Abstract
Telomere maintenance in cycling cells relies on both DNA replication and capping by the protein complex shelterin. Two single-stranded DNA (ssDNA)-binding proteins, replication protein A (RPA) and protection of telomere 1 (POT1) play critical roles in DNA replication and telomere capping, respectively. While RPA binds to ssDNA in a non-sequence-specific manner, POT1 specifically recognizes singlestranded TTAGGG telomeric repeats. Loss of POT1 leads to aberrant accumulation of RPA at telomeres and activation of the ataxia telangiectasia and Rad3-related kinase (ATR)-mediated checkpoint response, suggesting that POT1 antagonizes RPA binding to telomeric ssDNA. The requirement for both POT1 and RPA in telomere maintenance and the antagonism between the two proteins raises the important question of how they function in concert on telomeric ssDNA. Two interesting models were proposed by recent studies to explain the regulation of POT1 and RPA at telomeres. Here, we discuss how these models help unravel the coordination, and also the antagonism, between POT1 and RPA during the cell cycle.
Telomeres, the natural ends of chromosomes, are composed of repetitive DNA sequences and “capped” by both specific proteins and non-coding RNAs.Citation1–Citation3 One of the critical functions of telomeres is to prevent chromosomal ends from recognition by the DNA damage response machinery. Critically short or improperly capped telomeres lead to telomere dysfunction and are a major source of genomic instability.Citation4 While telomeres need to be properly capped to remain stable, they also need to be duplicated during each cell division by the DNA replication machinery. The requirement of these two seemingly competing processes for telomere maintenance suggests that the cell must coordinate DNA replication and capping of telomeres to ensure faithful telomere duplication yet avoid an inappropriate DNA damage response.
Telomeric DNA is unique in several ways. The bulk of each human telomere is comprised of double-stranded TTA GGG repeats. At the very end of each telomere, a stretch of single-stranded TTAGGG repeats exists as a 3′ overhang. The TTA GGG repeats in the telomeric single-stranded DNA (ssDNA) allow it to loop back and invade telomeric double-stranded DNA (dsDNA), forming a structure called the t-loop.Citation5 At the base of the t-loop, the TTAGGG strand of the telomeric dsDNA is displaced by the invading single-stranded 3′ overhang to form a single-stranded D-loop. Thus, the unique DNA sequence and structures of telomeres confer the ability to bind proteins in both sequence- and structure-specific manners, providing the basis for additional regulations.
In human cells, telomere capping is orchestrated by the protein complex shelterin, which contains TRF1, TRF2, RAP1, TIN2, TPP1 and POT1.Citation3 Among these shelterin components, TRF1 and TRF2 interact with telomeric dsDNA in a sequence-specific manner, whereas POT1, in a complex with TPP1, binds to telomeric ssDNA in a sequence-specific manner.Citation6–Citation8 While the human genome contains only one POT1 gene, the mouse genome contains two POT1-related genes, POT1a and POT1b.Citation9–Citation11 TIN2 functions to stabilize TRF1 and TRF2 DNA binding and also tethers the POT1-TPP1 heterodimer to the rest of the shelterin complex on telomeric dsDNA.Citation12,Citation13
Unlike the properly capped telomeres, double-stranded DNA breaks (DSBs) with ssDNA overhangs are known to activate the ATR checkpoint kinase.Citation14,Citation15 In a complex with its functional partner ATRIP, ATR is recruited to ssDNA by RPA, a non-sequence-specific ssDNA-binding protein complex.Citation16 In addition to the ATR-ATRIP kinase complex, several other checkpoint proteins involved in ATR activation are also recruited in the presence of RPA-ssDNA.Citation15 The structural resemblance between DSBs and telomeres and the presence of ssDNA at telomeres raise the important question as to how ATR activation is repressed at telomeres.
