Abstract
Voltage-gated Na+ channels (VGSCs) are macromolecular protein complexes containing a pore-forming α subunit and smaller non-pore-forming β subunits. VGSCs are expressed in metastatic cells from a number of cancers. In these cells, Na+ current carried by α subunits enhances migration, invasion and metastasis in vivo. In contrast, the β subunits mediate cellular adhesion and process extension. The prevailing hypothesis is that VGSCs are upregulated in cancer, in general favoring an invasive/metastatic phenotype, although the mechanisms are still not fully clear. Expression of the Nav1.5 α subunit associates with poor prognosis in clinical breast cancer specimens, suggesting that VGSCs may have utility as prognostic markers for cancer progression. Furthermore, repurposing existing VGSC-blocking therapeutic drugs may provide a new strategy to improve outcomes in patients suffering from metastatic disease, which is the major cause of cancer-related deaths, and for which there is currently no cure.
Introduction
Voltage-gated Na+ channels (VGSCs) are comprised of a pore-forming α subunit typically in association with one or more smaller β subunits ().Citation1 The β subunits regulate channel expression and gating, and are immunoglobulin (Ig) superfamily cell adhesion molecules (CAMs).Citation2 VGSCs are classically responsible for action potential initiation and conduction in excitable cells.Citation3 Both the α and β subunits have been shown to interact with a range of other signaling molecules (), enabling fine-tuning of channel activity on the one hand, and allowing VGSCs to participate in non-conducting signaling on the other.Citation4 VGSCs are also expressed in a range of cell types that are considered “non-excitable,” including glia, fibroblasts, immune cells and metastatic cancer cells.Citation5 The past 15 y have seen a rapid expansion in published studies documenting the expression VGSCs across a broadening number of cancers, their role in regulating cellular migration and invasion and, importantly, their potential utility as diagnostic and therapeutic targets. In particular, several recent studies have started to define a mechanistic role for VGSCs in regulating migration and invasion. The purpose of this review is to assimilate the current body of evidence ascribing a malignant role for VGSCs during metastasis (the spread of tumor cells from primary to distant sites), and consider the clinical implications.
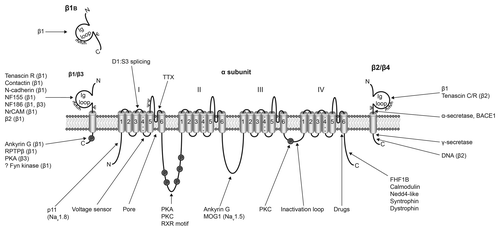
Figure 1. Multifunctional interactions of VGSCs. Basic topology of the pore-forming α subunit is shown, consisting of four homologous domains each containing six transmembrane segments. Segment four contains the voltage sensor.Citation6 The smaller β subunits contain an extracellular immunoglobulin (Ig) loop, transmembrane domain and an intracellular C-terminal domain.Citation88 β1 and β3 are non-covalently linked to the α subunit, whereas, β2 and β4 are covalently linked through disulfide bonds.Citation2 The alternative splice variant, β1B, lacks a transmembrane domain.Citation89 α subunits interact with a number of other signaling molecules, including p11,Citation90 protein kinase A (PKA),Citation91 protein kinase C (PKC),Citation92 ankyrin G,Citation93 MOG1,Citation94 fibroblast growth factor-homologous factor 1B (FHF1B),Citation95 calmodulin,Citation96 NEDD4,Citation97,Citation98 syntrophinCitation99 and dystrophin.Citation100 Several of the β subunits interact with other cell adhesion molecules and regulatory proteins, including tenascin C,Citation101 tenascin R,Citation101,Citation102 contactin,Citation103 N-cadherin,Citation104 neurofascin (NF)155,Citation105 NF186,Citation105 NrCAM,Citation105 ankyrin G,Citation104 receptor protein tyrosine phosphatase β (RPTPβ),Citation106 PKACitation107 and fyn kinase.Citation68 The β subunits are substrates for proteolytic cleavage by α-secretase, BACE1 and γ-secretase.Citation108,Citation109 The intracellular domain of β2 is proposed to regulate gene expression in the nucleus.Citation71 ψ, glycosylation sites. Figure was produced using Science Slides 2006 software.
VGSC Expression in Cancer
The VGSC α subunit family contains nine members, Nav1.1–Nav1.9, encoded by genes SCN1A–SCN11A ().Citation6 There are four β subunits, β1-β4, encoded by genes SCN1B–SCN4B.Citation2 VGSC expression has been predominantly reported to date in carcinomas (cancers of epithelial origin). The α subunits have been identified in cells from the following carcinomas: breast cancer,Citation7,Citation8 cervical cancer,Citation9,Citation10 colon cancer,Citation11 melanoma,Citation12,Citation13 mesothelioma,Citation14 neuroblastoma,Citation15 non-small cell lung cancer,Citation16 ovarian cancer,Citation17 prostate cancerCitation18-Citation21 and small-cell lung cancerCitation22,Citation23 (). α subunits are also expressed in gliomas,Citation24,Citation25 lymphomaCitation26 and leukemia cells,Citation27 the latter suggesting that VGSCs may be present in hematological malignancies, in addition to solid tumors. Although the majority of reports published to date have focused on cell lines, a number of studies now show that α subunits are also expressed in vivo, in patient biopsy material.Citation8-Citation11,Citation17,Citation28-Citation30
Table 1. VGSC subtype expression in cancer
In excitable cells/tissues, different α subunits have subtly individual, but often largely overlapping, tissue distributions, which are proposed to permit functional specializations as a result of subtle variations in electrophysiological properties.Citation31 In cancer cells, there appears to be a variable pattern of expression of different α subunits, such that a number of cancers express multiple α subunits, but not all subtypes are expressed in all cancers (). In several cancers expressing multiple α subunits, a predominant α subunit has been identified. For example, in lymphoma and breast cancer cells, the most highly expressed α subunit is Nav1.5 (gene: SCN5A),Citation8,Citation26 whereas in prostate cancer cells, the predominant α subunit is Nav1.7 (gene: SCN9A).Citation18
Alternative mRNA splicing enables further functional variation among α subunits.Citation32 An important developmentally regulated splicing event occurs in exon 6, encoding the domain I segment 3 (DI:S3) region, such that the 5′ “neonatal” variant is expressed at birth, whereas the 3′ “adult” variant is expressed later in postnatal development.Citation33 In lymphoma, neuroblastoma, breast and prostate cancer cells, SCN5A and SCN9A are mainly expressed in their DI:S3 5′ neonatal splice forms.Citation8,Citation15,Citation26,Citation28 In contrast, the adult SCN5A variant is expressed in colon cancer cells, and the neonatal variant is absent.Citation11 In certain cancers, the presence of α subunits may therefore be an example of oncofetal gene expression, where embryonic genes are pathologically re-expressed during oncogenesis.Citation34
In several cancers, α subunit mRNA and protein expression correlates with metastatic potential. For example, in breast cancer, the neonatal SCN5A splice variant is expressed ~1,800-fold higher in metastatic MDA-MB-231 cells than weakly metastatic MCF-7 cells. Na+ currents are detectable in MDA-MB-231 cells, but not in weakly metastatic MCF-7 cells.Citation7,Citation8 The expression of neonatal SCN5A mRNA in breast cancer biopsies correlates with occurrence of lymph node metastasis.Citation8 Furthermore, SCN5A mRNA is elevated in breast tumors from patients who had a recurrence, or died within five years, and associates with increased odds of developing metastasis.Citation30 A similar pattern has been observed in prostate cancer cells, where VGSC expression increases in line with metastatic potential in the LNCaP progression model.Citation35 In agreement with this, the predominant α subunit, SCN9A, is elevated in prostate cancer biopsies compared with non-cancerous prostate samples,Citation28 and is more highly expressed in strongly metastatic PC-3 and Mat-LyLu cells than weakly metastatic LNCaP and AT-2 cells.Citation18 Na+ currents have been detected only in the metastatic prostate cancer cell lines.Citation19,Citation21 The positive correlation between α subunit expression and metastatic potential has also been reported for colonCitation11 and ovarian cancers.Citation17 However, in gliomas, the mRNA level of α subunits is inversely correlated with malignancy grade.Citation25 There appears to be no association between α subunit mRNA expression and metastatic potential of small cell or non-small cell lung cancer cell lines.Citation16,Citation22 Therefore, the relationship between α subunit expression, tumor grade and metastatic potential may be cancer type-specific.
