Abstract
Casein Kinase I (CKI) is a conserved component of the Wnt signaling pathway, which regulates cell fate determination in metazoans. We show that post-embryonic asymmetric division and fate specification of C. elegans epidermal stem cells are controlled by a non-canonical Wnt/b-catenin signaling pathway, involving the b-catenins WRM-1 and SYS-1, and that C. elegans kin-19/CKIa functions in this pathway. Furthermore, we find that kin-19 is the only member of the Wnt asymmetry pathway that functions with, or in parallel to, the heterochronic temporal patterning pathway to control withdrawal from self-renewal and subsequent terminal differentiation of epidermal stem cells. We show that, except in the case of kin-19, the Wnt asymmetry pathway and the heterochronic pathway function separately and in parallel to control different aspects of epidermal stem cell fate specification. However, given the function of kin-19/CKIa in both pathways, and that CKI, Wnt signaling pathway and heterochronic pathway genes are widely conserved in animals, our findings suggest that CKIa may function as a regulatory hub through which asymmetric division and terminal differentiation are coordinated in adult stem cells of vertebrates.
Introduction
Asymmetric cell division (AD) and self-renewal (SR) are defining characteristics of stem cells. In asymmetric division, the progenitor stem cell undergoes mitosis to give rise to progeny cells that have different fates. One daughter cell differentiates, thus becoming more restricted in its cell fate potential, and typically exits the cell cycle. The fate of the other daughter cell is self-renewal, that is it retains proliferative capability and maintains the cell fate potential of the progenitor cell. The cell behaviors of AD and SR allow stem cells to populate or re-populate, the differentiated cells of a tissue while maintaining multipotent stem cell lineages, and these behaviors are fundamental to the processes of both tissue formation during development and tissue regeneration and repair.Citation1,Citation2 The C. elegans seam cells are multipotent epidermal stem cells that undergo AD and SR during postembryonic larval stages, and serve as a model for investigating stem cell development.Citation3,Citation4
The C. elegans seam cells (H0-H2, V1-6, T) undergo asymmetric divisions at each of the four larval stages (L1-L4) to produce self-renewing seam cells and differentiated hypodermal cells. For most seam cell lineages, the posterior daughter cell becomes the self-renewing seam cell while the anterior daughter cells terminally differentiates by fusing with the surrounding epidermal snycytium (Hyp7).Citation5 In the V5 and T lineages, the non-self renewing daughter cells undergo further divisions in the L2 stage and differentiate into neural cells and neuronal support cells.Citation5 The self-renewing seam cells themselves terminally differentiate at the end of larval development, as is the case for all somatic lineages in C. elegans. In addition to asymmetric divisions at every larval stage, the V1 to V6 seam cell lineages also undergo one round of symmetric division during the L2 stage, thus increasing the seam cell number.
Two genetic pathways have been shown to regulate seam cell development: The heterochronic pathway controls stage specific patterns of seam cell division at each larval stage, as well as seam cell loss of self-renewal at the end of larval development that results in terminal differentiation, while C. elegans homologs of the Wnt/β-catenin signaling have been implicated in the control of asymmetric division and cell fate determination of the V5.p and T seam cells.Citation6–Citation8 Additionally, ceh-16, the homolog of engrailed, is a transcriptional regulator that represses genes involved in seam cell terminal differentiation and activates genes that maintain seam cell fate.Citation9 The L2-specific divisions are regulated by RNT-1 and BRO-1 transcription factors, the C. elegans homologs of Runx and its binding partner CBFβ, respectively, which control hematopoietic stem cell development in vertebrates, and by sel-6 (also known as emb-4 and mal-2), which encodes a splicesomal protein that regulates germline chromosome condensation during embryonic development.Citation10–Citation12 The heterochronic pathway and the Wnt signaling pathways function at every larval stage, and mutations in either of these pathways affect both seam cell division and differentiation patterns, raising the possibility that there is cross-regulation between these pathways.Citation13,Citation14 In this study we sought to identify new components of seam cell Wnt signaling and heterochronic pathways, focusing especially on candidates with potential dual function in both pathways. Additionally, we sought to define the regulatory relationship, if any, between the Wnt signaling and heterochronic pathways in regulation of seam cell asymmetric division and self-renewal.
The Casein kinase I (CKI) family of serine/threonine kinases functions in β-catenin dependent Wnt signaling pathways, which are broadly conserved in animal species and regulate numerous aspects of cell fate determination, including stem cell development.Citation2,Citation15 In the canonical Wnt pathway of Drosophila and vertebrate species, CKIα functions as part of a cytoplasmic ‘destruction complex’ that includes the scaffold proteins AXIN and APC, and GSK3β kinase.Citation16 This ‘destruction complex’ regulates the stability of β-catenin, which is phosphorylated in the absence of Wnt signaling and thus targeted for ubiquitylation and degradation. When Wnt signals are detected by Frizzled/LRP co-receptor complexes, and the signal is transduced through cytoplasmic Dishevelled (Dsh), β-catenin phosphorylation by the ‘destruction complex’ is inhibited allowing stabilized β-catenin to enter the nucleus, where it functions with Tcf/Lef transcription factors to induce expression of Wnt signaling pathway target genes.Citation15 Thus, in the current model of canonical Wnt signaling, different cell fates in the daughter cells of asymmetric division depends on differential ability to receive and transduce Wnt signals, and on the differential regulation of Wnt pathway components, such as CKIα and CKIε/δ, which control the stability and function of β-catenin and Tcf/Lef.
In C. elegans, the asymmetric cell fate choice that occurs upon seam cell division is also controlled by β-catenin dependent Wnt signaling pathways, and specifically by differential levels of POP.1, the C. elegans Tcf/Lef homolog.Citation8,Citation14 Low levels of POP-1/Tcf in the nucleus characterize the posterior daughter cells, which assume seam cell fate, while high levels of nuclear POP-1/Tcf characterize anterior daughter cells, which assume hypodermal cell fate.Citation17 However, the molecular events upstream of POP-1/TCF differ from the canonical Wnt signaling pathway of vertebrates. Unlike the known animal Wnt signaling pathways, which utilize a single form of β-catenin, C. elegans Wnt signaling pathways utilize four different β-catenins encoded by bar-1, sys-1, wrm-1 and hmp-2.Citation18–Citation20 BAR-1 and SYS-1 bind directly to POP-1 and are termed canonical β-catenins, while WRM-1 and HMP-2, which do not directly bind POP-1, are termed non-canonical β-catenins. However, WRM-1, in complex with LIT-1, signals indirectly through POP-1 by regulating its nuclear export.Citation19,Citation21 The BAR-1/β-catenin dependent Wnt signaling pathway is similar in molecular components and mechanism to the vertebrate canonical Wnt pathway, and regulates a number of cell fate determination processes during larval development.Citation7,Citation8 For example, in the V5 seam cell lineage, the anterior daughter cell of one of the L2 divisions gives rise to a sensory structure called a postdereid rather than becoming a hypodermal cell, and this postdereid cell fate is dependent on bar-1/β-catenin.Citation22 The C. elegans BAR-1 dependent canonical Wnt signaling pathway appears to function primarily in cell fate specification that does not involve asymmetric division, which is instead controlled by a non-canonical Wnt signaling pathway that utilizes the WRM-1 and SYS-1 β-catenins.Citation8 In T seam cells, this ‘Wnt/β-catenin asymmetry pathway’ regulates seam versus hypodermal cell fate by controlling the ratio of nuclear POP-1 to SYS-1 ().Citation8 In the anterior cells, which will become hypodermis, nuclear export of WRM-1 and LIT-1 results in high nuclear POP-1 levels and a high POP-1 to SYS-1 ratio, enabling POP-1 function as a transcriptional repressor (). In the posterior cells, which retain seam cell identity, nuclear export of WRM-1 and LIT-1 is inhibited, and activation of the WRM-1/LIT-1 complex leads to nuclear export of POP-1. The resulting drop in the POP-1 to SYS-1 ratio allows the formation of a POP-1/SYS-1 complex that converts POP-1 from a transcriptional repressor to a transcriptional activator of Wnt signaling pathway genes ().Citation8,Citation20,Citation23 The Wnt/β-catenin asymmetry pathway has been modeled from studies in the T seam cells only. Thus, it is not yet known whether this wrm-1/sys-1 dependent pathway also controls the reiterative asymmetric divisions of the other seam cells, or is a T seam cell-specific mechanism.
The genes of the heterochronic developmental timing pathway control both stage-specific seam cell division patterns as well as loss of seam cell self-renewal capacity in terminal differentiation. A recurring molecular mechanism in the heterochronic pathway is the stage specific expression of microRNAs that downregulate translational expression of target gene products, and thus allows progression to the next developmental stage.Citation4,Citation13 There are three occurrences of a stage-specific ‘microRNA and target’ mechanism during larval development: lin-4 microRNA downregulates lin-14 and lin-28 during L1 and L2 stages, the mir-48, mir-84 and mir-241 let-7 family microRNAs function redundantly to downregulate hbl-1 during the L2 stage, and let-7 microRNA downregulates a number of gene transcripts, including lin-41, during the L4 stage.Citation4,Citation13 In each case, loss of function mutation of the microRNA results in retarded seam cell development that involves lack of developmental progression and reiteration of stage specific seam cell division, as well as a delay in or lack of terminal differentiation. For example, triple mutation of the mir-48, mir-84 and mir-241 let-7 family microRNAs results in L3 reiteration of the L2 stage specific seam cell amplifying divisions and loss of seam cell terminal differentiation.Citation24 Conversely, loss of function mutation of microRNA target genes results in precocious seam cell development, which involves exit from the self-renewal cell division cycle and premature terminal differentiation. For example, mutation of the let-7 microRNA target genes lin-41, hbl-1 or lin-42 results in premature terminal differentiation during the L3 and L4 stages. Thus, the normal function of the L3/L4 stage heterochronic genes that are microRNA targets appears to be the maintenance of seam cell identity, which includes self-renewal (). Vertebrate homologs of the C. elegans heterochronic genes also regulate stem cell development.Citation3,Citation4 For example, lin-28 is one of five factors that work together to induce stem cell pluripotency in differentiated somatic cells.Citation4,Citation25,Citation26 lin-28 specifies stem cell pluripotency and self-renewal by inhibiting microRNA processing, including that of let-7 microRNAs, thus preventing expression of mature microRNAs that induce cellular differentiation.Citation26–Citation28 let-7 microRNA target genes have known functions in stem cell regulation. The mouse homolog of LIN-41 (mLIN-41) is a ubiquitin ligase that functions with LIN-28 to suppress let-7 microRNA activity in stem cells.Citation29 In another example, both Drosophila hunchback and mouse ikaros, the homologs of C. elegans hbl-1, function to maintain multipotency of neural stem cells.Citation30,Citation31
In Drosophila and vertebrates, Casein kinase I epsilon and delta isoforms (CKIε/δ) function in cell cycle regulation and act as positive regulators of the canonical β-catenin dependent Wnt signaling pathway.Citation32 CKI function in the C. elegans Wnt signaling pathways has not been definitively determined, but kin-20, the sole CKIε/δ homolog in C. elegans, functions in the seam cell heterochronic pathway.Citation33 kin-20 loss of function mutation results in premature seam cell terminal differentiation during the L4 stage, and thus kin-20/CKIε/δ functions with or in parallel to the L3/L4 stage precocious heterochronic genes, lin-41, lin-42 and hbl-1 (). The vertebrate and Drosophila homologs of kin-20 and lin-42, CKIε/δ and period respectively, function together in the broadly conserved circadian timing pathway, and genetic analyses suggest that kin-20 and lin-42 may have a similar regulatory relationship in the seam cell heterochronic pathway.Citation33,Citation34 A further similarity between heterochronic and circadian timing pathway is that the circadian pathway has been shown to regulate the timing of hematopoetic and neural stem cell proliferation and maturation, although the mechanisms of this regulation are not known.Citation35–Citation37 Since both kin-20 and lin-42 are expressed in the seam cells throughout larval development, these CKIε/δ and period homologs may play a reiterative role in controlling seam cell asymmetric division and self-renewal, either together with or in parallel to the Wnt asymmetry pathway.
