPCs and to induce expression of BMP receptor-IB through BMP receptor-IA.Citation30,Citation57 When the amount of BMP receptor-IB reaches to a threshold level, BMPs induce mitotic arrest and neuronal differentiation through BMP receptor-IB.Citation58 This evidence suggests that BMPs function differently as a mitogen at the early stage and as a differentiation factor at the later stage of CNS formation.
Transcription factors in neuronal fate determination.
Proneural genes encode bHLH family transcription factors and play an important role in neurogenesis. Mutation analysis in the mouse showed clear proneural activity for Mash1, Ngn1 Ngn2, and possibly Mash1 and Mash5, whereas other neural bHLH genes are involved in specifying neuronal fates or in neuronal differentiation instead of proneural role.Citation55,Citation59 In the formation of spinal cord, Mash1, Ngn1 and Ngn2 are expressed in most progenitors, except in two domains which are located at the ventral and dorsal ends of the neural tube where Ngn3 and Math1 are expressed, respectively. Although the role of proneural genes has not yet been systematically examined in most regions of the brain, Mash1, Ngn1 and Ngn2 are co-expressed in the dorsal telencephalon, and account for the generation of all progenitors of the cerebral cortex.Citation60 In contrast, Mash1 is the only known proneural genes to be expressed in the ventral telencephalon and some progenitors persist and differentiate normally although a large fraction of progenitors is missing in this region in Mash1 mutants, indicating the presence of other genes with proneural activity.Citation61
Many of neuronal-differentiation genes are structurally related to proneural genes, suggesting that distinct bHLH genes acting in cascades underlie the sequential steps of cell determination and differentiation. In vertebrates, bHLH genes of the NeuroD family have the characteristics of differentiation genes as well. NeuroD is required for the proliferation, differentiation and survival of granule cells in the cerebellum and hippocampus. The genes of NeuroD family act downstream of vertebrate proneural genes in a manner very similar to ase and cato in Drosophila neurogenesis.Citation62 Activation of NeuroD by Ngns is likely direct, because of the activation of ase by ac and sc. Nieto M, et al. showed that vertebrate proneural genes promoted neuronal fates and inhibited glial fates simultaneously, indicating their important role in the transition from neurogenesis to gliogenesis.Citation63
Transcription factor and signaling interplays in neuronal fate determination.
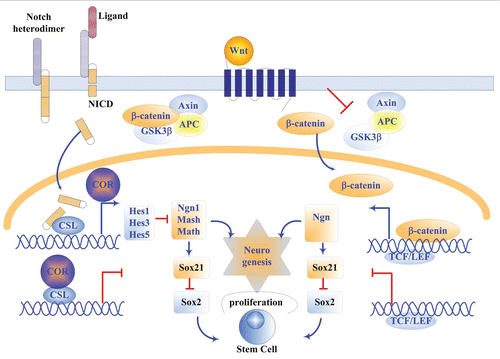
The balance of proliferation and differentiation of NSCs is mainly regulated by the opposing activities of repressor bHLH and activator bHLH as well as Sox1-3 and Sox21 which are largely determined by the activities of Notch and Wnt signalings. Notch signals triggers the expression of Hes family genes such as Hes1 and Hes5 that in turn inhibit Sox21 expression via downregulating the expression of proneural genes. Therefore, attenuation of Notch signaling is a critical event for neurogenic commitment of NSCs.Citation64,Citation65 Unlike Notch, Wnt signaling is regarded as one of the few signaling pathways that have been shown to promote the neuronal fate in an instructive manner. It induces neuronal differentiation through promoting proneural Ngn1 and Ngn2 expressionCitation49 and suppressing Hes1 and Hes5 expression in late embryonic and postnatal development stagesCitation14 ().
Regulatory Factors in Astroglial Fate Determination
Main signaling pathways for astroglial fate determination.
Signaling pathways involved in the induction of astrocyte specification are thought to be induced via cytokines such as CNTF, LIF and IL-6 as well as the signaling mediated by BMP and Notch proteins.Citation66,Citation67
STAT3 signaling. The typical effect of STAT3 signaling is to promote GFAP expression, a specific biomarker of glial cells. As a transcription factor, STAT3 induces GFAP expression via binding with STAT-responsive element in GFAP promoter region when activated by CNTF and LIF.Citation33 The capacity of STAT3 in triggering GFAP expression largely depends upon the stage where neural progenitor cells are in. When LIF and CNTF mediated phosphorylation occurs to STAT3, the phorsphorylated STAT3 molecules translocalize into nucleus and trigger GFAP expression in relatively late-(E14.5) but not early-stage (E11.5) NPCs.Citation49,Citation68 Similar phenomena could be observed in Notch-induced GFAP expression.Citation69 Instead of GFAP induction, Notch and LIF/CNTF-STAT3 promote self-renewal of early NPCs.Citation70 Interestingly, CNTF induces almost the same levels of STAT3 phosphorylation in early and late NPCs, but GFAP is induced only in the late NPCs, suggesting that the same stimuli result in different biological outcomes in early and later NPCs. Among the genes activated by STAT1/3 in late embryonic NEPs, there are several composition elements of the Jak-Stat pathway itself, including gp130, Jak1, Stat1 and Stat3. Thus, STAT signaling triggers an auto-regulatory loop that reinforces itself and presumably tends to consolidate and stabilize the astrocyte phenotype.Citation68
BMP signaling. Bone morphogenetic proteins have been implicated in the development of gliocytes. It has been proved that BMPs are involved in astrocytic differentiation of neural precursor cells. In the nuclei, BMP-downstream transcription factors, i.e., Smad proteins, induce expression of astrocyte-specific genes in cooperation with another cytokine signaling.Citation71 The typical example is that BMP signaling mutant mice exhibit glial cell maturation defects, indicating that BMP signaling is the critical element for astrocyte generation and maturation.Citation72
Notch signaling. Notch directly promotes the differentiation of many glial subtypes except oligodendrocytes in an instructive manner. Notch1 could be detected in the nuclei of radial glial cells but not in neuronal precursor cells positive in neuronal basic helix-loop-helix proteins or in the differentiating neurons of the embryonic forebrain.Citation73 Furthermore, activated Notch signaling in retinal progenitors blocks rat neuronal differentiation and promotes the expression of Müller glia markers.Citation70 In postnatal mouse retinae, Ectopic expression of Hes5 promotes gliogenesis at the expense of neurogenesis;Citation20 conversely, erasing Hes5 expression results in significant reduction of Müller glia.Citation74 These data thus suggest that Notch activation is essential for astrogenesis by inhibiting the tendency of neuronal formation. Notch appears to promote the astrocytic phenotype through its downstream effectors Hes1 and Hes5 which are known to promote astrocytic differentiation via suppressing the function of proneural bHLHs.Citation49
Wnt signaling. In addition to neuronal induction, Wnt signaling is able to promote astroglial differentiation and suppresses oligodendroglial differentiation in phase-dependent fashion through indirectly regulating gliogenesis by inducing BMPs in neuronal cells.Citation12,Citation75 Wnt-induced BMPs are then secreted to the extracellular space to promote astroglial differentiation and inhibit oligodendroglial differentiation of surrounding cells. Oligodendroglial differentiation can only proceed when attenuation of Wnt and BMP signaling occurs. In the aspects of astrogenic promotion and oligogenic suppression, the functional similarities remain not only between Wnt and BMP but also between them two and Notch signaling.Citation75 Therefore, a coordinative functioning of Wnt, BMP and Notch pathways is essential in determining the courses of neurogenesis and gliogenesis. In another word, Wnt signaling never plays its roles independently but through interplays with other signaling elements which provide positional information and induce stepwise cell fate specification.
