Abstract
Epidemiological data suggest an important role of vitamin D signaling in cancer development and progression, and experimental studies demonstrate that the active vitamin D metabolite 1α, 25-dihydroxyvitamin D₃ (1,25D₃) has broad spectrum antitumor activity. Hypercalcemia has often been suggested to limit the clinical application of these data. The 14-epi-analog of 1,25D₃, inecalcitol [19-nor-14-epi-23-yne-1,25-(OH)₂D₃; TX522], was developed to have superagonistic antitumor activities but low hypercalcemia potential. We examined the antitumor activity of inecalcitol and the underlying mechanisms in a murine squamous cell carcinoma (SCC) model system. In vitro, compared with 1,25D₃, inecalcitol showed enhanced vitamin D receptor (VDR)-mediated transcriptional activity. Inecalcitol suppressed SCC cell proliferation in a dose-dependent manner with an IC₅₀ value 30 times lower than that of 1,25D₃. Both inecalcitol and 1,25D₃ induced a comparable level of G₀/G₁ cell cycle arrest in SCC cells. The level of apoptosis induced by inecalcitol was markedly higher than that of 1,25D₃. Apoptosis was mediated through the activation of the caspase 8/10- caspase 3 pathway. Further, inecalcitol markedly inhibited the mRNA and protein expression of c-IAP1 and XIAP compared with 1,25D₃. In vivo, inecalcitol inhibits SCC tumor growth in a dose-dependent fashion. Notably, inecalcitol induced a significantly higher level of apoptosis in the SCC xenograft model. While in vitro inecalcitol demonstrates apparent enhanced VDR binding and antiproliferative effects compared to 1,25D₃, in vivo these advantages disappear; at doses of inecalcitol that have equivalent antitumor effects, similar hypercalcemia is seen. This may be explained by the pharmacokinetics of 1,25D₃ vs. inecalcitol and attributed to the much shorter serum half-life of inecalcitol.We show that inecalcitol has potent antitumor activity in the SCC model system, and this is associated with a strong induction of apoptosis. These findings support the further development of inecalcitol in cancer treatment.
Keywords: :
Introduction
The active vitamin D metabolite, 1α, 25-dihydroxyvitamin D3 (1,25D3), has shown antitumor activities in vitro and in vivo in a number of cancer types.Citation1 Hypercalcemia may be the dose-limiting factor for the application of 1,25D3 in the clinic, particularly when continuous dosing schedules are employed.Citation1 Therefore, efforts have been made to develop analogs of 1,25D3 with a dissociation of antiproliferative and calcemic effects.Citation1
1,25D3 and its analogs exert a genomic action by binding to the intracellular vitamin D receptor (VDR), which heterodimerizes with retinoid X receptor (RXR). The heterodimer subsequently recruits coactivators and binds to vitamin D response elements (VDRE) in the promoter region of target genes to regulate gene transcription.Citation2 Two 14-epi analogs, inecalcitol [TX522, 19-nor-14-epi-23-yne-1,25-(OH)2D3] and TX527 [19-nor-14,20-bisepi-23-yne-1,25(OH)2D3], have been discovered, and they exert superagonistic activity yet have low calcemic effects. Biochemical studies demonstrate that both inecalcitol and TX527 induce stronger binding of VDR-RXR heterodimers to a direct repeat 3 (DR3)-type VDRE than 1,25D3 in DNA-dependent assays.Citation3 Moreover, inecalcitol and TX527 induce stronger interaction between VDR and coactivators, including steroid receptor coactivator 1 (SRC-1), transcriptional intermediary factor 2 (Tif2) and vitamin D receptor-interacting protein 205 (DRIP205). These effects contribute to their superagonistic action.Citation4 Both 14-epi analogs induce transcriptional activation from a VDRE more efficiently than 1,25D3 in COS-1 fibroblasts and human breast cancer MCF-7 cells.Citation3 In addition, inecalcitol and TX527 each are more resistant to metabolic degradation through 24-hydroxylase (CYP24).Citation4 Together, these biochemical characteristics contribute to their enhanced in vitro activity compared with 1,25D3.
The enhanced transcriptional activity is associated with increased antitumor activity of the 14-epi analogs. Inecalcitol and TX527 have enhanced anti-proliferative effects compared with 1,25D3 in MCF-7 cells in vitro and in vivo.Citation5 In vivo, inecalcitol is less (at least 4-fold) hypercalcemic than TX 527.Citation5 Therefore, inecalcitol was further developed for clinical investigation. Inecalcitol shows stronger inhibition of cell growth and clonogenic survival in human prostate cancer cell lines in vitro and less calcemic effects in vivo, compared with 1,25D3.Citation6 However, the mechanisms for the antitumor effect of inecalcitol remain to be fully understood.
In the current study, we chose a squamous cell carcinoma (SCC) model system that has been well established in our laboratory to investigate the antitumor activity of inecalcitol and the underlying mechanisms in vitro and in vivo.
