Abstract
The final abscission event of cytokinesis is necessary for daughter cells to part ways from one another. Failure to properly divide has been indicated as a potential cancer initiating event due to an increase in cellular aneuploidy. However, the exact mechanisms of abscission have remained obscured by our inability to properly discern the spatiotemporal regulation of the various proteins and organelles required for cytokinesis. Three recent publications have taken slightly varied high resolution imaging approaches to visualize cytokinesis and abscission. As a result of this work, two differing, but not necessarily mutually exclusive, models have emerged. One model is ESCRT-centric and the other, recycling endosome-dependent, each describing the steps leading up to the final abscission event. Presently these models describe late cytokinesis events leading to abscission in greater detail than previously known.
As a single cell divides, the resulting daughter cells must separate from one another during the final step of cytokinesis. This abscission event occurs on either or both sides of the midbody within the intracellular bridge (ICB). While earlier studies thought this abscission event was simply a continuation of the constriction placed on the cleavage furrow by the actomyosin contractile ring, it was later discovered that abscission is a highly complex and organized event consisting of much more than a simple actin and non-muscle myosin constricting ring.Citation1 Vesicular trafficking, protein complex function and cytoskeletal reorganization were all shown to contribute to cytokinesis and abscission.Citation1,Citation2 Recent studies have established a requirement for recycling endosome delivery to the intracellular bridge during cytokinesis.Citation3,Citation4 Faulty recruitment and delivery of these recycling endosomes caused a failure in cytokinesis and abscission.Citation3,Citation4 Additionally, the ESCRT (endosomal sorting complex required for transport) complex was identified to be necessary for abscission.Citation5 The ESCRT complex function was hypothesized to be similar to its topologically equivalent scission roles in multi-vesicular body (MVB) biogenesis and enveloped viral budding. Due to a recent string of publications,Citation2,Citation6,Citation7 our knowledge of the final moments during cytokinesis and abscission is becoming clearer. We have shown that Rab11 effector protein, FIP3-positive endosome fusion is required for the generation of a secondary ingression site, prior to abscission.Citation2 Furthermore, two different labsCitation6,Citation7 have further clarified the role of the ESCRT complex function contributing to abscission. Here we discuss these recent publications and their implications in the field of cytokinesis.
Recycling Endosome Delivery is a Prerequisite for Cytokinesis
The delivery of recycling endosomes in many distinct organisms has been shown to play a role in cytokinesis and abscission. The family of monomeric Rab GTPases are known regulators of vesicle trafficking and two Rab proteins in particular, Rab11 and Rab35, have been demonstrated to function during cytokinesis.Citation8 When Rab11 and its effector protein FIP3,Citation4 have been depleted from mammalian cells, defects in cytokinesis and abscission became apparent, and led to an increase in the number of multi-nucleated cells. Observations by several laboratories have shown that the plasma membrane is highly dynamic during the later stages of cytokinesis,Citation2,Citation9 presumably as the result of FIP3-positive endosome fusion with the plasma membrane of the ICB. Furthermore, our recent work combining time-lapse microscopy and correlative high-resolution 3D tomography have demonstrated that localized microtubule severing and fusion of FIP3-endosomes within the ICB is required for, and leads to, the formation of a secondary ingression.Citation2 This secondary ingression results in the thinning of the ICB to a width of 100–200 nm and appears to be required for successful completion of abscission. What remains unclear is how FIP3-endosomes mediate the initiation and expansion of the secondary ingression.