Based on the observation that CKI isoforms function in both spatial and temporal patterning pathways to regulate stem cell development, we hypothesized that cross-regulation between the Wnt and heterochronic pathways may be mediated by CKI. To better understand the roles played by CKI in different cell fate determination pathways, we investigated the function of kin-19/CKIα during post-embryonic seam stem cell development in C. elegans.
Results
kin-19/CKIα is expressed in the epidermal seam cells throughout larval development.
The casein kinase I (CKI) family in C. elegans consists of four genes encoding three different isoforms: kin-19/CKIα, kin-20/CKIε/δ, csnk-1/CKIγ and spe-6/CKIγ. We have previously reported that kin-20/CKIε/δ is expressed in the seam cells throughout larval development and functions in the heterochronic pathway to regulate seam cell terminal differentiation, while csnk-1 and spe-6 either are not expressed, or do not function, in the seam cells.Citation33 Since both CKIα and CKIε/δ are known to function in the vertebrate canonical Wnt signaling pathway, we investigated whether kin-19/CKIα is also expressed in the seam cells and/or hypodermis. Drosophila and vertebrate CKIα genes encode multiple splice variants that have different functions,Citation38 but C. elegans kin-19 is expressed as a single transcript that is expressed at a relatively constant level throughout larval development (). We assayed KIN-19 protein expression using a kin-19p::kin-19::gfp translational reporter in wild-type N2 worms, and observed GFP expression in the epidermal seam cells, the vulval cells and in the ventral nerve cord at all larval stages (). We observed KIN-19::GFP expression in both nucleus and cytoplasm of seam cells, and we noticed the formation of a punctate GFP pattern in the cytoplasm. This punctate expression pattern may be indicative of the localization of KIN-19 to cytoplasmic macromolecular complexes, similar to the localization of CKIα to the ‘destruction complex’ in the canonical Wnt signaling pathway. We did not notice any obvious subcelluar asymmetry (nuclear versus cytosolic or anterior versus posterior of single seam cells) or cellular asymmetry (anterior versus posterior daughter cell after mitosis) in KIN-19::GFP expression during the L4 stage.
kin-19/CKIα regulates anterior-posterior spatial patterning of seam cell fates.
In the early C. elegans embryo, MOM-2/Wnt-signaling polarizes the EMS cell that divides asymmetrically into the E blastomere, which gives rise to endoderm, and the MS blastomere, which gives rise to mesoderm. Disruption of MOM-2/Wnt-signaling results in embryonic lethality due to the transformation of E blastomere to M blastomere, and the production of excess mesoderm at the expense of endoderm.Citation39 Embryonic lethality due to this ‘more mesoderm’ (mom) phenotype is also produced by kin-19 mutation or RNA interference (RNAi), and suggests that kin-19 is a positive regulator of the asymmetry Wnt signaling pathway in embryos.Citation40 In order to assess whether kin-19 plays a similar role during post-embryonic development, we by-passed embryonic lethality by kin-19(RNAi) starting in larval stage 1 (L1) animals, and observed the effect on gross development and on epidermal seam cell development. At the gross anatomical level, kin-19(RNAi) resulted in adult animals that were thin and elongated in shape, had protruding vulva (Pvl), and were sterile (data not shown). The Pvl phenotype suggested to us that improper specification of vulval cell fates occurs on knockdown of kin-19 in larval animals, while the elongated body phenotype could have arisen from cell fate transformation that resulted in duplication of the epidermal cell types that determine nematode body length.
For a more detailed analysis, we followed seam cell development using a C. elegans strain (wIs78) in which the seam cell nuclei and adherens junctions are marked by GFP in wildtype genetic background. We observed kin-19(RNAi) animals throughout larval development and detected the formation of excess seam cells during the fourth larval (L4) and adult stages ( and B and and B). In wIs78 worms, 16 GFP-marked seam cells enter the L4 stage and synchronously go through one round of mitosis during mid-L4 producing 29 cells ( and ). The anterior daughter cells migrate away from the seam and differentiate by fusing with the Hyp7 hypodermal syncytium, and thus lose seam cell specific GFP staining. The posterior daughter cells, which retain seam cell identity, terminally differentiate by fusing to form the seam syncytium, and secrete collagenous ridges called alae. Thus, at the end of the L4 stage in wIs78 animals, 16 GFP-marked seam cell nuclei are present within the seam syncytium, whose boundaries are also marked by GFP (). In kin-19(RNAi) animals, the wild type number of seam cells was observed during the early L3 stages, but an average of 19 seam cells, instead of 14, were seen to enter the L4 stage (). This number increased to an average of 28 seam cells in the late L4 stage kin-19(RNAi) animals and persisted into the adult stage ( and ).
The supernumerary seam cells in kin-19(RNAi) animals could have resulted from the failure of anterior daughter cells to migrate out of the seam, that is the Hyp7 fated daughters of seam cell division may have undergone an anterior to posterior cell fate transformation and thus aberrantly retained seam cell identity. This type of hypodermal-to-seam cell fate would result in 27 adult seam cells instead of 14 (). Indeed, we observed that an average of 28 GFP-marked cells persisted from the mid L4 into the adult stage, and that they behaved like normal seam cells, that is they fused during the late L4 and secreted alae during the early adult stage. However, since we occasionally observed kin-19(RNAi) worms with more than 28 seam cell nuclei (up to a maximum of 34 seam cell nuclei), a few of the seam cell must have divided again after the mid-L4 division. An alternative explanation to seam cell fate transformation could be that the normal developmental timing program was retarded in kin-19(RNAi) worms, such that the seam cells divided a second time immediately after the mid-L4 division instead of undergoing terminal differentiation. An second round of synchronized seam cell division in the mid L4 stage would result in approximately 40 GFP-marked nuclei being visible in the mid to late L4 stages, and retardation of terminal differentiation would also result in lack of seam cell fusion and lack of adult alae secretion. We observed a maximum of only 34 seam cell nuclei in kin-19(RNAi) animals ( and ). Moreover, seam cell fusion and alae secretion in kin-19(RNAi) animals took place at the appropriate time, during the late L4 transition to young adult ( and ). Therefore, the extra seam cells in kin-19(RNAi) animals most likely did not arise from an extra round of synchronized cell division in the L4 stage, and kin-19(RNAi) does not result in a retarded developmental timing defect in seam cell L4 division or terminal differentiation. Instead, our observations are consistent with the hypothesis that kin-19(RNAi) causes cell fate transformation from anterior hypodermal fate to posterior seam cell fate during postembryonic development (), similar to alteration of endodermal to mesodermal fate on kin-19(RNAi) during early embryonic development.
A non-canonical Wnt/β-catenin pathway dependent on wrm-1 and sys-1 β-catenins controls seam cell asymmetric division.
Seam cell fate specification is correlated with asymmetric nuclear distribution of POP-1/Tcf between anterior and posterior post-mitotic daughter cells: POP-1 accumulation in the nucleus of the anterior daughter cells confers non-seam cell/hypodermal identity, while lower levels of nuclear POP-1 in the posterior daughter cells specify seam cell fateCitation17 (). Thus, we would predict that net reduction of POP-1 in all seam cell progeny would cause cells that were normally fated for hypodermis to be transformed to the seam cell fate. Indeed, we observed excess post-embryonic seam cells formed on RNAi of pop-1/Tcf, very similar to the seam cell phenotype of kin-19(RNAi) animals ( and ). The supernumerary seam cell phenotype of pop-1(RNAi) was more pronounced than that of kin-19(RNAi), and an average of 52 seam cell nuclei were observed in the late L4 and adult stages ( and ). This doubling of the seam cell nuclei numbers in the mid L4 stage suggests that the seam cells undergo a second synchronous division after the mid-L4 division (). The observation that an average of 52 seam cell nuclei persisted into the adult stage, instead of decreasing in number, indicates that a hypodermal-to-seam cell fate transformation occurs in pop-1(RNAi) animals (). As with kin-19(RNAi), pop-1(RNAi) did not result in a developmental timing defect as seam cell fusion and alae secretion in these worms occurred with wild type timing ( and ). The similarity between the kin-19 and pop-1 RNAi seam cell phenotypes, together with previous reports that kin-19 acts in the early embryo Wnt/β-catenin asymmetry pathwayCitation40 and that CKIα functions as a negative regulator of Wnt signaling in vertebrates and invertebrates,Citation41 suggests that kin-19/CKIα functions with pop-1/TCF in a Wnt signaling pathway to regulate asymmetric division of post-embryonic seam cells.
In order to define the Wnt/β-catenin pathway that kin-19 functions in, we first sought to identify the Wnt signaling pathway responsible for controlling asymmetric division of V1-4,6 seam cells. Wnt signaling pathways have been shown to regulate asymmetric division of V5.p and T seam cells.Citation8,Citation14 In the V5.p lineage, a Wnt signaling pathway dependent on bar-1/β-catenin is required for the anterior daughter cell of the first L2 division to express postdeireid neural cell fate instead of hypodermal fate, while a Wnt signaling pathway dependent on wrm-1/β-catenin regulates asymmetric division of the T seam cell and its progeny in the L1 and L2 stages.Citation22,Citation42 However, it is not known which β-catenins are involved in regulating non-T and non-V5 seam cell fates. Using post-embryonic RNAi, we observed the effect on seam cell development of knocking-down each of the C. elegans β-catenins. RNAi of bar-1, sys-1 or hmp-2 did not have any discernable effect on V seam cell numbers divisions or terminal seam cell number and terminal differentiation (). However, we found that knockdown of the non-canonical β-catenin wrm-1, or of its associated MAP kinase lit-1, resulted in fewer than normal seam cell nuclei ( and ). The normal number of seam cells was observed to enter the L2 stage in both wrm-1(RNAi) and lit-1(RNAi) worms, but the number of GFP-marked cells dropped during the late L2, L3 and L4 stages, and only 1 to 2 seam cells were visible by the late L4 to adult stages. MOM-4/MAPKKK phosphorylates and activates the WRM-1/LIT-1 complex,Citation43 and mom-4(RNAi) also resulted in loss of seam cells, although the phenotype was less pronounced and seam cell loss was not observed until the L4 stage (). These phenotypes suggest that there was transformation of cell fate from posterior seam cell identity to anterior Hyp7 identity in wrm-1(RNAi), lit-1(RNAi) and mom-4(RNAi) worms (). In lit-1(RNAi) animals, the seam cell number increased modestly during the mid-L4 indicating that some of the seam cells had divided, but the observation that the final seam cell number dropped down to approximately two cells suggests that nearly all the posterior daughter cells of seam cell division underwent fate transformation to hypodermal fate (). This anteriorization of cell fate is opposite to the phenotype seen on RNAi of either kin-19 or pop-1, and is consistent with the current model of the WRM-1/SYS-1 dependent Wnt/β-catenin asymmetry pathway proposed for T seam cells ().Citation8 According to the current model, in the posterior post-mitotic daughter cell WRM-1, LIT-1 and MOM-4 function together to phosphorylate POP-1 and induce its nuclear export, thus reducing the POP-1 to SYS-1 nuclear ratio and allowing the formation of the POP-1/SYS-1 transcriptionally active complex ().Citation21,Citation44,Citation45 A prediction from this model is that RNAi knockdown of wrm-1, lit-1 or mom-4 would lead to net increase of nuclear POP-1 in all seam cell progeny, thus prevent the formation of the POP-1/SYS-1 transcriptional complex, and cause cells normally fated for seam cell identity to be transformed to hypodermis—as is observed in our experiments.