Transcription factors in astroglial fate determination.
Proneural bHLH transcription factors such as Ngn1 and Mash1 repress gliogenesis by inhibiting the activity of the Jak-STAT pathway.Citation76 They achieve this goal through two approaches: (1) competing with STAT1/3 for binding to p300 and CBP which are present in limiting quantities in the cells and (2) directly inhibiting STAT activation. Sun and his colleaguesCitation77 proposed a sequestration model, where Ngn1 inhibits glial differentiation by sequestering activator complexes away from glial-specific genes and by directly inhibiting STAT3 activation when the cortical precursor cells are neurogenic. These evidences largely explain how the switch of neurogenesis is turned to gliogenesis during development. As gestation proceeds, Ngn1 expression is downregulated and STAT3 can form a complex with Smad1, bridged by p300, to effectively induce astrogenesis. The factor(s) leading to Ngn downregulation remains to be determined. It has been shown that growth hormone (GH) acts through the Jak/STAT pathway to activate the inhibitory STAT5, resulting in the downregulation of Ngn1 expression in cultured neurospheres.Citation78 GH and its receptor are expressed in the rodent brain as early as embryonic day 10 and reach the peak just before birth. It is therefore conceivable that GH might play a role in the neuron-astrocyte switch. However, this could not easily explain the fact that neuronglial switching occurs approximately on schedule in primary NEPs in the absence of added GH. An alternative option is that robust Jak/STAT signaling by itself might inhibit Ngn expression.Citation79
Transcription factor and signaling interplays in astroglial fate determination.
Multiple signaling pathways participate in the initiation of astrogenesis. The onset of astrocyte formation requires activation of the gp130-JAK-STAT pathway that is thought to be induced by cytokines such as ciliary neurotrophic factor/CNTF, leukaemia inhibitory factor/LIF and interleukin-6/IL-6.Citation80 It has been noticed that the astrogenic induction is the outcome of interactions or crosstalks of STAT signaling with the ones mediated by BMP and Notch proteins.Citation35,Citation36,Citation69,Citation81 STAT3 might co-operate with BMP2 as well as Notch family members to induce glial fibrillary acidic protein (GFAP) expression. LIF and BMP2 were found to act in synergy on primary fetal neural progenitor cells to induce astrocyte formation by p300 bridged STAT3-Smad1 complex.Citation82 LIF can also consequently activate BMP and Smad1 by triggering STAT3 activation.Citation35 Although STAT3 signaling suppresses the expression of proneural bHLHs to inhibit neurogenesis by promote Notch signaling,Citation83 it can be activated reversely by Hes proteins that directly recruit the upstream kinase JAK2.Citation69 Additionally, Notch-Hes signaling is found to induce astrocytic differentiation through activation of STAT3 by directly recruiting the upstream kinase JAK2. In addition to the interaction with STAT3 signaling, Notch signaling also promotes astrogliogenesis via direct CSL-mediated GFAP activation.Citation84 Furthermore, recruitment of p300 to the Notch intracellular domain (NIC)-containing complex can be facilitated by activated Smad1, indicating the contribution of BMP2-mediated enhancement of Notch-induced Hes-5 expression.Citation85 Consequently, a functional coordination of Notch, BMP and STAT3 signaling is important in the accomplishment of astrogenesis by maintaining appropriate intensity of STAT3 siganling. It should be pointed out that, a positive autoregulatory loop of STAT signaling is formed during the onset of astrogliogenesis, by which STAT1/3 directly induces the expression of various components of the Jak-STAT pathway, including gp130, Jak1, STAT1 and STAT3.Citation86 By this way, STAT signaling is further strengthened, leading to upregulation of its downstream genes and consolidated astrocyte phenotype.
Regulatory Factors in Oligodendroglial Fate Determination
Main signaling pathways for oligodendroglial fate determination.
The origin of oligodendroglia has been debated for years. One of the well accepted notions is that oligodendroglia from both dorsal and ventral sources become active at different times during development. Oligodendrocyte precursors first arise in a restricted ventral part of the embryonic spinal cord and migrate laterally and dorsally from there; later on, secondary sources develop in the dorsal cord.Citation87 Apparently, oligodendrogenesis can be induced by several signaling pathways depending on different origins.
Shh pathway.
Shh signaling has been implicated in the specification and subsequent development of oligodendrocytes. Several studies have shown that Shh promotes the generation of pMN-derived OLPs (progenitor of motor neurons derived oligodendrocyte progenitor) in the spinal cord.Citation88 In the absence of Shh signaling, ventrally derived OLPs fail to form in the spinal cord or forebrain, while transplanting Shh-expressing tissue adjacent to the dorsal neural tube in vivo or adding Shh to spinal cord explants in vitro induces the formation of ectopic OLPs, highlighting the importance of spatially restricted Shh signalling in directing OLP generation.Citation89 Several studies demonstrated that sonic hedgehog (Shh) signaling was specificly required during oligodendrocyte development. For instance, Shh promotes NSCs to transform to oligodendrocyte progenitor/OLPCitation89,Citation90 and induces pMN-derived OLPs (progenitor of motor neurons derived oligodendrocyte progenitor) in primary cultures of dissociated forebrain NPCs.Citation91,Citation92
Shh-independent pathway. Although the activated Shh signaling is important for oligogenesis, a Shh-independent pathway to oligodendrocyte specification is evidenced because NSCs from Shh or Smo null mice can still generate OLPs in culture.Citation93 It was demonstrated that FGF2,Citation93,Citation94 also participate in oligodendrocyte specification according to different origin of oligodendroglia.Citation92,Citation95 NSCs from the cortex and the dorsal spinal cord can generate OLPs when they are exposed to FGF2 and cyclopamine, a potent inhibitor of Shh signaling. It is therefore possible that FGF might be responsible for oligodendrocyte induction in the dorsal spinal cord and/or forebrain, which are unlikely to be within the range of Shh from the floor plate (spinal cord) or MGE (forebrain).
