Abstract
Resveratrol (RSV, trans-3, 4, 5-Trihydroxystilbene), a type of polyphenol originally found in red wines, shows a great promise for the treatment of cancer, aging, type 2 diabetes, and cardiovascular diseases. Recent studies suggest that suppressing the signaling pathway mediated by mTOR, a well-known energy sensor that integrates various hormonal, nutrient, and environmental signals to regulate cell growth, metabolism and survival, could play an important role in mediating the beneficial effect of RSV. The underlying mechanisms by which RSV inhibits mTOR signaling remain elusive, but our recent studies show that RSV inhibits amino acid-stimulated mTOR signaling in C2C12 fibroblasts via a Sirt1- and AMPK-independent mechanism. RSV treatment has no effect on the expression levels of mTOR, raptor and DEPTOR, but greatly promotes the interaction between mTOR and its inhibitor DEPTOR. Our results reveal a novel mechanism by which RSV inhibits mTOR signaling and its function.
Resveratrol (RSV) is a natural-polyphenol compound that exerts multiple beneficial effects on cardiovascular, metabolic and immune functions.Citation1–Citation5 The mechanisms underlying the protective effects of RSV on various cardiovascular and metabolic disorders remain elusive, but there is some evidence suggesting that inhibition of the mammalian target of rapamycin (mTOR) signaling pathway could play a role.Citation1–Citation3
mTOR is a member of the PI 3-kinase-related protein kinase (PIKK) family that plays a critical role in the regulation of cell homeostasis in response to various upstream stimuli such as growth factors, nutrients and stress.Citation6,Citation7 mTOR exists in two distinct complexes, TORC1 and TORC2, which differ in subunit compositions and biological functions.Citation8 The rapamycin-sensitive mTORC1, which consists of five components including mTOR, the regulatory-associated protein of mTOR (Raptor), mammalian lethal with Sec13 protein 8 (mLST8, also known as GβL), proline-rich Akt substrate 40 kDa (PRAS40), and the DEP-domain-containing and mTOR-interactive protein (DEPTOR),Citation9 regulates protein synthesis and cell growth by phosphorylating downstream target proteins such as p70 ribosomal S6 kinase 1 (S6K) and the eukaryotic initiation factor 4E-BP1.Citation6,Citation10 The rapamycin-insensitive TORC2 contains similar components except that Raptor is replaced with a unique protein named rapamycin-insensitive companion of mTOR (Rictor).
The mechanism by which RSV inhibits mTOR signaling remains elusive. Some studies suggest that activation of the Sirt1 signaling pathway is essential for RSV action.Citation11–Citation13 However, a number of recent studies indicate that many of the protective effects of RSV could be mediated by Sirt1-independent mechanisms.Citation14–Citation16 In agreement with this, our recent study showed that RSV inhibits insulin- and leucine-stimulated mTOR signaling in a Sirt1-independent manner.Citation17
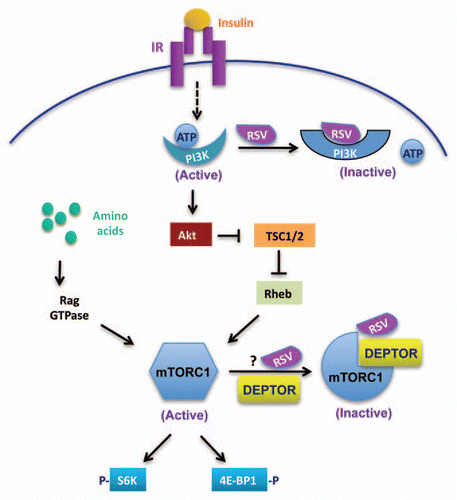
RSV has been shown to directly bind to the ATP-binding site of PI3K and thus function as a class 1A PI3K inhibitor.Citation16,Citation18 Since activation of the PI3K/PI3K-dependent kinase 1 (PDK1)/Akt signaling pathway is necessary for insulin-stimulated mTOR activation,Citation19,Citation20 it is possible that RSV inhibits mTOR signaling by inhibiting the PI3K/PDK1/Akt signaling pathway. Consistent with this, we found that treating C2C12 fibroblasts with RSV greatly inhibited insulin-stimulated activation of the PI3K and the mTOR signaling pathways.Citation17 However, inhibition of Akt or disruption of PDK1 expression had no significant effect on leucine-stimulated mTOR signaling, suggesting that the mechanisms by which insulin and amino acids promote mTOR activation are distinct.Citation17
The tumor suppressor tuberous sclerosis complex (TSC1/2) plays a crucial role in the negative regulation of Rheb, a small GTPase that activates mTOR.Citation21,Citation22 While it is possible that RSV may inhibit mTOR signaling by targeting TSC1/2, we found that suppression or disruption of TSC1/2 expression had little effect on leucine-stimulated mTOR signaling in C2C12 cells or MEFs,Citation17 suggesting that RSV negatively regulates mTOR signaling by targeting at a site downstream of TSC1/2. Consistent with this view, we found that RSV treatment markedly increased the association between mTOR and DEPTOR, a newly identified negative regulator of mTOR.Citation23,Citation24 Since the binding of DEPTOR to mTOR is critical for the negative regulation of mTOR activity,Citation23,Citation24 enhancing the interaction between DEPTOR and mTOR could thus provide a mechanism by which RSV inhibits mTORC1 signaling. In agreement with this view, suppressing the expression levels of DEPTOR by RNAi greatly diminished the inhibitory effect of RSV on leucine-stimulated mTORC1 signaling.Citation17 Our study also showed that RSV treatment negatively regulates Akt activity.Citation17 Since Akt stimulates mTOR activity by suppressing the negative effect of TSC1/2,Citation25 these findings indicate that RSV is able to negatively regulate mTOR activity via distinct mechanisms in response to different upstream stimuli.