Results
Inecalcitol induces stronger transcriptional activity than 1,25D3 in SCC cells
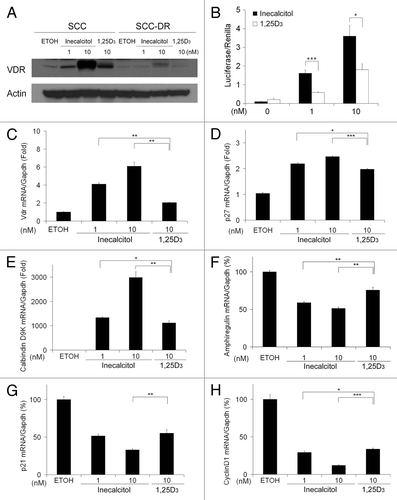
1,25D3 and its analogs exert most of their activities through binding to VDR and regulating the transcription of target genes, including VDR itself. Therefore, we first examined the ability of inecalcitol to promote VDR signaling in SCC cells or 1,25D3-resistant SCC-DR cells, generated through continuous culture of SCC cells in 1,25D3-containing media.Citation7 SCC or SCC-DR cells were treated with vehicle control ethanol (EtOH), 1 or 10 nM inecalcitol or 10 nM 1,25D3 for 48 h and VDR protein expression was examined by immunoblot analysis. VDR was induced by inecalcitol in a dose-dependent manner (), and 10 nM inecalcitol resulted in markedly higher induction of VDR in comparison to 10 nM 1,25D3 (). Nominal induction of VDR was observed in SCC-DR cells by 10 nM inecalcitol (), indicating the critical involvement of vitamin D signaling. To compare VDR-mediated transcriptional activity induced by inecalcitol or 1,25D3 in SCC cells, a luciferase reporter assay was used, as previously described.Citation8 Cells were infected with an adenoviral vector that carries the luciferase reporter gene driven by the CYP24A1 promoter. Treatment with 1 or 10 nM inecalcitol produced significantly higher VDR-mediated transcription of the CYP24A1 promoter compared with the same concentrations of 1,25D3 (). Further, mRNA expression of the known VDR target genes was examined. Consistent with the VDR protein expression results, inecalcitol induced the mRNA expression of VDR in a dose-dependent manner, and the induction was significantly higher than the induction with 1,25D3 (). Likewise, inecalcitol induced the mRNA expression of p27 () and calbindin D9K (), while suppressing the expression of amphiregulin (), p21 () and cyclin D1 () dose-dependently and to a greater extent than 1,25D3. At equal molar dose and exposure time, inecalcitol-modulated mRNA expression was significantly stronger than 1,25D3 for all the target genes (). These results indicate that inecalcitol is a more potent inducer of VDR transcriptional activities in SCC.
Figure 1. Inecalcitol induces stronger transcriptional activity than 1,25D3 in SCC cells. (A) SCC or SCC-DR cells were treated with vehicle control EtOH, 1 or 10 nM inecalcitol, or 10 nM 1,25D3 for 48 h. VDR expression was assessed by immunoblot analysis. Actin was the loading control. Results are representative of two independent experiments. (B) 72 h of treatment with inecalcitol induces higher VDR-mediated transcription of the CYP24A1 promoter compared with induction with 1,25D3. Results were normalized to renilla activity as a control for infection. (C–H) SCC cells were treated with EtOH, 1 or 10 nM inecalcitol, or 10 nM 1,25D3 for 24 h unless specified otherwise. mRNA expression of VDR target genes vdr (C), 16 h treatment, p27 (D), 6 h treatment, calbindin D9K (E), amphiregulin (F), p21 (G) and cyclin D1 (H) were assessed by real-time PCR and normalized to mouse gapdh. Results are representative of two independent experiments. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Inecalcitol has a stronger growth inhibitory effect on SCC than 1,25D3 in vitro.
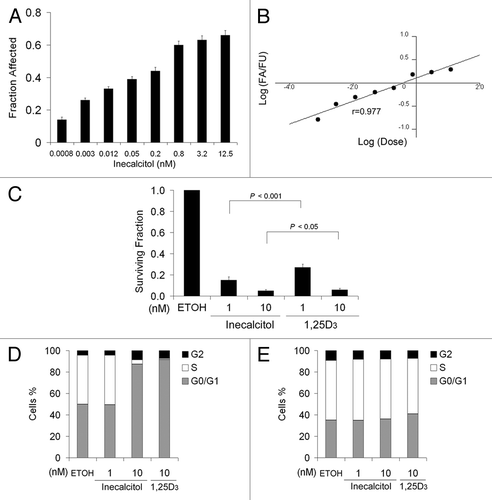
To determine the effect of inecalcitol on SCC cell growth, MTT and in vitro clonogenic assays were employed. SCC cells were treated with increasing concentrations of inecalcitol for 48 h and subjected to MTT assay. Inecalcitol inhibited the growth of SCC cells in a dose-dependent manner (). The IC50 of inecalcitol was determined to be 0.38 nM [95% confidence interval (CI) of 0.17 to 0.83 nM] by Calcusyn software (). The IC50 for 1,25D3 in SCC cells (12 nM)Citation9 was ~30 fold higher than the IC50 of inecalcitol. The more sensitive clonogenic assay revealed that inecalcitol treatment resulted in significantly stronger inhibition of the clonogenic capacity of SCC cells than 1,25D3 treatment at either 1 nM (p < 0.001) or 10 nM (p < 0.05) final concentration (). These results indicate that inecalcitol has greater anti-proliferative effects than 1,25D3 in SCC cells in vitro.
Figure 2. Inecalcitol has greater growth inhibitory effect on SCC than 1,25D3 in vitro. (A) SCC cells were treated with increasing doses of inecalcitol (0.0008–12.5 nM) for 48 h and subjected to MTT assays. Fraction affected (FA) was calculated as 1-(MTT value of the treatment cells)/(MTT value of EtOH-treated cells). (B) Median-effect blot generated using CalcuSyn software to determine the IC50 of inecalcitol. (C) SCC cells were treated with EtOH, inecalcitol, or 1,25D3 and subjected to in vitro clonogenic assaysf. The clones were fixed, stained and counted on day 9. Results are the mean ± SD of triplicate determinations and are representative of three independent experiments. (D and E) SCC (D) or SCC-DR (E) cells were treated with EtOH, 1 or 10 nM inecalcitol, or 10 nM 1,25D3 for 48 h. Cell cycle was examined by PI staining using flow cytometry. Results are representative of three independent experiments.