In addition to Shh and FGF2, insulin-like growth factor I (IGF-I) has also been implicated to influence oligodendrocyte specification, because multipotent adult neural progenitors can be induced to generate oligodendrocytes in vitro after treating IGF-I, IGF-II or insulin.Citation96 The oligodendrogenic activity of IGF-I is thought to be mediated at least in part by inhibition of BMP signaling.Citation96
Notch signaling is essential at the early stages of specification of the oligodendrocyte lineage, because constitutive activation of Notch signaling results in excess OLPs at the expense of motor neurons.Citation97 On the other hand, persistent Notch signaling can inhibit in vitro oligodendrocyte maturation, and loss of Notch signaling leads to premature differentiation of OLPs to oligodendrocytes in vivo.Citation98 These data thus suggest that Notch signaling might regulate both the specification of OLPs and oligodendrocytes maturation.
Opposite effects of BMPs on oligodendrogenesis. BMP signaling is another element that influences oligodendrocytic fate determination but in an opposite way of Shh signaling. The members of the BMP family have been proposed to act as negative regulators of oligodendrocyte specification because BMPs induce expression of Id proteins that interact with OLIG proteins to inhibit OLP differentiation and maturation.Citation58 In vitro study revealed that when forebrain or spinal cord neural stem cells were incubated with BMPs, they failed to develop oligodendrocytes, irrespective to the presence of activated Shh or FGF2 signaling.Citation99,Citation100 Apparently, dynamic transition of signaling patterns with imbalanced but nonrandom signaling pressure is essential for NSC fate determination.
Transcription factors in oligodendroglial fate determination.
bHLH genes. A possible molecular link between Shh signaling and oligodendrocyte specification was revealed by cloning a subclass of neural basic helix-loop-helix (bHLH) transcription factors, Olig1 and Olig2.Citation91 Induced expression of Olig2 is sufficient for oligodendrocyte specification but not for motor neuron specification and astrocyte repression. On the other hand, when Olig1 and Olig2 are doubly knockout, the mice have no oligodendrocytes at E18.5, indicating that these two Shh-target genes are both necessary for oligodendrocyte formation.Citation101 It has been found that ES cell-derived Olig2+ cells can give rise to both motor neurons and oligodendrocytes, depending on the time/phase at which differentiation is initiated. It should be noted that Shh conducts its functions such as motor neuron specification much earlier than the time of oligodentrocyte specification. Therefore, in order to keep CNS development in a steady pace, other intrinsic and extrinsic factors may exist at that time to inhibit the oligogenic effects of Shh.
Olig1 and Olig2 were originally identified as two bHLH transcription factors which mark the oligodendrocyte lineage from very early stages.Citation101 In the spinal cord, these two transcripts can be detected from embryonic day 9 onwards and known to be induced by Shh signaling. The transcripts are initially observed in the third ventral of the spinal cord but soon become restricted to the pMN domain. Olig2 expression persists in pMN throughout motor neurons development; it is rapidly extinguished in post-mitotic MNs but remains in OLPs.Citation102 Expression of Olig1 is more dynamic at early times prior to the appearance of OLPs.Citation103 Both Olig1 and Olig2 genes are expressed throughout the oligodendrocyte lineage including the mature oligodendrocytes. Functional studies on mice have demonstrated the significance of Olig2 in either motor neuron or oligodendrocyte specification.Citation103 In a mouse embryonic stem (ES) cell line, Olig2 requires the cooperation of retinoic acid (RA) and Shh for motor neuron specification.Citation104 Olig2 bypasses the need of Shh in oligodendrocyte specification but can not bypass the CNTF-STAT signaling to repress astrocyte differentiation.Citation104 Unlike Olig2 that seems essential for oligodendrocyte development, initial evidence indicates a redundant, non-essential function for Olig1.Citation105 However, more recent work with the Olig1 mutant mice questions the previous data and suggests that Olig1 is, in fact, a central regulator of oligodendrocyte maturation and myelinogenesis.Citation106
Another bHLH gene that has recently been found to play a role in oligodendrocyte development is the proneural gene Mash1. In the early telencephalon, Mash1 is co-expressed with Olig2 at the time of oligodendrocyte specification in the medial ganglionic eminence (MGE)/anterior entopeduncular area (AEP) and its expression is briefly maintained in initial populations of migrating OLPs.Citation61 Although the data indicate both Mash1-dependent and Mash1-independent OLPs in the telencephalon, the precise function of Mash1 in the oligodendrocyte lineage remains to be further investigated.
Nkx 6.1, Nkx 6.2 and Nkx 2. Nkx 6.1 and Nkx 6.2 are two homeodomain transcription factors which are induced by Shh in the ventral spinal cord.Citation107 Within the pMN domains, Nkx6.1 and Nkx6.2 appear to regulate the expression of Olig2 in a dose-dependent manner. In Nkx6.1/Nkx6.2 mutants, all the motor neurons and oligodendrocytes generated from the pMN domain are eliminated, demonstrating an essential role for these two genes in oligodendrogenesis from this region;Citation100 on the other hand, oligodendrocyte development from dorsal precursors is unaffected, indicating oligodendrocyte specification from dorsal and ventral precursors are significant different.Citation108 Loss-of-function studies in mouse have suggested a redundant or non-essential function for Nkx 6.2 in the spinal cord, whereas, in the absence of Nkx6.1, expression of Olig2 is delayed and greatly diminished and the generation of MNs is severely impaired. Therefore, Nkx6.1 seems more responsible for Olig2 expression, while Nkx6.2 is expressed in myelinating oligodendrocytes where it appears to be involved in regulating axon-glial interactions at myelin paranodes.Citation109,Citation110
Nkx2.2 is another homeobox transcription factor involved in oligodendrogenesis in the neural tube.Citation111 The expression of Nkx2.2 is induced by Shh at the time of neural plate induction and is restricted to a small region of the VZ adjacent to the floor plate. The expression domains of Olig2 and Nkx2.2 are overlapping till the end of neurogenesis. Nkx2.2 expands dorsally into the pMN domain at the onset of oligodendrogenesis. It has become clear that Nkx2.2 is more important during the maturation of oligodendrocytes rather than in oligodendrocyte specification.Citation87,Citation112
Sox genes. The Sox E family, comprising Sox 9, Sox 8 and Sox 10, has a prominent role in oligodendrocyte development. They are expressed at different stages of oligodendrocyte development.Citation113,Citation114 In loss-of-function experiments, Sox9 had to be ablated in a tissue-specific manner. The mice lacking Sox9 exhibit severe reduced numbers of oligodendrocytes as well as astrocytes, indicating Sox 9 as a major component of the neuron-glia switch on.Citation113,Citation115 Sox 8 overlap partially the function of Sox 9 in oligodendrocytes.Citation116 On the other hand, the loss of Sox10 results in normal numbers of oligodendrocyte progenitors but a failure to maturation, indicating the requirement of this protein for terminal differentiation.Citation117,Citation118
Mechanisms for Timing Neural Specification
The mechanisms underlying mammalian NSC commitment have not yet been fully characterized, but are thought to include extrinsic and intrinsic factors, such as the accumulation of signaling and transcription factors just discussed above and dynamic epigenetic modifications. Sequential generation of neurons and glial cells is fundamental features in vertebrates. Some studies on this area have shed light on the essential signaling pathways in controlling the stepwise transition from NSC to proliferation; neurogenesis to gliogenesis.