To test if C. elegans Wnt pathway genes interact in a manner consistent with function of the T seam cell Wnt/β-catenin asymmetry pathway in V seam cells, we assayed the effect of simultaneous or double RNAi (dRNAi) of pairs of Wnt pathway genes on seam cell asymmetric division (). Like kin-19(RNAi), single RNAi of pop-1, apr-1 or pry-1 resulted in excess seam cells, albeit to varying degrees, and we thus termed these genes seam cell inducers. Double RNAi of pop-1 with pry-1 or apr-1 resulted in pop-1(RNAi) phenotype, while dRNAi of pry-1 and apr-1 resulted in apr-1(RNAi) phenotype. These genetic interactions indicate that APR-1 may function downstream of PRY-1, and that PRY-1 and APR-1 normally function to increase nuclear POP-1 levels in the anterior daughter cell. Indeed, PRY-1 is localized to the cortex and serves as a scaffold for cortical APR-1, WRM-1 and LIT-1, while APR-1 has been observed to shuttle between the cortex and nucleus to aid in the nuclear export of WRM-1 and thus inhibit nuclear export of POP-1.Citation44,Citation46 According to the current model of the Wnt/β-catenin asymmetry pathway, hypodermal fate is determined by high nuclear POP-1 and a high POP-1:SYS-1 nuclear ratio, which can be generated by either increasing the amount of POP-1 or decreasing the amount of SYS-1 in the nucleus. Thus, the model predicts that loss of SYS-1 would result in fate transformation from seam cell to hypodermal fate. We did not observe any effect on seam cell asymmetric division with RNAi of sys-1 alone ( and and ). However, dRNAi of sys-1 with pop-1, apr-1 or pry-1 suppressed the excess seam cell phenotype of the seam cell inducer genes, while dRNAi of sys-1 with mom-4 further enhanced the loss of seam cell phenotype of mom-4(RNAi). Both these sets of dRNAi results indicate that sys-1 normally functions in the V seam cells to positively regulate seam cell fate, and is consistent with SYS-1 function as a transcriptional coactivator of POP-1 in T seam cells. We also found that single RNAi of the Dishevelled homologs dsh-1 and dsh-2 did not have any effect on seam cell asymmetric division ( and ). However, dRNAi of these genes in combination with sys-1(RNAi) resulted in a small decrease in the terminal number of seam cells, while dRNAi of these genes in combination with apr-1(RNAi) resulted in suppression of the excess seam cell phenotype (). Both these interactions indicate transformation of cell fate from seam cell to hypodermal cell, and are consistent with a model in which nuclear SYS-1 levels decrease due to inhibition of nuclear import that is normally mediated via Dishevelled molecules.Citation47,Citation48 We further determined that neither bar-1/β-catenin nor hmp-2/β-catenin interact with any of the Wnt asymmetry pathway genes or other known C. elegans Wnt pathway genes to regulate seam cell asymmetric division (). Together, our data indicate that asymmetric division of both V seams cells and the T seam cells is controlled by a non-canonical Wnt/β-catenin that involves WRM-1 and SYS-1 β-catenins as well as the atypical MAPK pathway kinases LIT-1 and MOM-4.
The non-canonical Wnt/β-catenin asymmetry pathway in seam cells may function cell autonomously.
In the canonical model of the Wnt signaling pathway in vertebrates, inhibition of the ‘destruction complex’, subsequent stabilization and nuclear import of β-catenin and expression of Wnt pathway target genes by the β-catenin/Tcf transcription complex all depend on reception and transduction of WNT signals by WNT receptors. In C. elegans, there are 5 known Wnt genes (cwn-1, cwn-2, egl-20, lin-44, mom-2) and 4 known Wnt receptors (lin-17, lin-18, mig-1 and mom-5). While the downstream molecules of the Wnt/β-catenin asymmetry pathway are conserved in the EMS blastomere and T seam cell, asymmetric division in the EMS blastomere is signaled through reception of MOM-2/Wnt by MOM-5/Wnt receptor, while asymmetric division in the T seam cell is signaled through reception of LIN-44/Wnt by LIN-17/Wnt receptor.Citation39,Citation49,Citation50 In both cell types, proteins that localize asymmetrically in the cell cortex of the progenitor cell before mitosis, or in one of the daughter cells after mitosis, are thought to connect the Wnt signal received by the receptor to downstream transcriptional regulation mediated by POP-1. Specifically, Wnt signaling in the T seam cell asymmetry pathway has been postulated to activate Dsh-homolog mediated nuclear import of SYS-1, and inhibit PRY-1 and APR-1 mediated export and cortical localization of WRM-1, thus in effect decreasing the nuclear POP-1:SYS-1 ratio and allowing transcription by the POP-1/SYS-1 complex ().Citation20,Citation44–Citation46 In order to determine which Wnts and Wnt receptors regulate the Wnt/β-catenin asymmetry pathway in the V seam cells, we assayed the effect of single and double RNAi of Wnt, Wnt receptor and Wnt signaling pathway genes on seam cell asymmetric division. In addition, we tested the effect of RNAi of mig-14, which is expressed in Wnt secreting cells.Citation51 On single RNAi in wIs78 animals, none of the Wnts, Wnt receptors or mig-14 affected seam cell asymmetric division (). Double RNAi with various combinations of the Wnt and Wnt receptor genes also had no effect on normal seam cell asymmetric division (not shown). On double RNAi with sys-1, RNAi of the Wnt receptors genes lin-17, lin-18 and mom-5 resulted in a modest reduction in terminal seam cell numbers, while double RNAi of the same Wnt receptors genes with mom-4 weakly enhanced the loss of seam cell phenotype (significance 0.15 < p < 0.3). These results together suggest, but do not conclusively demonstrate, that cell extrinsic Wnt signaling via Wnt and Wnt receptors does not regulate the seam cell non-canonical Wnt/β-catenin asymmetry pathway, which may function cell autonomously.
kin-19/CKIα functions in the seam cell non-canonical Wnt/β-catenin asymmetry pathway.
In order to determine whether kin-19 functions with wrm-1 and lit-1, and what the functional genetic position of kin-19 is relative to these Wnt pathway genes, we performed semi-epistasis experiments using simultaneous RNAi of kin-19 with each of the Wnt pathway genes (). Consistent with our findings that the canonical β-catenin bar-1 and the non-canonical β-catenin hmp-2 do not appear to regulate V seam cell fate, RNAi of either of these genes had no effect on the supernumerary seam cell phenotype of kin-19(RNAi). An evolutionarily conserved feature of the β-catenin dependent canonical Wnt signaling pathway is the dual kinase regulation of β-catenin stability by CKIα and GK3β in the ‘destruction complex’.Citation2,Citation52 In vertebrates, CKIα phosphorylation of β-catenin is primed by CKIα phosphorylation by GK3β, which also phosphorylates β-catenin. Indeed the conjunction of GK3β and CKI activities has been detected in a number of other pathways, such the Hedgehog (Hh) pathway in which GK3β and CKI control the stability of Cubitus (Ci) transcription factor in response to Hh signaling, and the Drosophila and vertebrate circadian pathways.Citation53,Citation54 However, gsk-3(RNAi) resulted in the normal terminal seam cell number, and dRNAi of kin-19 and gsk-3 resulted in the kin-19(RNAi) phenotype (). Similarly, dRNAi of gsk-3 with other genes of the asymmetric Wnt pathway did not have any effect on seam cell development beyond the phenotypes of single Wnt gene RNAi (). Thus, gsk-3 does not appear to function, either alone, with kin-19 or with Wnt signaling pathway genes, to regulate seam cell asymmetric division.
Single RNAi of the sys-1/β-catenin alone did not have a discernable effect on seam cell development, but dRNAi of sys-1 and kin-19 resulted in partial suppression of the kin-19(RNAi) phenotype, suggesting that sys-1 functions downstream of kin-19 ( and ). Dishevelled molecules are thought to mediate nuclear import of SYS-1, thus enabling SYS-1 transcriptional function in complex with POP-1.Citation47,Citation48 Consistent with this function of Dsh, and with the epistatic relationship between sys-1 and kin-19, dRNAi of dsh-1 or dsh-2 and kin-19 resulted in suppression of the kin-19(RNAi) phenotype (). We further found that RNAi of wrm-1, lit-1 or mom-4 completely or partially suppressed the seam cell fate posteriorization phenotype of kin-19(RNAi), indicating that these Wnt pathway genes also function downstream of kin-19 ( and ). On the other hand, dRNAi of kin-19 with pop-1 resulted in the pop-1(RNAi) phenoype, dRNAi with apr-1 resulted in modest enhancement of the kin-19(RNAi) phenotype, and dRNAi with pry-1 resulted in kin-19(RNAi) phenotype ( and ). These results suggest that pop-1 functions downstream of kin-19, while apr-1 functions in parallel with, and pry-1 upstream of kin-19. Simultaneous RNAi of kin-19 with any of the five Wnt genes, with any of the four Wnt receptor genes, or with mig-14 resulted in the kin-19(RNAi) asymmetric seam cell division phenotype ( and ), suggesting that kin-19 regulation of seam cell asymmetric division is not dependent on cell extrinsic Wnt signaling. Together, the results from our dRNAi experiments of kin-19 with Wnt signaling pathway genes indicate that during C. elegans post-embryonic development, asymmetric division of seam cells is regulated by kin-19 function in a non-canonical Wnt/β-catenin asymmetry signaling pathway that involves the sys-1 canonical β-catenin, the wrm-1 non-canonical β-catenin, and the lit-1 and mom-4 MAP kinases, but does not involve gsk-3 kinase.
Heterochronic genes do not regulate seam cell asymmetric division.
kin-19/CKIα is closely related to kin-20/CKIε/δ, a C. elegans heterochronic gene that determines the correct timing of seam cell terminal differentiation.Citation33 In vertebrate and invertebrate cells, CKIε and CKIα can work additively or in opposition to regulate Wnt signaling pathways.Citation40,Citation55,Citation56 Thus, we investigated whether kin-20/CKIε, or other heterochronic genes, interacts with kin-19/CKI, or other Wnt pathway genes, to regulate seam cell asymmetric division. Since the effects of Wnt pathway gene RNAi on seam cell division are mainly observed during late larval development, we focused our analysis on the late larval heterochronic genes, which function during the L3 and L4 stages.
The late larval heterochronic genes regulate seam cell terminal differentiation, which occurs during the L4 to adult transition and consists of exit from the cell cycle and fusion of the seam cells into a syncytium, followed by secretion of collagenous ridges called alae. let-7 microRNA functions to allow the acquisition of adult seam cell fates after the L4 stage, and thus let-7 loss-of-function mutants have retarded terminal differentiation and show delayed and incomplete alae formation as adult animals.Citation57 lin-41, hbl-1, lin-42 and kin-20 function downstream of, or in parallel to, let-7 to specify larval seam cell identity and prevent terminal differentiation. Thus, these ‘precocious’ heterochronic genes need to be downregulated at the end of L4 to allow transition to adult fates, and loss of function of any of these let-7 target genes results in precocious terminal differentiation during the L3 or L4 stage ().Citation58–Citation61 One of the functions of the L4 ‘precocious’ genes appears to be inhibition of lin-29, whose function is required for terminal differentiation.Citation62 Thus, loss of function of the ‘precocious’ L4 genes allows LIN-29 function that results in terminal differentiation while loss of function of lin-29 prevents terminal differentiation, and results in incomplete seam cell fusion and alae secretion.