In vitro kinase assay experiments strongly suggest that RSV may suppress PI3K activity through a direct binding to the ATP-binding site of PI3K ().Citation18 The 4′ hydroxyl group of RSV has been shown to form a hydrogen bond with the catalytic lysine residues in the ATP-binding site of p110α (or p110β), thus leading to the suppression of PI3K activity.Citation16,Citation18 mTOR is a class IV PI3K protein kinase,Citation26,Citation27 and its C-terminal catalytic domain is homologous to the p110α catalytic subunit of the class I PI3K.Citation28 The structural similarity between mTOR and PI3K has promoted the identification of highly potent and effective PI3K/mTOR dual inhibitors for the treatment of cancer.Citation29–Citation31 An interesting question that remains unanswered is whether RSV also targets the ATP-binding pocket of mTOR and if it does, whether the binding provides a mechanism underlying the negative effect of RSV on mTOR activity ().
DEPTOR has been shown to interact with the C-terminal portion of mTOR (aa 1,483–2,000) that is upstream of its kinase domain (aa 2,430–2,549) including the ATP-binding site (aa 2,519–2,549).Citation32 It is possible that the interaction between RSV and mTOR leads to a conformational change that promotes the interaction between mTOR and DEPTOR. Another possibility is that RSV may interact with DEPTOR and the formation of the RSV/DEPTOR complex is essential for interaction with mTOR. Further studies will be necessary to address these possibilities. Nevertheless, our finding that RSV promotes the interaction between DEPTOR and mTOR reveals a novel mechanism by which RSV negatively regulates mTOR signaling and function.
Figures and Tables
Figure 1 Mechanisms by which RSV inhibits insulin and amino acid-induced mTOR activation. (1) RSV binds to the ATP-binding site of the p110 subunit of PI3K and competitively inhibits the binding of ATP to the enzyme, leading to suppression of mTOR activity and downstream signaling. (2) RSV promotes the interaction between mTOR and its inhibitor DEPTOR, resulting in an inhibition of mTOR signaling.
Acknowledgments
This work is supported by National Institutes of Health Grants RO1 DK76902 (to F.L.).
References
- Baur JA. Resveratrol, sirtuins, and the promise of a DR mimetic. Mech Ageing Dev 2010; 131:261 - 269
- Lagouge M, Argmann C, Gerhart-Hines Z, Meziane H, Lerin C, Daussin F, et al. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1alpha. Cell 2006; 127:1109 - 1122
- Chen H, Yang Z, Gibbs D, Yang X, Hau V, Zhao P, et al. Association of HTRA1 polymorphism and bilaterality in advanced age-related macular degeneration. Vision Res 2008; 48:690 - 694
- Acharya SH, Philip S, Viswanath AK, Boroujerdi M, Waugh NR, Pearson DW. Glycaemic control and body mass index in late-adolescents and young adults with Type 1 diabetes mellitus: a population-based study. Diabet Med 2008; 25:360 - 364
- Baur JA, Pearson KJ, Price NL, Jamieson HA, Lerin C, Kalra A, et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006; 444:337 - 342
- Wullschleger S, Loewith R, Hall MN. TOR signaling in growth and metabolism. Cell 2006; 124:471 - 484
- Yang Q, Guan KL. Expanding mTOR signaling. Cell Res 2007; 17:666 - 681
- Loewith R, Jacinto E, Wullschleger S, Lorberg A, Crespo JL, Bonenfant D, et al. Two TOR complexes, only one of which is rapamycin sensitive, have distinct roles in cell growth control. Mol Cell 2002; 10:457 - 468
- Laplante M, Sabatini DM. mTOR signaling at a glance. J Cell Sci 2009; 122:3589 - 3594
- Guertin DA, Sabatini DM. Defining the role of mTOR in cancer. Cancer Cell 2007; 12:9 - 22
- Wood JG, Rogina B, Lavu S, Howitz K, Helfand SL, Tatar M, Sinclair D. Sirtuin activators mimic caloric restriction and delay ageing in metazoans. Nature 2004; 430:686 - 689