Regulating the transition from NSC to proliferation.
As referred above, the transition of proliferation to differentiation of NSCs is mainly regulated by the opposing activities of repressor bHLH and activator bHLH as well as Sox1-3 and Sox21 which are largely determined by the activities of Notch and Wnt signalings. Among these signalings, the Wnt/β-catenin pathway is noticeable because it could promotes proliferation via upregulation of cyclin D1, cyclin D2, c-Myc and c-jun expression in other systems, while it appears to promote neuronal differentiation via upregulating Ngn1 and Ngn2 expression in the neocortex.Citation52 Wnt signaling together with its nuclear effectors, β-catenin and T-cell factor (TCF) DNA-binding proteins, form a regulatory system, by which β-catenin/TCF complexes distinguish precisely between genes to be activated and genes to remain silent in a given cell system and/or at a given time. It has been known that activated Wnt signaling is a major initiation and promotion factor of neurogenesis because of its ability to selectively triggering a panel of neuronal-asscoiated gene expression in NSC differentiation. It therefore suggests that the transition of NSC to neuron is determined to a certain signaling such as Wnt by selectively triggering a panel of neural fate-determination genes instead of genes directed proliferation. The possibility of the dynamic features of the gene responses to Wnt signaling may be the influence of epigenetic factors including DNA methylation and chromatin re-modeling.Citation119,Citation120
It has been found that only Wnt-responsive promoters are bound by specific subsets of TCFs, whereas nonresponsive Wnt target promoters remain unoccupied.Citation121 It is also found that Wnt-responsive, TCF-bound states correlate with DNA hypomethylation, histone H3 hyperacetylation, and H3K4 trimethylation, whereas inactive, nonresponsive promoter chromatin shows DNA hypermethylation, is devoid of active histone marks and additionally can show repressive H3K27 trimethylation. These data suggest that epigenetic modifications of promoter chromatin and differential promoter occupancy by functionally distinct TCF proteins jointly determine susceptibility to Wnt signaling.Citation121 This result begins to unveil why Wnt signaling play different roles in neural stem cell self-renewal and neurogenesis which exist a long time.Citation122
Controlling stepwise transition from neurogenesis to gliogenesis.
JAK-STAT pathway is a main pathway controlling the onset of astrocyte formation. The neurobiological functions of STAT signaling are phase dependent. CNTF can induce almost the same levels of STAT3 phosphorylation in early and late NPCs, but GFAP is induced only in the later cells,Citation123 indicating that the capacity of STAT3 in triggering GFAP expression largely depends upon the developmental stage of neural progenitor cells.Citation49 This phenomenon again strongly suggests that the same stimuli may result in different biological outcomes during stepwise CNS development. It is therefore possible that different sets of intrinsic factors may present in early and late NPCs, which determine the ways to respond external stimuli such as STAT3 and Wnt signaling via modulating the status of gene accessibility.Citation14,Citation49,Citation124 In this context, the influence of epigenetic factors in NSC fate determination should be taken into account.
STAT3 promotes astrogenesis by promoting GFAP expression. A CpG motif within the STAT-binding element has been found in the promoter region of GFAP gene. It is methylated in early NPCs, but becomes demethylated and accessible to STAT3 later during development or culture.Citation123 Fan, et al. indicated that DNA methylation was a key mechanism in inhibiting the expression of various components of the astrogliogenic JAK-STAT pathway,Citation125 suggesting a causal relationship between DNMT1 (DNA methyltransferase 1) expression and the control of the onset of astrogliogenesis in NPCs. It should be addressed that DNMT1 molecule has also been shown to directly inhibit gene transcription with its N-terminal transcription repression domain, which interacts with other transcription repressor components, including histone deacetylases and histone lysinemethyltransferases for inactive chromatin remodeling.Citation126 DNA methylation not only inhibits astroglial marker genes but also the genes which are essential for JAK-STAT signaling. Thus, demethylation of these two groups of genes and subsequent elevation of STAT activity are key mechanisms which control the timing and magnitude of astroglial differentiation. JAK-STAT activation in neural precursor cells is under the tight control of several independent mechanisms.Citation125 Additionally, the transcriptional activity of GFAP promoter can also be regulated via modification of histone H3.Citation127
This stage-specific DNA methylation and chromatin modification of the important DNA elements might be part of a general mechanism that accounts for some of the intrinsic differences among different stages of NPCs. Though GFAP stands as a specific biomarker for glial cells, it is merely the Type III of intermediary filament proteins and by no means a crucial protein for astrocytic differentiation. Therefore, more important astrocytic determinant genes may be epigenetically regulated during stepwise brain development, leading to differential cellular and molecular responses to the same signaling.
Concluding Remarks
Neural cell fate determination is complicated, which requires diverse signal pathways, transcription factors and epigenetic machineries to participate in. Multiple extracellular signals and corresponding intrinsic transcriptional factors may exert different effects in time- and stage-dependent fashions. Wnt and Notch pathways are important in the specification of neural fates; the former is thought to be a positive regulator, while the later plays the opposite roles. The mediators connecting Wnt or Notch signaling with the fates of neural stem cells are proneural genes which encode bHLH family transcription factors, such as Ngn1, Ngn2 and Mash1. The BMPs and STAT signaling pathways can act independently of one another but also synergize in some circumstances to specify astrocytes. A coordinative function of Wnt, BMP, Notch and STAT signaling is essential in determining the courses of neurogenesis and gliogenesis. Shh signaling has been implicated in the specification and subsequent development of oligodendrocytes. FGF2, insulin-like growth factor I (IGF-I) has also been suggested to influence oligodendrocyte specification according to different origin of oligodendroglia. Transcriptional factors play important roles as mediators between different signaling pathways and neural cell fate determination. Given the availability of the above extracellular and interior molecular elements, the responsiveness of NSCs to these signals is the key point for determining the direction of NSC differentiation, which is largely controlled by epigenetic mechanisms such as DNA methylation, histone modulations and non-coding RNA. As a matter of fact, the involvement of epigenetic mechanisms in embryonic CNS development has been increasingly documented, especially in the recent years. The manners of molecular response to neural-associated signaling are largely regulated by the epigenetic factors which determine the accessibility of the corresponding genes. And the enzymes which regulate epigenetic program often interact with transcription factors to form a panel of complexes to control global gene expression pattern. By this way, local and overall chromatin identities are created, which suit for distinct fate determination of NSCs. The continuous progress in epigenetics of developmental neuroscience may help us to answer how cell fate is precisely determined, remembered and inherited through disclosing the manners of local and global epigenetic programming and re-editing during the transitions of NSC differentiation.
Figures and Tables
Figure 1 Three proposed models for the genesis of CNS cell components. (A) Model1, NRP/GRP model; (B) Model2, MNOP model; (C) Model3, Sequential model. NRPs, neuronal restricted progenitors; GRPs, glial restricted progenitors; MNOP, motoneuron and oligodendrocyte common progenitor; NA, neuron and astrocyte common progenitor; OPC, oligodendroctye precursor; pA, astrocyte precursor; pM, motoneuron precursor.