The expression of β subunits in cancer cells has been less extensively studied. β subunits are expressed in prostate,Citation36,Citation37 breast,Citation38 non-small cell lungCitation16 and cervical cancersCitation10 (). As with α subunits, the β subunit expression profile appears to vary between cancers. For example, β3 is present in prostate and non-small cell lung cancer cells,Citation16,Citation36 but is absent in breast and cervical cancer cells.Citation10,Citation38 However, β1 is the most abundant β subunit in breast, prostate and cervical cancer cells.Citation10,Citation36,Citation38 Interestingly, β1 appears to be inversely correlated with SCN5A and metastatic potential in breast cancer cells: SCN1B mRNA (encoding β1) is significantly higher in weakly metastatic MCF-7 cells than in metastatic MDA-MB-231 cells.Citation38 This suggests that β subunits may be performing certain functions in cancer cells independent of the pore-forming α subunits. In contrast, a recent study has shown that β2 expression increases in line with metastatic potential in the LNCaP prostate cancer progression model.Citation37 Therefore, as with α subunits, different β subunits may be expressed at varying levels in different cancer types, and may perform distinct functions.
Functional Role
VGSC α subunits potentiate a number of cellular behaviors associated with metastasis (). In breast, prostate and lung cancer cell lines, the VGSC pore-blocker tetrodotoxin (TTX) inhibits behaviors including process outgrowth/extension,Citation39 galvanotaxis,Citation8,Citation40 migration,Citation8,Citation14,Citation17,Citation41-Citation44 endocytosis,Citation8,Citation22,Citation45 vesicular patterning,Citation46,Citation47 detachment from substrate,Citation48 gene expressionCitation43,Citation49 and invasion.Citation7,Citation8,Citation10-Citation12,Citation16,Citation17,Citation19,Citation21,Citation26 TTX does not inhibit proliferation of cancer cells,Citation7,Citation8,Citation10,Citation16,Citation17,Citation41 suggesting that VGSCs may be involved mainly in metastatic progression, rather than tumorigenesis.Citation50 However, recent evidence has shown that VGSCs also regulate angiogenic properties of endothelial cells, including vascular endothelial growth factor (VEGF)-induced proliferation, tubular differentiation and adhesion.Citation51 Therefore, the exact functional contribution of VGSCs to the cancer process may depend on the cell type, fate and state of the tumor.
Table 2. Metastatic cell behaviors regulated by VGSCs
Silencing SCN5A with siRNA reduces in vitro invasion of MDA-MB-231 breast cancer cells.Citation52 Further, specifically targeting the neonatal splice variant of SCN5A reduces the migration and invasion of MDA-MB-231 cells, suggesting that the neonatal form itself may be responsible for VGSC-dependent potentiation of metastatic cell behavior in breast cancer cells.Citation42 Similarly, siRNA targeting SCN8A (encoding Nav1.6) and SCN9A reduces invasion and endocytic activity in PC-3 metastatic prostate cancer cells.Citation53 Finally, the Nav1.6-specific toxin Cn2 inhibits the invasion of cervical cancer cells.Citation10 Thus, in different cancers, different α subunits appear to promote metastatic cell behaviors. However, it is not yet clear whether expression of a specific α subunit in a particular cancer provides a specific functional advantage, or is related to the natural history of the disease. The fact that any α subunit is present in a cancer cell may be more important than which α subunit it is. For example, overexpression of Nav1.4 in weakly metastatic LNCaP prostate cancer cells is necessary and sufficient to increase their invasiveness, even though the predominant α subunit expressed in metastatic prostate cancer cells is Nav1.7.Citation18,Citation35
Several studies have indicated that a number of therapeutically relevant small molecule VGSC blockers can also inhibit cell behaviors associated with metastasis. For example, the anticonvulsants phenytoin and carbamazepine inhibit secretion of prostate-specific antigen and interleukin-6 by prostate cancer cells.Citation54 Phenytoin and the local anesthetic lidocaine also inhibit endocytic activity in small cell lung cancer cells.Citation22 In addition, phenytoin suppresses migration of prostate cancer cells.Citation41 Furthermore, we have recently shown that therapeutically relevant concentrations of phenytoin inhibit Na+ current, migration and invasion in metastatic breast cancer cells.Citation30
Persistent Na+ Current
Cancer cells typically have a relatively depolarized membrane potential compared with terminally differentiated cells, e.g., epithelia, neurons.Citation55 For example, the resting membrane potential of a typical neuron may be around -65 mV,Citation56 whereas, we and others have shown that metastatic MDA-MB-231 breast cancer cells have membrane potentials between ~-15–30 mV.Citation7,Citation8,Citation30 Following depolarization, VGSCs open and rapidly inactivate within a few milliseconds, remaining inactivated until the membrane repolarizes.Citation3 Therefore, in cancer cells with depolarized resting potentials, the majority of VGSCs will be inactivated. However, several VGSCs, including Nav1.5, do not inactivate completely, and a steady-state Na+ current persists, which is typically a few percent of the peak transient current.Citation57,Citation58 We recently proposed that the persistent Na+ current is likely to be predominant in cancer cells expressing VGSCs, and this component of the Na+ current may specifically potentiate the cells’ migration and invasion.Citation30
Mechanisms of Action
The obvious question is: how does Na+ influx through VGSCs potentiate metastatic behaviors, including invasion? Over the years, there has been much speculation in the literature (reviewed in ref. Citation50). Three models are considered below, based on recently published experimental data.
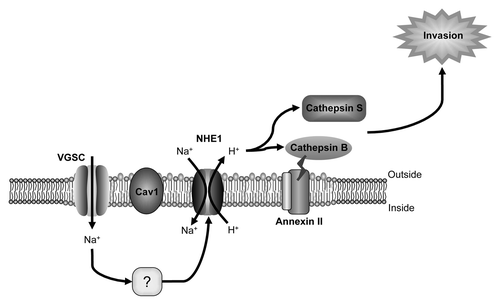
(1) Regulation of pH. In MDA-MB-231 cells, Na+ influx through Nav1.5 results in intracellular alkalinization, and extracellular acidification adjacent to the plasma membrane.Citation52 The Na+/H+ exchanger NHE1, which is an important regulator of H+ efflux, is co-expressed with Nav1.5 in lipid rafts contained within the caveolae of MDA-MB-231 cells.Citation59 Na+ influx through Nav1.5 increases H+ efflux through NHE1, thus enhancing pH-dependent extracellular matrix degradation and invasion ().Citation59 However, the precise mechanism by which Nav1.5 activity enhances NHE1 is not yet clear. The resultant (Nav1.5/NHE1-dependent) perimembrane acidification is proposed to favor the proteolytic activity of cysteine cathepsins B and S, the function of which has been shown to depend, at least in part, on VGSC activity.Citation52 A similar mechanism has been identified by which Nav1.5 expressed on intracellular endosomal membranes of primed macrophages acts as a charge sink, permitting Na+ efflux from the endosome, resulting in H+ influx, likely via the vesicular ATPase, and subsequent endosomal acidification.Citation60
Figure 2. α subunit involvement in pH-dependent cellular invasion. Na+ influx through Nav1.5 is proposed to activate the Na+/H+ exchanger NHE1, co-expressed with Nav1.5 in lipid rafts contained within the caveolae of invasive breast cancer cells.Citation59 Increased NHE1 activity results in increased H+ efflux, which, in turn, enhances proteolytic activity of cysteine cathepsins B and S, which degrade the extracellular matrix, permitting invasion.Citation52,Citation59 The mechanism by which Nav1.5 enhances NHE1 is not yet clear. Figure was produced using Science Slides 2006 software.