Single RNAi of the late larval heterochronic genes did not affect terminal seam cell number suggesting that these heterochronic genes do not regulate seam cell asymmetric division (). However, dRNAi of a heterochronic gene with a Wnt asymmetry pathway gene resulted in suppression of the terminal seam cell number phenotype produced on RNAi of pop-1, kin-19, apr-1 or pry-1 (). This result suggests that seam cell hyperplasia, resulting from disruption of asymmetric division and fate specification, depends on function of the ‘precocious’ L4 heterochronic genes. However, a model in which the function of the Wnt asymmetry pathway depends on that of the heterochronic pathway predicts that loss of function of heterochronic genes would affect seam cell asymmetrical division, which is not observed. An alternate and simpler explanation becomes apparent when changes in seam cell numbers are followed over larval development instead of assaying only terminal seam cell number (). RNAi mediated reduction of function of the precocious heterochronic genes led to a decrease in the maximum number of seam cell nuclei observed during the L4 stage, although initial seam cell number entering the L4 and the terminal seam cell were wild-type (). The decrease in maximum seam cell number indicates that fewer seam cells undergo division, probably due to seam cells that had precociously terminally differentiated and thus had exited the cell cycle. We have previously reported this phenotype (decrease in maximum number of seam cells) for kin-20 and lin-42 loss or reduction of function.Citation33 Therefore, it is likely that the suppression of seam cell hyperplasia, seen on dRNAi of precocious heterochronic genes with Wnt asymmetry pathway genes, occurs because some seam cells terminally differentiate during the early L4, resulting in net reduction of seam cells that are capable of undergoing division (–H). If the Wnt asymmetry pathway regulating seam cell specification was dependent on heterochronic gene function (as suggested by suppression of seam cell hyperplasia), one might also expect an enhancement of the seam cell hypoplasia phenotype. However, we did not observe an effect on seam cell hypoplasia in dRNAi experiments between the precocious heterochronic genes and Wnt asymmetry pathway genes, wrm-1, lit-1, sys-1 or mom-4 (). Finally, we also found that loss of function of the heterochronic genes let-7 or lin-29, either alone or in dRNAi with Wnt asymmetry pathway genes, did not affect seam cell asymmetric division as measured by terminal seam cell number (not shown). Therefore, our data indicate that heterochronic genes do not regulate seam cell asymmetric division, at least in the L4 stage.
Wnt asymmetry pathway genes do not regulate seam cell terminal differentiation.
Seam cell terminal differentiation involves permanent withdrawal from the cell cycle and loss of self-renewal capacity. pop-1(RNAi) and kin-19(RNAi) result in dramatic seam cell hyperplasia that involves extra cell divisions, and suggests that seam cell self-renewal capacity is extended, and terminal differentiation prevented, in these animals. We tested whether the Wnt asymmetry pathway genes play a role in regulating seam cell terminal differentiation by assaying the effect of Wnt pathway gene RNAi on seam cell fusion and adult alae expression ( and ). Single RNAi of apr-1, pry-1, sys-1, mom-4, bar-1 and hmp-2 resulted in wildtype seam cell terminal differentiation ( and ). However, RNAi of kin-19 or pop-1 and wrm-1 or lit-1 resulted in retarded development of adult alae (), although seam cell fusion occurred with normal timing (not shown). Since kin-19 or pop-1 have the opposite effect on seam cell asymmetric division (RNAi produces seam cell hyperplasia) compared to wrm-1 or lit-1 (RNAi produces seam cell hypoplasia), it is difficult to propose a mechanism by which both pairs of genes cause a retarded heterochronic phenotype. However, the observation that the timing of seam cell fusion, which precedes alae secretion, occurs with normal timing in animals on kin-19, pop-1, wrm-1 or lit-1 RNAi argues that adult alae formation is not strictly coupled to seam cell fusion, and that the lack of adult alae in these animals is not a true heterochronic phenotype. In pop-1(RNAi) and kin-19(RNAi) animals, the retarded alae formation phenotype is modest, and the areas along the seam that lack alae correlate well with areas where seam cell hyperplasia is most severe. However, in all cases that we observed, the seam cells had laterally fused into a syncytium by the late L4 and young adult stage, and we did not observe division of seam cell nuclei after syncytium formation. Therefore, the mis-specified extra seam cells in kin-19(RNAi) and pop-1(RNAi) animals fuse to form the seam syncytium at the normal developmental time, and the extra seam cell nuclei within the syncytia do not undergo nuclear division after syncytium formation. The reduction in adult alae in these animals may simply reflect a loss of competence of the seam cells to activate alae synthesis pathways, even though the seam cells have fused. In wrm-1(RNAi) and lit-1(RNAi) animals, the retarded alae formation phenotype is severe (), and the extent of lack of adult alae correlates with the loss of seam cells due to misspecification as hypodermal cells. We also observed that contiguous seam cells in these RNAi animals fused into mini-syncytia and secreted alae at the normal developmental time. Therefore the severe, seemingly retarded heterochronic phenotype of lack of adult alae is a consequence of misregulated seam cell asymmetric division and cell fate misspecification, and not due to delayed seam cell terminal differentiation.
We assayed for enhancement of the retarded terminal differentiation phenotypes of let-7(n2853) mutant animals and suppression of the precocious terminal differentiation phenotypes of lin-42(n1089), lin-41(ma104), hbl-1(mg285) and kin-20(RNAi) animals. animals. For all heterochronic mutants tested, RNAi of apr-1, pry-1, sys-1, mom-4, bar-1 or hmp-2 did not significantly alter the heterochronic phenotypes of the mutant or RNAi animals in terms of either seam cell fusion or adult alae expression (–F). RNAi of pop-1, wrm-1 or lit-1 enhanced the retarded alae phenotype of the let-7(n2853) mutant and suppressed the precocious alae phenotypes of the other heterochronic gene mutants and kin-20(RNAi) animals (–F). However, retardation or enhancement of adult alae formation was not mirrored by similar effects on seam cell fusion, which occurred with normal developmental timing in all cases (). Therefore, pop-1, wrm-1 and lit-1 do not affect the timing of seam cell terminal differentiation, as indicated by normal seam cell fusion. The effect of RNAi of these genes on heterochronic adult alae phenotypes can be explained by the inability of excess seam cells in pop-1(RNAi) animals to fully undergo terminal differentiation and form adult alae, and by the loss of alae producing seam cells in wrm-1(RNAi) and lit-1(RNAi) animals (). The results of double RNAi experiments of heterochronic genes with Wnt pathway genes, and RNAi of Wnt pathway genes in heterochronic gene mutants indicate that the Wnt asymmetry pathway genes, as well as bar-1 and hmp-2, do not regulate the timing of seam cell terminal differentiation.
kin-19 has dual function in Wnt asymmetric pathway and heterochronic pathway.
We propose a model in which the Wnt asymmetry genes and the L4 heterochronic genes function in separate and parallel pathways to control seam cell asymmetric division and terminal differentiation, respectively. RNAi phenotypes of seam cell development for all of the Wnt pathway genes and heterochronic genes tested are consistent with this ‘separate and parallel’ model, with the notable exception of kin-19. Among the Wnt asymmetry pathway genes, only RNAi of kin-19, in combination with loss of function of the L4 heterochronic genes, resulted in suppression of both precocious alae formation and precocious seam cell fusion ( and ). These results indicate that loss of kin-19 results in bona fide alteration to the timing of seam cell terminal differentiation. kin-19(RNAi) also enhanced the retarded heterochronic phenotype of let-7(n2853) mutant animals, and is consistent with kin-19 function either downstream or parallel to let-7 and the L4 precocious heterochronic genes (). The results of the dRNAi experiments indicate that kin-19 normally functions as a positive regulator of seam stem cell terminal differentiation (). However, kin-19(RNAi) alone does not produce a discernable heterochronic phenotype. Therefore, the primary function of kin-19, as part of the Wnt asymmetry pathway in the seam cells, appears to be the regulation of asymmetric division. But kin-19 has a secondary function, outside of the Wnt asymmetry pathway and most likely in parallel with the heterochronic pathway, in regulating seam cell terminal differentiation.
Discussion
We present the first example of a single CKI isoform, CKIα, acting to regulate both asymmetric division and terminal differentiation in the C. elegans epidermal stem cells. We report that kin-19/CKIα regulates seam stem cell fate based on asymmetric division, and functions in a non-canonical Wnt pathway that involves the β-catenins wrm-1 and sys-1. Furthermore, we show that kin-19 also interacts with the late larval heterochronic genes to regulate seam stem cell withdrawal from self-renewal and allow terminal differentiation. However, other known components of C. elegans canonical and non-canonical Wnt signaling pathways do not show a similar interaction with heterochronic genes and do not appear to play a role in seam cell terminal differentiation. Similarly, we found that the late larval heterochronic genes did not play a role in regulating seam cell asymmetric division. The seam cell Wnt/β-catenin asymmetry pathway and the heterochronic temporal identity pathway thus appear to function separately and in parallel to regulate different aspects of seam stem cell fate. These two pathways appear to have at least one point of molecular intersection at kin-19/CKIα, whose regulation and function may serve to integrate signaling through the spatial and temporal pathways that regulate seam stem cell development.
The role of kin-19/CKIα in the seam cell Wnt/β-catenin asymmetry pathway.
CKIα is a conserved component of the canonical Wnt/β-catenin signaling pathway in Drosophila and vertebrates, and most studies indicate that it functions as a negative regulator of β-catenin stability, and thus as a negative regulator of Tcf/Lef-mediated transcription of Wnt pathway target genes.Citation16,Citation63 Asymmetric division in the post-embryonic seam cells is controlled by a non-canonical Wnt/β-catenin pathway that utilizes β-catenin nuclear localization instead of β-catenin stability (). Therefore, a direct comparison between the canonical Wnt/β-catenin and non-canonical Wnt/β-catenin asymmetry pathways is inappropriate. However, our studies indicate that kin-19/CKIα normally functions as a negative regulator of nuclear localization, or nuclear activity, of both wrm-1/β-catenin and sys-1/β-catenin in the seam cells. Downregulation of wrm-1 or sys-1 function has the opposite effect on seam cell specification (seam cell hypoplasia) compared to downregulation of pop-1 or kin-19 function (seam cell hyperplasia) (, and ). Additionally, semi-epistasis analysis by double RNAi suggests that wrm-1 and sys-1 function downstream of kin-19 ( and ). Thus, kin-19/CKIα normally functions in the postembryonic seam cells to decrease the POP-1:SYS-1 ratio and thus activate the POP-1:SYS-1 transcriptional complex. There appears to be developmental and/or cell specific variations in kin-19/CKIα function. In contrast to its post-embryonic role, kin-19/CKIα functions as a positive regulator of the Wnt/β-catenin asymmetry pathway during asymmetric division of the EMS blastomere. Loss of function of kin-19 or of wrm-1 or pop-1 results in the same mom phenotype, in which excess mesoderm is specified at the expense of endoderm.Citation39 These differences in embryonic versus post-embryonic function may be due to differences in the upstream signaling cascades that converge on WRM-1 and SYS-1 β-catenins and POP-1. In the EMS blastomere, the asymmetry pathway is signaled by cell extrinsic MOM-2/Wnt, which is detected by the transmembrane MOM-5/Wnt receptor, and the signal is transduced through DSH-2 and MIG-5 Dishevelled molecules and GSK-3β kinase.Citation39,Citation64,Citation65 In contrast, we find that GSK-3β, Dsh molecules, Wnt receptors and Wnt molecules do not appear to have a function in the post-embryonic seam cell Wnt asymmetry pathway (). Indeed, the V seam cell Wnt asymmetry pathway may be regulated differently from the analogous pathway in T-seam cells, in which asymmetric division is signaled by cell extrinsic LIN-44/Wnt that is received by the transmembrane LIN-17/Wnt receptor.Citation39,Citation49 Because of functional redundancy among Wnt molecules,Citation66–Citation68 we cannot unequivocally state from the results of our single RNAi experiments that Wnt signaling does not activate the V seam cell Wnt asymmetry pathway. However, the modest effect or lack of effect on asymmetric division with RNAi of Wnt receptor genes suggests that the V seam cell Wnt asymmetry pathway may function cell autonomously, while asymmetric division of the T seam cell and embryonic EMS blastomere is regulated cell extrinsically by Wnt signaling.