POT1 is a Critical Repressor of ATR Activation at Telomeres
One important clue to the mechanism of ATR repression at telomeres came from the studies on mouse cells lacking POT1a. When POT1a was conditionally deleted from mouse cells, telomere capping was compromised, leading to activation of the ATR checkpoint.Citation10,Citation17,Citation18 Furthermore, upon POT1a depletion, increased amounts of RPA accumulated at telomeres, suggesting that POT1a antagonizes the binding of RPA to telomeric ssDNA.Citation19 Similar to mouse cells, knockdown of POT1 in human cells also resulted in accumulation of RPA at telomeres.Citation20,Citation21 Together, these studies convincingly showed that, POT1, which binds to telomeric ssDNA in a sequence-specific manner, represses ATR activation in vivo by antagonizing the non-sequence-specific binding of RPA to telomeric ssDNA.
Although it is clear that POT1 antagonizes the binding of RPA to telomeres and represses ATR activation in vivo, exactly how POT1 executes this role is less clear. Two recent studies by Takai et al. and Flynn et al. showed that in vitro RPA bound to telomeric ssDNA more efficiently than, or at least as efficiently as, the POT1-TPP1 complex.Citation21,Citation22 Furthermore, we showed that when RPA and POT1-TPP1 were both present at similar concentrations, RPA outcompeted POT1-TPP1 for binding to telomeric ssDNA.Citation21 In addition to the relative affinity of RPA and POT1 to telomeric ssDNA, their relative abundance in vivo is another important factor that influences their binding to telomeric ssDNA. While RPA is known to be an abundant protein in cells,Citation23,Citation24 the abundance of POT1 is quite low.Citation25 Takei et al. estimated that RPA and POT1 are present at 3–5 × 106 and 1–2 × 104 molecules/cell, respectively.Citation22 Taken together, these results suggest that POT1 has to rely on additional regulatory mechanisms in vivo to antagonize RPA binding to telomeric ssDNA.
In addition to its function in ATR activation, RPA is also a critical component of DNA replication machinery.Citation26,Citation27 When DNA replication forks travel through telomeres, RPA likely interacts with singlestranded TTAGGG repeats to support DNA synthesis. Indeed, in both yeast and humans, RPA was found to transiently associate with telomeres during S phase.Citation28–Citation31 Furthermore, genetic studies in yeast have clearly implicated RPA in telomere maintenance.Citation32–Citation36 While persistent activation of ATR at telomeres would likely trigger cell cycle arrest, senescence or apoptosis, ATR is also known to play a positive role in telomere maintenance. In mouse cells, ATR is required for suppression of telomere fragility and recombination and promotes elongation of short telomeres by telomerase.Citation29,Citation37,Citation38 Like RPA, ATR and its yeast homolog Mec1 were both found to bind telomeres transiently during the cell cycle.Citation29,Citation31 These findings suggest that the competition between RPA and POT1 on telomeric ssDNA may not be a simple one. Furthermore, the interplay between RPA and POT1 may be important for the coordination of telomere replication and telomere capping during the cell cycle.
Tethering POT1 to Telomeres
POT1 binds to telomeric ssDNA directly via its OB folds.Citation10,Citation11,Citation39,Citation40 However, although the OB folds of POT1 are important for the function of POT1 in telomere end protection,Citation20,Citation41,Citation42 they are neither necessary nor sufficient for the localization of POT1 to telomeres.Citation8,Citation20,Citation43 Indeed, while a point mutation in the OB fold abolished POT1a binding to telomeric ssDNA in vitro, this mutant efficiently localized to telomeres in vivo.Citation10 These results suggest that POT1 needs to be recruited to telomeres by another mechanism to enable its direct binding to telomeric ssDNA. In both human and mouse, the abilities of POT1 and POT1a/b to bind TPP1 are critical for their functions in telomere protection.Citation8,Citation13,Citation42–Citation44 A POT1 mutant lacking the TPP1-binding domain failed to localize to telomeres.Citation8 Furthermore, a POT1 mutant unable to bind TPP1 failed to prevent RPA accumulation at telomeres,Citation20 suggesting that the role of POT1 in antagonizing RPA binding is dependent upon its interaction with TPP1.