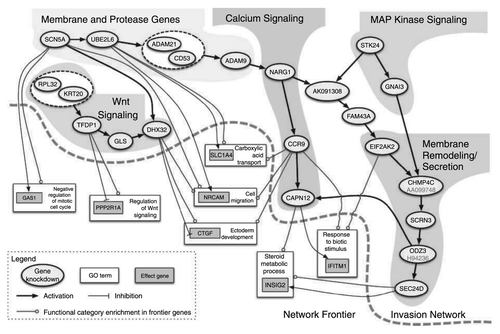
(2) Regulation of gene expression. Several studies have shown that VGSCs regulate gene expression, both in excitable cells, e.g., neurons and cardiomyocytes, and in cancer cells.Citation38,Citation43,Citation49,Citation61,Citation62 In silico factor graph nested effects modeling of gene expression in colon cancer cell lines has revealed a novel network of gene interactions that are implicated in cancer invasion.Citation11 Strikingly, SCN5A is a key regulator of this invasion gene network, suggesting that VGSCs, in particular Nav1.5, may function as early entry points in signaling mechanisms regulating invasion. Downstream gene ontology categories include Wnt signaling, cell migration, ectoderm development, response to biotic stimulus, steroid metabolic process and cell cycle regulation ().Citation11 These data suggest that, at least in colon cancer, Nav1.5 may regulate invasion by mechanism(s) in addition to/instead of H+ efflux.Citation59 The challenge now is to understand how Na+ current mediated by Nav1.5 may regulate transcription in cancer cells, or, indeed, whether the effect is mediated by mechanism(s) independent of ion conduction.
Figure 3. Nav1.5-regulated gene transcriptional network controlling invasion. Oval nodes represent regulatory genes, gray boxes represent effector genes and white boxes represent gene ontology categories. Arrows indicate activation and tees show repression. Reprinted by permission from the American Association for Cancer Research: House CD et al., Voltage-gated Na+ channel SCN5A is a key regulator of a gene transcriptional network that controls colon cancer invasion. Cancer Res 2010; 70:6957-67; PMID:20651255.
(3) Regulation of intracellular Ca2+. In excitable cells, Na+ current carried by VGSCs can result in an increase in intracellular Ca2+ level, e.g., by activating voltage-gated Ca2+ channels.Citation63,Citation64 Similar VGSC-dependent elevation of intracellular Ca2+ has been reported in non-excitable cells. For example, Na+ current carried by Nav1.5 is essential for the sustained Ca2+ entry into CD4+ T cells that occurs during positive selection.Citation65 In addition to being expressed at the plasma membrane, α subunits are also present on internal membranes of cancer cells and macrophages.Citation12,Citation30,Citation60 In THP-1 macrophages and HTB-66 melanoma cells, Nav1.6 is expressed on vesicular structures adjacent to podosomes.Citation12 Agonist-mediated activation of VGSCs in these cells causes Na+ release from cationic intracellular stores, followed by rapid Na+ uptake by anionic mitochondria and subsequent Ca2+ release into the cytosol.Citation12 It is proposed that this Ca2+ release then enhances podosome/invadopodia formation, leading to increased invasion.Citation12 However, it is not yet clear how VGSCs present on vesicular membranes are gated, and/or whether they interact with VGSCs present at the plasma membrane. In vascular endothelial cells, VGSC-mediated Na+ influx is required for VEGF-induced membrane depolarization and elevation of intracellular Ca2+, which, in turn, activates PKC and extracellular signal-regulated kinase (ERK)1/2, potentiating angiogenic functions including proliferation, differentiation and adhesion.Citation51 In contrast to cancer cells and macrophages, the Ca2+ rise in endothelial cells occurs as a result of Ca2+ influx through reverse-mode operation of the Na+/Ca2+ exchanger NCX.Citation66
In summary, several models have recently emerged, supported by experimental data, suggesting that VGSC α subunits may enhance cellular invasion by a variety of mechanisms. Further studies will no doubt contribute further data in order to clarify whether the mechanisms discussed above are widely applicable to all cancers in which VGSCs are expressed, or whether different VGSCs perform distinct functions in different cells in different cancers.
The Role of β Subunits
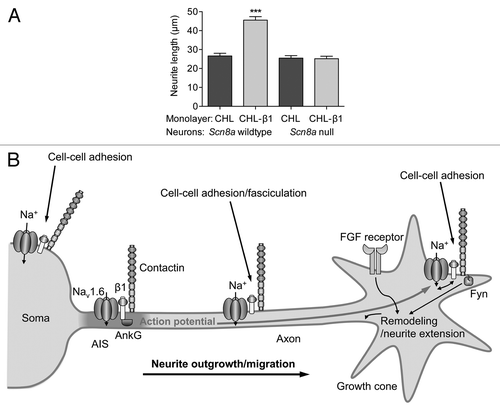
In breast cancer, β1 is highly expressed in weakly metastatic MCF-7 cells, where it enhances adhesion and retards transwell migration.Citation38 β1 is expressed at a much lower level in strongly metastatic MDA-MB-231 cells, and stable overexpression of β1 in this line increases cell-cell adhesion, induces process outgrowth and reduces migration in wound-healing assays.Citation38 β2 appears to play a slightly different role in prostate cancer cells. Overexpression of β2 in LNCaP cells increases adhesion, process outgrowth, migration and invasion.Citation37 However, the same study reported that β2-overexpressing LNCaP cells have a reduced tumor take and smaller tumor volume when subcutaneously injected into nude mice. Thus, not only do β1 and β2 play subtly different functional roles in different cancer cells, these subunits may function differently in tumors in vivo, compared with cells in culture. This highlights the critical importance of studying the functional role of VGSCs in cancer in vivo, in addition to using in vitro models. Nonetheless, given that β subunits are Ig family cell adhesion molecules, it is plausible to speculate that their major contribution to the metastatic behaviors of cancer cells may be through regulated adhesion/detachment. Of special note is that β subunits play a critical role in regulating adhesion and migration in excitable cells, where they are normally expressed.Citation2 For example, β1 enhances neurite outgrowth, neuronal pathfinding and fasciculation during early postnatal central nervous system development.Citation67,Citation68 β1-mediated neurite outgrowth in cerebellar granule neurons requires fyn kinase, the cell adhesion molecule contactin, and is dependent on Na+ current carried by Nav1.6Citation2,Citation68 (). In turn, Nav1.6-dependent resurgent Na+ current and action potential firing are dependent on the presence of β1Citation56 (). Thus, in neurons, there is a functional relationship between α and β subunits, such that both are required for neurite outgrowth, migration and electrical excitability. This is an interesting contrast with β1 and Nav1.5 in breast cancer cells, which appear to play opposing, antagonistic roles in regulating adhesion and migration.Citation38
Figure 4. Neurite outgrowth and excitability regulated by β1 and Nav1.6. (A) β1-mediated neurite outgrowth is inhibited by the Scn8a null-mutation. Neurite lengths of wild-type and Scn8a-null cerebellar granule neurons grown on monolayers of control and b1-expressing fibroblasts. Data are mean and SEM ***p < 0.001. (B) Proposed model for Na+ current reciprocal involvement in β1-mediated neurite outgrowth. Complexes containing Nav1.6, β1 and contactin are present throughout the neuronal membrane in the soma, neurite and growth cone. Na+ influx is required for β1-mediated neurite extension and migration. VGSC complexes along the neurite participate in cell-cell adhesion and fasciculation. β1 is also required for Nav1.6 expression at the axon initial segment, and subsequent high-frequency action potential firing. Electrical activity may further promote β1-mediated neurite outgrowth at or near the growth cone. Figure reproduced with permission.Citation56
β subunits regulate the mRNA levels of α subunits, including Nav1.5 and other β subunits, in neurons and cardiomyocytes.Citation61,Citation69-Citation71 Downregulation of SCN1B in MCF-7 cells results in an increase in the mRNA level of the neonatal SCN5A splice variant,Citation38 raising the possibility that β1 may regulate gene transcription, and supporting the notion that β1 and Nav1.5 may function antagonistically in breast cancer cells.Citation30 However, their expression may not be mutually exclusive, and both subunits colocalize in lamellipodia.Citation30
Less is known about the expression/function of the other β subunits (β3 and β4) in cancer cells. However, SCN3B contains two response elements to the tumor suppressor p53, and DNA damaging agents and p53 overexpression upregulate SCN3B mRNA in cancer cell lines, suggesting that β3 may be involved in p53-dependent apoptosis.Citation72 The function of β4 in cancer cells has not been investigated. However, a recent study has shown that, together with Nav1.5, β4 is required for CD4+ T cell development,Citation65 suggesting that β4 may play an as-yet-unexplored role(s) in non-excitable cells.
In summary, VGSC β subunits appear to play a role in regulating a number of cell behaviors associated with metastasis, including adhesion, migration and gene expression. Importantly, some of these functions oppose the effects of α subunitsCitation38 (). Further work is required to delineate the respective functions of α and β subunits in cancer cells. A major challenge will be to establish whether or not β subunits are functioning (e.g., to regulate adhesion and migration) independently of α subunits.
