Abstract
Messenger RNA translation, or protein synthesis, is a fundamental biological process affecting cell growth, survival and proliferation. Initiation is the rate limiting and hence the most regulated step of translation. In eukaryotes, translation initiation is facilitated by multiple protein factors collectively called eIFs (for eukaryotic translation initiation factors). The complex consisting of the eIF4 group factors including the mRNA cap-binding eIF4E protein, large scaffolding protein eIF4G and RNA helicase eIF4A is assisted by the eIF4B co-factor to unwind local secondary structures and create a ribosome landing pad on mRNA. Recruitment of the ribosome and augmentation in the mRNA scanning process culminates in the positioning of the ribosome over the start codon. Deregulated translational control is believed to play an important role in oncogenic transformation. Indeed, many eIFs are bona fide proto-oncogenes. In many types of human cancers, eIFs are either overexpressed or ectopically activated by Ras-MAPK and PI3K-mTOR signaling cascades, resulting in increased survival and accelerated proliferation. In this review we will analyze the bulk of data describing eIF4B and its role in cell survival and proliferation. Recent studies have shown that eIF4B is phosphorylated and activated by Ras-MAPK and PI3K-mTOR signaling cascades. In addition, eIF4B regulates translation of proliferative and pro-survival mRNAs. Moreover, eIF4B depletion in cancer cells attenuates proliferation, sensitizes them to genotoxic stress driven apoptosis. Taken together, these findings identify eIF4B as a potential target for development of anti-cancer therapies.
Introduction to Mammalian eIF4B
The translation initiation factor 4B has been purified and identified in the late 70's as a translation-stimulating activity in vitro. Later studies have shown that it functionally interacts with eIF4F and eIF4A potentiating their RNA helicase activities in vitro.Citation1 eIF4B is an RNA binding protein and is the least evolutionarily conserved translation initiation factor. Despite low sequence similarity, the eIF4B homologues from different species share overall comparable domain structures: N-terminal RRM (RNA Recognition Motif), domain, middle DRYG region and C-terminal ARM (Arginine Rich Motif) domain (). The RRM and ARM domains contribute to RNA binding, whereas the DRYG motif is important for interaction with eIF3 and selfassociation of eIF4B.Citation2,Citation3
The stable physical interaction between eIF4A and its co-factor eIF4B in the presence of RNA and an ATP analog AMPPNP has been described.Citation4 The region important for eIF4A activity resides within the C-terminal region of eIF4B adjacent to the ARM domain, however deletion of DRYG or point mutations in N-terminal RRM domains drastically reduce the ability of eIF4B to stimulate eIF4A helicase activity in vitro. eIF4B also has been reported to interact with the eIF3a subunit and 18S rRNA.Citation3,Citation5 These findings illustrate a model whereby eIF4B forms auxiliary bridges between the mRNA and the 40S ribosomal subunit and then facilitates the scanning process. An alternative hypothesis proposes that eIF4B is essential for unidirectional 5′ to 3′ mRNA scanning by the 43S complex.Citation6 This ‘Brownian Ratchet’ model assigns eIF4B the role of a pawl that prevents sliding of the ribosome in the opposite direction. A recent study demonstrated that eIF4E may be crosslinked at most, 2 nucleotides from the mRNA 5′cap structure, whereas crosslinking of eIF4B or its paralogue eIF4H could be observed up to 52 nucleotides downstream. These findings raised the possibility that eIF4B and eIF4H are chaperone-like factors seeded on mRNA to stabilize unwound linear stretches of the mRNA template.Citation7 The eIF4F complex was proposed to play the role of nucleation, since the crosslinking is sensitive to cap-analog addition and hence is 5′ cap-dependent. As previously mentioned eIF4H is a paralogue of eIF4B. eIF4H is shorter than eIF4B and both proteins share significant sequence homology over the entire length of eIF4H. Either eIF4B or eIF4H can stimulate eIF4A helicase activity and their binding to eIF4A is mutually exclusive.Citation4 More recently, site-directed mutagenesis and biophysical experiments assessed the topology of the eIF4A/eIF4G/eIF4H helicase complex using solution NMR.Citation8 This study provides insight into the mechanisms of regulation of the eIF4A helicase activity through modulating its affinity for ATP and conformational changes augmented by eIF4G and eIF4H. Since eIF4B and eIF4H appear to share a common binding site on eIF4A and have a similar function, these findings likely can be extrapolated to helicase complexes containing eIF4B instead of eIF4H.