Post-Golgi secretory vesicles also are known to traffic to the ICB. It was suggested that secretory vesicles may be required for cytokinesis,Citation10 as treatment with Brefeldin A inhibits cytokinesis is some organisms. However, in other organisms, the inhibition of secretory vesicle trafficking has no effect on cytokinesis.Citation10 Presently the exact function of secretory vesicles during cytokinesis of mammalian cells remains unclear, but it is unlikely that they play a key role in the abscission event. As we demonstrated in tomographic reconstructions of the ICBs of HeLa cells during early and late cytokinesis, spherical vesicles, likely to be secretory vesicles, are present in the ICB during early telophase. Interestingly, these vesicles could no longer be seen inside the ICB during late telophase, when FIP3-endosomes and lytic endosomes are now the dominant population of organelles inside the ICB.Citation2
The Requirement of the ESCRT Complex during Abscission
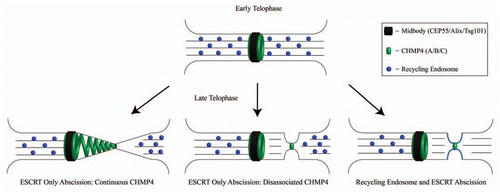
The ESCRT complex has well known roles in MVB biogenesis. However, it was not until Carlton and Martin-SerranoCitation5 found two proteins involved in HIV-1 budding, tumor susceptibility gene 101 (Tsg101) and Alix, that ESCRT complex proteins were demonstrated to be required for cytokinesis. The localization of Tsg101 and Alix requires the presence of CEP55, a centrosomal and midbody protein, yet the necessary ESCRT complex members involved during cytokinesis are unique from their other functions within the cell. Similarly, an ESCRT-III homologue from Archaea of the genus Sulpholobus is cell cycle regulated, positioned between segregating nucleoids of the daughter cells, and is required for cell division.Citation11,Citation12 The model of ESCRT recruitment to the midbody by CEP55 is as follows: Alix, an ESCRT complex accessory protein and Tsg101, an ESCRT-I member, are recruited to the midbody by binding CEP55. These components then recruit various ESCRT-III members culminating in the recruitment of CHMP4B to the midbody. CHMP4B has the ability to form ∼5 nm filaments in vitroCitation13 and can form multi-filament polymers which can appear as spiral shaped conformations.Citation13 These studies have served as the basis for the ESCRT abscission model, with membrane constriction and abscission occurring via polymerizing CHMP4B spirals ().
Recently, two papers have provided further evidence in support of the ESCRT model of abscission. Guizetti and colleagues noted the appearance of 17 nm filaments within the ICB during late cytokinesis by performing high-resolution tomography. Based on these data it was proposed that CHMP4 is recruited to the midbody, where it polymerizes to form continuous spiral filaments spanning from the midbody to the secondary ingression site ( Continuous CHMP4 Model). Despite all of these discoveries, a few questions remain unanswered. The 17 nm filaments shown in the tomographic reconstructions were alluded to be CHMP4B filaments within the ICB. Unfortunately, the authors did not perform immunoelectron microscopy on these 17 nm filaments. As the result, it remains to be shown whether these filaments actually represent ESCRT-III. An interesting alternative could be that these filaments are actually composed of septins. Septin filaments form multi-filament structures from their single filament base diameter of 8 nm,Citation14 and could also form the visible ∼17 nm filaments. Septins have been demonstrated to form filamentous structures within the budneck of yeasts,Citation14 and are required for cytokinesis in both yeast and mammalian cells.Citation14
The involvement of the ESCRT-III complex in the abscission event was also confirmed by a study by Elia and colleagues, which also used structural illumination microscopy to analyze the spatiotemporal changes in CHMP4B dynamics during late telophase.Citation6 Interestingly, while this study clearly demonstrated the accumulation of CHMP4B at the ICB during late telophase, a continuous distribution of CHMP4B filaments from the midbody to the secondary ingression and abscission site was not observed.Citation6 While one possibility is that these filaments could not be detected due to the low expression levels of tagged CHMP4B, the authors also speculate that the midbody may be a “staging area” for CHMP4B activation, with the activated CHMP4B then diffusing to the site of abscission ( Disassociated CHMP4 Model). What remain unclear are the mechanisms that determine the location of the abscission site, and how the activated CHMP4B is targeted away from the midbody to the forming secondary ingression.
The Cross-Talk between the ESCRT Complex and FIP3-Endosomes
What is the relationship between the roles of the ESCRT complex and endosomes during abscission? Perhaps the two proposed cytokinesis models, the ESCRT abscission model and the endosome-dependent secondary ingression model, are not mutually exclusive ( Recycling Endosome and ESCRT Abscission Model). One intriguing possibility is that CHMP4B is first recruited and activated at the midbody “staging area.” Upon narrowing the ICB to 100–200 nm through microtubule severing and endosome fusion-induced secondary ingression, CHMP4B is targeted to the secondary ingression to complete abscission. While MVB biogenesis, enveloped viral budding, and cytokinesis are all topologically similar events, the plasma membrane necks of MVB biogenesis and enveloped viral budding are on a different scale compared to the ICB during cytokinesis, which is approximately 20 times larger. Can CHMP4B filaments physically handle the actin and microtubule rich 2–3 µm thick ICB during cytokinesis, compared to the 50–100 nm vesicle necks of MVB biogenesis and enveloped viral budding? Perhaps the formation of the secondary ingression is needed to “cut-down” the size of the ICB to a more manageable 100 nm. This thinning of the ICB may also be a trigger allowing the recruitment of activated CHMP4B to the abscission site. Finally, fusion of the endosomes with the ICB's plasma membrane may also deliver proteins and/or lipids to modify the abscission site and allow CHMP4B filament formation. Furrow-enriched recycling endosomes have already been implicated in the delivery of PI(3)P phospholipids as well as regulatory proteins, such as BRUCE and FYVE-CENT, to the ICB during late telophase.Citation15,Citation16 Thus, while a combined model of endosomes and the ESCRT complex roles during cytokinesis is beginning to emerge, more studies are needed to decipher the mechanisms of the abscission step of cytokinesis.