Table 3. Complementary functions of VGSC α and β subunits in neurons and cancer cells
Regulation of Expression
How are VGSCs upregulated in cancer cells? VGSCs are macromolecular signaling complexes and their activity can be regulated at multiple levels by a vast array of different interacting partners and mechanisms, which have been studied extensively in excitable cells, e.g., neurons and cardiomyocytes (; reviewed in refs. Citation2 and Citation73). However, the mechanisms by which expression of VGSC α and β subunits are regulated in cancer cells are not yet well understood. Serum has been shown to modulate expression and activity of α subunits in Mat-LyLu prostate cancer cells.Citation74 Subsequent studies have shown that several components of serum have various effects on VGSC function. For example, epidermal growth factor (EGF) increases Nav1.7 expression, Na+ current and VGSC-dependent migration and invasion of Mat-LyLu and PC-3M prostate cancer cells.Citation44,Citation75 In addition, nerve growth factor (NGF) increases Na+ current in Mat-LyLu cells via protein kinase A (PKA) activation.Citation76 Although NGF also increases migration in these cells, its effect is independent of VGSC activity,Citation76 suggesting that different growth factors may regulate different pools of VGSCs, which may in turn have different downstream effects on metastatic cell behavior.
Steroid hormones have also been shown to regulate VGSC expression and activity in cancer cells. In MDA-MB-231 breast cancer cells, the estrogen β-estradiol increases Na+ current via the G-protein-coupled estrogen receptor GPR30 and PKA.Citation77 In addition, SCN1B expression is negatively regulated by the androgen dihydrotestosterone (DHT) in LNCaP and PC-3 prostate cancer cells.Citation36 However, there is no association between SCN5A expression and estrogen receptor α (ER), progesterone receptor (PR) or human epidermal growth factor 2 (HER2) status in clinical breast cancer specimens,Citation30 suggesting that any relationship between serum factors and VGSC expression in vivo may be highly complex.
Finally, α subunit expression and activity in metastatic cancer cells appears to be auto-regulated via a positive feedback mechanism. In Mat-LyLu cells, Na+ current activates PKA, which in turn upregulates SCN9A mRNA levels, and promotes externalization of α subunits to the plasma membrane.Citation43 Similarly, in MDA-MB-231 breast cancer cells, expression of the neonatal Nav1.5 splice variant is maintained by positive feedback involving PKA.Citation78 In both cases, inhibition of Na+ current with TTX effectively collapses the positive feedback loop, suppressing VGSC-dependent migration.Citation43,Citation78 In summary, expression and function of VGSC α and β subunits in cancer cells is regulated by serum components, including steroid hormones and growth factors.
Therapeutic Potential
The observation that VGSCs play a pro-invasive role in metastatic cells, together with patient data associating α subunit expression with poor clinical outcome, suggests that VGSCs may be important diagnostic and/or therapeutic targets in the metastatic setting. Given that α subunits are regulated by activity-dependent positive feedback, pharmacological blockade of channel conductance may be an ideal intervention to inhibit VGSC-dependent metastatic cell behaviors. Indeed, a recent study shows that local injection of TTX into subcutaneous tumors of Mat-LyLu cells in the Copenhagen rat significantly reduces lung metastases and improves overall survival.Citation79 However, the toxicity of TTX would preclude its use as a systemic anti-metastatic treatment.
We have proposed that VGSC-blocking drugs FDA-approved for treating other conditions, e.g., anticonvulsants, antiarrhythmics, local anesthetics and tricyclic antidepressants, may warrant investigation for repurposing to metastatic disease.Citation30 In support of this, several of these agents inhibit metastatic cell behaviors in vitro models.Citation8,Citation22,Citation30,Citation41,Citation54 Application of local anesthetics during radical prostatectomy surgery associates with significantly reduced recurrence and metastasis.Citation80 In addition, a recent retrospective case-control analysis using the General Practice Research Database suggests that use of tricyclic antidepressants may have potential for prevention of glioma and colorectal cancer.Citation81 Furthermore, riluzole, which is both a VGSC blocker and metabotropic glutamate receptor inhibitor, reduces breast cancer tumor volume in mice, and suppresses metabolic activity of tumors in patients with resectable stage III and IV melanoma.Citation82,Citation83 Finally, a mexiletine analog (RS100642) reduces oxidative stress associated with tumor burden, and increases overall survival in the experimentally induced 7,12-dimethylbenz(a) anthracene (DMBA) rat breast cancer model.Citation84 In summary, several studies suggest that VGSC α subunits may be useful therapeutic targets in cancer.
Conclusion and Perspectives
A growing body of evidence suggests that VGSCs play an important pathological role during cancer progression toward metastasis. However, the role of individual α and β subunits appears to be complex. Nonetheless, a common theme is that Na+ current carried by α subunits favors an invasive phenotype, whereas β subunits may regulate adhesion. Future work is required to establish how widely VGSCs are expressed across different types of cancer, and the extent of contribution(s) of different α and β subunits to disease progression. The ultimate goal should be to definitively evaluate the potential for both α and β subunits as diagnostic and therapeutic targets. In respect of the former, repurposing FDA-approved channel blockers may be a cost-effective intervention in metastatic disease, which is the major cause of cancer-related deaths,Citation85,Citation86 and treatment of which is still largely limited to palliation.Citation87
| Abbreviations: | ||
| CAM | = | cell adhesion molecule |
| DHT | = | dihydrotestosterone |
| DMBA | = | 7,12-dimethylbenz(a) anthracene |
| EGF | = | epidermal growth factor |
| ER | = | estrogen receptor α ERK, extracellular signal-regulated kinase |
| FHF1B | = | fibroblast growth factor-homologous factor 1B |
| HER2 | = | human epidermal growth factor 2 |
| Ig | = | immunoglobulin |
| NF | = | neurofascin |
| NGF | = | nerve growth factor |
| PKA | = | protein kinase A |
| PKC | = | protein kinase C |
| PR | = | progesterone receptor |
| RPTPβ | = | receptor protein tyrosine phosphatase β |
| TTX | = | tetrodotoxin |
| VEGF | = | vascular endothelial growth factor |
| VGSC | = | voltage-gated Na+ channel |
Acknowledgments
This work was supported by the Medical Research Council [Fellowship number G1000508(95657)].
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- Catterall WA. Cellular and molecular biology of voltage-gated sodium channels. Physiol Rev 1992; 72:S15 - 48; PMID: 1332090
- Brackenbury WJ, Isom LL. Na Channel β Subunits: Overachievers of the Ion Channel Family. Front Pharmacol 2011; 2:53; http://dx.doi.org/10.3389/fphar.2011.00053; PMID: 22007171
- Hille B. Ionic channels of excitable membranes. Sunderland (Massachusetts): Sinauer Associates Inc., 1992.