TPP1 contributes to the function of POT1 at telomeres in at least two distinct ways. First, TPP1 enhances the binding of POT1 to telomeric ssDNA.Citation6,Citation8 However, given that RPA binds to telomeric ssDNA more efficiently than the POT1-TPP1 complex in vitro,Citation21 this function of TPP1 alone is insufficient to fully explain the requirement of POT1-TPP1 interaction for RPA exclusion from telomeres. Second, recent observations suggest that the tethering of POT1-TPP1 by TIN2 is important for its function in antagonizing RPA binding at telomeres.Citation22 Deletion of TIN2 from mouse cells compromised the localization of POT1 and TPP1 to telomeres.Citation22 Furthermore, like deletion of POT1a and TPP1, deletion of TIN2 also resulted in RPA accumulation at telomeres and ATR activation.Citation22 Consistent with the idea that POT1-TPP1 needs to be tethered by TIN2 to antagonize RPA binding, we found that overexpression of the TPP1ΔRD mutant,Citation8 which competes with endogenous TPP1 for TIN2 binding but is unable to bind POT1, induced RPA foci at telomeres (). Thus, by increasing the local concentration of POT1-TPP1 at telomeres, TIN2 plays a critical role in the repression of ATR at telomeres. Quantification of chromatin-bound shelterin components suggested that TRF1, TRF2, RAP1 and TIN2 are much more abundant than POT1 and TTP1 on DNA.Citation25 It is plausible that TRF1, TRF2, RAP1 and TIN2 spread across the telomeric dsDNA regions, whereas the POT1-TPP1 complex may associate with both telomeric dsDNA and ssDNA through TIN2 and the OB folds of POT1, respectively. However, although TIN2 may bring POT1-TPP1 to the vicinity of telomeric ssDNA and enhance its ability to antagonize RPA binding, this mechanism alone does not adequately explain how RPA and ATR can access telomeres during the cell cycle and execute their functions during DNA replication.
A Cell Cycle-Regulated RPA-to-POT1 Switch on Telomeric ssDNA
To understand how RPA and POT1 can both function at telomeres, we analyzed how these proteins associated with telomeric ssDNA in human cell extracts.Citation21 To our surprise, we found that in extracts, RPA, but not POT1 was actively displaced from telomeric ssDNA in a DNA sequence-specific manner. Using a biochemical approach, we traced the RPA-displacing activity to the heterogeneous nuclear ribonucleoproteins (hnRNPs) A1 and A2/B1, which were known to bind telomeric ssDNA in vitro and associate with telomeres in vivo.Citation45–Citation48 In vitro, purified hnRNP A1 recapitulated the DNA sequence- and protein-specific RPA displacement from telomeric ssDNA.Citation21
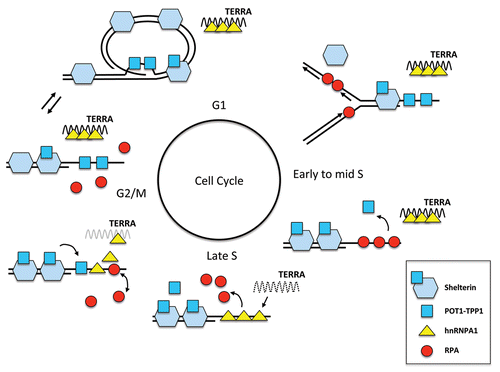
Importantly, the telomere-specific RPA-displacing activity is cell cycle-regulated. This activity is low during early to mid S phase, but is upregulated in late S phase. Interestingly, the cell cycle fluctuation of RPA-displacing activity inversely correlates with the presence of the noncoding RNA TERRA (telomere repeatcontaining RNA) at telomeres. TERRA associates with telomeres during early to mid S phase, disappears in late S phase and reaccumulates in G2.Citation49 In vitro, TERRA binds to hnRNPs and is a potent inhibitor of the RPA-displacing activity, suggesting that TERRA may inhibit RPA displacement during S phase by sequestering hnRNPs. Furthermore, the decline of TERRA in late S phase may provide a window for hnRNPs to displace RPA from telomeric ssDNA.Citation21
In vitro, TERRA not only inhibits the displacement of RPA by hnRNP A1, but also removes hnRNP A1 from telomeric ssDNA. When TERRA reaccumulates in G2 cells, it may gradually sequester hnRNPs away from telomeric ssDNA. During this transition period, hnRNPs may shuttle between telomeric ssDNA and TERRA, and both POT1 and RPA may gain access to telomeric ssDNA. However, because RPA would be specifically displaced by hnRNPs before hnRNPs are fully sequestered by TERRA, only POT1 but not RPA would remain stably bound to telomeric ssDNA. Thus, this dynamic process would eventually promote an RPA-to-POT1 switch on telomeric ssDNA.Citation21
Coordination of Telomere Replication and Telomere Capping
The model above provides a possible explanation for how DNA replication and capping of telomeres are coordinated during the cell cycle. During early and mid S phase, hnRNPs are sequestered by TERRA at telomeres, allowing RPA to outcompete POT1 for binding to single-stranded TTAGGG repeats at the replication forks traveling through telomeres (). In the presence of TERRA, RPA may bind to 3′ telomere overhangs upon the arrival of DNA replication forks, during resection of telomere ends, or during telomere extension by telomerase. When TERRA declines in late S phase, hnRNPs are unleashed to displace RPA from telomeric ssDNA (). In G2, as TERRA reaccumulates at telomeres, hnRNPs gradually move from telomeric ssDNA to TERRA, promoting the occupancy of POT1 on telomeric ssDNA and completing the capping of telomeres ().