Figures and Tables
Figure 1 ESCRT-dependent and endosome dependent models of cytokinesis and abscission. (Top row) ICB during early telophase has been hypothesized to undergo abscission by the three models shown in the bottom row (Bottom Left and Center- ESCRT-dependent abscission models, Bottom right- ESCRT and endosome dependent abscission model).
Acknowledgments
We are grateful to Carly Willenborg (University of Colorado Denver) and Dr. Gwyn Gould (University of Glasgow) for critical reading of the manuscript. Work in Dr. Prekeris' laboratory has been funded by the National Institute of Health, DK064380 and Susan G. Komen Breast Cancer Research Foundation.
Addendum to:
References
- Eggert US, Mitchison TJ, Field CM. Animal cytokinesis: from parts list to mechanisms. Annu Rev Biochem 2006; 75:543 - 566
- Schiel JA, Park K, Morphew MK, Reid E, Hoenger A, Prekeris R. Endocytic membrane fusion and buckling-induced microtubule severing mediate cell abscission. J Cell Sci 2011; 124:1411 - 1424
- Fielding AB, Schonteich E, Matheson J, Wilson G, Yu X, Hickson GR, et al. Rab11-FIP3 and FIP4 interact with Arf6 and the exocyst to control membrane traffic in cytokinesis. EMBO J 2005; 24:3389 - 3399
- Wilson GM, Fielding AB, Simon GC, Yu X, Andrews PD, Hames RS, et al. The FIP3-Rab11 protein complex regulates recycling endosome targeting to the cleavage furrow during late cytokinesis. Mol Biol Cell 2005; 16:849 - 860
- Carlton JG, Martin-Serrano J. Parallels between cytokinesis and retroviral budding: a role for the ESCRT machinery. Science 2007; 316:1908 - 1912
- Elia N, Sougrat R, Spurlin TA, Hurley JH, Lippincott-Schwartz J. Dynamics of endosomal sorting complex required for transport (ESCRT) machinery during cytokinesis and its role in abscission. Proc Natl Acad Sci USA 2011; 108:4846 - 4851
- Guizetti J, Schermelleh L, Mantler J, Maar S, Poser I, Leonhardt H, et al. Cortical constriction during abscission involves helices of ESCRT-III-dependent filaments. Science 2011; 331:1616 - 1620
- Schiel JA, Prekeris R. Making the final cut—mechanisms mediating the abscission step of cytokinesis. Sci World J 2010; 10:1424 - 1434
- Byers B, Abramson DH. Cytokinesis in HeLa: post-telophase delay and microtubule-associated motility. Protoplasma 1968; 66:413 - 435
- Baluska F, Menzel D, Barlow PW. Cytokinesis in plant and animal cells: endosomes ‘shut the door’. Dev Biol 2006; 294:1 - 10
- Lindas AC, Karlsson EA, Lindgren MT, Ettema TJ, Bernander R. A unique cell division machinery in the Archaea. Proc Natl Acad Sci USA 2008; 105:18942 - 18946
- Samson RY, Obita T, Freund SM, Williams RL, Bell SD. A role for the ESCRT system in cell division in archaea. Science 2008; 322:1710 - 1713
- Hanson PI, Roth R, Lin Y, Heuser JE. Plasma membrane deformation by circular arrays of ESCRT-III protein filaments. J Cell Biol 2008; 180:389 - 402
- Cao L, Yu W, Wu Y, Yu L. The evolution, complex structures and function of septin proteins. Cell Mol Life Sci 2009; 66:3309 - 3323
- Pohl C, Jentsch S. Final stages of cytokinesis and midbody ring formation are controlled by BRUCE. Cell 2008; 132:832 - 845
- Sagona AP, Nezis IP, Pedersen NM, Liestol K, Poulton J, Rusten TE, et al. PtdIns(3)P controls cytokinesis through KIF13A-mediated recruitment of FYVE-CENT to the midbody. Nat Cell Biol 2010; 12:362 - 371