What are the regulatory targets and mechanistic function of KIN-19 in the seam cell Wnt asymmetry pathway? Since kin-19(RNAi) results in specification of excess seam cells at the expense of Hyp7 cells, the normal function of KIN-19 must be as a positive regulator of Hyp7 cells, in which the nuclear POP-1:SYS-1 ratio is high, and/or as a negative regulator of seam cells, in which the nuclear POP-1:SYS-1 ratio is low. The simplest model of KIN-19 function is that KIN-19 positively regulates nuclear export of WRM-1, and does so possibly in complex with APR-1 and/or PRY-1. Thus, kin-19(RNAi) would result in increased levels of WRM-1/LIT-1, decreased levels of POP-1, decrease in the POP-1:SYS-1 ratio and specification of seam cell fate—as is experimentally observed (). APR-1 is though to enable WRM-1 nuclear export and localization to the cell cortex, where it forms a complex with PRY-1.Citation8,Citation46,Citation69 In our study, apr-1(RNAi) resulted in an excess seam cell phenotype that is similar to, although less severe than, kin-19(RNAi), suggesting that APR-1 and KIN-19 regulate the Wnt asymmetry pathway in the same manner ( and ). Furthermore, the genetic epistatic relationship between pry-1 and kin-19 is the same as that between pry-1 and apr-1, which are known to function in parallel ().Citation8,Citation46,Citation69 Double RNAi of apr-1 and kin-19 did not substantially enhance one or the other phenotype, also suggesting that APR-1 and KIN-19 function in parallel (). This model of KIN-19 function predicts that KIN-19 should co-localize with APR-1 and/or PRY-1, and that KIN-19 may shuttle between the nucleus and the cell cortex, as has been observed for APR-1.Citation69 While we have not yet rigorously tested these predictions, we observed that KIN-19::GFP is expressed in both nucleus and cytoplasm. Furthermore, cytoplasmic KIN-19::GFP in seam cells formed a punctate expression pattern, which could indicate the presence of macromolecular complexes, such as that of KIN-19 with APR-1, PRY-1 and other molecules. Additional support for our model of kin-19 function comes from the fact that the association of CKIα and Axin and APC to regulate β-catenin is a conserved mechanism in the vertebrate and Drosophila canonical Wnt pathways.Citation2,Citation15
The function of KIN-19 may be to regulate the amount of nuclear SYS-1, in addition to or as alternative to regulating nuclear POP-1 levels via WRM-1 localization. A well-documented function of Casein Kinase I is regulation of a target protein's stability via phosphorylation that leads to Ubiquitin-mediated proteolysis.Citation16 For example, in the canonical Wnt signaling pathway of vertebrates, CKIα phosphorylation is the key step that results in β-catenin degradation.Citation16 In the C. elegans non-canonical Wnt asymmetry pathway, SYS-1 β-catenin is primarily regulated by the protein degradation system.Citation70 KIN-19 may be localized to the anterior seam daughter cell, which becomes specified as Hyp7, and function to target SYS-1 β-catenin for degradation. Therefore, kin-19(RNAi) would result in higher nuclear SYS-1 levels, a lower POP-1:SYS-1 nuclear ratio and misspecification of the anterior daughter cell as seam cell. In both models of KIN-19 activity in the seam cell asymmetry pathway, KIN-19 primarily functions in the anterior daughter cell that is fated to become hypodermis (). We did not observe asymmetric localization of KIN-19:GFP during seam cell division, but anterior cell-specific localization of KIN-19 may not be necessary as its postulated interactors, APR-1 and PRY-1, are localized to the anterior cortex before cell division.Citation8,Citation44
The relationship of kin-19 to the heterochronic genes in regulating seam cell terminal differentiation.
Our studies indicate that kin-19 has dual and independent function in Wnt/β-catenin asymmetry and heterochronic pathways. While the retarded or precocious terminal differentiation phenotypes, caused by reduction of function of L4 heterochronic genes, are partially dependent on kin-19/CKIα, other components of the Wnt asymmetry pathway do not interact genetically with the L4 heterochronic genes ( and ). Therefore, it appears that kin-19 does not signal through POP-1 or any of the C. elegans β-catenins to exert its regulation of terminal differentiation. Our semi-epistasis RNAi experiments did not involve null alleles and thus it is not possible to accurately define the genetic relationship between kin-19 and the late larval heterochronic genes. While there are no reports of CKI involvement in microRNA expression, biogenesis or function, in yeast and a number of mammalian cell lines, CKI has been shown to co-localize and function with nuclear RNA processing factors, and also to regulate activity of eIF-4E mRNA cap-binding protein.Citation71,Citation72 Thus, KIN-19/CKIα may have a function in microRNA processing that has not yet been defined. However, since kin-19 loss of function alone does not have a discernable heterochronic phenotype, and kin-19 loss of function phenotype is unaffected by the L4 heterochronic genes (–), it is more likely that kin-19 does not function directly in the heterochronic pathway, and instead functions in parallel to the heterochronic genes to control the timing of terminal differentiation ().
How does KIN-19/CKIα regulate terminal differentiation and loss of self-renewal in seam stem cells?
Seam cell terminal differentiation is characterized by a sequence of cellular events including mitotic cell cycle withdrawal, cell fusion to form a syncytium and the expression of adult seam cell specific genetic programs, such as for alae secretion. CKIα in animals systems other than C. elegans is known to regulate cell cycle progression and cell adhesion, as well as regulate the function of transcription factors involved in differentiation.Citation32 For example, studies in yeast and mammalian cell culture show that CKIα prevents progression from G2 to mitosis by directly phosphorylating Cdc25 phosphatase, which a key regulator of this transition.Citation73,Citation74 Therefore, in these systems CKIα acts as a positive regulator of cell cycle withdrawal by preventing cell cycle progression. C. elegans has four cdc-25 genes, two of which have well-defined, but partially redundant, functions in regulating cell cycle progression in embryonic germline stem cells.Citation75–Citation77 Post-embryonic functions of the C. elegans cdc-25 genes have not been rigorously tested, and thus it is possible that these genes may function redundantly to regulate cell cycle progression in somatic cells, such as the seam cells. Another point at which KIN-19/CKIα could regulate seam cell withdrawal from mitosis is through the C. elegans homologs of the calcium sensor, calmodulin (CaM) and/or FADD (Fas-associated protein with Death Domain). In vertebrates, FADD and Calmodulin act together to regulate cell cycle progression, cell proliferation and apoptosis. Studies in mice and vertebrate cell lines show that CKIα directly phosphorylates FADD and CaM in vitro, and that loss of CKIα phosphorylation function leads to lack of FADD binding to CaM, and results in cell proliferation due to lack of checkpoint arrest at the G2 to M transition.Citation78,Citation79 KIN-19/CKIα could similarly interact with the products of the C. elegans CaM homologs (cmd-1, cal-1, cal-2, cal-3, cal-4) and/or with the C. elegans FADD homolog. Therefore, kin-19 loss of function could result in lack of CaM and FADD interaction, loss of normal G2 to M checkpoint control and thus prevent cell cycle withdrawal and inhibit terminal differentiation. In C. elegans, both CDC-25 and CaM are encoded by multiple genes whose products may function redundantly. This possible functional redundancy in control of mitotic cell cycle progression may be a reason for why kin-19/CKIα loss of function only has an effect on terminal differentiation in the sensitized backgrounds of heterochronic gene mutants. A final model for KIN-19 regulation of the seam cell cycle is based on the relationship between CKIα and the p53 tumor suppressor. p53 integrates various environmental signals and responds to cellular stress by inducing cell cycle arrest and apoptosis, thus minimizing genetic instability.Citation80 In undamaged mammalian cells, CKIα has been found to phosphorylate and stabilize p53.Citation81,Citation82 Therefore, KIN-19 may also normally function to stabilize CEP-1, the C. elegans p53 homolog.Citation80 Reduction in CEP-1/p53 due to kin-19 loss of function may result in reduction of cell cycle arrest and thus prevent terminal differentiation.
In addition to, or instead of, controlling cell cycle progression, KIN-19 may regulate key transcription factors that control terminal differentiation programs. In vertebrates, the FoxG1 (Forkhead family) transcriptional repressor functions to maintain neural stem cell identity during forebrain neurogenesis.Citation83,Citation84 Terminal differentiation of the neural stem cell is triggered by nuclear export of FoxG1, and CKI phosphorylation enables nuclear localization and stabilization of nuclear FoxG1.Citation85 The heterochronic gene product KIN-20/CKIε/δ also functions as a positive regulator of self-renewal and a negative regulator of terminal differentiation in seam cells,Citation33 and CKIε/δ and CKIα play antagonistic roles in the vertebrate canonical Wnt signaling pathway.Citation32 Therefore, KIN-20/CKIε/δ and KIN-19/CKIα may function antagonistically to regulate function of one or more of the 15 C. elegans Forkhead transcription factors or other transcription factors, which control self-renewal versus terminal differentiation in the seam cells.
In this study, seam cell terminal differentiation is assayed by seam cell fusion and adult alae expression, and KIN-19/CKIα may directly regulate the process of cell fusion. Vertebrate CKIα has been shown to destabilize adherens junctions and reduce cell-to-cell contacts by phosphorylating E-cadherin.Citation86 If, KIN-19/CKIα plays a similar role in the seam cells, then loss of kin-19 function would inhibit seam cell fusion and thus delay terminal differentiation.
In summary, our study indicates that C. elegans KIN-19/CKIα functions in a Wnt/β-catenin pathway to regulate seam stem cell asymmetric division, and also has a separate function, in parallel with the heterochronic/temporal identity pathway, in regulating seam stem cell terminal differentiation. The dual but independent roles of KIN-19 suggest that different aspects of seam stem cell development may be coordinately regulated through KIN-19. Distinguishing among the proposed models of KIN-19 action, and defining the exact mode of KIN-19 function in the Wnt/β-catenin asymmetry pathway, as well as in regulating terminal differentiation, will depend on characterizing KIN-19 subcellular localization and identifying direct phosphorylation targets in the seam cells. The conserved function of Wnt/β-catenin and heterochronic pathway molecules and CKI in regulating cellular differentiation in a variety of animal systems suggests that definition of KIN-19 function and regulation in C. elegans seam cell development will illuminate the fundamental process of stem cell development in higher vertebrates. In particular, the study of kin-19 regulation presents the opportunity to define how multiple aspects of stem cell development may be coordinated through a single molecule.