In contrast to RPA, POT1 remains stably bound to telomeric ssDNA in cell extracts, suggesting that once POT1 is loaded onto telomeric ssDNA, it cannot be easily removed by RPA or hnRNPs.Citation21 Although POT1 was detected at telomeres throughout the cell cycle,Citation37,Citation50 its specific associations with telomeric ssDNA and dsDNA-bound shelterin are difficult to discern. It is possible that once POT1 binds to telomeric ssDNA in G2, it remains bound until the arrival of DNA replication machinery in the next S phase (). In this scenario, even during S phase, 3′ telomere overhangs are still occupied by POT1 until the arrival of DNA replication machinery. It is also possible that the lagging-strand DNA polymerases at replication forks are unable to reach the ends of 3′ telomere overhangs, leaving some POT1 on telomeric ssDNA throughout S phase. Both of these possibilities would explain why rapid depletion of POT1a in S-phase mouse cells resulted in RPA accumulation at telomeres.Citation19
In cells outside of S phase, POT1 is also important for telomere protection. Rapid degradation of POT1a in mouse cells led to RPA accumulation at telomeres and ATR activation in both G1 and S/G2.Citation19 Interestingly, upon POT1a depletion, RPA was more readily detected at telomeres in S/G2 than in G1,Citation19 suggesting that either the ability of POT1a to antagonize RPA binding or the structure of telomeres alters between the two cell cycle phases.Citation14 When human cells were synchronized in S phase and POT1 levels were gradually reduced by siRNA, RPA accumulation at telomeres was not evident. However, when cells with reduced levels of POT1 were synchronously released into G2, RPA foci were readily detected at telomeres.Citation21 Similarly, when the tethering of POT1-TPP1 by TIN2 was compromised by the TPP1ΔRD mutant, RPA accumulated at telomeres as cells progress from S phase to G2 (). These results suggest that while low levels of POT1 may be sufficient to maintain telomere capping in S phase, POT1 needs to be present at a higher concentration to re-establish telomere capping after DNA replication.
More than One Way to Regulate POT1
The two models proposed to explain how POT1 antagonizes RPA binding at telomeres are not mutually exclusive. The tethering model of POT1 emphasizes that the local concentration of POT1-TPP1 at telomeres is critical for its ability to compete with RPA.Citation22 On the other hand, the RPA-to-POT1 switch model provides an explanation for how RPA can access single-stranded TTAGGG repeats during DNA replication without compromising telomere capping.Citation21 The cell cycle-regulated fluctuation of TERRA and sequestration of hnRNPs at telomeres present a window for RPA to transiently bind telomeric ssDNA and a window for POT1 to stably cap telomere ends. This RPA-to-POT1 switch may only occur when POT1-TPP1 is tethered to telomeric dsDNA by TIN2 and the local concentration of POT1 at telomeres is sufficiently high.