The reports regarding the effects of eIF4B overexpression on translation rates in mammalian systems are controversial. Transient overexpression of eIF4B increases translation rates in some studiesCitation9,Citation10 whereas others have shown inhibition.Citation11–Citation13 Since eIF4B may have numerous binding partners, differences in expression levels may explain such discrepancies. A high level of eIF4B may disrupt endogenous protein complexes and thus create translationally inactive pools of eIF4A, eIF3 and 40S ribosomal subunits. Besides, a small molecule inhibitor of translation (Pateamine A) has been recently shown to act through excessive stimulation of eIF4A-associated activities.Citation14 As an eIF4A cofactor, overexpressed eIF4B is very likely to disturb the proper control of eIF4A helicase activity, which is necessary for efficient ribosome binding and translation initiation.Citation15
eIF4B Regulation by Signaling Cascades
Toe printing studies have underscored an important role for eIF4B in 48S initiation complex assembly even on messages with relatively unstructured 5′UTRs.Citation16 Interestingly, the recombinant factor purified from bacteria poorly substitutes for the native protein, suggesting that eIF4B activity depends on posttranslational modifications that are unique to eukaryotic cells. Indeed, eIF4B becomes phosphoryled in rabbit reticulocyte lysates.Citation17 Furthermore, eIF4B phosphorylation status changes in a serum and mitogen-dependent manner.Citation18,Citation19 For example, Ser422 on eIF4B can be phosphorylated by S6K1 through a ramapycin-senstive pathway.Citation13 The phosphomimetic mutant (Ser422Glu) and wild type exogenous proteins behave similarly in translation assays, whereas the non-phosphorylatable eIF4B Ser422Ala mutant is inert. Subsequently, it was demonstrated that the MAPK-RSK module is an additional regulator of Ser422 phosphorylation.Citation20 Given the very similar substrate consensus motif phosphorylated by S6K and RSK proteins, the involvement of RSKs in eIF4B Ser422 phosphorylation was confirmed by several pharmacological, biochemical, RNAi and genetic approaches. It was then shown that eIF4B phosphorylation was a result of the MAPK/RSK and mTOR/S6K pathways' interplay and could be subdivided in two phases: (1) an early but transient MAPK/RSK-dependent phase and (2) an mTOR/S6K-dependent phase which is latent but sustained (). Different groups have shown the significance of eIF4B Ser422 phosphorylation for eIF3 binding and stimulation of cap-dependent translation.Citation9,Citation10,Citation20,Citation21 The effects of interferon-α on translational activation of specific mRNAs and subsequent growth inhibition were also shown to be mediated by eIF4B Ser422 phosphorylation.Citation21 Interferon-α induces eIF4B Ser422 phosphorylation and interaction with eIF3a in normal MEFs but was ineffective in S6K1/2 double knockout MEFs. Interferon-α also induces ATPase activity associated with eIF4B in a S6K1/2-dependent manner. Mentioning that eIF4B was previously established to increase eIF4A ATPase and helicase activities, the authors raised the possibility that eIF4A could be the source of the measured ATPase activity. Noteworthy, eIF3a RNAi abolished the ATPase activity induction and the translation interferon-α-responsive mRNAs, suggesting that eIF4B phosphorylation and interaction with eIF3 were necessary for these events.
