Abstract
Despite a long history of anatomical mapping of neuronal networks, we are only beginning to understand the detailed three-dimensional (3D) organization of the cortical micro-circuitry. This is in part due to the lack of complete reconstructions of individual cortical neurons. Morphological studies are either performed on incomplete cells in vitro, or when performed in vivo, lack the necessary cellular resolution. We recently reconstructed the in vivo axonal and dendritic morphology of two types of L(ayer) 5 neurons from vibrissal cortex. The 3D profiles of short-range as well as long-range projections indicate that L5 slender-tufted and L5 thick-tufted neurons represent very different building blocks of the cortical circuitry. In this addendum to Oberlaender et al, PNAS 2011, we motivate our technical approach and the advancements this may give in reconstructing the cortical micro-circuitry.
Due to its well-defined columnar and laminar organization, the vibrissal cortex in rodents is a highly suitable model to study cortical circuits.Citation1–Citation3 In particular, the segregation into column and septum related regions of the vibrissal cortex allows one to align and compare 3D morphologies of neurons from different animals. In our in vivo approach, the individual neuron reconstructions are therefore always performed in the context of a common reference frame containing three anatomical landmarks: (1) the barrel contours in granular layer 4, (2) the pia surface and (3) the white matter. Biological variability in brain size or dimensions of the vibrissal area can then be accounted for by linear remodelling of the common reference frame.
Such reconstructions revealed that 3D axonal morphologies in vivoCitation4,Citation5 can be an order of magnitude larger compared to previously reported values obtained in brain slices.Citation6,Citation7 This could be due to limited visibility of axons in 300–400 µm thick slices,Citation8 or more plausible, axonal structures in in vitro preparations represent not more than 10% of the possible axonal projections present in vivo. The vast innervation volumes of in vivo filled axons also imply that manual reconstructions are highly labor intensive. This makes it virtually impossible to get the high throughput required to reconstruct representative amounts of all the different neuronal cell types found in the cortexCitation9 and thus asks for more automated approaches.Citation10 These issues aside, perhaps the most important problem of manual reconstructions is that intricate morphologies makes the chance for human error very likely, even for the very skilled human tracer. This motivated us to develop a semi-automated reconstruction pipeline.Citation11–Citation13 This method allows algorithm-based (and therefore reliable) reconstructions and reduces manual labor for individual reconstructions from ∼90 hours to 8–10 hours (the manual labor involves splicing of serial sections and not reconstruction). In short, this technique allows highly accurate quantitative axonal reconstructions that can be achieved in relatively little time.
The first quantitative study we performed using this technique was to reconstruct the axonal projection patterns of two types of layer 5 pyramidal neurons filled with biocytin in vivo ().Citation4 These slender-and thick-tufted neurons can be classified based on their dendritic morphology and respond differently to passive whisker touch or active whisker movements.Citation14,Citation15 We found that axonal projections of slender-tufted and thick-tufted neurons target different layers and thus represent functionally and anatomically distinct units of the cortical micro-circuitry. L5 slender-tufted neurons displayed wide-spreading axonal projections (86.8 ± 5.5 mm), which primarily innervated supragranular layers of the entire vibrissal cortex and higher order cortices (dysgranular zone, posterior parietal cortex). L5 thick-tufted neurons in turn, are characterized by shorter and less complex axonal projections (31.6 ± 14.3 mm), which primarily innervated nearby infragranular layers. These results indicate that the use of semi-automated reconstructions of axonal projection profiles provides detailed new insights into the putative postsynaptic targets of individual neurons. Furthermore, full 3D axonal reconstructions are a crucial step in generating hypotheses about the pathways of cortical information processing. There is virtually no limit to the semi-automated reconstruction pipeline and it may be used to reconstruct neurons from any brain area of choice.
Ultimately, the combination of anatomical reconstructions and electrophysiological recordings of the activity of individual neurons during different behavioral states will generate insight into the functions of different cell types and facilitate the reconstruction of cortical circuits. These experimental approaches will pave the way to conduct simulations of anatomically realistic networks and may lay the foundation for future studies on brain degenerative diseases, such as Alzheimer's. This research will thus not only help to advance our basic understanding of cortical circuits, but may eventually provide a starting point to study the influences of brain diseases on neuronal morphology and function.