Inecalcitol induces cell cycle arrest in SCC cells
To elucidate the potential mechanisms for the anti-proliferative effects of inecalcitol, its role in cell cycle progression was studied. Inecalcitol had no effect at 1 nM but induced profound G0/G1 cell cycle arrest at 10 nM, to a similar extent as 1,25D3 at the same dose (). Neither agent affected cell cycle progression in SCC-DR cells (). These data suggest that the induction of cell cycle arrest is associated with the anti-proliferative effects of inecalcitol, but does not explain its enhanced activity compared with 1,25D3.
Inecalcitol induces significantly greater apoptosis than 1,25D3 in SCC cells
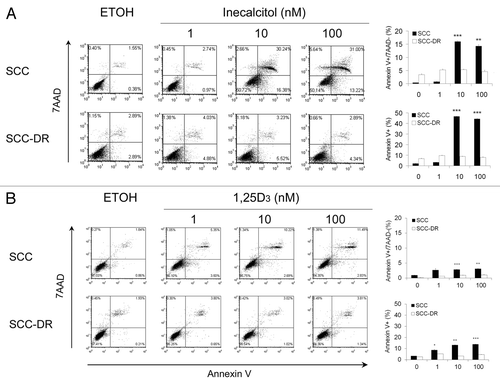
To examine whether apoptosis contributes to inecalcitol-induced cytotoxicity, SCC or SCC-DR cells were treated with EtOH, inecalcitol or 1,25D3 for 48 h, and apoptosis was assessed by annexin V/7AAD staining. Inecalcitol at 10 or 100 nM induced ~15% of early apoptosis (annexin V+/7AAD−) and 40–50% of total apoptosis (annexin V+) in SCC cells (). This was markedly higher than the levels induced by 1,25D3 (2–3% of early apoptosis and 10–15% of total apoptosis) (). Neither agent induced apoptosis in SCC-DR cells ().
Figure 3. Inecalcitol induces more apoptosis than 1,25D3 in SCC cells. SCC or SCC-DR cells were treated with EtOH, 1 to 100 nM inecalcitol (A) or 1,25D3 (B) for 48 h. Apoptosis was assessed by annexin V-PE and 7AAD staining by flow cytometry. The populations of annexin V−/7AAD−, annexin V+/7AAD− and annexin V+/7AAD+ correspond to live cells, early apoptotic cells and late apoptotic cells, respectively. Representative histograms are presented. The results were summarized in bar graphs as mean ± s.e.m. of annexin V+/7AAD- cells or annexin V+ cells in a representative experiment. *, p < 0.05; **, p < 0.01; ***, p < 0.001. Treatment vs. control “0.”
Inecalcitol induces the activation of caspases 8/10 and 3
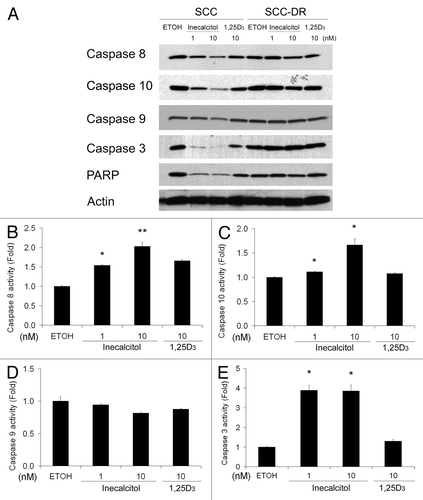
We subsequently investigated the mechanisms for apoptosis induction by inecalcitol. To investigate whether the intrinsic and/or extrinsic apoptotic pathways are involved, the activation of the major caspases was studied by immunoblot analysis. Compared with EtOH treatment, inecalcitol promoted the cleavage of pro-caspases 8, 10, 3 and poly (ADP-ribose) polymerase (PARP) in SCC cells in a dose-dependent manner, and more intensely than 1,25D3 at the same 10 nM dosage (). The level of pro-caspase 9 was unaffected by either agent (). Caspase cleavage was not observed in SCC-DR cells (). These results were confirmed using substrate-based caspase activity assays, which showed that inecalcitol treatment resulted in increased activities of caspases 8, 10 and 3 (), but not that of caspase 9 (). 1,25D3 induced modest increases in the activities of caspases 8 and 3 (). These results indicate that both inecalcitol and 1,25D3 promote apoptosis through the activation of the extrinsic pathway mediated by caspase 8/10 and caspase 3.
Figure 4. Inecalcitol induces the activation of caspases 8/10 and 3. SCC or SCC-DR cells were treated with EtOH, 1 or 10 nM inecalcitol, or 10 nM 1,25D3 for 48 h. (A) The levels of caspases 8, 10, 9, 3 and PARP were evaluated by immunoblot analysis. Actin was the loading control. (B–E) The activities of caspase 8 (B), 10 (C), 9 (D) and 3 (E) were examined using substrate-based caspase activity assays. Results are representative of three independent experiments. *, p < 0.05; **, p < 0.01.