Materials and Methods
C. elegans strains.
Britol N2 was used as the wild type strain. Strains JR1000/wIs78 (ajm-1::gfp/MH27::GFP and scm-1::gfp), JR667/wIs51 (unc-119(e2498)::Tc1;wIs51), and PS3729/syIs78 (unc-119(ed4); syIs78[ajm-1::GFP + unc-119(+)]) were used to visualize seam cell nuclei and seam cell junctions. Additional strains used in this work are listed by chromosome as follows: LGI: lin-41(ma104), LGII: lin-42(n1089), LGIII: wrm-1(ne1982), lit-1(ne1991), LGX, let-7(n2853), hbl-1(mg285). All strains were propagated on NGM plates containing E. coli OP50 bacterial lawns. Synchronized starved L1 populations were obtained by hatching eggs, isolated from gravid adults, and starving larvae in M9 medium. All strains were cultured at 20°C, unless otherwise noted.
Microscopy.
Light and fluorescence microscopy were carried out on Zeiss AxioPlan 2 and Zeiss AxioimagerZ1 microscopes and images were captured and analyzed using Zeiss AxioVision software and Hamatzu cameras.
RNAi analysis.
RNAi constructs for the following genes were obtained from the Ahringher RNAi library: bar-1, hmp-2, sys-1, pop-1, lit-1, gsk-3, pry-1, apr-1, mig-5, egl-20. Additional RNAi constructs were created using the pL4440 vector, and were made for the following genes: kin-20, kin-19, wrm-1, mom-4, dsh-1, dsh-2, mig-5, lin-17, lin-18, mom-5, cwn-1, cwn-2, lin-44, mom-2, mig-14. The specific gene sequence of all RNAi constructs was sequenced to confirm the identity of the insert. RNAi was induced by feeding, starting with L1 stage starved and synchronized animals. RNAi constructs were expressed from E. coli HT115(DE3) bacteria, which were grown on NGM media containing 1 mM IPTG to induce expression from the convergent T7 polymerase promoters on the pL4440 vector. All strains feeding on RNAi bacteria were cultured at 20°C unless otherwise noted. Simultaneous RNAi was carried out by growing the two different RNAi bacterial strains to the same optical density and mixing equal volumes of the cultures before plating onto NGM plates. Mock RNAi consisted of feeding animals on bacteria carrying the empty pL4440 vector (EV). The extent of target gene expression knockdown by RNAi was measured by quantitative real time PCR using SYBR green. Target gene expression level was standardized to ama-1 transcript levels from the same population, and the ratio of target gene:ama-1 from RNAi and control populations was compared. RNAi efficiencies ranged from 48% to 83% depending on the RNAi construct.
Plasmids and molecular analysis.
kin-19::gfp transcriptional fusion construct. 4.5 kb of the kin-19 promoter sequence was PCR amplified from C. elegans genomic DNA, 0.9 kb of kin-19 coding sequence was PCR amplified from N2 cDNA, and gfp and unc-54 3′UTR sequences were PCR amplified from the plasmid pPD95.Citation75. The kin-19 translational reporter was constructed by fusing the three PCR fragments by overlap-extension PCR. Primers used are available upon request. The resulting kin-19p::kin-19::gfp::unc-54 3′UTR fragment was injected into N2 animals at 50 ng/µl along with rol-6 at 75 ng/µl as a coinjection marker. Five transgenic lines were identified by the rolling phenotype caused by rol-6. Rolling animals were re-examined to confirm the presence of GFP, and spatial and temporal expression patterns of the GFP were examined for all developmental stages in all five transgenic lines.
Quantitative real time PCR (qRT-PCR) of kin-19. Synchronized populations of N2/wildtype were grown at 25°C on OP50, and harvested every 2 hours post-plating, starting at 4 hrs and extending to 40 hrs. Total RNA was extracted from each population using Trizol, cDNA was synthesized from the RNA and used for qRT-PCR. kin-19 and ama-1 specific primers were used for each qRT-PCR run with SYBR Green on Biorad CFX96 RT-PCR detection system.
Figures and Tables
Figure 1 kin-19/CKIα expression during C. elegans larval development. kin-19::GFP translational reporter expression in the (A) epidermal seam cells (SC), (B) vulval cells (V) and neuronal cells (N) of the ventral nerve cord in L4 stage wildtype animals. Images taken at 430×. (C) Expression levels of kin-19 transcript, relative to ama-1 transcript, during larval stages in wildtype C. elegans grown at 25°C.
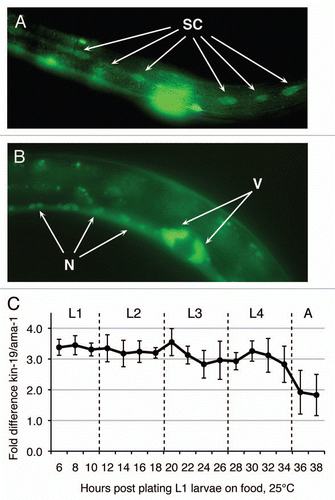
Figure 2 Anomalous terminal seam cell numbers result from RNAi knockdown of kin-9, pop-1 or wrm-1. (A–D) Young adult or L4 wIs78 worms, in which GFP expression marks the seam cell nuclei (SCM::GFP) and adherens junctions (ajm-1::gfp/MH27::GFP). Images are composites of 3 to 4 photographs of the same animal in each case, taken at a magnification of 200×. (A) Young adult wIs78 animal on mock RNAi showing 14 seam cell nuclei within the continuous syncytium of the fused seam cells. (B) Young adult wIs78 animal on kin-19(RNAi) showing 27 seam cell nuclei. (C) Young adult wIs78 animal on pop-1(RNAi) showing 48 seam cell nuclei. (D) Mid L4 stage wIs78 animal on wrm-1(RNAi) showing 9 seam cell nuclei (SC) within the discontinuous seam cell syncytium. Also visible in the plane of the seam cells are the nuclei of cells (Hyp) that have not fused with the seam cell syncytium.
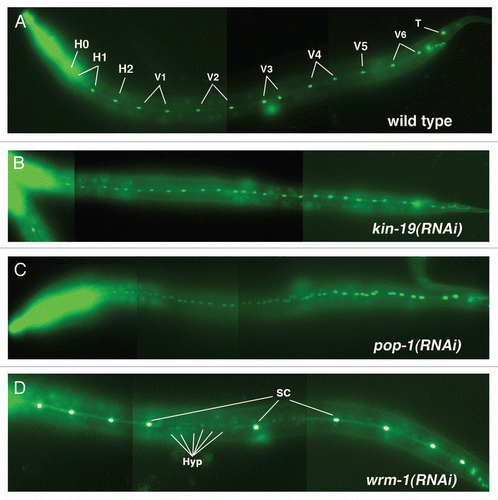
Figure 3 (A–H) Anomalous seam cell numbers result from RNAi knockdown of kin-19 and Wnt signaling pathway genes. For each RNAi condition, n = 45–50 animals were observed for each time point, and data points are averages of three trials. Error bars indicate standard deviation. (A) wIs78 animals on mock RNAi, and RNAi of bar-1, hmp-2 or sys-1 showed wildtype seam cell numbers and pattern of division. (B) RNAi knockdown of kin-19, pop-1 or apr-1 resulted in seam cell hyperplasia starting in the early L4 stage and persisting into the adult stage. (C) RNAi knockdown of wrm-1, lit-1 or mom-4 resulted in seam cell hypoplasia, starting during the late L2 stage and persisting into the adult stage. (D) RNAi knockdown of the heterochronic genes kin-20, lin-42 or lin-41 resulted in the normal terminal seam cell number, but fewer than normal seam cells underwent division in the L4 stage. hbl-1(RNAi) produced a similar seam cell pattern of division and terminal seam cell number (not shown).
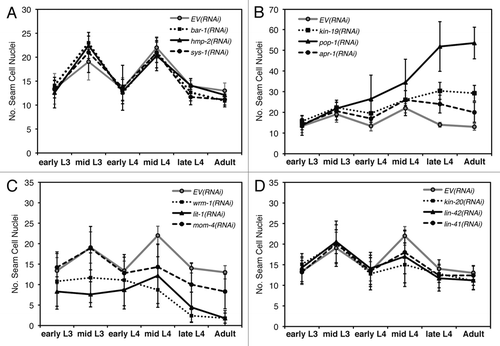
Figure 4 Post-embryonic lineages of epidermal seam cells V1–4 and V6 on RNAi knockdown of wnt signaling pathway genes and/or heterochronic genes. H: hypodermal cell, SC: seam cell, SC boxed in black indicates terminally differentiated seam cells that have fused into a syncytium. L1–L4: larval stages 1 to 4, A: adult. Numbers in parentheses at the bottom of each lineage indicate the number of terminal seams cells within the lineage shown (first number), and the range of terminal seam cells that results if all V-seam cells undergo the lineage shown (second number). (A) Normal pattern of seam cell (V1–4, 6) division and differentiation during larval development. (2/14) Each of the V1–4, V6 lineages produce 2 terminal seam cells, and the total number of visible terminal seam cells is 14. (B–D) Models of misspecification of V seam cell lineages in worms on (B) kin-19(RNAi), (C) pop-1(RNAi) and (D) wrm-1(RNAi) or lit-1(RNAi). (E–H) Models of misspecification of V seam cell lineages in worms on (E) RNAi of precocious heterochronic genes (PHG-kin-20, lin-42, lin-41 or hbl-1), and (F–H) simultaneous RNAi of a PHG and a wnt signaling pathway gene—(F) kin-19, (G) pop-1, (H) wrm-1 or lit-1.
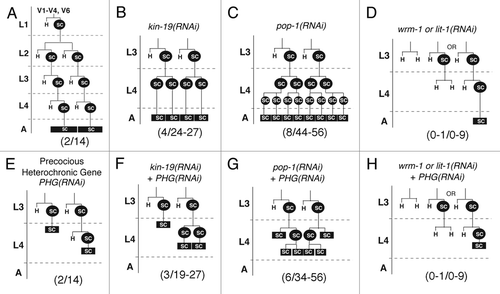
Figure 5 Effect of genetic interaction between kin-19 and Wnt signaling pathway genes on terminal seam cell number, a measure of asymmetric division. (A and B) The number of terminal seam cell nuclei was observed in wIs78 animals fed on simultaneous RNAi of kin-19 and the indicated Wnt signaling pathway gene, except for the following in (A): wrm-1(ne1982ts);wIs78 or lit-1(ne1991ts);wIs78 mutant animals were grown at and subjected to RNAi of the indicated Wnt signaling pathway gene at 25°C. Results shown are averages of three trials carried out with n=50–65 animals for each RNAi condition. Error bars indicate standard deviation. (**) P< 0.05 and (*) P<0.1, indicates statistically significant suppression of kin-19/EV(RNAi) phenotype, determined using students t-test. The dashed line indicates the average terminal seam cell number on kin-19(RNAi). The dotted line indicates the wildtype terminal seam cell number.