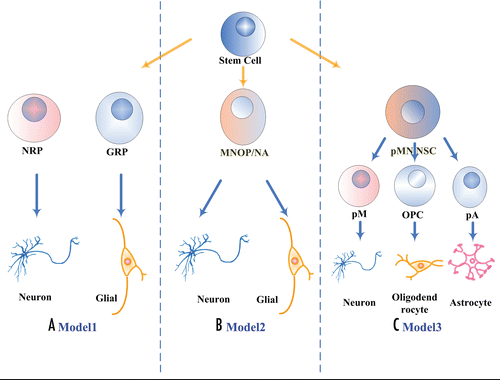
Figure 2 Signaling pathways involved in CNS development. The blue arrows indicate activations, the red lines represent repression.
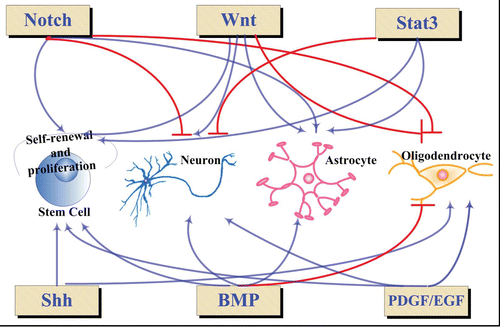
Figure 3 The involvement of Notch and Wnt signaling pathways and their relevant transcription factors in neurogenesis. The blue arrows indicate activating effects and the red lines represent repressive effects. Notch is a negative signal while Wnt is a positive signal in neurogenesis through the mediation by bHLH and SOX transcription factors.
Acknowledgements
This work is supported in part by grants from the National Natural Science Foundation of China (30370384, 30527002 and 30670946) and the special grants for creative research team (2007-7026) and for the key laboratory (20060193) from Liaoning Department of Education.
References
- Merkle FT, Alvarez-Buylla A. Neural stem cells in mammalian development. Curr Opin Cell Biol 2006; 18:704 - 709
- Parnavelas JG, Nadarajah B. Radial glial cells: are they really glia?. Neuron 2001; 31:881 - 884
- Merkle FT, Tramontin AD, García-Verdugo JM, Alvarez-Buylla A. Radial glia give rise to adult neural stem cells in the subventricular zone. Proc Natl Acad Sci USA 2004; 101:17528 - 17532
- Garcia AD, Doan NB, Imura T, Bush TG, Sofroniew MV. GFAP-expressing progenitors are the principal source of constitutive neurogenesis in adult mouse forebrain. Nat Neurosci 2004; 7:1233 - 1241
- Rao MS. Multipotent and restricted precursors in the central nervous system. Anat Rec 1999; 257:137 - 148
- Wu S, Wu Y, Capecchi MR. Motoneurons and oligodendrocytes are sequentially generated from neural stem cells but do not appear to share common lineage-restricted progenitors in vivo. Development 2006; 133:581 - 590
- Liu Y, Rao M. Oligodendrocytes, GRPs and MNOPs. Trends Neurosci 2003; 26:410 - 412
- Chapouton P, Jagasia R, Bally-Cuif L. Adult neurogenesis in non-mammalian vertebrates. BioEssays 2007; 29:745 - 757
- Cheng XX, Wang ZC, Chen XY, Sun Y, Kong QY, Liu J, Li H. Correlation of Wnt-2 expression and beta-catenin intracellular accumulation in Chinese gastric cancers: relevance with tumour dissemination. Cancer Lett 2005; 223:339 - 347
- Clevers H. Wnt/beta-catenin signaling in development and disease. Cell 2006; 127:469 - 480
- Zechner D, Fujita Y, Hülsken J, Müller T, Walther I, Taketo MM, et al. beta-Catenin signals regulate cell growth and the balance between progenitor cell expansion and differentiation in the nervous system. Dev Biol 2003; 258:406 - 418
- Kasai M, Satoh K, Akiyama T. Wnt signaling regulates the sequential onset of neurogenesis and gliogenesis via induction of BMPs. Genes Cells 2005; 10:777 - 783
- Machon O, Backman M, Machonova O, Kozmik Z, Vacik T, Andersen L, et al. A dynamic gradient of Wnt signaling controls initiation of neurogenesis in the mammalian cortex and cellular specification in the hippocampus. Dev Biol 2007; 311:223 - 237
- Hirsch C, Campano LM, Wöhrle S, Hecht A. Canonical Wnt signaling transiently stimulates proliferation and enhances neurogenesis in neonatal neural progenitor cultures. Exp Cell Res 2007; 313:572 - 587
- Grandbarbe L, Bouissac J, Rand M, Hrabé de Angelis M, Artavanis-Tsakonas S, Mohier E. Delta-Notch signaling controls the generation of neurons/glia from neural stem cells in a stepwise process. Development 2003; 130:1391 - 1402
- Lai EC. Notch cleavage: Nicastrin helps Presenilin make the final cut. Curr Biol 2002; 12:200 - 202
- Song W, Nadeau P, Yuan M, Yang X, Shen J, Yankner BA. Proteolytic release and nuclear translocation of Notch-1 are induced by presenilin-1 and impaired by pathogenic presenilin-1 mutations. Proc Natl Acad Sci USA 1999; 96:6959 - 6963
- Yoon K, Gaiano N. Notch signaling in the mammalian central nervous system: insights from mouse mutants. Nature Neurosci 2005; 8:709 - 715
- Anthony TE, Mason HA, Gridley T, Fishell G, Heintz N. Brain lipid-binding protein is a direct target of Notch signaling in radial glial cells. Genes Dev 2005; 19:1028 - 1033
- Louvi A, Artavanis-Tsakonas S. Notch signalling in vertebrate neural development. Nat Rev Neurosci 2006; 7:93 - 102
- Balemans W, Van Hul W. Extracellular regulation of BMP signaling in vertebrates: a cocktail of modulators. Dev Biol 2002; 250:231 - 250
- Farkas LM, Jászai J, Unsicker K, Krieglstein K. Characterization of bone morphogenetic protein family members as neurotrophic factors for cultured sensory neurons. Neuroscience 1999; 92:227 - 235
- Miyazono K, Maeda S, Imamura T. BMP receptor signaling: transcriptional targets, regulation of signals and signaling cross-talk. Cytokine Growth Factor Rev 2005; 16:251 - 263
- Wang W, Mariani FV, Harland RM, Luo K. Ski represses bone morphogenic protein signaling in Xenopus and mammalian cells. Proc Natl Acad Sci USA 2000; 97:14394 - 14399
- Luo K. Negative regulation of BMP signaling by the ski oncoprotein. J Bone Joint Surg Am 2003; 85:39 - 43
- Cheng LE, Reed RR. Zfp423/OAZ participates in a developmental switch during olfactory neurogenesis. Neuron 2007; 54:547 - 557
- Hata A, Seoane J, Lagna G, Montalvo E, Hemmati-Brivanlou A, Massagué J. OAZ uses distinct DNA- and protein-binding zinc fingers in separate BMP-Smad and Olf signaling pathways. Cell 2000; 100:229 - 240
- Keren A, Tamir Y, Bengal E. The p38 MAPK signaling pathway: a major regulator of skeletal muscle development. Mol Cell Endocrinol 2006; 252:224 - 230
- Cahoy JD, Emery B, Kaushal A, Foo LC, Zamanian JL, Christopherson KS, et al. A transcriptome database for astrocytes, neurons and oligodendrocytes: a new resource for understanding brain development and function. J Neurosci 2008; 28:264 - 278