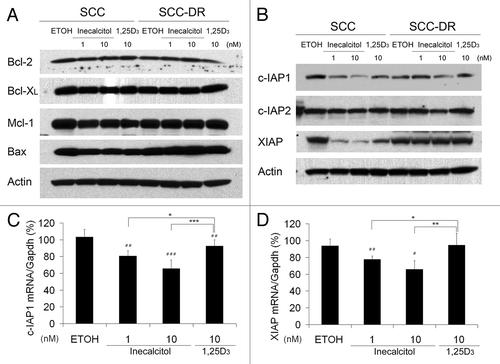
Inecalcitol reduces the expression of c-IAP1 and XIAP
Bcl-2 and the inhibitor of apoptosis (IAP) family members are the major regulators of apoptosis.Citation10,Citation11 Therefore, we next examined whether they are involved in inecalcitol or 1,25D3-mediated apoptosis. Immunoblot analysis showed that neither inecalcitol nor 1,25D3 had any effect on the protein levels of anti-apoptotic Bcl-2, Bcl-XL and Mcl-1 and the pro-apoptotic Bax (). Among IAP proteins, the expression levels of c-IAP1 and XIAP were reduced by inecalcitol in a dose-dependent manner (). 1,25D3 reduced the level of c-IAP1 and XIAP, but to a lesser extent as compared with inecalcitol at the same concentration (10 nM) (). c-IAP2 levels were not affected by either agent (). The regulation of c-IAP1 and XIAP occurred at the transcriptional level, because qRT-PCR results showed that inecalcitol treatment for 24 h resulted in a significant reduction of the mRNA levels of c-IAP1 and XIAP (p < 0.01) (, respectively). 1,25D3 modestly reduced the mRNA level of c-IAP1 but not that of XIAP (, respectively). Taken together, these data suggest that both inecalcitol and 1,25D3-promoted apoptosis were associated with the selective downregulation of anti-apoptotic IAP proteins.
Figure 5. Inecalcitol reduces the expression of c-IAP1 and XIAP. SCC or SCC-DR cells were treated with EtOH, 1 or 10 nM inecalcitol, or 10 nM 1,25D3 for 48 h (A and B) or 24 h (C and D). (A) The protein levels of Bcl-2, Bcl- XL, Mcl-1 and Bax were evaluated by immunoblot analysis. Actin was the loading control. (B) c-IAP1, c-IAP2 and XIAP levels were evaluated by immunoblot analysis. Actin was the loading control. (C and D) mRNA expression levels of c-IAP1 (C) and XIAP (D) were assessed by qRT-PCR and the value is normalized to the mRNA level of Gapdh. Results are representative of three independent experiments. *, p < 0.05; **, p < 0.01; ***, p < 0.001. Treatment vs. EtOH: #, p < 0.05; ##; p < 0.01.
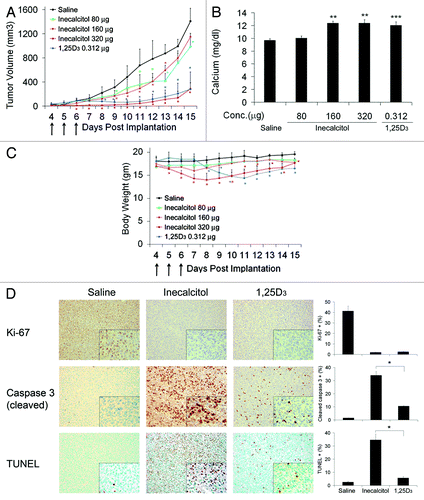
In vivo antitumor activity of inecalcitol and 1,25D3.
To evaluate whether the observations in vitro translate to antitumor effects in vivo, 4 d post-inoculation, SCC tumor-bearing mice were treated with saline, 80, 160 or 320 μg of inecalcitol or 0.312 μg of 1,25D3 daily for 3 d. Eighty or 160 μg inecalcitol treatment resulted in modest but significant (p < 0.05) suppression of tumor growth, while inecalcitol (320 μg) or 1,25D3 (0.312 μg) almost completely inhibited tumor growth (). To study the safety profiles of inecalcitol and 1,25D3, serum calcium levels and body weight were monitored. 1,25D3 treatment resulted in elevated calcium level (). Inecalcitol at 80 μg dosage did not affect serum calcium level, but at 160 or 320 μg dosage, inecalcitol increased the calcium level (). Weight loss was not observed in mice treated with 80 or 160 μg inecalcitol, but was observed upon treatment with 320 μg inecalcitol or 0.312 μg 1,25D3 (). To study the mechanisms for the antitumor activity in vivo, the effect of inecalcitol or 1,25D3 on SCC cell proliferation and apoptosis was assessed by immunohistochemical analysis of tumor tissues. The dose 80 μg was chosen for inecalcitol, since this dose had significant antitumor activity without side effects. Ki-67 staining revealed that inecalcitol and 1,25D3 had a similar strong anti-proliferative effect on SCC tumors (). Staining of cleaved caspase 3 and the in situ TUNEL assay showed that inecalcitol induced significantly more evidence of apoptosis by these measures than 1,25D3 in vivo (). These data indicate that inecalcitol and 1,25D3 in vivo each exerted potent antitumor effects at 320 μg and 0.312 μg QD × 3, respectively, and inecalcitol was a potent inducer of apoptosis in vivo in the SCC system.
Figure 6. In vivo antitumor activity of inecalcitol and 1,25D3. Four days post inoculation, SCC tumor-bearing C3H mice were treated with saline, 80, 160 or 320 μg/mouse inecalcitol or 0.312 μg/mouse 1,25D3 daily for 3 d. (A) Tumor growth was monitored and measurements taken on the days indicated. Arrows indicate the days animals received treatment. Tumor volumes were calculated by the following formula: volume = (length × width2)/2. (B) Blood was collected 24 h after the last treatment, and serum calcium levels were detected using a colorimetric calcium assay kit. (C) Body weights of the mice were monitored on the days indicated. Arrows indicate the days animals received treatment. (D) SCC tumors in mice treated with 80 μg inecalcitol or 0.312 μg 1,25D3 were harvested at the end of the study and tissues stained with antibodies for Ki-67 and cleaved caspase 3 for immunohistochemistry study or subjected to TUNEL assay. The percentage of positive stained cells were quantified and presented in bar graphs next to the representative images. (×200 and ×400) *, p < 0 0.05; **, p < 0.01; ***, p < 0.001, Treatment vs. saline.