- Brackenbury WJ, Isom LL. Voltage-gated Na+ channels: potential for beta subunits as therapeutic targets. Expert Opin Ther Targets 2008; 12:1191 - 203; http://dx.doi.org/10.1517/14728222.12.9.1191; PMID: 18694383
- Brackenbury WJ, Djamgoz MB, Isom LL. An emerging role for voltage-gated Na+ channels in cellular migration: regulation of central nervous system development and potentiation of invasive cancers. Neuroscientist 2008; 14:571 - 83; http://dx.doi.org/10.1177/1073858408320293; PMID: 18940784
- Catterall WA. From ionic currents to molecular mechanisms: the structure and function of voltage-gated sodium channels. Neuron 2000; 26:13 - 25; http://dx.doi.org/10.1016/S0896-6273(00)81133-2; PMID: 10798388
- Roger S, Besson P, Le Guennec JY. Involvement of a novel fast inward sodium current in the invasion capacity of a breast cancer cell line. Biochim Biophys Acta 2003; 1616:107 - 11; http://dx.doi.org/10.1016/j.bbamem.2003.07.001; PMID: 14561467
- Fraser SP, Diss JK, Chioni AM, Mycielska ME, Pan H, Yamaci RF, et al. Voltage-gated sodium channel expression and potentiation of human breast cancer metastasis. Clin Cancer Res 2005; 11:5381 - 9; http://dx.doi.org/10.1158/1078-0432.CCR-05-0327; PMID: 16061851
- Diaz D, Delgadillo DM, Hernández-Gallegos E, Ramírez-Domínguez ME, Hinojosa LM, Ortiz CS, et al. Functional expression of voltage-gated sodium channels in primary cultures of human cervical cancer. J Cell Physiol 2007; 210:469 - 78; http://dx.doi.org/10.1002/jcp.20871; PMID: 17051596
- Hernandez-Plata E, Ortiz CS, Marquina-Castillo B, Medina-Martinez I, Alfaro A, Berumen J, et al. Overexpression of NaV 1.6 channels is associated with the invasion capacity of human cervical cancer. Int J Cancer 2012; 130:2013 - 23; http://dx.doi.org/10.1002/ijc.26210; PMID: 21630263
- House CD, Vaske CJ, Schwartz AM, Obias V, Frank B, Luu T, et al. Voltage-gated Na+ channel SCN5A is a key regulator of a gene transcriptional network that controls colon cancer invasion. Cancer Res 2010; 70:6957 - 67; http://dx.doi.org/10.1158/0008-5472.CAN-10-1169; PMID: 20651255
- Carrithers MD, Chatterjee G, Carrithers LM, Offoha R, Iheagwara U, Rahner C, et al. Regulation of podosome formation in macrophages by a splice variant of the sodium channel SCN8A. J Biol Chem 2009; 284:8114 - 26; http://dx.doi.org/10.1074/jbc.M801892200; PMID: 19136557
- Allen DH, Lepple-Wienhues A, Cahalan MD. Ion channel phenotype of melanoma cell lines. J Membr Biol 1997; 155:27 - 34; http://dx.doi.org/10.1007/s002329900155; PMID: 9002422
- Fulgenzi G, Graciotti L, Faronato M, Soldovieri MV, Miceli F, Amoroso S, et al. Human neoplastic mesothelial cells express voltage-gated sodium channels involved in cell motility. Int J Biochem Cell Biol 2006; 38:1146 - 59; http://dx.doi.org/10.1016/j.biocel.2005.12.003; PMID: 16458569
- Ou SW, Kameyama A, Hao LY, Horiuchi M, Minobe E, Wang WY, et al. Tetrodotoxin-resistant Na+ channels in human neuroblastoma cells are encoded by new variants of Nav1.5/SCN5A. Eur J Neurosci 2005; 22:793 - 801; http://dx.doi.org/10.1111/j.1460-9568.2005.04280.x; PMID: 16115203
- Roger S, Rollin J, Barascu A, Besson P, Raynal PI, Iochmann S, et al. Voltage-gated sodium channels potentiate the invasive capacities of human non-small-cell lung cancer cell lines. Int J Biochem Cell Biol 2007; 39:774 - 86; http://dx.doi.org/10.1016/j.biocel.2006.12.007; PMID: 17307016
- Gao R, Shen Y, Cai J, Lei M, Wang Z. Expression of voltage-gated sodium channel alpha subunit in human ovarian cancer. Oncol Rep 2010; 23:1293 - 9; PMID: 20372843
- Diss JK, Archer SN, Hirano J, Fraser SP, Djamgoz MB. Expression profiles of voltage-gated Na(+) channel alpha-subunit genes in rat and human prostate cancer cell lines. Prostate 2001; 48:165 - 78; http://dx.doi.org/10.1002/pros.1095; PMID: 11494332
- Laniado ME, Lalani EN, Fraser SP, Grimes JA, Bhangal G, Djamgoz MB, et al. Expression and functional analysis of voltage-activated Na+ channels in human prostate cancer cell lines and their contribution to invasion in vitro. Am J Pathol 1997; 150:1213 - 21; PMID: 9094978
- Smith P, Rhodes NP, Shortland AP, Fraser SP, Djamgoz MB, Ke Y, et al. Sodium channel protein expression enhances the invasiveness of rat and human prostate cancer cells. FEBS Lett 1998; 423:19 - 24; http://dx.doi.org/10.1016/S0014-5793(98)00050-7; PMID: 9506834
- Grimes JA, Fraser SP, Stephens GJ, Downing JE, Laniado ME, Foster CS, et al. Differential expression of voltage-activated Na+ currents in two prostatic tumour cell lines: contribution to invasiveness in vitro. FEBS Lett 1995; 369:290 - 4; http://dx.doi.org/10.1016/0014-5793(95)00772-2; PMID: 7649275
- Onganer PU, Djamgoz MB. Small-cell lung cancer (human): potentiation of endocytic membrane activity by voltage-gated Na(+) channel expression in vitro. J Membr Biol 2005; 204:67 - 75; http://dx.doi.org/10.1007/s00232-005-0747-6; PMID: 16151702
- Blandino JK, Viglione MP, Bradley WA, Oie HK, Kim YI. Voltage-dependent sodium channels in human small-cell lung cancer cells: role in action potentials and inhibition by Lambert-Eaton syndrome IgG. J Membr Biol 1995; 143:153 - 63; http://dx.doi.org/10.1007/BF00234661; PMID: 7731034
- Joshi AD, Parsons DW, Velculescu VE, Riggins GJ. Sodium ion channel mutations in glioblastoma patients correlate with shorter survival. Mol Cancer 2011; 10:17; http://dx.doi.org/10.1186/1476-4598-10-17; PMID: 21314958
- Schrey M, Codina C, Kraft R, Beetz C, Kalff R, Wölfl S, et al. Molecular characterization of voltage-gated sodium channels in human gliomas. Neuroreport 2002; 13:2493 - 8; http://dx.doi.org/10.1097/00001756-200212200-00023; PMID: 12499855
- Fraser SP, Diss JK, Lloyd LJ, Pani F, Chioni AM, George AJ, et al. T-lymphocyte invasiveness: control by voltage-gated Na+ channel activity. FEBS Lett 2004; 569:191 - 4; http://dx.doi.org/10.1016/j.febslet.2004.05.063; PMID: 15225632
- Yamashita N, Hamada H, Tsuruo T, Ogata E. Enhancement of voltage-gated Na+ channel current associated with multidrug resistance in human leukemia cells. Cancer Res 1987; 47:3736 - 41; PMID: 2439197
- Diss JK, Stewart D, Pani F, Foster CS, Walker MM, Patel A, et al. A potential novel marker for human prostate cancer: voltage-gated sodium channel expression in vivo. Prostate Cancer Prostatic Dis 2005; 8:266 - 73; http://dx.doi.org/10.1038/sj.pcan.4500796; PMID: 16088330
- Onganer PU, Seckl MJ, Djamgoz MB. Neuronal characteristics of small-cell lung cancer. Br J Cancer 2005; 93:1197 - 201; http://dx.doi.org/10.1038/sj.bjc.6602857; PMID: 16265346
- Yang M, Kozminski DJ, Wold LA, Modak R, Calhoun JD, Isom LL, et al. Therapeutic potential for phenytoin: targeting Na(v)1.5 sodium channels to reduce migration and invasion in metastatic breast cancer. Breast Cancer Res Treat 2012; 134:603 - 15; http://dx.doi.org/10.1007/s10549-012-2102-9; PMID: 22678159