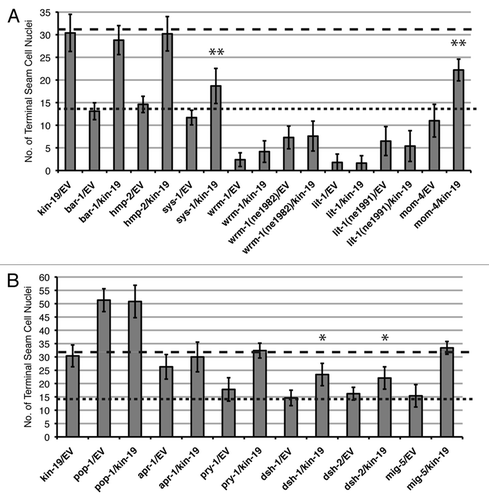
Figure 6 Effect of genetic interaction between kin-19 and heterochronic pathway genes on terminal seam cell number, a measure of asymmetric division. The number of terminal seam cell nuclei was observed in wIs78 animals fed on simultaneous RNAi of kin-19 and the indicated heterochronic pathway gene. Results shown are averages of three trials carried out with n = 50–65 animals for each RNAi condition. Error bars indicate standard deviation. **p < 0.05 and *p < 0.1, indicates statistically significant suppression of kin-19/EV(RNAi) phenotype, determined using students t-test.
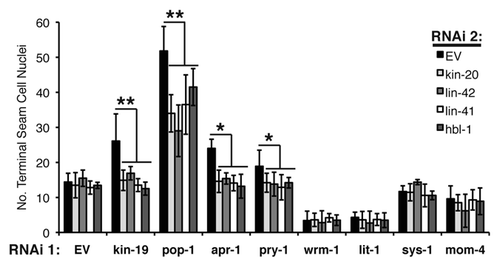
Figure 7 Effect of genetic interaction between kin-19 and heterochronic pathway genes on alae formation, a measure of terminal differentiation. The extent of alae expression was observed in young adult wIs78 (A) or let-7(n2853) (B) worms on RNAi of the indicated Wnt signaling pathways genes. The extent of alae expression was observed in early L4 stage lin-42(n1089) (C), lin-41(ma104) (D) hbl-1(mg285) (E) and wIs78 (F) animals on RNAi of the indicated Wnt signaling pathways genes or double RNAi with kin-20 (F). Results shown are the average of three trials of each experiment carried out with n = 30–40 animals for each RNAi condition. Error bars indicate standard deviation. **p < 0.05 and *p < 0.1, indicates statistically significant suppression compared to EV(RNAi), determined using students t-test.
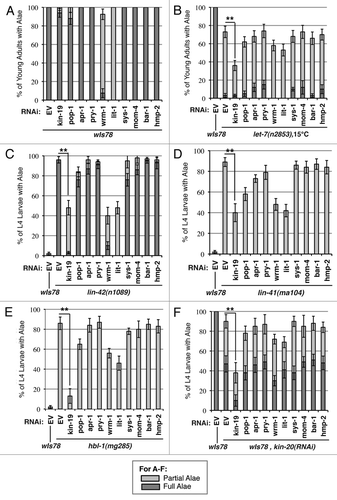
Figure 8 Effect of genetic interaction between kin-19 and heterochronic pathway genes on seam cell fusion, a measure of terminal differentiation. (A) Seam cell fusion in early and mid-L4 animals of the following strains: wIs78, lin-42(n1089), lin-42(ma104), hbl-1(mg285) on RNAi of the indicated Wnt asymmetry pathway genes. (B) Seam cell fusion in early and mid-L4 wIs78 animals on simultaneous RNAi of kin-20 and the indicated Wnt asymmetry pathway gene. For (B), the dashed line indicates the average number of L4 stage wIs78 animals on kin-20(RNAi) showing precocious seam cell fusion. Error bars indicate standard deviation. **p < 0.05 and *p < 0.1, indicates statistically significant suppression compared to EV(RNAi), determined using students t-test.
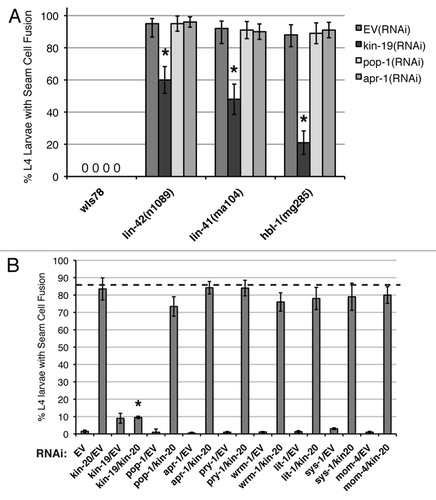
Figure 9 Models of kin-19 function in C. elegans seam stem cells. (A) A simplified model of differential fate specification in daughter cells mediated by the Wnt/β-catenin asymmetry pathway, and the possible roles of KIN-19 in this pathway. FZ: Frizzled, Wnt receptor, PM: plasma membrane, NM: nuclear membrane, P: phosphorylation. Pathway components that do not appear to function in the V seam cell asymmetry pathway (Wnt, FZ) are shown in grey. Indirect and speculative interactions are indicated by dashed, grey lines. (B) The relationship between kin-19 and the late larval heterochronic genes in regulating seam cell terminal differentiation. kin-19 likely functions downstream of and in parallel to the heterochronic genes to act as a positive regulator of terminal differentiation.
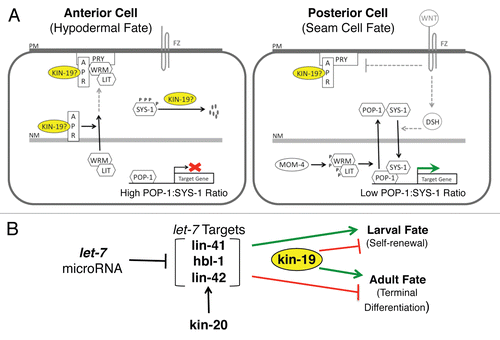
Table 1 Terminal seam cell number resulting from genetic interaction between Wnt singaling pathway genes
Acknowledgements
We thank the CGC for nematode strains, and the C. elegans and C. briggsae genome sequencing consortia for sequence data. This work was supported by Yale Biological Sciences Postdoctoral Fellowship, Anna B. Fuller Fellowship, and Virginia Tech start-up grant to D.B. and NIH R01 grant (GM64701) to F.J.S.
References
- Staal FJT, Luis TC. Wnt signaling in hematopoiesis: Crucial factors for self-renewal, proliferation and cell fate decisions. J Cell Biochem 2010; 109:844 - 849
- Nusse R. Wnt signaling and stem cell control. Cell Res 2008; 18:523 - 527
- Joshi PM, Riddle MR, Djabrayan NJV, Rothman JH. Caenorhabditis elegans as a model for stem cell biology. Dev Dyn 2010; 239:1539 - 1554
- Nimmo RA, Slack FJ. An elegant miRror: microRNAs in stem cells, developmental timing and cancer. Chromosoma 2009; 14
- Sulston JE, Horvitz HR. Post-embryonic cell lineages of the nematode Caenorhabditis elegans. Dev Biol 1977; 56:110 - 156
- Moss E. Heterochronic Genes and the Nature of Developmental Time. Curr Biol 2007; 17:425 - 434
- Eisenmann DM. Wnt signaling. WormBook 2005; 1 - 17
- Mizumoto K, Sawa H. Two betas or not two betas: regulation of asymmetric division by beta-catenin. Trends in Cell Biology 2007; 17:465 - 473
- Huang X, Tian E, Xu Y, Zhang H. The C. elegans engrailed homolog ceh-16 regulates the self-renewal expansion division of stem cell-like seam cells. Developmental Biology 2009; 333:337 - 347
- Nimmo R, Antebi A, Woollard A. mab-2 encodes RNT-1, a C. elegans Runx homologue essential for controlling cell proliferation in a stem cell-like developmental lineage. Development 2005; 132:5043 - 5054
- Xia D, Zhang Y, Huang X, Sun Y, Zhang H. The C. elegans CBFβ homolog, BRO-1, regulates the proliferation, differentiation and specification of the stem cell-like seam cell lineages. Dev Biol 2007; 309:259 - 272
- Kagoshima H, Shigesada K, Kohara Y. RUNX regulates stem cell proliferation and differentiation: Insights from studies of C. elegans. J Cell Biochem 2007; 100:1119 - 1130
- Resnick TD, Mcculloch KA, Rougvie AE. miRNAs give worms the time of their lives: Small RNAs and temporal control in Caenorhabditis elegans. Dev Dyn 2010; 239:1477 - 1489
- Van Hoffelen S, Herman MA. Analysis of Wnt signaling during Caenorhabditis elegans postembryonic development. Methods Mol Biol 2008; 469:87 - 102
- Clevers H. Wnt/beta-catenin signaling in development and disease. Cell 2006; 127:469 - 480
- Knippschild U, Gocht A, Wolff S, Huber N, Lohler J, Stoter M. The casein kinase 1 family: Participation in multiple cellular processes in eukaryotes. Cell Signal 2005; 17:675 - 689
- Lin R, Hill RJ, Priess JR. POP-1 and anterior-posterior fate decisions in C. elegans embryos. Cell 1998; 92:229 - 239
- Korswagen HC, Herman MA, Clevers HC. Distinct beta-catenins mediate adhesion and signalling functions in C. elegans. Nature 2000; 406:527 - 532
- Natarajan L, Witwer NE, Eisenmann DM. The divergent Caenorhabditis elegans beta-catenin proteins BAR-1, WRM-1 and HMP-2 make distinct protein interactions but retain functional redundancy in vivo. Genetics 2001; 159:159 - 172
- Kidd AR 3rd, Miskowski JA, Siegfried KR, Sawa H, Kimble J. A beta-catenin identified by functional rather than sequence criteria and its role in Wnt/MAPK signaling. Cell 2005; 121:761 - 772
- Rocheleau CE, Yasuda J, Shin TH, Lin R, Sawa H, Okano H, et al. WRM-1 activates the LIT-1 protein kinase to transduce anterior/posterior polarity signals in C. elegans. Cell 1999; 97:717 - 726
- Hunter CP, Harris JM, Maloof JN, Kenyon C. Hox gene expression in a single Caenorhabditis elegans cell is regulated by a caudal homolog and intercellular signals that inhibit wnt signaling. Development 1999; 126:805 - 814
- Phillips BT, Kidd AR, King R, Hardin J, Kimble J. Reciprocal asymmetry of SYS-1/beta-catenin and POP-1/TCF controls asymmetric divisions in Caenorhabditis elegans. Proc Natl Acad Sci USA 2007; 104:3231 - 3236
- Abbott A, Alvarezsaavedra E, Miska E, Lau N, Bartel D, Horvitz H, et al. The let-7 microRNA family members mir-48, mir-84 and mir-241 function together to regulate developmental timing in Caenorhabditis elegans. Dev Cell 2005; 9:403 - 414
- Liao J, Wu Z, Wang Y, Cheng L, Cui C, Gao Y, et al. Enhanced efficiency of generating induced pluripotent stem (iPS) cells from human somatic cells by a combination of six transcription factors. Cell Res 2008; 18:600 - 603
- Piskounova E, Viswanathan SR, Janas M, LaPierre RJ, Daley GQ, Sliz P, et al. Determinants of microRNA processing inhibition by the developmentally regulated RNA-binding protein Lin28. J Biol Chem 2008; 283:21310 - 21314
- Rybak A, Fuchs H, Smirnova L, Brandt C, Pohl E, Nitsch R, et al. A feedback loop comprising lin-28 and let-7 controls pre-let-7 maturation during neural stem-cell commitment. Nat Cell Biol 2008; 19
- Viswanathan SR, Daley GQ, Gregory RI. Selective blockade of microRNA processing by Lin28. Science 2008; 320:97 - 100
- Rybak A, Fuchs H, Hadian K, Smirnova L, Wulczyn EA, Michel G, et al. The let-7 target gene mouse lin-41 is a stem cell specific E3 ubiquitin ligase for the miRNA pathway protein Ago2. Nat Cell Biol 2009; 11:1411 - 1420
- Elliott J, Jolicoeur C, Ramamurthy V, Cayouette M. Ikaros confers early temporal competence to mouse retinal progenitor cells. Neuron 2008; 60:26 - 39
- Urban J, Mettler U. Connecting temporal identity to mitosis: the regulation of Hunchback in Drosophila neuroblast lineages. Cell Cycle 2006; 5:950 - 952
- Knippschild U, Wolff S, Giamas G, Brockschmidt C, Wittau M, Würl PU, et al. The role of the casein kinase 1 (CK1) family in different signaling pathways linked to cancer development. Onkologie 2005; 28:508 - 514
- Banerjee D, Kwok A, Lin SY, Slack FJ. Developmental timing in C. elegans is regulated by kin-20 and tim-1, homologs of core circadian clock genes. Dev Cell 2005; 8:287 - 295