Discussion
Epidemiological and experimental studies provide justification for the study of 1,25D3 in cancer prevention and treatment.Citation12,Citation13 However, hypercalcemia remains a limitation of 1,25D3 when used at higher doses. Inecalcitol is a 1,25D3 analog optimized to have strong anti-proliferative activity coupled with low calcemic effect. Structurally, inecalcitol differs from 1,25D3 through epimerization of C14, deletion of C19 and 23-yne modification in the side chain. The greater interaction between VDR and its coactivators, the improved docking of inecalcitol in the ligand binding domain of VDR and the delayed metabolism by 24-hydroxylase all explain the superagonistic activity of inecalcitol.Citation4 Inecalcitol has shown more enhanced anti-proliferative activity than 1,25D3 in several tumor model systems.Citation3,Citation5,Citation6 However, the underlying mechanisms for the antitumor activity are incompletely understood.
We have previously shown that 1,25D3 has growth inhibitory activity in the SCC model system in vitro and in vivo.Citation14 We chose this SCC model system to study the antitumor effect of inecalcitol in comparison to 1,25D3. Inecalcitol shows stronger regulation of VDR transcriptional activity than 1,25D3, as indicated by increased VDR-mediated transcription of CYP24A1 promoter and enhanced modulation of a number of known VDR target genes. The enhanced VDR signaling induced by inecalcitol translates into stronger in vitro anti-proliferative effects, as indicated by a 30-fold lower IC50 than that of 1,25D3 and stronger anti-clonogenic capacity.
We further evaluated the mechanisms for the enhanced antitumor activity of inecalcitol. Both inecalcitol and 1,25D3 induce similar level of G0/G1 cell cycle arrest in SCC cells. In vivo, the Ki-67 staining indicated comparable inhibition of cell cycle progression by each agent. These results indicate that the induction of cell cycle arrest is one of the mechanisms for antitumor activity of both inecalcitol and 1,25D3, but does not explain the enhanced activity of inecalcitol in vitro.
Next, we studied the role of apoptosis in the antitumor effect of inecalcitol. Notably, inecalcitol induces significantly more apoptosis than 1,25D3 in SCC cells in vitro. Apoptosis occurred through the caspase 8/10-caspase 3 pathway. Further investigation of the potential mediators of apoptosis reveals that inecalcitol treatment results in strong inhibition of the expression of two IAP family members, c-IAP1 and XIAP, at both mRNA and protein level. 1,25D3 only modestly reduces the level of c-IAP1. In vivo, inecalcitol induces markedly more apoptosis in SCC tumor tissues than 1,25D3 as indicated by the staining of cleaved caspase 3 and the in situ TUNEL assay. These data indicate that inecalcitol is a stronger inducer of apoptosis than 1,25D3 in SCC cells in vitro and in vivo, which may explain the enhanced antitumor effect of inecalcitol.
1,25D3-resistant SCC-DR cells express very low levels of VDR.Citation7 In SCC-DR cells, inecalcitol has no effect on cell cycle progression or apoptosis, indicating that VDR signaling is crucial in inecalcitol activity.
Collectively, studies from our laboratory suggest that inecalcitol and 1,25D3 have different mechanisms for their antitumor activity in SCC cells. The anti-proliferative effect of inecalcitol involves both cell cycle arrest and intense apoptosis. The mechanisms for the antitumor activity of 1,25D3 seem to involve the induction of G0/G1 cell cycle arrest associated with the reduction of p21 protein level.Citation14,Citation15 Little or no apoptosis is induced by 1,25D3 in SCC cells when the entire population is assessed.Citation14,Citation16 In the fraction of floating SCC cells following 1,25D3 treatment, apoptosis can be detected,Citation17 suggesting that the floating cells account for a small part of the whole population. The ability of inecalcitol to induce apoptosis appears to be cell type-specific. Inecalcitol treatment for 72 h induces less than 10% apoptosis in human breast cancer MCF-7 cells.Citation5 Supporting our findings, a recent study shows that inecalcitol treatment results in profound apoptosis in LNCaP xenograft tumors in vivo.Citation6
The mechanisms for inecalcitol-mediated apoptosis have not been reported previously. The apoptosis-inducing mechanisms for 1,25D3 or other analogs remain to be fully understood and appear to be cancer and cell type-specific. 1,25D3 induces caspase-independent apoptosis in MCF-7 cells by interrupting mitochondrial function, which is accompanied by Bax translocation and the production of reactive oxygen species.Citation18 1,25D3 or its analog EB1089 promotes apoptosis in MCF-7 cells by enhancing the intracellular calcium level, which results in the activation of the calcium-dependent cysteine protease μ-calpain.Citation19 1,25D3 or EB1089-induced apoptosis in colorectal cancer cell lines is associated with promoted protein level of Bak.Citation20 In prostate cancer LNCaP and ALVA-31 cells, 1,25D3 promotes apoptosis by the reduction of the expression of Bcl-2, Bcl-xL, Mcl-1 and IAP proteins.Citation21 In ovarian cancer OVCAR3 cells, 1,25D3-induced apoptosis involves the downregulation of telomerase by reducing the stability of the human telomerase reverse transcriptase mRNA.Citation22 In peripheral blood lymphocytes isolated from B-cell chronic lymphocytic leukemia (B-CLL) patients, EB1089 induces apoptosis through the activation of p38 MAP kinase, suppression of ERK activation and the levels of Bcl-2 and Mcl-1.Citation23
Further, in the SCC xenograft model, inecalcitol inhibits tumor growth in a dose-dependent manner. Notably, at the 80 μg daily dose for 3 d, inecalcitol shows significant tumor growth inhibition and profound apoptosis without affecting serum calcium level or body weight. It requires a higher dose for inecalcitol (320 μg) than 1,25D3 (0.312 μg) to achieve comparable antitumor effect, which is not consistent with the in vitro results showing that inecalcitol has significantly stronger anti-proliferative effect than 1,25D3 at the same dose (10 nM). This observation can be explained, at least in part, by their different pharmacokinetic profiles. Inecalcitol has a half-life of 18.3 min and clearance of 53.3 ml/h in mice.Citation6 In contrast, 1,25D3 has a much longer half-life of 4.4 h and clearance of 192 ml/h.Citation24 Therefore, a higher dose of inecalcitol is necessary to deliver antitumor effect. These observations indicate that inecalcitol (80 μg) is able to achieve the dissociation of antitumor and calcemic effect and support its application in combinational chemotherapy.