- Clare JJ, Tate SN, Nobbs M, Romanos MA. Voltage-gated sodium channels as therapeutic targets. Drug Discov Today 2000; 5:506 - 20; http://dx.doi.org/10.1016/S1359-6446(00)01570-1; PMID: 11084387
- Diss JK, Fraser SP, Djamgoz MB. Voltage-gated Na+ channels: multiplicity of expression, plasticity, functional implications and pathophysiological aspects. Eur Biophys J 2004; 33:180 - 93; http://dx.doi.org/10.1007/s00249-004-0389-0; PMID: 14963621
- Onkal R, Mattis JH, Fraser SP, Diss JK, Shao D, Okuse K, et al. Alternative splicing of Nav1.5: an electrophysiological comparison of ‘neonatal’ and ‘adult’ isoforms and critical involvement of a lysine residue. J Cell Physiol 2008; 216:716 - 26; http://dx.doi.org/10.1002/jcp.21451; PMID: 18393272
- Monk M, Holding C. Human embryonic genes re-expressed in cancer cells. Oncogene 2001; 20:8085 - 91; http://dx.doi.org/10.1038/sj.onc.1205088; PMID: 11781821
- Bennett ES, Smith BA, Harper JM. Voltage-gated Na+ channels confer invasive properties on human prostate cancer cells. Pflugers Arch 2004; 447:908 - 14; http://dx.doi.org/10.1007/s00424-003-1205-x; PMID: 14677067
- Diss JK, Fraser SP, Walker MM, Patel A, Latchman DS, Djamgoz MB. Beta-subunits of voltage-gated sodium channels in human prostate cancer: quantitative in vitro and in vivo analyses of mRNA expression. Prostate Cancer Prostatic Dis 2008; 11:325 - 33; http://dx.doi.org/10.1038/sj.pcan.4501012; PMID: 17893701
- Jansson KH, Lynch JE, Lepori-Bui N, Czymmek KJ, Duncan RL, Sikes RA. Overexpression of the VSSC-associated CAM, β-2, enhances LNCaP cell metastasis associated behavior. Prostate 2012; 72:1080 - 92; http://dx.doi.org/10.1002/pros.21512; PMID: 22127840
- Chioni AM, Brackenbury WJ, Calhoun JD, Isom LL, Djamgoz MB. A novel adhesion molecule in human breast cancer cells: voltage-gated Na+ channel β1 subunit. Int J Biochem Cell Biol 2009; 41:1216 - 27; http://dx.doi.org/10.1016/j.biocel.2008.11.001; PMID: 19041953
- Fraser SP, Ding Y, Liu A, Foster CS, Djamgoz MB. Tetrodotoxin suppresses morphological enhancement of the metastatic MAT-LyLu rat prostate cancer cell line. Cell Tissue Res 1999; 295:505 - 12; http://dx.doi.org/10.1007/s004410051256; PMID: 10022970
- Mycielska M, Madeja Z, Fraser SP, Korohoda W, Korohoda W, Djamgoz MBA. Directional movement of rat prostate cancer cells in direct-current electric field: involvement of voltagegated Na+ channel activity. J Cell Sci 2001; 114:2697 - 705; PMID: 11683396
- Fraser SP, Salvador V, Manning EA, Mizal J, Altun S, Raza M, et al. Contribution of functional voltage-gated Na+ channel expression to cell behaviors involved in the metastatic cascade in rat prostate cancer: I. Lateral motility. J Cell Physiol 2003; 195:479 - 87; http://dx.doi.org/10.1002/jcp.10312; PMID: 12704658
- Brackenbury WJ, Chioni AM, Diss JK, Djamgoz MB. The neonatal splice variant of Nav1.5 potentiates in vitro invasive behaviour of MDA-MB-231 human breast cancer cells. Breast Cancer Res Treat 2007; 101:149 - 60; http://dx.doi.org/10.1007/s10549-006-9281-1; PMID: 16838113
- Brackenbury WJ, Djamgoz MB. Activity-dependent regulation of voltage-gated Na+ channel expression in Mat-LyLu rat prostate cancer cell line. J Physiol 2006; 573:343 - 56; http://dx.doi.org/10.1113/jphysiol.2006.106906; PMID: 16543264
- Uysal-Onganer P, Djamgoz MB. Epidermal growth factor potentiates in vitro metastatic behaviour of human prostate cancer PC-3M cells: involvement of voltage-gated sodium channel. Mol Cancer 2007; 6:76; http://dx.doi.org/10.1186/1476-4598-6-76; PMID: 18036246
- Mycielska ME, Fraser SP, Szatkowski M, Djamgoz MB. Contribution of functional voltage-gated Na+ channel expression to cell behaviors involved in the metastatic cascade in rat prostate cancer: II. Secretory membrane activity. J Cell Physiol 2003; 195:461 - 9; http://dx.doi.org/10.1002/jcp.10265; PMID: 12704656
- Krasowska M, Grzywna ZJ, Mycielska ME, Djamgoz MB. Patterning of endocytic vesicles and its control by voltage-gated Na+ channel activity in rat prostate cancer cells: fractal analyses. Eur Biophys J 2004; 33:535 - 42; http://dx.doi.org/10.1007/s00249-004-0394-3; PMID: 15024523
- Krasowska M, Grzywna ZJ, Mycielska ME, Djamgoz MB. Fractal analysis and ionic dependence of endocytotic membrane activity of human breast cancer cells. Eur Biophys J 2009; 38:1115 - 25; http://dx.doi.org/10.1007/s00249-009-0516-z; PMID: 19618177
- Palmer CP, Mycielska ME, Burcu H, Osman K, Collins T, Beckerman R, et al. Single cell adhesion measuring apparatus (SCAMA): application to cancer cell lines of different metastatic potential and voltage-gated Na+ channel expression. Eur Biophys J 2008; 37:359 - 68; http://dx.doi.org/10.1007/s00249-007-0219-2; PMID: 17879092
- Mycielska ME, Palmer CP, Brackenbury WJ, Djamgoz MB. Expression of Na+-dependent citrate transport in a strongly metastatic human prostate cancer PC-3M cell line: regulation by voltage-gated Na+ channel activity. J Physiol 2005; 563:393 - 408; http://dx.doi.org/10.1113/jphysiol.2004.079491; PMID: 15611019
- Onkal R, Djamgoz MB. Molecular pharmacology of voltage-gated sodium channel expression in metastatic disease: clinical potential of neonatal Nav1.5 in breast cancer. Eur J Pharmacol 2009; 625:206 - 19; http://dx.doi.org/10.1016/j.ejphar.2009.08.040; PMID: 19835862
- Andrikopoulos P, Fraser SP, Patterson L, Ahmad Z, Burcu H, Ottaviani D, et al. Angiogenic functions of voltage-gated Na+ Channels in human endothelial cells: modulation of vascular endothelial growth factor (VEGF) signaling. J Biol Chem 2011; 286:16846 - 60; http://dx.doi.org/10.1074/jbc.M110.187559; PMID: 21385874
- Gillet L, Roger S, Besson P, Lecaille F, Gore J, Bougnoux P, et al. Voltage-gated Sodium Channel Activity Promotes Cysteine Cathepsin-dependent Invasiveness and Colony Growth of Human Cancer Cells. J Biol Chem 2009; 284:8680 - 91; http://dx.doi.org/10.1074/jbc.M806891200; PMID: 19176528
- Nakajima T, Kubota N, Tsutsumi T, Oguri A, Imuta H, Jo T, et al. Eicosapentaenoic acid inhibits voltage-gated sodium channels and invasiveness in prostate cancer cells. Br J Pharmacol 2009; 156:420 - 31; http://dx.doi.org/10.1111/j.1476-5381.2008.00059.x; PMID: 19154441
- Abdul M, Hoosein N. Inhibition by anticonvulsants of prostate-specific antigen and interleukin-6 secretion by human prostate cancer cells. Anticancer Res 2001; 21:2045 - 8; PMID: 11497296
- Kunzelmann K. Ion channels and cancer. J Membr Biol 2005; 205:159 - 73; http://dx.doi.org/10.1007/s00232-005-0781-4; PMID: 16362504
- Brackenbury WJ, Calhoun JD, Chen C, Miyazaki H, Nukina N, Oyama F, et al. Functional reciprocity between Na+ channel Nav1.6 and β1 subunits in the coordinated regulation of excitability and neurite outgrowth. Proc Natl Acad Sci USA 2010; 107:2283 - 8; http://dx.doi.org/10.1073/pnas.0909434107; PMID: 20133873
- Crill WE. Persistent sodium current in mammalian central neurons. Annu Rev Physiol 1996; 58:349 - 62; http://dx.doi.org/10.1146/annurev.ph.58.030196.002025; PMID: 8815799