- Virshup DM, Eide EJ, Forger DB, Gallego M, Harnish EV. Reversible protein phosphorylation regulates circadian rhythms. Cold Spring Harb Symp Quant Biol 2007; 72:413 - 420
- Borgs L, Beukelaers P, Vandenbosch R, Nguyen L, Moonen G, Maquet P, et al. Period 2 regulates neural stem/progenitor cell proliferation in the adult hippocampus. BMC Neurosci 2009; 10:30
- Mendez-Ferrer S, Lucas D, Battista M, Frenette PS. Haematopoietic stem cell release is regulated by circadian oscillations. Nature 2008; 452:442 - 447
- Moriya T, Hiraishi K, Horie N, Mitome M, Shinohara K. Correlative association between circadian expression of mousePer2 gene and the proliferation of the neural stem cells. Neuroscience 2007; 146:494 - 498
- Green CL, Bennett GS. Identification of four alternatively spliced isoforms of chicken casein kinase Ialpha that are all expressed in diverse cell types. Gene 1998; 216:189 - 195
- Korswagen HC. Canonical and non-canonical Wnt signaling pathways in Caenorhabditis elegans: variations on a common signaling theme. Bioessays 2002; 24:801 - 810
- Peters JM, McKay RM, McKay JP, Graff JM. Casein kinase I transduces Wnt signals. Nature 1999; 401:345 - 350
- Price MA. CKI, there's more than one: casein kinase I family members in Wnt and Hedgehog signaling. Genes Dev 2006; 20:399 - 410
- Herman M. C. elegans POP-1/TCF functions in a canonical Wnt pathway that controls cell migration and in a noncanonical Wnt pathway that controls cell polarity. Development 2001; 128:581 - 590
- Shin TH, Yasuda J, Rocheleau CE, Lin R, Soto M, Bei Y, et al. MOM-4, a MAP kinase kinase kinase-related protein, activates WRM-1/LIT-1 kinase to transduce anterior/posterior polarity signals in C. elegans. Molecular Cell 1999; 4:275 - 280
- Nakamura K, Kim S, Ishidate T, Bei Y, Pang K, Shirayama M, et al. Wnt signaling drives WRM-1/beta-catenin asymmetries in early C. elegans embryos. Genes Dev 2005; 19:1749 - 1754
- Takeshita H, Sawa H. Asymmetric cortical and nuclear localizations of WRM-1/beta-catenin during asymmetric cell division in C. elegans. Genes Dev 2005; 19:1743 - 1748
- Mizumoto K, Sawa H. Cortical beta-catenin and APC regulate asymmetric nuclear beta-catenin localization during asymmetric cell division in C. elegans. Dev Cell 2007; 12:287 - 299
- Hingwing K, Lee S, Nykilchuk L, Walston T, Hardin J, Hawkins N. CWN-1 functions with DSH-2 to regulate C. elegans asymmetric neuroblast division in a β-catenin independent Wnt pathway. Dev Biol 2009; 328:245 - 256
- King RS, Maiden SL, Hawkins NC, Kidd AR 3rd, Kimble J, Hardin J, et al. The N- or C-terminal domains of DSH-2 can activate the C. elegans Wnt/beta-catenin asymmetry pathway. Dev Biol 2009; 328:234 - 244
- Herman MA. Control of cell polarity by noncanonical Wnt signaling in C. elegans. Semin Cell Dev Biol 2002; 13:233 - 241
- Thorpe CJ, Schlesinger A, Carter JC, Bowerman B. Wnt signaling polarizes an early C, elegans blastomere to distinguish endoderm from mesoderm. Cell 1997; 90:695 - 705
- Yang PT, Lorenowicz MJ, Silhankova M, Coudreuse DY, Betist MC, Korswagen HC. Wnt signaling requires retromer-dependent recycling of MIG-14/Wntless in Wnt-producing cells. Dev Cell 2008; 14:140 - 147
- Zeng X, Tamai K, Doble B, Li S, Huang H, Habas R, et al. A dual-kinase mechanism for Wnt coreceptor phosphorylation and activation. Nature 2005; 438:873 - 877
- Smelkinson MG, Zhou Q, Kalderon D. Regulation of Ci-SCFSlimb binding, Ci proteolysis and hedgehog pathway activity by Ci phosphorylation. Dev Cell 2007; 13:481 - 495
- Harms E, Young MW, Saez L. CK1 and GSK3 in the Drosophila and mammalian circadian clock. Novartis Found Symp 2003; 253:267 - 277
- Sakanaka C. Phosphorylation and regulation of betacatenin by casein kinase I epsilon. J Biochem (Tokyo) 2002; 132:697 - 703
- Knippschild U, Wolff S, Giamas G, Brockschmidt C, Wittau M, Wurl PU, et al. The role of the casein kinase 1 (CK1) family in different signaling pathways linked to cancer development. Onkologie 2005; 28:508 - 514
- Reinhart BJ, Slack FJ, Basson M, Pasquinelli AE, Bettinger JC, Rougvie AE, et al. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 2000; 403:901 - 906
- Lin SY, Johnson SM, Abraham M, Vella MC, Pasquinelli A, Gamberi C, et al. The C. elegans hunchback homolog, hbl-1, controls temporal patterning and is a probable microRNA target. Dev Cell 2003; 4:639 - 650
- Abrahante JE, Daul AL, Li M, Volk ML, Tennessen JM, Miller EA, et al. The Caenorhabditis elegans hunchback-like gene lin-57/hbl-1 controls developmental time and is regulated by microRNAs. Dev Cell 2003; 4:625 - 637
- Slack FJ, Basson M, Liu Z, Ambros V, Horvitz HR, Ruvkun G. The lin-41 RBCC gene acts in the C. elegans heterochronic pathway between the let-7 regulatory RNA and the LIN-29 transcription factor. Mol Cell 2000; 5:659 - 669
- Jeon M. Similarity of the C. elegans developmental timing protein LIN-42 to circadian rhythm proteins. Science 1999; 286:1141 - 1146
- Rougvie AE, Ambros V. The heterochronic gene lin-29 encodes a zinc finger protein that controls a terminal differentiation event in Caenorhabditis elegans. Development 1995; 121:2491 - 2500
- Price M. CKI, there's more than one: Casein kinase I family members in Wnt and Hedgehog signaling. Genes Dev 2006; 20:399 - 410
- Shetty P, Lo MC, Robertson SM, Lin R. C. elegans TCF protein, POP-1, converts from repressor to activator as a result of Wnt-induced lowering of nuclear levels. Dev Biol 2005; 285:584 - 592
- Calvo D, Victor M, Gay F, Sui G, Luke MP, Dufourcq P, et al. A POP-1 repressor complex restricts inappropriate cell type-specific gene transcription during Caenorhabditis elegans embryogenesis. EMBO J 2001; 20:7197 - 7208
- Zinovyeva AY, Yamamoto Y, Sawa H, Forrester WC. Complex network of Wnt signaling regulates neuronal migrations during Caenorhabditis elegans development. Genetics 2008; 179:1357 - 1371
- Green JL, Inoue T, Sternberg PW. Opposing Wnt pathways orient cell polarity during organogenesis. Cell 2008; 134:646 - 656
- Gleason JE, Szyleyko EA, Eisenmann DM. Multiple redundant Wnt signaling components function in two processes during C. elegans vulval development. Dev Biol 2006; 298:442 - 457
- Rosin-Arbesfeld R, Cliffe A, Brabletz T, Bienz M. Nuclear export of the APC tumor suppressor controls beta-catenin function in transcription. EMBO J 2003; 22:1101 - 1113
- Huang G, Chen S, Li S, Cha J, Long C, Li L, et al. Protein kinase A and casein kinases mediate sequential phosphorylation events in the circadian negative feedback loop. Genes Dev 2007; 21:3283 - 3295
- Gross SD, Loijens JC, Anderson RA. The casein kinase Ialpha isoform is both physically positioned and functionally competent to regulate multiple events of mRNA metabolism. J Cell Sci 1999; 112:2647 - 2656
- Haas DW, Hagedorn CH. Casein kinase I phosphorylates the 25 kDa mRNA cap-binding protein. Arch Biochem Biophys 1991; 284:84 - 89
- Honaker Y, Piwnica-Worms H. Casein kinase 1 functions as both penultimate and ultimate kinase in regulating Cdc25A destruction. Oncogene 2031:1 - 11
- Pal G, Paraz MT, Kellogg DR. Regulation of Mih1/Cdc25 by protein phosphatase 2A and casein kinase 1. J Cell Biol 2008; 180:931 - 945
- Kim J, Kawasaki I, Shim YH. cdc-25.2, a C. elegans ortholog of cdc25, is required to promote oocyte maturation. J Cell Sci 2010; 123:993 - 1000
- Kim J, Lee AR, Kawasaki I, Strome S, Shim YH. A mutation of cdc-25.1 causes defects in germ cells but not in somatic tissues in C. elegans. Mol Cells 2009; 28:43 - 48
- Kipreos ET. C. elegans cell cycles: Invariance and stem cell divisions. Nat Rev Mol Cell Biol 2005; 6:766 - 776
- Papoff G, Trivieri N, Crielesi R, Ruberti F, Marsilio S, Ruberti G. FADD-calmodulin interaction: a novel player in cell cycle regulation. Biochim Biophys Acta 2010; 1803:898 - 911
- Alappat EC, Feig C, Boyerinas B, Volkland J, Samuels M, Murmann AE, et al. Phosphorylation of FADD at serine 194 by CKIalpha regulates its nonapoptotic activities. Mol Cell 2005; 19:321 - 332
- Derry WB, Putzke AP, Rothman JH. Caenorhabditis elegans p53: Role in apoptosis, meiosis and stress resistance. Science 2001; 294:591 - 595
- Huart AS, MacLaine NJ, Meek DW, Hupp TR. CK1alpha plays a central role in mediating MDM2 control of p53 and E2F-1 protein stability. J Biol Chem 2009; 284:32384 - 32394
- Maclaine NJ, Hupp TR. The regulation of p53 by phosphorylation: a model for how distinct signals integrate into the p53 pathway. Aging (Albany NY) 2009; 1:490 - 502
- Arden KC. FoxO: Linking new signaling pathways. Mol Cell 2004; 14:416 - 418
- Katoh M. Human FOX gene family (Review). Int J Oncol 2004; 25:1495 - 1500
- Regad T, Roth M, Bredenkamp N, Illing N, Papalopulu N. The neural progenitor-specifying activity of FoxG1 is antagonistically regulated by CKI and FGF. Nat Cell Biol 2007; 9:531 - 540
- Dupre-Crochet S, Figueroa A, Hogan C, Ferber EC, Bialucha CU, Adams J, et al. Casein kinase 1 is a novel negative regulator of E-cadherin-based cell-cell contacts. Mol Cell Biol 2007; 27:3804 - 3816