Inecalcitol has been utilized in clinical studies. Inecalcitol is well tolerated at 4 mg daily by mouth. There was neither toxicity nor hypercalcemia in a single oral dose phase I clinical trial in healthy volunteers up to 320 μg (Delansorne R, personal communication). Results of the subsequent phase II trial in castration-resistant prostate cancer showed that inecalcitol in combination with docetaxel had a better PSA response compared with docetaxel alone (> 30% PSA decline in 3 mo: 85% vs. 67%; > 50% PSA decline anytime: 76% vs. 45%) [Medioni J, et al. J Clin Oncol 29: 2011 (suppl; abstr 4605)]. These data provide support for further evaluation of inecalcitol in cancer therapy.
In summary, this study reports that a 14-epi analog of 1,25D3, inecalcitol, shows enhanced growth inhibitory effect in vitro and strong antitumor activity in vivo in comparison to 1,25D3 in a SCC model system. The mechanism for the enhanced antitumor effect of inecalcitol is based on potent induction of apoptosis, which is mediated by the caspase 8/10–3 pathway and is associated with the reduction of c-IAP1 and XIAP levels.
Materials and Methods
Materials
1,25D3 was provided by Hoffmann-LaRoche and inecalcitol by Hybrigenics. Annexin V-PE and 7AAD were purchased from BD Biosciences. Anti-caspase 3, anti-caspase 9, anti-caspase 10, anti-PARP and anti-XIAP were from Cell Signaling Technology. Anti-VDR, anti-Bcl-2, anti-Bcl-XL, anti-Bax, anti-Mcl-1, anti-c-IAP1 and anti-c-IAP2 were purchased from Santa Cruz Biotechnology. Anti-caspase 8 (552038) was from BD PharMingen. Anti-actin was purchased from Calbiochem.
Cell culture and tumor model systems
The murine SCC (SCCVII-SF) cells were used as described previously.Citation14 1,25D3-resistant SCC-DR cells were generated by continuously culturing SCC cells in media containing 10 nM 1,25D3, as described previously.Citation7 SCC cells were maintained in 6–10-week-old female C3H/HeJ mice purchased from Jackson Laboratory. The murine protocol used was approved by the Roswell Park Cancer Institutional Animal Care and Use Committee.
Luciferase reporter assay
VDR-mediated transcription was assessed in the presence of 1,25D3 or inecalcitol by using an adenovirus carrying the firefly luciferase gene driven by the CYP24A1 promoter, which contains two VDREs.Citation8,Citation25 Luciferase reporter assay was performed as described previously.Citation8
Immunoblot analysis
SCC and SCC-DR cells were treated with vehicle control EtOH, 1 or 10 nM inecalcitol, or 10 nM 1,25D3 for 48 h. Cell were then harvested and lysates prepared for immunoblot analysis as previously described.Citation16
In vitro cytotoxicity assay and dose-effect analysis
SCC cells were plated in 96-well tissue culture plates. Cells were treated with EtOH or varying concentrations (0.0008–12.5 nM) of inecalcitol for 48 h. Cell growth was assessed by MTT assay. Dose-effect was analyzed by the median-dose effect method, and IC50 values were calculated using CalcuSyn software (Biosoft).
In vitro clonogenic assay
SCC cells were treated with EtOH or 1 or 10 nM inecalcitol or 1,25D3 and subjected to in vitro clonogenic assays, as described previously.Citation26
Cell cycle analysis
SCC cells were harvested, fixed in 70% EtOH and stained with propidium iodide (PI) (50 μg/ml). The cell cycle profile was determined by flow cytometry utilizing Becton Dickinson FACScan and FCS Express software (De Novo Software).
Apoptosis assay
SCC and SCC-DR cells were collected and stained with PE-conjugated annexin V and 7AAD, according to manufacturer’s instructions, as described previously.Citation16
Caspase activity assays
The activities of caspases-3, 8, 9 (R&D Systems) and caspase 10 (EMD Millipore) were measured using the caspase fluorometric assay kit following the manufacturer’s protocols. Caspase activity was normalized to protein concentration.
Quantitative reverse transcriptase PCR (qRT-PCR)
Total RNA was isolated from SCC cells using the RNeasy kit (Qiagen) according to manufacturer's instructions. cDNA was obtained by reverse transcription of 1 μg of total RNA using First Strand cDNA Synthesis Kit (Roche Applied Science) following the manufacturer’s protocol. mRNA expression of VDR, p27, calbindin D9K, Amphiregulin, p21, cyclinD1, XIAP and c-IAP1 was assessed by qRT-PCR using TaqMan® Gene expression assays. Assays were run on a 7300 Real-Time PCR System (Applied Biosystems), as described previously.Citation27 Data was analyzed using the RQ study software (Applied Biosystems).
Tumor regrowth delay
SCC cells (1.5 × 105) were inoculated s.c. into the flank of C3H mice. Studies were initiated when tumors were palpable, 4 d post implantation. Mice were treated in five groups (10 per group): saline, inecalcitol (80, 160, or 320 μg/mouse) or 1,25D3 (0.312 μg/mouse). The doses were determined in pilot dose finding experiments. Mice were treated for 3 d with single, daily i.p. injections. Body weights were measured daily after tumor implantation. Tumor measurements were taken as described.Citation28 Serum calcium levels were measured 24 h after the last injection of inecalcitol or 1,25D3 using the QuantiChrom™ calcium assay kit (BioAssay Systems).