- Ju YK, Saint DA, Gage PW. Hypoxia increases persistent sodium current in rat ventricular myocytes. J Physiol 1996; 497:337 - 47; PMID: 8961179
- Brisson L, Gillet L, Calaghan S, Besson P, Le Guennec JY, Roger S, et al. Na(V)1.5 enhances breast cancer cell invasiveness by increasing NHE1-dependent H(+) efflux in caveolae. Oncogene 2011; 30:2070 - 6; http://dx.doi.org/10.1038/onc.2010.574; PMID: 21170089
- Carrithers MD, Dib-Hajj S, Carrithers LM, Tokmoulina G, Pypaert M, Jonas EA, et al. Expression of the voltage-gated sodium channel NaV1.5 in the macrophage late endosome regulates endosomal acidification. J Immunol 2007; 178:7822 - 32; PMID: 17548620
- Lopez-Santiago LF, Meadows LS, Ernst SJ, Chen C, Malhotra JD, McEwen DP, et al. Sodium channel Scn1b null mice exhibit prolonged QT and RR intervals. J Mol Cell Cardiol 2007; 43:636 - 47; http://dx.doi.org/10.1016/j.yjmcc.2007.07.062; PMID: 17884088
- Tolón RM, Sánchez-Franco F, López Fernández J, Lorenzo MJ, Vázquez GF, Cacicedo L. Regulation of somatostatin gene expression by veratridine-induced depolarization in cultured fetal cerebrocortical cells. Brain Res Mol Brain Res 1996; 35:103 - 10; http://dx.doi.org/10.1016/0169-328X(95)00188-X; PMID: 8717345
- Dravid SM, Baden DG, Murray TF. Brevetoxin activation of voltage-gated sodium channels regulates Ca dynamics and ERK1/2 phosphorylation in murine neocortical neurons. J Neurochem 2004; 89:739 - 49; http://dx.doi.org/10.1111/j.1471-4159.2004.02407.x; PMID: 15086530
- Fekete A, Franklin L, Ikemoto T, Rózsa B, Lendvai B, Sylvester Vizi E, et al. Mechanism of the persistent sodium current activator veratridine-evoked Ca elevation: implication for epilepsy. J Neurochem 2009; 111:745 - 56; http://dx.doi.org/10.1111/j.1471-4159.2009.06368.x; PMID: 19719824
- Lo WL, Donermeyer DL, Allen PM. A voltage-gated sodium channel is essential for the positive selection of CD4(+) T cells. Nat Immunol 2012; 13:880 - 7; http://dx.doi.org/10.1038/ni.2379; PMID: 22842345
- Andrikopoulos P, Baba A, Matsuda T, Djamgoz MB, Yaqoob MM, Eccles SA. Ca2+ influx through reverse mode Na+/Ca2+ exchange is critical for vascular endothelial growth factor-mediated extracellular signal-regulated kinase (ERK) 1/2 activation and angiogenic functions of human endothelial cells. J Biol Chem 2011; 286:37919 - 31; http://dx.doi.org/10.1074/jbc.M111.251777; PMID: 21873429
- Davis TH, Chen C, Isom LL. Sodium channel β1 subunits promote neurite outgrowth in cerebellar granule neurons. J Biol Chem 2004; 279:51424 - 32; http://dx.doi.org/10.1074/jbc.M410830200; PMID: 15452131
- Brackenbury WJ, Davis TH, Chen C, Slat EA, Detrow MJ, Dickendesher TL, et al. Voltage-gated Na+ channel β1 subunit-mediated neurite outgrowth requires fyn kinase and contributes to central nervous system development in vivo. J Neurosci 2008; 28:3246 - 56; http://dx.doi.org/10.1523/JNEUROSCI.5446-07.2008; PMID: 18354028
- Lopez-Santiago LF, Pertin M, Morisod X, Chen C, Hong S, Wiley J, et al. Sodium channel beta2 subunits regulate tetrodotoxin-sensitive sodium channels in small dorsal root ganglion neurons and modulate the response to pain. J Neurosci 2006; 26:7984 - 94; http://dx.doi.org/10.1523/JNEUROSCI.2211-06.2006; PMID: 16870743
- Chen C, Westenbroek RE, Xu X, Edwards CA, Sorenson DR, Chen Y, et al. Mice lacking sodium channel beta1 subunits display defects in neuronal excitability, sodium channel expression, and nodal architecture. J Neurosci 2004; 24:4030 - 42; http://dx.doi.org/10.1523/JNEUROSCI.4139-03.2004; PMID: 15102918
- Kim DY, Carey BW, Wang H, Ingano LA, Binshtok AM, Wertz MH, et al. BACE1 regulates voltage-gated sodium channels and neuronal activity. Nat Cell Biol 2007; 9:755 - 64; http://dx.doi.org/10.1038/ncb1602; PMID: 17576410
- Adachi K, Toyota M, Sasaki Y, Yamashita T, Ishida S, Ohe-Toyota M, et al. Identification of SCN3B as a novel p53-inducible proapoptotic gene. Oncogene 2004; 23:7791 - 8; http://dx.doi.org/10.1038/sj.onc.1208067; PMID: 15334053
- Abriel H. Cardiac sodium channel Na(v)1.5 and interacting proteins: Physiology and pathophysiology. J Mol Cell Cardiol 2010; 48:2 - 11; http://dx.doi.org/10.1016/j.yjmcc.2009.08.025; PMID: 19744495
- Ding Y, Djamgoz MB. Serum concentration modifies amplitude and kinetics of voltage-gated Na+ current in the Mat-LyLu cell line of rat prostate cancer. Int J Biochem Cell Biol 2004; 36:1249 - 60; http://dx.doi.org/10.1016/j.biocel.2003.10.010; PMID: 15109569
- Ding Y, Brackenbury WJ, Onganer PU, Montano X, Porter LM, Bates LF, et al. Epidermal growth factor upregulates motility of Mat-LyLu rat prostate cancer cells partially via voltage-gated Na+ channel activity. J Cell Physiol 2008; 215:77 - 81; http://dx.doi.org/10.1002/jcp.21289; PMID: 17960590
- Brackenbury WJ, Djamgoz MB. Nerve growth factor enhances voltage-gated Na+ channel activity and Transwell migration in Mat-LyLu rat prostate cancer cell line. J Cell Physiol 2007; 210:602 - 8; http://dx.doi.org/10.1002/jcp.20846; PMID: 17149708
- Fraser SP, Ozerlat-Gunduz I, Onkal R, Diss JK, Latchman DS, Djamgoz MB. Estrogen and non-genomic upregulation of voltage-gated Na(+) channel activity in MDA-MB-231 human breast cancer cells: role in adhesion. J Cell Physiol 2010; 224:527 - 39; http://dx.doi.org/10.1002/jcp.22154; PMID: 20432453
- Chioni AM, Shao D, Grose R, Djamgoz MB. Protein kinase A and regulation of neonatal Nav1.5 expression in human breast cancer cells: activity-dependent positive feedback and cellular migration. Int J Biochem Cell Biol 2010; 42:346 - 58; http://dx.doi.org/10.1016/j.biocel.2009.11.021; PMID: 19948241
- Yildirim S, Altun S, Gumushan H, Patel A, Djamgoz MB. Voltage-gated sodium channel activity promotes prostate cancer metastasis in vivo. Cancer Lett 2012; 323:58 - 61; http://dx.doi.org/10.1016/j.canlet.2012.03.036; PMID: 22484465
- Biki B, Mascha E, Moriarty DC, Fitzpatrick JM, Sessler DI, Buggy DJ. Anesthetic technique for radical prostatectomy surgery affects cancer recurrence: a retrospective analysis. Anesthesiology 2008; 109:180 - 7; http://dx.doi.org/10.1097/ALN.0b013e31817f5b73; PMID: 18648226
- Walker AJ, Card T, Bates TE, Muir K. Tricyclic antidepressants and the incidence of certain cancers: a study using the GPRD. Br J Cancer 2011; 104:193 - 7; http://dx.doi.org/10.1038/sj.bjc.6605996; PMID: 21081933
- Yip D, Le MN, Chan JL, Lee JH, Mehnert JA, Yudd A, et al. A phase 0 trial of riluzole in patients with resectable stage III and IV melanoma. Clin Cancer Res 2009; 15:3896 - 902; http://dx.doi.org/10.1158/1078-0432.CCR-08-3303; PMID: 19458050
- Speyer CL, Smith JS, Banda M, DeVries JA, Mekani T, Gorski DH. Metabotropic glutamate receptor-1: a potential therapeutic target for the treatment of breast cancer. Breast Cancer Res Treat 2012; 132:565 - 73; http://dx.doi.org/10.1007/s10549-011-1624-x; PMID: 21681448