While both models offer attractive explanations for the regulation of RPA and POT1 at telomeres, they each raise a set of new questions. For example, it is still unclear how the POT1-TPP1 complexes tethered by TIN2 distribute on telomeric dsDNA and how they engage 3′ telomere overhangs. How hnRNPs displace RPA from telomeric ssDNA and how POT1 resists this displacement are also unknown. In addition to the two models discussed above, there may be other mechanisms that contribute to the regulation of RPA and POT1 at telomeres. A better understanding of the concerted action of RPA, POT1, and their regulators in telomere maintenance requires further investigations of their biochemical properties, their cell cycle dynamics, and their abundance and distribution at telomeres.
Figures and Tables
Figure 1 Expression of the TPP1ΔRD mutant leads to RPA accumulation at telomeres. HeLa cells were transfected with plasmids expressing wild-type TPP1, the TPP1ΔRD mutant or mock treated. Cells were synchronized in S phase with thymidine and subsequently released from the thymidine block. As the majority of cells entering G2 (8 h after thymidine release), they were analyzed by immunofluorescence staining with RPA32 and TRF2 antibodies.
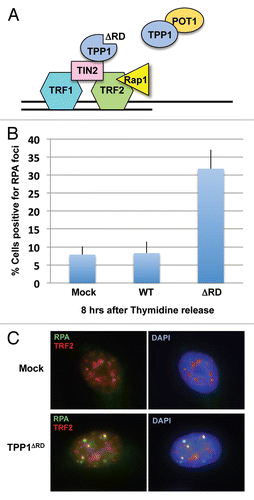
Figure 2 A model for coordinated functions of RPA and POT1 at telomeres. Tethering of POT1-TPP1 by TI N2 to the shelterin bound to telomeric dsDNA is required for POT1 to reach a critical concentration at telomeres. The cell cycle-regulated RPA-to-POT1 switch on telomeric ssDNA orchestrated by TERRA and hnRNPs enables RPA to support DNA replication of telomeres without compromising telomere capping.
Acknowledgements
R.L.F. is supported the ACS fellowship 0902501. S.C. is supported by the NIH grant CA129037. L.Z. is supported by the NIH grant GM076388. L.Z. is the Jim and Ann Orr MGH Research Scholar and a Scholar of the Ellison Medical Foundation.
References
- Azzalin CM, Reichenbach P, Khoriauli L, Giulotto E, Lingner J. Telomeric repeat containing RNA and RNA surveillance factors at mammalian chromosome ends. Science 2007; 318:798 - 801; PMID: 17916692; http://dx.doi.org/10.1126/science.1147182
- Schoeftner S, Blasco MA. Developmentally regulated transcription of mammalian telomeres by DNA-dependent RNA polymerase II. Nat Cell Biol 2008; 10:228 - 236; PMID: 18157120; http://dx.doi.org/10.1038/ncb1685
- Palm W, de Lange T. How shelterin protects mammalian telomeres. Annu Rev Genet 2008; 42:301 - 334; PMID: 18680434; http://dx.doi.org/10.1146/annurev.genet.41.110306.130350
- Artandi SE, DePinho RA. Telomeres and telomerase in cancer. Carcinogenesis 2010; 31:9 - 18; PMID: 19887512; http://dx.doi.org/10.1093/carcin/bgp268
- Griffith JD, Comeau L, Rosenfield S, Stansel RM, Bianchi A, Moss H, et al. Mammalian telomeres end in a large duplex loop. Cell 1999; 97:503 - 514; PMID: 10338214; http://dx.doi.org/10.1016/S0092-8674(00)80760-6
- Wang F, Podell ER, Zaug AJ, Yang Y, Baciu P, Cech TR, et al. The POT1-TPP1 telomere complex is a telomerase processivity factor. Nature 2007; 445:506 - 510; PMID: 17237768; http://dx.doi.org/10.1038/nature05454
- Baumann P, Cech TR. Pot1, the putative telomere end-binding protein in fission yeast and humans. Science 2001; 292:1171 - 1175; PMID: 11349150; http://dx.doi.org/10.1126/science.1060036