Immunohistochemistry
SCC tumor tissues were harvested and immunohistochemical detection of Ki-67 and cleaved caspase 3 was performed as described previously.Citation29 TUNEL assay was performed using the TACS-XL In Situ Apoptosis Detection Kit (R&D Systems) according to manufacturer’s instruction.
Statistical analyses
The two-tailed Student's t-test was used to determine statistical significance between groups.
| Abbreviations: | ||
| 1,25D3 | = | 1,α, 25-dihydroxyvitamin D3 |
| SCC | = | squamous cell carcinoma |
| VDR | = | vitamin D receptor |
| RXR | = | retinoid X receptor |
| VDRE | = | vitamin D response elements |
| SRC-1 | = | steroid receptor coactivator 1 |
| Tif2 | = | transcriptional intermediary factor 2 |
| DRIP205 | = | vitamin D receptor-interacting protein 205 |
| CYP24 | = | 24-hydroxylase |
| EtOH | = | ethanol |
| IAP | = | inhibitor of apoptosis |
Acknowledgments
We thank Dr Josephia R. Muindi for his scientific contribution, Ms Rui-Xian Kong for her excellent technical support and Dr Pamela A. Hershberger for her critical review of the manuscript. This study was supported by NIH/NCI grants CA067267 and CA085142 to Dr Candace S. Johnson, and CA095045 to Dr Donald L. Trump. It was also supported, in part, by the NCI Cancer Center Support Grant to the Roswell Park Cancer Institute (CA016056).
Disclosure of Potential Conflicts of Interest
Remi Delansorne owns 1.3% of Hybrigenics’shares. There are no financial disclosures from any other authors.
Related Research Data
References
- Deeb KK, Trump DL, Johnson CS. Vitamin D signalling pathways in cancer: potential for anticancer therapeutics. Nat Rev Cancer 2007; 7:684 - 700; http://dx.doi.org/10.1038/nrc2196; PMID: 17721433
- Brown AJ, Dusso A, Slatopolsky E. Vitamin D. Am J Physiol 1999; 277:F157 - 75; PMID: 10444570
- Verlinden L, Verstuyf A, Quack M, Van Camp M, Van Etten E, De Clercq P, et al. Interaction of two novel 14-epivitamin D3 analogs with vitamin D3 receptor-retinoid X receptor heterodimers on vitamin D3 responsive elements. J Bone Miner Res 2001; 16:625 - 38; http://dx.doi.org/10.1359/jbmr.2001.16.4.625; PMID: 11315990
- Eelen G, Verlinden L, Rochel N, Claessens F, De Clercq P, Vandewalle M, et al. Superagonistic action of 14-epi-analogs of 1,25-dihydroxyvitamin D explained by vitamin D receptor-coactivator interaction. Mol Pharmacol 2005; 67:1566 - 73; http://dx.doi.org/10.1124/mol.104.008730; PMID: 15728261
- Verlinden L, Verstuyf A, Van Camp M, Marcelis S, Sabbe K, Zhao XY, et al. Two novel 14-Epi-analogues of 1,25-dihydroxyvitamin D3 inhibit the growth of human breast cancer cells in vitro and in vivo. Cancer Res 2000; 60:2673 - 9; PMID: 10825140
- Okamoto R, Delansorne R, Wakimoto N, Doan NB, Akagi T, Shen M, et al. Inecalcitol, an analog of 1α,25(OH)(2) D(3), induces growth arrest of androgen-dependent prostate cancer cells. Int J Cancer 2012; 130:2464 - 73; http://dx.doi.org/10.1002/ijc.26279; PMID: 21732345
- Ma Y, Yu WD, Hershberger PA, Flynn G, Kong RX, Trump DL, et al. 1alpha,25-Dihydroxyvitamin D3 potentiates cisplatin antitumor activity by p73 induction in a squamous cell carcinoma model. Mol Cancer Ther 2008; 7:3047 - 55; http://dx.doi.org/10.1158/1535-7163.MCT-08-0243; PMID: 18790784
- Hidalgo AA, Deeb KK, Pike JW, Johnson CS, Trump DL. Dexamethasone enhances 1alpha,25-dihydroxyvitamin D3 effects by increasing vitamin D receptor transcription. J Biol Chem 2011; 286:36228 - 37; http://dx.doi.org/10.1074/jbc.M111.244061; PMID: 21868377
- Hershberger PA, McGuire TF, Yu WD, Zuhowski EG, Schellens JH, Egorin MJ, et al. Cisplatin potentiates 1,25-dihydroxyvitamin D3-induced apoptosis in association with increased mitogen-activated protein kinase kinase kinase 1 (MEKK-1) expression. Mol Cancer Ther 2002; 1:821 - 9; PMID: 12492115
- Li L, Ittmann MM, Ayala G, Tsai MJ, Amato RJ, Wheeler TM, et al. The emerging role of the PI3-K-Akt pathway in prostate cancer progression. Prostate Cancer Prostatic Dis 2005; 8:108 - 18; http://dx.doi.org/10.1038/sj.pcan.4500776; PMID: 15724144
- Song G, Ouyang G, Bao S. The activation of Akt/PKB signaling pathway and cell survival. J Cell Mol Med 2005; 9:59 - 71; http://dx.doi.org/10.1111/j.1582-4934.2005.tb00337.x; PMID: 15784165
- Garland CF, Garland FC, Gorham ED, Lipkin M, Newmark H, Mohr SB, et al. The role of vitamin D in cancer prevention. Am J Public Health 2006; 96:252 - 61; http://dx.doi.org/10.2105/AJPH.2004.045260; PMID: 16380576