- Batcioglu K, Uyumlu AB, Satilmis B, Yildirim B, Yucel N, Demirtas H, et al. Oxidative Stress in the in vivo DMBA Rat Model of Breast Cancer: Suppression by a Voltage-gated Sodium Channel Inhibitor (RS100642). Basic Clin Pharmacol Toxicol 2012; 111:137 - 41; PMID: 22429688
- Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin 2011; 61:69 - 90; http://dx.doi.org/10.3322/caac.20107; PMID: 21296855
- Rugo HS. The importance of distant metastases in hormone-sensitive breast cancer. Breast 2008; 17:S3 - 8; http://dx.doi.org/10.1016/S0960-9776(08)70002-X; PMID: 18279764
- Suva LJ, Griffin RJ, Makhoul I. Mechanisms of bone metastases of breast cancer. Endocr Relat Cancer 2009; 16:703 - 13; http://dx.doi.org/10.1677/ERC-09-0012; PMID: 19443538
- Isom LL, De Jongh KS, Catterall WA. Auxiliary subunits of voltage-gated ion channels. Neuron 1994; 12:1183 - 94; http://dx.doi.org/10.1016/0896-6273(94)90436-7; PMID: 7516685
- Patino GA, Brackenbury WJ, Bao Y, Lopez-Santiago LF, O’Malley HA, Chen C, et al. Voltage-gated Na+ channel β1B: a secreted cell adhesion molecule involved in human epilepsy. J Neurosci 2011; 31:14577 - 91; http://dx.doi.org/10.1523/JNEUROSCI.0361-11.2011; PMID: 21994374
- Okuse K, Malik-Hall M, Baker MD, Poon WY, Kong H, Chao MV, et al. Annexin II light chain regulates sensory neuron-specific sodium channel expression. Nature 2002; 417:653 - 6; http://dx.doi.org/10.1038/nature00781; PMID: 12050667
- Cantrell AR, Smith RD, Goldin AL, Scheuer T, Catterall WA. Dopaminergic modulation of sodium current in hippocampal neurons via cAMP-dependent phosphorylation of specific sites in the sodium channel alpha subunit. J Neurosci 1997; 17:7330 - 8; PMID: 9295379
- Numann R, Catterall WA, Scheuer T. Functional modulation of brain sodium channels by protein kinase C phosphorylation. Science 1991; 254:115 - 8; http://dx.doi.org/10.1126/science.1656525; PMID: 1656525
- Lemaillet G, Walker B, Lambert S. Identification of a conserved ankyrin-binding motif in the family of sodium channel alpha subunits. J Biol Chem 2003; 278:27333 - 9; http://dx.doi.org/10.1074/jbc.M303327200; PMID: 12716895
- Wu L, Yong SL, Fan C, Ni Y, Yoo S, Zhang T, et al. Identification of a new co-factor, MOG1, required for the full function of cardiac sodium channel Nav 1.5. J Biol Chem 2008; 283:6968 - 78; http://dx.doi.org/10.1074/jbc.M709721200; PMID: 18184654
- Liu Cj, Dib-Hajj SD, Waxman SG. Fibroblast growth factor homologous factor 1B binds to the C terminus of the tetrodotoxin-resistant sodium channel rNav1.9a (NaN). J Biol Chem 2001; 276:18925 - 33; http://dx.doi.org/10.1074/jbc.M101606200; PMID: 11376006
- Deschênes I, Neyroud N, DiSilvestre D, Marbán E, Yue DT, Tomaselli GF. Isoform-specific modulation of voltage-gated Na(+) channels by calmodulin. Circ Res 2002; 90:E49 - 57; http://dx.doi.org/10.1161/01.RES.0000012502.92751.E6; PMID: 11884381
- van Bemmelen MX, Rougier JS, Gavillet B, Apothéloz F, Daidié D, Tateyama M, et al. Cardiac voltage-gated sodium channel Nav1.5 is regulated by Nedd4-2 mediated ubiquitination. Circ Res 2004; 95:284 - 91; http://dx.doi.org/10.1161/01.RES.0000136816.05109.89; PMID: 15217910
- Abriel H, Kamynina E, Horisberger JD, Staub O. Regulation of the cardiac voltage-gated Na+ channel (H1) by the ubiquitin-protein ligase Nedd4. FEBS Lett 2000; 466:377 - 80; http://dx.doi.org/10.1016/S0014-5793(00)01098-X; PMID: 10682864
- Gee SH, Madhavan R, Levinson SR, Caldwell JH, Sealock R, Froehner SC. Interaction of muscle and brain sodium channels with multiple members of the syntrophin family of dystrophin-associated proteins. J Neurosci 1998; 18:128 - 37; PMID: 9412493
- Gavillet B, Rougier JS, Domenighetti AA, Behar R, Boixel C, Ruchat P, et al. Cardiac sodium channel Nav1.5 is regulated by a multiprotein complex composed of syntrophins and dystrophin. Circ Res 2006; 99:407 - 14; http://dx.doi.org/10.1161/01.RES.0000237466.13252.5e; PMID: 16857961
- Srinivasan J, Schachner M, Catterall WA. Interaction of voltage-gated sodium channels with the extracellular matrix molecules tenascin-C and tenascin-R. Proc Natl Acad Sci USA 1998; 95:15753 - 7; http://dx.doi.org/10.1073/pnas.95.26.15753; PMID: 9861042
- Xiao ZC, Ragsdale DS, Malhotra JD, Mattei LN, Braun PE, Schachner M, et al. Tenascin-R is a functional modulator of sodium channel beta subunits. J Biol Chem 1999; 274:26511 - 7; http://dx.doi.org/10.1074/jbc.274.37.26511; PMID: 10473612
- Kazarinova-Noyes K, Malhotra JD, McEwen DP, Mattei LN, Berglund EO, Ranscht B, et al. Contactin associates with Na+ channels and increases their functional expression. J Neurosci 2001; 21:7517 - 25; PMID: 11567041
- Malhotra JD, Thyagarajan V, Chen C, Isom LL. Tyrosine-phosphorylated and nonphosphorylated sodium channel beta1 subunits are differentially localized in cardiac myocytes. J Biol Chem 2004; 279:40748 - 54; http://dx.doi.org/10.1074/jbc.M407243200; PMID: 15272007
- McEwen DP, Isom LL. Heterophilic interactions of sodium channel beta1 subunits with axonal and glial cell adhesion molecules. J Biol Chem 2004; 279:52744 - 52; http://dx.doi.org/10.1074/jbc.M405990200; PMID: 15466474
- Ratcliffe CF, Qu Y, McCormick KA, Tibbs VC, Dixon JE, Scheuer T, et al. A sodium channel signaling complex: modulation by associated receptor protein tyrosine phosphatase beta. Nat Neurosci 2000; 3:437 - 44; http://dx.doi.org/10.1038/74805; PMID: 10769382
- Merrick EC, Kalmar CL, Snyder SL, Cusdin FS, Yu EJ, Sando JJ, et al. The importance of serine 161 in the sodium channel beta3 subunit for modulation of Na(V)1.2 gating. Pflugers Arch 2010; 460:743 - 53; http://dx.doi.org/10.1007/s00424-009-0739-y; PMID: 19806359
- Wong HK, Sakurai T, Oyama F, Kaneko K, Wada K, Miyazaki H, et al. beta subunits of voltage-gated sodium channels are novel substrates of BACE1 and gamma -secretase. J Biol Chem 2005; 280:23009 - 17; http://dx.doi.org/10.1074/jbc.M414648200; PMID: 15824102
- Kim DY, Ingano LA, Carey BW, Pettingell WH, Kovacs DM. Presenilin/gamma-secretase-mediated cleavage of the voltage-gated sodium channel beta2-subunit regulates cell adhesion and migration. J Biol Chem 2005; 280:23251 - 61; http://dx.doi.org/10.1074/jbc.M412938200; PMID: 15833746
- Diss JK, Stewart D, Fraser SP, Black JA, Dib-Hajj S, Waxman SG, et al. Expression of skeletal muscle-type voltage-gated Na+ channel in rat and human prostate cancer cell lines. FEBS Lett 1998; 427:5 - 10; http://dx.doi.org/10.1016/S0014-5793(98)00378-0; PMID: 9613589
- Patino GA, Isom LL. Electrophysiology and beyond: multiple roles of Na+ channel β subunits in development and disease. Neurosci Lett 2010; 486:53 - 9; http://dx.doi.org/10.1016/j.neulet.2010.06.050; PMID: 20600605
- Malhotra JD, Kazen-Gillespie K, Hortsch M, Isom LL. Sodium channel β subunits mediate homophilic cell adhesion and recruit ankyrin to points of cell-cell contact. J Biol Chem 2000; 275:11383 - 8; http://dx.doi.org/10.1074/jbc.275.15.11383; PMID: 10753953