- Xin H, Liu D, Wan M, Safari A, Kim H, Sun W, et al. TPP1 is a homologue of ciliate TEBP-beta and interacts with POT1 to recruit telomerase. Nature 2007; 445:559 - 562; PMID: 17237767; http://dx.doi.org/10.1038/nature05469
- Hockemeyer D, Daniels JP, Takai H, de Lange T. Recent expansion of the telomeric complex in rodents: Two distinct POT1 proteins protect mouse telomeres. Cell 2006; 126:63 - 77; PMID: 16839877; http://dx.doi.org/10.1016/j.cell.2006.04.044
- Wu L, Multani AS, He H, Cosme-Blanco W, Deng Y, Deng JM, et al. Pot1 deficiency initiates DNA damage checkpoint activation and aberrant homologous recombination at telomeres. Cell 2006; 126:49 - 62; PMID: 16839876; http://dx.doi.org/10.1016/j.cell.2006.05.037
- He H, Multani AS, Cosme-Blanco W, Tahara H, Ma J, Pathak S, et al. POT1b protects telomeres from end-to-end chromosomal fusions and aberrant homologous recombination. EMBO J 2006; 25:5180 - 5190; PMID: 17053789; http://dx.doi.org/10.1038/sj.emboj.7601294
- Ye JZ, Donigian JR, van Overbeek M, Loayza D, Luo Y, Krutchinsky AN, et al. TIN2 binds TRF1 and TRF2 simultaneously and stabilizes the TRF2 complex on telomeres. J Biol Chem 2004; 279:47264 - 47271; PMID: 15316005; http://dx.doi.org/10.1074/jbc.M409047200
- Ye JZ, Hockemeyer D, Krutchinsky AN, Loayza D, Hooper SM, Chait BT, et al. POT1-interacting protein PIP1: a telomere length regulator that recruits POT1 to the TIN2/TRF1 complex. Genes Dev 2004; 18:1649 - 1654; PMID: 15231715; http://dx.doi.org/10.1101/gad.1215404
- Shiotani B, Zou L. Single-stranded DNA orchestrates an ATM-to-ATR switch at DNA breaks. Mol Cell 2009; 33:547 - 558; PMID: 19285939; http://dx.doi.org/10.1016/j.molcel.2009.01.024
- Flynn RL, Zou L. ATR: a master conductor of cellular responses to DNA replication stress. Trends Biochem Sci 2011; 36:133 - 140; PMID: 20947357; http://dx.doi.org/10.1016/j.tibs.2010.09.005
- Zou L, Elledge SJ. Sensing DNA damage through ATRIP recognition of RPA-ssDNA complexes. Science 2003; 300:1542 - 1548; PMID: 12791985; http://dx.doi.org/10.1126/science.1083430
- Denchi EL, de Lange T. Protection of telomeres through independent control of ATM and ATR by TRF2 and POT1. Nature 2007; 448:1068 - 1071; PMID: 17687332; http://dx.doi.org/10.1038/nature06065
- Guo X, Deng Y, Lin Y, Cosme-Blanco W, Chan S, He H, et al. Dysfunctional telomeres activate an ATM-ATR-dependent DNA damage response to suppress tumorigenesis. EMBO J 2007; 26:4709 - 4719; PMID: 17948054; http://dx.doi.org/10.1038/sj.emboj.7601893
- Gong Y, de Lange TA. A Shld1-controlled POT1a provides support for repression of ATR signaling at telomeres through RPA exclusion. Mol Cell 2010; 40:377 - 387; PMID: 21070964; http://dx.doi.org/10.1016/j.molcel.2010.10.016
- Barrientos KS, Kendellen MF, Freibaum BD, Armbruster BN, Etheridge KT, Counter CM. Distinct functions of POT1 at telomeres. Mol Cell Biol 2008; 28:5251 - 5264; PMID: 18519588; http://dx.doi.org/10.1128/MCB.00048-08
- Flynn RL, Centore RC, O'Sullivan RJ, Rai R, Tse A, Songyang Z, et al. TERRA and hnRNPA1 orchestrate an RPA-to-POT1 switch on telomeric single-stranded DNA. Nature 2011; 471:532 - 536; PMID: 21399625; http://dx.doi.org/10.1038/nature09772