- Liu A, Niswander LA. Bone morphogenetic protein signalling and vertebrate nervous system development. Nat Rev Neurosci 2005; 6:945 - 954
- Sarkar L, Cobourne M, Naylor S, Smalley M, Dale T, Sharpe PT. Wnt/Shh interactions regulate ectodermal boundary formation during mammalian tooth development. Proc Natl Acad Sci USA 2000; 97:4520 - 4524
- Stipursky J, Gomes FC. TGFbeta1/SMAD signaling induces astrocyte fate commitment in vitro: implications for radial glia development. Glia 2007; 55:1023 - 1033
- Moon C, Yoo JY, Matarazzo V, Sung YK, Kim EJ, Ronnett GV. Leukemia inhibitory factor inhibits neuronal terminal differentiation through STAT3 activation. Proc Natl Acad Sci USA 2002; 99:9015 - 9020
- Yanagisawa M, Nakashima K, Taga T. STAT3-mediated astrocyte differentiation from mouse fetal neuroepithelial cells by mouse oncostatin M. Neurosci Lett 1999; 269:169 - 172
- Fukuda S, Abematsu M, Mori H, Yanagisawa M, Kagawa T, Nakashima K, et al. Potentiation of astrogliogenesis by STAT3-mediated activation of bone morphogenetic protein-Smad signaling in neural stem cells. Mol Cell Biol 2007; 27:4931 - 4937
- Taylor MK, Yeager K, Morrison SJ. Physiological Notch signaling promotes gliogenesis in the developing peripheral and central nervous systems. Development 2007; 134:2435 - 2447
- Gong L, Yao F, Hockman K, Heng HH, Morton GJ, Takeda K, et al. Signal transducer and activator of transcription-3 is required in hypothalamic agouti-related protein/neuropeptide Y neurons for normal energy homeostasis. Endocrinology 2008; 149:3346 - 3354
- Yu LJ, Wu ML, Li H, Chen XY, Wang Q, Sun Y, Kong QY, Liu J. Inhibition of STAT3 expression and signaling in resveratrol-differentiated medulloblastoma cells. Neoplasia 2008; 10:736 - 744
- Martí E, Bovolenta P. Sonic hedgehog in CNS development: one signal, multiple outputs. Trends Neurosci 2002; 25:89 - 96
- Jeong J, McMahon AP. Cholesterol modification of Hedgehog family proteins. J Clin Invest 2007; 110:591 - 596
- Wilson CW, Chuang PT. New “hogs” in Hedgehog transport and signal reception. Cell 2006; 125:435 - 438
- Hatton BA, Knoepfler PS, Kenney AM, Rowitch DH, de Alborán IM, Olson JM, et al. N-myc is an essential downstream effector of Shh signaling during both normal and neoplastic cerebellar growth. Cancer Res 2006; 66:8655 - 8661
- Oliver TG, Grasfeder LL, Carroll AL, Kaiser C, Gillingham CL, Lin SM, et al. Transcriptional profiling of the Sonic hedgehog response: a critical role for N-myc in proliferation of neuronal precursors. Proc Natl Acad Sci USA 2003; 100:7331 - 7336
- Fuccillo M, Joyner Al, Fishell G. Morphogen to mitogen: the multiple roles of hedgehog signalling in vertebrate neural development. Nature Rev Neurosci 2006; 7:772 - 783
- Yang ZJ, Ellis T, Markant SL, Read TA, Kessler JD, Bourboulas M, et al. Medulloblastoma can be initiated by deletion of Patched in lineage-restricted progenitors or stem cells. Cancer Cell 2008; 14:135 - 145
- Schlessinger J. Cell signaling by receptor tyrosine kinases. Cell 2000; 103:211 - 225
- Campos LS, Decker L, Taylor V, Skarnes W. Notch, epidermal growth factor receptor, and beta1-integrin pathways are coordinated in neural stem cells. J Biol Chem 2006; 281:5300 - 5309
- Mondal D, Pradhan L, LaRussa VF. Signal transduction pathways involved in the lineage-differentiation of NSCs: can the knowledge gained from blood be used in the brain?. Cancer Invest 2004; 22:925 - 943
- Hirabayashi Y, Gotoh Y. Stage-dependent fate determination of neural precursor cells in mouse forebrain. Neurosci Res 2005; 51:331 - 344
- Israsena N, Hu M, Fu W, Kan L, Kessler JA. The presence of FGF2 signaling determines whether beta-catenin exerts effects on proliferation or neuronal differentiation of neural stem cells. Dev Biol 2004; 268:220 - 231
- Bouhon IA, Joannides A, Kato H, Chandran S, Allen ND. Embryonic stem cell-derived neural progenitors display temporal restriction to neural patterning. Stem Cells 2006; 24:1908 - 1913
- Hirabayashi Y, Itoh Y, Tabata H, Nakajima K, Akiyama T, Masuyama N, et al. The Wnt/beta-catenin pathway directs neuronal differentiation of cortical neural precursor cells. Development 2004; 131:2791 - 2801
- Israsena N, Hu M, Fu W, Kan L, Kessler JA. The presence of FGF2 signaling determines whether beta-catenin exerts effects on proliferation or neuronal differentiation of neural stem cells. Dev Biol 2004; 268:220 - 231
- Lee HY, Kleber M, Hari L, Brault V, Suter U, Taketo M M, Kemler R, Sommer L. Instructive role of Wnt/beta-catenin in sensory fate specification in neural crest stem cells. Science 2004; 303:1020 - 1023
- Bertrand N, Castro DS, Guillemot F. Proneural genes and the specification of neural cell types. Nat Rev Neurosci 2002; 3:517 - 530
- Redmond L, Oh SR, Hicks C, Weinmaster G, Ghosh A. Nuclear Notch1 signaling and the regulation of dendritic development. Nat Neurosci 2000; 3:30 - 40
- Nishimoto M, Furuta A, Aoki S, Kudo Y, Miyakawa H, Wada K. PACAP/PAC1 autocrine system promotes proliferation and astrogenesis in neural progenitor cells. Glia 2007; 55:317 - 327
- Samanta J, Kessler JA. Interactions between ID and OLIG proteins mediate the inhibitory effects of BMP4 on oligodendroglial differentiation. Development 2004; 131:4131 - 4142
- Sommer L, Rao M. Neural stem cells and regulation of cell number. Prog Neurobiol 2002; 66:1 - 18
- Cau E, Casarosa S, Guillemot F. Mash1 and Ngn1 control distinct steps of determination and differentiation in the olfactory sensory neuron lineage. Development 2002; 129:1871 - 1880