Phosphorylation of another eIF4B site, Ser406, is regulated by MAPK and PI3K/mTOR signaling cascades and is important for optimal translational activity.Citation10 eIF4B is phosphorylated on many other sites (see www.phosphosite.org) but the physiological significance of these modifications is yet to be discovered.
Effect of eIF4B on Cell Survival and Proliferation
Ectopic overexpression of eIF4B in Drosophila cultured cells and in developing eye imaginal discs stimulates cell proliferation, whereas eIF4B siRNA results in a minor inhibition of general translation but affected the survival of insect cells grown in reduced serum-containing media.Citation22 In mammalian cells, eIF4B silencing by RNAi in mammalian cells leads to polysome depletion and translational repression.Citation23 Both protein expression and mRNA polysomal distribution analysis support a model where mRNAs (such as bcl-2, XIAP, Cdc25C, c-myc and ODC) that harbor structured 5′UTRs are highly sensitive to eIF4B expression levels (). Consistent with the observed effects on specific mRNA translation, eIF4B knockdown leads to decreased cell survival and proliferation.Citation23 Compromised survival in eIF4B-silenced cells is reflected by molecular hallmarks of apoptosis such as pro-caspase-9 and PARP cleavage, DNA fragmentation and annexin V labeling. Interestingly, expression of the exogenous bcl-2 protein alone almost completely prevents DNA fragmentation in eIF4B-silenced cells. The latter effect suggests that bcl-2 mRNA is the major eIF4B-controlled translational target that plays a crucial role in cell survival. eIF4B knockdown leads to attenuated proliferation and this could be attributed to the combined effect of eIF4B depletion on survival and proliferative mRNAs. Noteworthy, eIF4B-silencing sensitizes cancer cells to low doses of the cytotoxic agent camptothecin. This effect also could be explained by dramatically decreased expression of anti-apoptotic proteins bcl-2 and XIAP. Depletion of either of these proteins has been previously proposed to compromise the viability of cancerous cells and sensitize them to cytotoxic drugs, allowing treatment with lower doses and hence decreasing systemic toxicity. Hence, it is plausible that development of small inhibitor molecules targeting eIF4B could have therapeutic value in cancer treatment.
Concluding Remarks
eIF4B is regulated by two major proto-oncogenic signaling pathways, Ras-MAPK and PI3-K/mTOR, which drive cell proliferation and control survival. When phosphorylated on Ser422, eIF4B binds more avidly to complexes containing ribosome bound eIF3. eIF4B phosphorylation induces ATPase activity, which most likely involves eIF4A. These effects alone are bound to activate translation of otherwise silent mRNAs harboring structured 5′UTRs. Intriguingly, all the identified mRNAs affected by eIF4B siRNA are involved in the control of either proliferation (Cdc25C, c-myc, ODC) or survival (bcl-2, XIAP). Hence, eIF4B represents one of the focal points whereby MAPK and mTOR signaling cascades converge to exert their effects on major biological processes such as proliferation and survival. Increased proliferation and survival are hallmarks of oncogenic transformation. Hence, eIF4B offers a potentially attractive target for development of anti-cancer therapies.
Future Directions
eIF4B plays a major role in the translation initiation most likely through stimulation of eIF4A helicase and ATPase activities, but potentially also through other mechanisms such as preventing initiation complex 3′–5′ movement and assisting in ribosome landing pad formation. The detailed mechanism of how this helicase activity occurs has not yet been elucidated and requires additional structural and kinetic experimentation. The vast majority of mRNAs harbor secondary structures in their 5′UTRs and hence require the energy-consuming helicase activity for efficient translation initiation. Many mRNAs with relatively structured 5′UTRs are transcribed from survival- and proliferation-related genes. The energy spent by helicases to unwind the 5′UTR of an mRNA is proportional to the extent of its structural complexity.Citation15 eIF4B has been shown to be dispensable for translation initiation on mRNAs with unstructured 5′UTR in vitro.Citation24 It is however unclear, whether ‘translationally strong’ mRNAs with relatively unstructured 5′UTRs (such as β-actin and GAPDH) can be translated in the complete absence of eIF4B in vivo. All these data emphasize an importance of eIF4B in translation initiation and suggest further studies targeting this protein in prevention and treatment of cancer.