- Giovannucci E, Liu Y, Rimm EB, Hollis BW, Fuchs CS, Stampfer MJ, et al. Prospective study of predictors of vitamin D status and cancer incidence and mortality in men. J Natl Cancer Inst 2006; 98:451 - 9; http://dx.doi.org/10.1093/jnci/djj101; PMID: 16595781
- Hershberger PA, Modzelewski RA, Shurin ZR, Rueger RM, Trump DL, Johnson CS. 1,25-Dihydroxycholecalciferol (1,25-D3) inhibits the growth of squamous cell carcinoma and down-modulates p21(Waf1/Cip1) in vitro and in vivo. Cancer Res 1999; 59:2644 - 9; PMID: 10363987
- Light BW, Yu WD, McElwain MC, Russell DM, Trump DL, Johnson CS. Potentiation of cisplatin antitumor activity using a vitamin D analogue in a murine squamous cell carcinoma model system. Cancer Res 1997; 57:3759 - 64; PMID: 9288784
- Ma Y, Yu WD, Kong RX, Trump DL, Johnson CS. Role of nongenomic activation of phosphatidylinositol 3-kinase/Akt and mitogen-activated protein kinase/extracellular signal-regulated kinase kinase/extracellular signal-regulated kinase 1/2 pathways in 1,25D3-mediated apoptosis in squamous cell carcinoma cells. Cancer Res 2006; 66:8131 - 8; http://dx.doi.org/10.1158/0008-5472.CAN-06-1333; PMID: 16912191
- McGuire TF, Trump DL, Johnson CS. Vitamin D(3)-induced apoptosis of murine squamous cell carcinoma cells. Selective induction of caspase-dependent MEK cleavage and up-regulation of MEKK-1. J Biol Chem 2001; 276:26365 - 73; http://dx.doi.org/10.1074/jbc.M010101200; PMID: 11331275
- Narvaez CJ, Welsh J. Role of mitochondria and caspases in vitamin D-mediated apoptosis of MCF-7 breast cancer cells. J Biol Chem 2001; 276:9101 - 7; http://dx.doi.org/10.1074/jbc.M006876200; PMID: 11053435
- Mathiasen IS, Sergeev IN, Bastholm L, Elling F, Norman AW, Jäättelä M. Calcium and calpain as key mediators of apoptosis-like death induced by vitamin D compounds in breast cancer cells. J Biol Chem 2002; 277:30738 - 45; http://dx.doi.org/10.1074/jbc.M201558200; PMID: 12072431
- Díaz GD, Paraskeva C, Thomas MG, Binderup L, Hague A. Apoptosis is induced by the active metabolite of vitamin D3 and its analogue EB1089 in colorectal adenoma and carcinoma cells: possible implications for prevention and therapy. Cancer Res 2000; 60:2304 - 12; PMID: 10786699
- Guzey M, Kitada S, Reed JC. Apoptosis induction by 1alpha,25-dihydroxyvitamin D3 in prostate cancer. Mol Cancer Ther 2002; 1:667 - 77; PMID: 12479363
- Jiang F, Bao J, Li P, Nicosia SV, Bai W. Induction of ovarian cancer cell apoptosis by 1,25-dihydroxyvitamin D3 through the down-regulation of telomerase. J Biol Chem 2004; 279:53213 - 21; http://dx.doi.org/10.1074/jbc.M410395200; PMID: 15485861
- Pepper C, Thomas A, Hoy T, Milligan D, Bentley P, Fegan C. The vitamin D3 analog EB1089 induces apoptosis via a p53-independent mechanism involving p38 MAP kinase activation and suppression of ERK activity in B-cell chronic lymphocytic leukemia cells in vitro. Blood 2003; 101:2454 - 60; http://dx.doi.org/10.1182/blood-2002-07-1984; PMID: 12446453
- Muindi JR, Modzelewski RA, Peng Y, Trump DL, Johnson CS. Pharmacokinetics of 1alpha,25-dihydroxyvitamin D3 in normal mice after systemic exposure to effective and safe antitumor doses. Oncology 2004; 66:62 - 6; http://dx.doi.org/10.1159/000076336; PMID: 15031600
- Hidalgo AA, Trump DL, Johnson CS. Glucocorticoid regulation of the vitamin D receptor. J Steroid Biochem Mol Biol 2010; 121:372 - 5; http://dx.doi.org/10.1016/j.jsbmb.2010.03.081; PMID: 20398752
- Yu WD, McElwain MC, Modzelewski RA, Russell DM, Smith DC, Trump DL, et al. Enhancement of 1,25-dihydroxyvitamin D3-mediated antitumor activity with dexamethasone. J Natl Cancer Inst 1998; 90:134 - 41; http://dx.doi.org/10.1093/jnci/90.2.134; PMID: 9450573
- Luo W, Karpf AR, Deeb KK, Muindi JR, Morrison CD, Johnson CS, et al. Epigenetic regulation of vitamin D 24-hydroxylase/CYP24A1 in human prostate cancer. Cancer Res 2010; 70:5953 - 62; http://dx.doi.org/10.1158/0008-5472.CAN-10-0617; PMID: 20587525
- Hershberger PA, Yu WD, Modzelewski RA, Rueger RM, Johnson CS, Trump DL. Calcitriol (1,25-dihydroxycholecalciferol) enhances paclitaxel antitumor activity in vitro and in vivo and accelerates paclitaxel-induced apoptosis. Clin Cancer Res 2001; 7:1043 - 51; PMID: 11309356
- Muindi JR, Yu WD, Ma Y, Engler KL, Kong RX, Trump DL, et al. CYP24A1 inhibition enhances the antitumor activity of calcitriol. Endocrinology 2010; 151:4301 - 12; http://dx.doi.org/10.1210/en.2009-1156; PMID: 20591973