- Parras CM, Hunt C, Sugimori M, Nakafuku M, Rowitch D, Guillemot F. The proneural gene Mash1 specifies an early population of telencephalic oligodendrocytes. J Neurosci 2007; 27:4233 - 4242
- Miyata T, Maeda T, Lee JE. NeuroD is required for differentiation of the granule cells in the cerebellum and hippocampus. Genes Dev 1999; 13:1647 - 1652
- Nieto M, Schuurmans C, Britz O, Guillemot F. Neural bHLH genes control the neuronal versus glial fate decision in cortical progenitors. Neuron 2001; 29:401 - 413
- Yoon K, Gaiano N. Notch signaling in the mammalian central nervous system: insights from mouse mutants. Nat Neurosci 2005; 8:709 - 715
- Nelson BR, Hartman BH, Georgi SA, Lan MS, Reh TA. Transient inactivation of Notch signaling synchronizes differentiation of neural progenitor cells. Dev Biol 2007; 304:479 - 498
- Nagao M, Sugimori M, Nakafuku M. Cross talk between notch and growth factor/cytokine signaling pathways in neural stem cells. Mol Cell Biol 2007; 27:3982 - 3994
- Bauer S, Kerr BJ, Patterson PH. The neuropoietic cytokine family in development, plasticity, disease and injury. Nat Rev Neurosci 2007; 8:221 - 232
- Bonni A, Sun Y, Nadal-Vicens M, Bhatt A, Frank DA, Rozovsky I, et al. Regulation of gliogenesis in the central nervous system by the JAK-STAT signaling pathway. Science 1997; 278:477 - 483
- Kamakura S, Oishi K, Yoshimatsu T, Nakafuku M, Masuyama N, Gotoh Y. Hes binding to STAT3 mediates crosstalk between Notch and JAK-STAT signalling. Nat Cell Biol 2004; 6:547 - 554
- Gaiano N, Fishell G. The role of notch in promoting glial and neural stem cell fates. Ann Rev Neurosci 2002; 25:471 - 490
- Fukuda S, Taga T. Roles of BMP in the development of the central nervous system. Clin Calcium 2006; 16:781 - 785
- See J, Mamontov P, Ahn K, Wine-Lee L, Crenshaw EB 3rd, Grinspan JB. BMP signaling mutant mice exhibit glial cell maturation defects. Mol Cell Neurosci 2007; 35:171 - 182
- Tokunaga A, Kohyama J, Yoshida T, Nakao K, Sawamoto K, Okano H. Mapping. Spatiotemporal activation of Notch signaling during neurogenesis and gliogenesis in the developing mouse brain. J Neurochem 2004; 90:142 - 154
- Lundkvist J, Lendahl U. Notch and the birth of glial cells. Trends Neurosci 2001; 24:492 - 494
- Kasai M, Satoh K, Akiyama T. Wnt signaling regulates the sequential onset of neurogenesis and gliogenesis via induction of BMPs. Genes Cells 2005; 10:777 - 783
- Ross SE, Greenberg ME, Stiles CD. Basic helix-loop-helix factors in cortical development. Neuron 2003; 39:13 - 25
- Sun Y, Nadal-Vicens M, Misono S, Lin MZ, Zubiaga A, Hua X, et al. Neurogenin promotes neurogenesis and inhibits glial differentiation by independent mechanisms. Cell 2001; 104:365 - 376
- Turnley AM, Faux CH, Rietze RL, Coonan JR, Bartlett PF. Suppressor of cytokine signaling 2 regulates neuronal differentiation by inhibiting growth hormone signaling. Nat Neurosci 2002; 5:1155 - 1162
- Guillemot F. Cell fate specification in the mammalian telencephalon. Prog Neurobiol 2007; 83:37 - 52
- Heinrich PC, Behrmann I, Müller-Newen G, Schaper F, Graeve L. Interleukin-6-type cytokine signalling through the gp130/Jak/STAT pathway. Biochem J 1998; 334:297 - 314
- Bonaguidi MA, McGuire T, Hu M, Kan L, Samanta J, Kessler JA. LIF and BMP signaling generate separate and discrete types of GFAP-expressing cells. Development 2005; 132:5503 - 5514
- Nakashima K, Yanagisawa M, Arakawa H, Kimura N, Hisatsune T, Kawabata M, et al. Synergistic signaling in fetal brain by STAT3-Smad1 complex bridged by p300. Science 1999; 284:479 - 482
- Gu F, Hata R, Ma YJ, Tanaka J, Mitsuda N, Kumon Y, et al. Suppression of Stat3 promotes neurogenesis in cultured neural stem cells. J Neurosci Res 2005; 81:163 - 171
- Ge W, Martinowich K, Wu X, He F, Miyamoto A, Fan G, et al. Notch signaling promotes astrogliogenesis via direct CSL-mediated glial gene activation. J Neurosci Res 2002; 69:848 - 860
- Takizawa T, Ochiai W, Nakashima K, Taga T. Enhanced gene activation by Notch and BMP signaling cross-talk. Nucleic Acids Res 2003; 31:5723 - 5731
- He F, Ge W, Martinowich K, Becker-Catania S, Coskun V, Zhu W, et al. A positive autoregulatory loop of Jak-STAT signaling controls the onset of astrogliogenesis. Nat Neurosci 2005; 8:616 - 625
- Richardson WD, Kessaris N, Pringle N. Oligodendrocyte wars. Nat Rev Neurosci 2006; 7:11 - 18
- Nery S, Wichterle H, Fishell G. Sonic hedgehog contributes to oligodendrocyte specification in the mammalian forebrain. Development 2001; 128:527 - 540
- Danesin C, Agius E, Escalas N, Ai XB, Emerson C, Cochard P, et al. Ventral neural progenitors switch toward an oligodendroglial fate in response to increased Sonic hedgehog (Shh) activity: involvement of Sulfatase 1 in modulating Shh signaling in the ventral spinal cord. J Neurosci 2006; 26:5037 - 5048
- Oh S, Huang X, Chiang C. Specific requirements of sonic hedgehog signaling during oligodendrocyte development. Dev Dyn 2005; 234:489 - 496
- Tekki-Kessaris N, Woodruff R, Hall AC, Gaffield W, Kimura S, Stiles CD, et al. Hedgehog-dependent oligodendrocyte lineage specification in the telencephalon. Development 2001; 128:2545 - 2554
- Kessaris N, Jamen F, Rubin LL, Richardson WD. Cooperation between sonic hedgehog and fibroblast growth factor/MAPK signalling pathways in neocortical precursors. Development 2004; 131:1289 - 1298
- Chandran S, Kato H, Gerreli D, Compston A, Svendsen CN, Allen ND. FGF-dependent generation of oligodendrocytes by a hedgehog-independent pathway. Development 2003; 130:6599 - 6609
- Gabay L, Lowell S, Rubin LL, Anderson DJ. Deregulation of dorsoventral patterning by FGF confers trilineage differentiation capacity on CNS stem cells in vitro. Neuron 2003; 40:485 - 499