Figures and Tables
Figure 1 eIF4B domain structure. Amino acid positions flanking the RRM, DRYG and ARM domains are indicated.
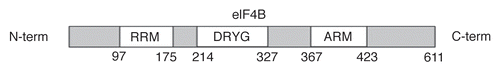
Figure 2 eIF4B activation by Ras-MAPK and PI3-K/mTOR signaling pathways.
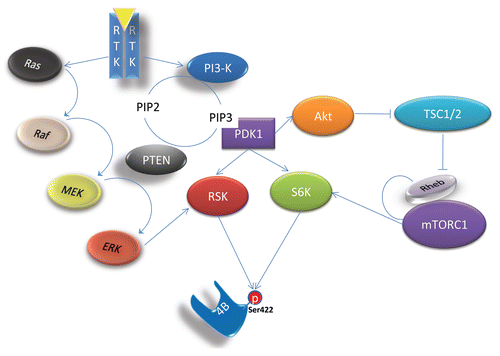
Figure 3 mRNAs harboring unstructured UTRs (A) are less dependent on elF4B than messages with complex 5′UTRs (B).
Acknowledgements
We apologize to our colleagues whose work was not included in this review due to the space limitations. We thank Colin Lister and Pam Kirk for technical support. A.P. is a recipient of Fonds de la Recherche en Santé du Québec postdoctoral award. N.S. is a Howard Hughes Medical Institute International scholar. This work was supported by a grant from the Canadian Cancer Research Institute to N.S.
References
- Rozen F, Edery I, Meerovitch K, Dever TE, Merrick WC, Sonenberg N. Bidirectional RNA helicase activity of eucaryotic translation initiation factors 4A and 4F. Mol Cell Biol 1990; 10:1134 - 1144
- Methot N, Pause A, Hershey JW, Sonenberg N. The translation initiation factor eIF-4B contains an RNA-binding region that is distinct and independent from its ribonucleoprotein consensus sequence. Mol Cell Biol 1994; 14:2307 - 2316
- Methot N, Song MS, Sonenberg N. A region rich in aspartic acid, arginine, tyrosine and glycine (DRYG) mediates eukaryotic initiation factor 4B (eIF4B) self-association and interaction with eIF3. Mol Cell Biol 1996; 16:5328 - 5334
- Rozovsky N, Butterworth AC, Moore MJ. Interactions between eIF4AI and its accessory factors eIF4B and eIF4H. RNA 2008; 14:2136 - 2148
- Methot N, Pickett G, Keene JD, Sonenberg N. In vitro RNA selection identifies RNA ligands that specifically bind to eukaryotic translation initiation factor 4B: the role of the RNA remotif. RNA 1996; 2:38 - 50
- Spirin AS. How does a scanning ribosomal particle move along the 5′-untranslated region of eukaryotic mRNA? Brownian Ratchet model. Biochemistry 2009; 48:10688 - 10692
- Lindqvist L, Imataka H, Pelletier J. Cap-dependent eukaryotic initiation factor-mRNA interactions probed by cross-linking. RNA 2008; 14:960 - 969
- Marintchev A, Edmonds KA, Marintcheva B, Hendrickson E, Oberer M, Suzuki C, et al. Topology and regulation of the human eIF4A/4G/4H helicase complex in translation initiation. Cell 2009; 136:447 - 460
- Holz MK, Ballif BA, Gygi SP, Blenis J. mTOR and S6K1 mediate assembly of the translation preinitiation complex through dynamic protein interchange and ordered phosphorylation events. Cell 2005; 123:569 - 580
- van Gorp AG, van der Vos KE, Brenkman AB, Bremer A, van den Broek N, Zwartkruis F, et al. AGC kinases regulate phosphorylation and activation of eukaryotic translation initiation factor 4B. Oncogene 2009; 28:95 - 106