- Parker JA, Arango M, Abderrahmane S, Lambert E, Tourette C, Catoire H, Neri C. Resveratrol rescues mutant polyglutamine cytotoxicity in nematode and mammalian neurons. Nat Genet 2005; 37:349 - 350
- Han YS, Zheng WH, Bastianetto S, Chabot JG, Quirion R. Neuroprotective effects of resveratrol against beta-amyloid-induced neurotoxicity in rat hippocampal neurons: involvement of protein kinase C. Br J Pharmacol 2004; 141:997 - 1005
- Um JH, Park SJ, Kang H, Yang S, Foretz M, McBurney MW, et al. AMP-activated protein kinase-deficient mice are resistant to the metabolic effects of resveratrol. Diabetes 2010; 59:554 - 563
- Chan AY, Dolinsky VW, Soltys CL, Viollet B, Baksh S, Light PE, Dyck JR. Resveratrol inhibits cardiac hypertrophy via AMP-activated protein kinase and Akt. J Biol Chem 2008; 283:24194 - 24201
- Jiang H, Shang X, Wu H, Gautam SC, Al-Holou S, Li C, et al. Resveratrol downregulates PI3K/Akt/mTOR signaling pathways in human U251 glioma cells. Resveratrol downregulates PI3K/Akt/mTOR signaling pathways in human U251 glioma cells. J Exp Ther Oncol 2009; 8:25 - 33
- Liu M, Wilk SA, Wang A, Zhou L, Wang RH, Ogawa W, et al. Resveratrol inhibits mTOR signaling by promoting the interaction between mTOR and DEPTOR. J Biol Chem 2010; 285:36387 - 36394
- Frojdo S, Cozzone D, Vidal H, Pirola L. Resveratrol is a class IA phosphoinositide-3-kinase inhibitor. Biochem J 2007; 406:511 - 518
- Scott PH, Brunn GJ, Kohn AD, Roth RA, Lawrence JC Jr. Evidence of insulin-stimulated phosphorylation and activation of the mammalian target of rapamycin mediated by a protein kinase B signaling pathway. Proc Natl Acad Sci USA 1998; 95:7772 - 7777
- Taha C, Liu Z, Jin J, Al-Hasani H, Sonenberg N, Klip A. Opposite translational control of GLUT1 and GLUT4 glucose transporter mRNAs in response to insulin. Role of mammalian target of rapamycin, protein kinase b and phosphatidylinositol-3-kinase in GLUT1 mRNA translation. J Biol Chem 1999; 274:33085 - 33091
- Garami A, Zwartkruis FJ, Nobukuni T, Joaquin M, Roccio M, Stocker H, et al. Insulin activation of Rheb, a mediator of mTOR/S6K/4E-BP signaling, is inhibited by TSC1 and 2. Mol Cell 2003; 11:1457 - 1466
- Saucedo LJ, Gao X, Chiarelli DA, Li L, Pan D, Edgar BA. Rheb promotes cell growth as a component of the insulin/TOR signalling network. Nat Cell Biol 2003; 5:566 - 571
- Proud CG. Dynamic balancing: DEPTOR tips the scales. J Mol Cell Biol 2009; 1:61 - 63
- Peterson TR, Laplante M, Thoreen CC, Sancak Y, Kang SA, Kuehl WM, et al. DEPTOR is an mTOR inhibitor frequently overexpressed in multiple myeloma cells and required for their survival. Cell 2009; 137:873 - 886
- Harrington LS, Findlay GM, Gray A, Tolkacheva T, Wigfield S, Rebholz H, et al. The TSC1-2 tumor suppressor controls insulin-PI3K signaling via regulation of IRS proteins. J Cell Biol 2004; 166:213 - 223
- Vivanco I, Sawyers CL. The phosphatidylinositol-3-Kinase AKT pathway in human cancer. Nat Rev Cancer 2002; 2:489 - 501
- Strimpakos AS, Karapanagiotou EM, Saif MW, Syrigos KN. The role of mTOR in the management of solid tumors: an overview. Cancer Treat Rev 2009; 35:148 - 159
- Harris TE, Lawrence JC Jr. TOR signaling. Sci STKE 2003; 15
- Serra V, Markman B, Scaltriti M, Eichhorn PJ, Valero V, Guzman M, et al. NVP-BEZ235, a dual PI3K/ mTOR inhibitor, prevents PI3K signaling and inhibits the growth of cancer cells with activating PI3K mutations. Cancer Res 2008; 68:8022 - 8030
- Knight ZA, Gonzalez B, Feldman ME, Zunder ER, Goldenberg DD, Williams O, et al. A pharmacological map of the PI3-K family defines a role for p110alpha in insulin signaling. Cell 2006; 125:733 - 747
- Yap TA, Garrett MD, Walton MI, Raynaud F, de Bono JS, Workman P. Targeting the PI3K-AKT-mTOR pathway: progress, pitfalls and promises. Curr Opin Pharmacol 2008; 8:393 - 412
- Lempiainen H, Halazonetis TD. Emerging common themes in regulation of PIKKs and PI3Ks. EMBO J 2009; 28:3067 - 3073