- Gangemi RM, Perera M, Corte G. Regulatory genes controlling cell fate choice in embryonic and adult neural stem cells. J Neurochem 2004; 89:286 - 306
- Hsieh J, Aimone JB, Kaspar BK, Kuwabara T, Nakashima K, Gage FH. IGF-I instructs multipotent adult neural progenitor cells to become oligodendrocytes. J Cell Biol 2004; 164:111 - 122
- Rowitch DH. Glial specification in the vertebrate neural tube. Nat Rev Neurosci 2004; 5:409 - 419
- Wang S, Sdrulla AD, diSibio G, Bush G, Nofziger D, Hicks C, et al. Notch receptor activation inhibits oligodendrocyte differentiation. Neuron 1998; 21:63 - 75
- Mekki-Dauriac S, Agius E, Kan P, Cochard P. Development Bone morphogenetic proteins negatively control oligodendrocyte precursor specification in the chick spinal cord. Development 2002; 129:5117 - 5130
- Vallstedt A, Klos JM, Ericson J. Multiple dorsoventral origins of oligodendrocyte generation in the spinal cord and hindbrain. Neuron 2005; 45:55 - 67
- Zhou Q, Anderson DJ. The bHLH transcription factors OLIG2 and OLIG1 couple neuronal and glial subtype specification. Neuron 2002; 109:61 - 73
- Nicolay DJ, Doucette JR, Nazarali AJ. Hoxb4 in oligodendrogenesis. Cell Mol Neurobiol 2004; 24:357 - 366
- Sun T, Echelard Y, Lu R, Yuk DI, Kaing S, Stiles CD, et al. Olig bHLH proteins interact with homeodomain proteins to regulate cell fate acquisition in progenitors of the ventral neural tube. Curr Biol 2001; 11:1413 - 1420
- Du ZW, Li XJ. Induced expression of Olig2 is sufficient for oligodendrocyte specification but not for motoneuron specification and astrocyte repression. Mol Cell Neurosci 2006; 33:371 - 380
- Kessaris N, Pringle N, Richardson WD. Specification of CNS glia from neural stem cells in the embryonic neuroepithelium. Philos Trans R Soc Lond B Biol Sci 2008; 363:71 - 85
- Xin M, Yue T, Ma Z, Wu FF, Gow A, Lu QR. Myelinogenesis and axonal recognition by oligodendrocytes in brain are uncoupled in Olig1-null mice. J Neurosci 2005; 25:1354 - 1365
- Cai J, St Amand T, Yin H, Guo H, Li G, Zhang Y, et al. Expression and regulation of the chicken Nkx-6.2 homeobox gene suggest its possible involvement in the ventral neural patterning and cell fate specification. Dev Dyn 1999; 216:459 - 468
- Pattyn A, Vallstedt A, Dias JM, Sander M, Ericson J. Complementary roles for Nkx6 and Nkx2 class proteins in the establishment of motoneuron identity in the hindbrain. Development 2003; 130:4149 - 4159
- Wegner M. A matter of identity: transcriptional control in oligodendrocytes. J Mol Neurosci 2008; 35:3 - 12
- Southwood C, He C, Garbern J, Kamholz J, Arroyo E, Gow A. CNS myelin paranodes require Nkx6-2 homeoprotein transcriptional activity for normal structure. J Neurosci 2004; 24:11215 - 11225
- Qi Y, Cai J, Wu Y, Wu R, Lee J, Fu H, et al. Control of oligodendrocyte differentiation by the Nkx2.2 homeodomain transcription factor. Development 2001; 128:2723 - 2733
- Sussman CR, Dyer KL, Marchionni M, Miller RH. Local control of oligodendrocyte development in isolated dorsal mouse spinal cord. J Neurosci Res 2000; 59:413 - 420
- Stolt CC, Lommes P, Friedrich RP, Wegner M. Transcription factors Sox8 and Sox10 perform non-equivalent roles during oligodendrocyte development despite functional redundancy. Development 2004; 131:2349 - 2358
- Cheung M, Briscoe J. Neural crest development is regulated by the transcription factor Sox9. Development 2003; 130:5681 - 5693
- Stolt CC, Lommes P, Sock E, Chaboissier MC, Schedl A, Wegner M. The Sox9 transcription factor determines glial fate choice in the developing spinal cord. Genes Dev 2003; 17:1677 - 1689
- Stolt CC, Schmitt S, Lommes P, Sock E, Wegner M. Impact oftranscription factor Sox8 on oligodendrocyte specification in the mouseembryonic spinal cord. Dev Biol 2005; 281:309 - 317
- Stolt CC, Rehberg S, Ader M, Lommes P, Riethmacher D, Schachner M, et al. Terminal differentiation of myelin-forming oligodendrocytes depends on the transcription factor Sox10. Genes Dev 2002; 16:165 - 170
- Liu Z, Hu X, Cai J, Liu B, Peng X, Wegner M, Qiu M. Induction of oligodendrocyte differentiation by Olig2 and Sox10: evidence for reciprocal interactions and dosage-dependent mechanisms. Develop Biol 2007; 302:683 - 693
- De Calisto J, Araya C, Marchant L, Riaz CF, Mayor R. Essential role of non-canonical Wnt signalling in neural crest migration. Development 2005; 132:2587 - 2597
- Ille F, Sommer L. Wnt signaling: multiple functions in neural development. Cell Mol Life Sci 2005; 62:1100 - 1108
- Wohrle S, Wallmen B, Hecht A. Differential control of Wnt target genes involves epigenetic mechanisms and selective promoter occupancy by T-cell factors. Mol Cell Biol 2007; 27:8164 - 8177
- Wen S, Li H, Liu J. Epigenetic background of neuronal fate determination. Prog Neurobiol 2009; 87:98 - 117
- Takizawa T, Nakashima K, Namihira M, Ochiai W, Uemura A, Yanagisawa M, et al. DNA methylation is a critical cell-intrinsic determinant of astrocyte differentiation in the fetal brain. Deve Cell 2001; 1:749 - 758
- Shimozaki K, Namihira M, Nakashima K, Taga T. Stage- and site-specific DNA demethylation during neural cell development from embryonic stem cells. J Neurochem 2005; 93:432 - 439
- Fan G, Martinowich K, Chin MH, He F, Fouse SD, Hutnick L, et al. DNA methylation controls the timing of astrogliogenesis through regulation of JAK-STAT signaling. Development 2005; 132:3345 - 3356
- Lachner M, O'Sullivan RJ, Jenuwein T. An epigenetic road map for histone lysine methylation. J Cell Sci 2003; 116:2117 - 2124
- Fukuda S, Taga T. Cell fate determination regulated by a transcriptional signal network in the developing mouse brain. Anat Sci Intl 2005; 80:12 - 18 2005