- Takai KK, Kibe T, Donigian JR, Frescas D, de Lange T. Telomere protection by TPP1/POT1 requires tethering to TIN2. Mol Cell 2011; 44:647 - 659; PMID: 22099311; http://dx.doi.org/10.1016/j.molcel.2011.08.043
- Seroussi E, Lavi S. Replication protein A is the major single-stranded DNA binding protein detected in mammalian cell extracts by gel retardation assays and UV cross-linking of long and short single-stranded DNA molecules. J Biol Chem 1993; 268:7147 - 7154; PMID: 8463251
- Kenny MK, Schlegel U, Furneaux H, Hurwitz J. The role of human single-stranded DNA binding protein and its individual subunits in simian virus 40 DNA replication. J Biol Chem 1990; 265:7693 - 7700; PMID: 2159011
- Takai KK, Hooper S, Blackwood S, Gandhi R, de Lange T. In vivo stoichiometry of shelterin components. J Biol Chem 2010; 285:1457 - 1467; PMID: 19864690; http://dx.doi.org/10.1074/jbc.M109.038026
- Wold MS. Replication protein A: a heterotrimeric, single-stranded DNA-binding protein required for eukaryotic DNA metabolism. Annu Rev Biochem 1997; 66:61 - 92; PMID: 9242902; http://dx.doi.org/10.1146/annurev.biochem.66.1.61
- Fanning E, Klimovich V, Nager AR. A dynamic model for replication protein A (RPA) function in DNA processing pathways. Nucleic Acids Res 2006; 34:4126 - 4137; PMID: 16935876; http://dx.doi.org/10.1093/nar/gkl550
- Déjardin J, Kingston RE. Purification of proteins associated with specific genomic Loci. Cell 2009; 136:175 - 186; PMID: 19135898; http://dx.doi.org/10.1016/j.cell.2008.11.045
- Verdun RE, Karlseder J. Replication and protection of telomeres. Nature 2007; 447:924 - 931; PMID: 17581575; http://dx.doi.org/10.1038/nature05976
- Moser BA, Subramanian L, Chang YT, Noguchi C, Noguchi E, Nakamura TM. Differential arrival of leading and lagging strand DNA polymerases at fission yeast telomeres. EMBO J 2009; 28:810 - 820; PMID: 19214192; http://dx.doi.org/10.1038/emboj.2009.31
- McGee JS, Phillips JA, Chan A, Sabourin M, Paeschke K, Zakian VA. Reduced Rif2 and lack of Mec1 target short telomeres for elongation rather than double-strand break repair. Nat Struct Mol Biol 2010; 17:1438 - 1445; PMID: 21057524; http://dx.doi.org/10.1038/nsmb.1947
- Smith J, Zou H, Rothstein R. Characterization of genetic interactions with RFA1: the role of RPA in DNA replication and telomere maintenance. Biochimie 2000; 82:71 - 78; PMID: 10717390; http://dx.doi.org/10.1016/S0300-9084(00)00183-8
- Ono Y, Tomita K, Matsuura A, Nakagawa T, Masukata H, Uritani M, et al. A novel allele of fission yeast rad11 that causes defects in DNA repair and telomere length regulation. Nucleic Acids Res 2003; 31:7141 - 7149; PMID: 14654689; http://dx.doi.org/10.1093/nar/gkg917
- Schramke V, Luciano P, Brevet V, Guillot S, Corda Y, Longhese MP, et al. RPA regulates telomerase action by providing Est1p access to chromosome ends. Nat Genet 2004; 36:46 - 54; PMID: 14702040; http://dx.doi.org/10.1038/ng1284
- Kibe T, Ono Y, Sato K, Ueno M. Fission yeast Taz1 and RPA are synergistically required to prevent rapid telomere loss. Mol Biol Cell 2007; 18:2378 - 2387; PMID: 17429064; http://dx.doi.org/10.1091/mbc.E06-12-1084
- Chen C, Umezu K, Kolodner RD. Chromosomal rearrangements occur in S. cerevisiae rfa1 mutator mutants due to mutagenic lesions processed by double-strand-break repair. Mol Cell 1998; 2:9 - 22; PMID: 9702187; http://dx.doi.org/10.1016/S1097-2765(00)80109-4