- Milburn SC, Hershey JW, Davies MV, Kelleher K, Kaufman RJ. Cloning and expression of eukaryotic initiation factor 4B cDNA: sequence determination identifies a common RNA recognition motif. EMBO J 1990; 9:2783 - 2790
- Naranda T, Strong WB, Menaya J, Fabbri BJ, Hershey JW. Two structural domains of initiation factor eIF-4B are involved in binding to RNA. J Biol Chem 1994; 269:14465 - 14472
- Raught B, Peiretti F, Gingras AC, Livingstone M, Shahbazian D, Mayeur GL, Sonenberg N. Phosphorylation of eucaryotic translation initiation factor 4B Ser422 is modulated by S6 kinases. EMBO J 2004; 23:1761 - 1769
- Bordeleau ME, Matthews J, Wojnar JM, Lindqvist L, Novac O, Jankowsky E, Sonenberg N. Stimulation of mammalian translation initiation factor eIF4A activity by a small molecule inhibitor of eukaryotic translation. Proc Natl Acad Sci USA 2005; 102:10460 - 10465
- Svitkin YV, Pause A, Haghighat A, Pyronnet S, Witherell G, Belsham GJ, Sonenberg N. The requirement for eukaryotic initiation factor 4A (elF4A) in translation is in direct proportion to the degree of mRNA 5′ secondary structure. RNA 2001; 7:382 - 394
- Dmitriev SE, Terenin IM, Dunaevsky YE, Merrick WC, Shatsky IN. Assembly of 48S translation initiation complexes from purified components with mRNAs that have some base pairing within their 5′ untranslated regions. Mol Cell Biol 2003; 23:8925 - 8933
- Benne R, Edman J, Traut RR, Hershey JW. Phosphorylation of eukaryotic protein synthesis initiation factors. Proc Natl Acad Sci USA 1978; 75:108 - 112
- Duncan R, Hershey JW. Regulation of initiation factors during translational repression caused by serum depletion. Covalent modification. J Biol Chem 1985; 260:5493 - 5497
- Morley SJ, Traugh JA. Phorbol esters stimulate phosphorylation of eukaryotic initiation factors 3, 4B and 4F. J Biol Chem 1989; 264:2401 - 2404
- Shahbazian D, Roux PP, Mieulet V, Cohen MS, Raught B, Taunton J, et al. The mTOR/PI3K and MAPK pathways converge on eIF4B to control its phosphorylation and activity. EMBO J 2006; 25:2781 - 2791
- Kroczynska B, Kaur S, Katsoulidis E, Majchrzak-Kita B, Sassano A, Kozma SC, et al. Interferon-dependent engagement of eukaryotic initiation factor 4B via S6 kinase (S6K)- and ribosomal protein S6K-mediated signals. Mol Cell Biol 2009; 29:2865 - 2875
- Hernandez G, Vazquez-Pianzola P, Zurbriggen A, Altmann M, Sierra JM, Rivera-Pomar R. Two functionally redundant isoforms of Drosophila melanogaster eukaryotic initiation factor 4B are involved in cap-dependent translation, cell survival and proliferation. Eur J Biochem 2004; 271:2923 - 2936
- Shahbazian D, Parsyan A, Petroulakis E, Topisirovic I, Martineau Y, Gibbs BF, et al. Control of cell survival and proliferation by mammalian eukaryotic initiation factor 4B. Mol Cell Biol 2010; 30:1478 - 1485
- Pestova TV, Kolupaeva VG. The roles of individual eukaryotic translation initiation factors in ribosomal scanning and initiation codon selection. Genes Dev 2002; 16:2906 - 2922