Figures and Tables
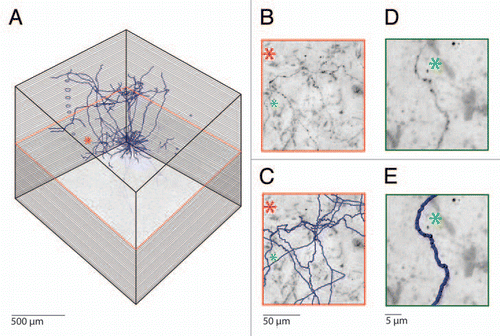
Figure 1 Semi-automated reconstruction of a Layer 5 pyramidal neuron filled with biocytin in vivo. (A) Mosaic scanning and subsequent serial reconstruction is performed on consecutive 100 µm thick tangential sections, which results in high resolution 3D images representing cubic millimeters of cortical volumes. The consecutive slices are aligned by using blood vessels that run perpendicular to the cortical surface. (B) Magnification of the area indicated by the asterisk. Note the abundant axons running through this area. (C) Automated detection of biocytin labeled processes allows fast and reliable reconstruction of axonal morphology. Axonal reconstructions from additional Z values are also visible in this part. (D) Example images of a single axonal branch labeled with biocytin and (E) the subsequent automated reconstruction.
Acknowledgments
This work was supported by the Max-Planck Society, the Center for Neurogenomics and Cognitive Research at Vrije Universiteit Amsterdam, and a Veni Grant from The Netherlands Organization for Scientific Research (to C.P.J.d.K.).
Addendum to:
References
- Helmstaedter M, de Kock CP, Feldmeyer D, Bruno RM, Sakmann B. Reconstruction of an average cortical column in silico. Brain Res Rev 2007; 55:193 - 203
- Markram H. The blue brain project. Nat Rev Neurosci 2006; 7:153 - 160
- O'Connor DH, Huber D, Svoboda K. Reverse engineering the mouse brain. Nature 2009; 461:923 - 929
- Oberlaender M, Boudewijns ZSRM, Kleele T, Mansvelder HD, Sakmann B,, de Kock CPJ. Three-dimensional axon morphologies of individual layer 5 neurons indicate cell type-specific intracortical pathways for whisker motion and touch. Proc Natl Acad Sci USA 2011; 108:4188 - 4193
- Bruno RM, Hahn TT, Wallace DJ, de Kock CP, Sakmann B. Sensory experience alters specific branches of individual corticocortical axons during development. J Neurosci 2009; 29:3172 - 3181
- Schubert D, Kotter R, Luhmann HJ, Staiger JF. Morphology, electrophysiology and functional input connectivity of pyramidal neurons characterizes a genuine layer Va in the primary somatosensory cortex. Cereb Cortex 2006; 16:223 - 236
- Lubke J, Roth A, Feldmeyer D, Sakmann B. Morphometric analysis of the columnar innervation domain of neurons connecting layer 4 and layer 2/3 of juvenile rat barrel cortex. Cereb Cortex 2003; 13:1051 - 1063
- Romand S, Wang Y, Toledo-Rodriguez M, Markram H. Morphological development of thick-tufted layer V pyramidal cells in the rat somatosensory cortex. Front Neuroanat 2011; 5:5
- Svoboda K. The past, present and future of single neuron reconstruction. Neuroinformatics 2011; 9:97 - 98
- Gillette TA, Brown KM, Svoboda K, Liu Y, Ascoli GA. DIADEMchallenge.Org: a compendium of resources fostering the continuous development of automated neuronal reconstruction. Neuroinformatics 2011; 9:303 - 304
- Oberlaender M, Bruno RM, Sakmann B, Broser PJ. Transmitted light brightfield mosaic microscopy for three-dimensional tracing of single neuron morphology. J Biomed Opt 2007; 12:064029
- Dercksen VJ, Oberlaender M, Sakmann B, Hege HC. Interactive visualization—a key prerequisite for reconstruction of anatomically realistic neural network. Proceedings of the 2009 Workshop on Visualization in Medicine and Life Sciences Springer Verlag 2010;
- Dercksen VJ, Weber B, Guenther D, Oberlaender M, Prohaska S, Hege HC. Automatic alignment of stacks of filament data. IEEE International Symposium on Biomedical Imaging: From Nano to Macro (ISBI) 2009; 971 - 974
- de Kock CP, Bruno RM, Spors H, Sakmann B. Layer- and cell-type-specific suprathreshold stimulus representation in rat primary somatosensory cortex. J Physiol 2007; 581:139 - 154
- de Kock CP, Sakmann B. Spiking in primary somatosensory cortex during natural whisking behavior in the awake rat. Proc Natl Acad Sci USA 2009; 106:16446 - 16450