- Verdun RE, Crabbe L, Haggblom C, Karlseder J. Functional human telomeres are recognized as DNA damage in G2 of the cell cycle. Mol Cell 2005; 20:551 - 561; PMID: 16307919; http://dx.doi.org/10.1016/j.molcel.2005.09.024
- McNees CJ, Tejera AM, Martínez P, Murga M, Mulero F, Fernandez-Capetillo O, et al. ATR suppresses telomere fragility and recombination but is dispensable for elongation of short telomeres by telomerase. J Cell Biol 2010; 188:639 - 652; PMID: 20212315; http://dx.doi.org/10.1083/jcb.200908136
- Lei M, Podell ER, Cech TR. Structure of human POT1 bound to telomeric single-stranded DNA provides a model for chromosome end-protection. Nat Struct Mol Biol 2004; 11:1223 - 1229; PMID: 15558049; http://dx.doi.org/10.1038/nsmb867
- Loayza D, Parsons H, Donigian J, Hoke K, de Lange T. DNA binding features of human POT1: a nonamer 5′-TAG GGT TAG-3′ minimal binding site, sequence specificity and internal binding to multimeric sites. J Biol Chem 2004; 279:13241 - 13248; PMID: 14715659; http://dx.doi.org/10.1074/jbc.M312309200
- Loayza D, De Lange T. POT1 as a terminal transducer of TRF1 telomere length control. Nature 2003; 423:1013 - 1018; PMID: 12768206; http://dx.doi.org/10.1038/nature01688
- Palm W, Hockemeyer D, Kibe T, de Lange T. Functional dissection of human and mouse POT1 proteins. Mol Cell Biol 2009; 29:471 - 482; PMID: 18955498; http://dx.doi.org/10.1128/MCB.01352-08
- Hockemeyer D, Palm W, Else T, Daniels JP, Takai KK, Ye JZ, et al. Telomere protection by mammalian Pot1 requires interaction with Tpp1. Nat Struct Mol Biol 2007; 14:754 - 761; PMID: 17632522; http://dx.doi.org/10.1038/nsmb1270
- Liu D, Safari A, O'Connor MS, Chan DW, Laegeler A, Qin J, et al. PTOP interacts with POT1 and regulates its localization to telomeres. Nat Cell Biol 2004; 6:673 - 680; PMID: 15181449; http://dx.doi.org/10.1038/ncb1142
- McKay SJ, Cooke H. hnRNP A2/B1 binds specifically to single stranded vertebrate telomeric repeat TTA GGGn. Nucleic Acids Res 1992; 20:6461 - 6464; PMID: 1282701; http://dx.doi.org/10.1093/nar/20.24.6461
- Ishikawa F, Matunis MJ, Dreyfuss G, Cech TR. Nuclear proteins that bind the pre-mRNA 3′ splice site sequence r(UUAG/G) and the human telomeric DNA sequence d(TTA GGG)n. Mol Cell Biol 1993; 13:4301 - 4310; PMID: 8321232
- LaBranche H, Dupuis S, Ben-David Y, Bani MR, Wellinger RJ, Chabot B. Telomere elongation by hnRNP A1 and a derivative that interacts with telomeric repeats and telomerase. Nat Genet 1998; 19:199 - 202; PMID: 9620782; http://dx.doi.org/10.1038/575
- Zhang QS, Manche L, Xu RM, Krainer AR. hnRNP A1 associates with telomere ends and stimulates telomerase activity. RNA 2006; 12:1116 - 1128; PMID: 16603717; http://dx.doi.org/10.1261/rna.58806
- Porro A, Feuerhahn S, Reichenbach P, Lingner J. Molecular dissection of telomeric repeat-containing RNA biogenesis unveils the presence of distinct and multiple regulatory pathways. Mol Cell Biol 2010; 30:4808 - 4817; PMID: 20713443; http://dx.doi.org/10.1128/MCB.00460-10
- Wei C, Price CM. Cell cycle localization, dimerization and binding domain architecture of the telomere protein cPot1. Mol Cell Biol 2004; 24:2091 - 2102; PMID: 14966288; http://dx.doi.org/10.1128/MCB.24.5.2091-102.2004