Abstract
The Golgi apparatus is the central protein sorting station inside eukaryotic cells. Although many regulators of Golgi trafficking have been identified, little is known about their crosstalk. Both the Arf activation cycle and phosphatidylinositol 4-phosphate metabolism have been recognized as key processes in the regulation of vesicular transport from this organelle. However, the mechanism ensuring the proper co-regulation of these processes has eluded our understanding thus far. We recently identified a physical interaction between the late yeast Golgi Arf activator Sec7p and the PI4-kinase Pik1p, and showed that the two proteins cooperate in the formation of clathrin-coated vesicles. This finding gives the first insight on the coordinated generation of a dual key signal by a small GTPase and a signaling phospholipid at the Golgi. In addition, it opens new perspectives for a better understanding of Golgi maturation through coordinated regulation of highly dynamic lipid and protein composition of this organelle.
The Golgi apparatus is the main crossroad of intracellular traffic events in eukaryotic cells. This central position confers a crucial role to this organelle in maintaining proper cellular organization and provides an important template for the integration of regulatory signals. Hence, besides its central role in protein sorting, the Golgi is also an important site for protein and lipid modification as well as a platform for several signal transduction pathways.Citation1-Citation3 To perform these functions, the Golgi apparatus is composed of a succession of sub-compartments or cisternae recognizable by their biochemical content.Citation4,Citation5 In most cells, Golgi cisternae are organized as a series of flattened stacks, even though this structure is not universal. One of the most noticeable exceptions is the budding yeast S. cerevisiae, where the Golgi apparatus is composed of scattered and mobile cisternae able to maintain their sub-compartmentalization despite their discrete spatial localization.Citation6 In general, newly synthesized proteins transported from the ER to the Golgi enter this organelle on the cis-side and progress through the different compartments before leaving the organelle from the trans-Golgi network. How this progression is achieved without altering the overall structure of the Golgi is still a matter of debate. On one hand, the cisternal maturation model considers single Golgi stacks as dynamic entities changing with time, and limits the role of vesicular transport to the recycling of resident proteins to earlier compartments. On the other hand, in the vesicular transport model, small vesicles carry material between stable cisternae in both the anterograde and retrograde direction. A considerable body of evidence supporting either model, or variations thereof, has been presented but no study could yet unambiguously prove the validity of either of these models.Citation7-Citation12
Recently, however, two parallel studies in S.cerevisiae following the distribution of early and late Golgi markers over time demonstrated that Golgi cisternae do mature over time in this model organism.Citation13,Citation14 Since the discovery of the Golgi apparatus, many factors involved in its function, from traffic regulators to resident glycosylation enzymes have been identified. Although the molecular characterization of these proteins has unraveled a number of mechanisms underlying vesicular transport and protein modification, the fundamental processes maintaining Golgi homeostasis and regulating protein sorting at this organelle are still enigmatic. In addition, the role of lipids and the interplay between lipid and protein regulators at the Golgi are largely missing.
Both Arf regulation and phosphatidylinositol 4-phosphate (PI4P) metabolism are essential for Golgi function. Activation of Arf GTPases at the Golgi promotes the formation of both clathrin and COPI coated vesicles and is catalyzed by Arf guanine nucleotide exchange factors (Arf-GEFs) of the BIG/GBF subfamily.Citation15 However, the recruitment of a subset of Arf effectors requires the presence of PI4-phosphate in addition to activated Arf. How the simultaneous generation of two membrane recruitment signals is controlled is therefore a central question for our understanding of the regulation of vesicular membrane transport from the Golgi. In our efforts to unravel the interplay between key players of Golgi membrane transport in the small budding yeast S. cerevisiae we recently identified a physical interaction between the Arf-GEF Sec7p and the phosphatidylinositol (PI) 4-kinase Pik1p.Citation16 We demonstrated that Pik1p specifically interacts with only one of the three Golgi Arf-GEFs, and the two proteins share a common function in regulating the formation of clathrin-coated vesicle. We therefore proposed that the physical interaction of Pik1p and Sec7p coordinates Arf activation with PI4P production to generate a dual key recognition system controlling the specific recruitment of clathrin coats to the late Golgi. In addition, the extensive co-localization studies performed in the course of this study suggest that Gea1p and Gea2p, an alternative pair of Arf activators at the Golgi that do not bind to Pik1p, function at a different Golgi compartment and affect an earlier transport step than Sec7p and Pik1p. In agreement with the maturation model, our experiments suggested a minor overlap between the two kinds of compartments, which could reflect the evolution from one stage to the next by the progressive replacement of membrane proteins.
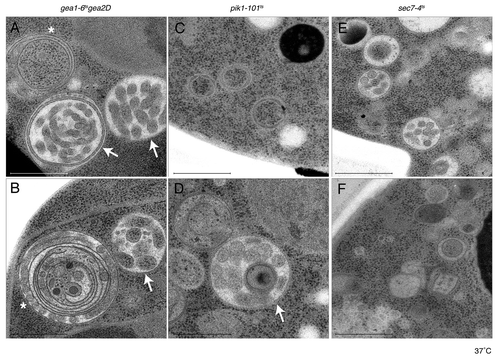
The interaction between Sec7p and Pik1p is also interesting to consider from the perspective of the maturation model. As lipid composition changes and membrane asymmetry increases between ER and plasma membrane, maturation of the Golgi apparatus does not only require a profound change in the protein but also the lipid composition of the Golgi membrane. This implies an important role for lipid modifying enzymes as Pik1p not only in vesicle formation but also in the maturation process itself. In the course of our studies, we prepared electron microscopy samples from different mutants (pik1–101ts, gea1–6ts∆gea2 and sec7–4ts) to directly compare the abnormal membrane accumulation described for each of those mutants.Citation17-Citation24 As expected, both sec7–4ts and pik1–101ts mainly accumulated Berkeley bodies (, , ), while gea1–6ts∆gea2 double mutants were devoid of them. These latter mutants preferentially accumulated various large and intricate membrane aggregates (, ). Interestingly, some of these larger structures were also observed in the pik1–101ts sample although less frequently (). As previously reported, a certain degree of ER swelling was also observed in all gef mutants, but not pik1–101ts. The large multi-lamellar structures seen in pik1–101ts and gea1–6ts∆gea2 are unlikely to be multivesicular bodies (MVBs) (arrows in , , ). Even if their shape sometimes reminds of these late endosomal intermediates, the structures observed here are more heterogeneous and comparison between different sections suggests a fenestrated structure rather than internalized vesicles. The origin of the large fenestrated compartments accumulating in both gea and pik1–101ts mutants is not clear. Due to their proximity to ER membranes and the involvement of Gea’s in retrograde transport from the Golgi, it is tempting to speculate that there could be a connection between these structures and early Golgi compartments. However, additional work is required to identify proteins associated with these structures. The small involuted structures in the sec7–4ts strain never reach a size comparable to the ones in pik1–101ts or gea1–6ts∆gea2 and no extensive tubulation of the internal structures was observed. Berkeley bodies were never obviously associated with the ER or any other membranes and have previously been observed in a variety of mutants affecting TGN exit and are likely to derive from late Golgi compartments.Citation18,Citation25 Hence, although Pik1p collaborates with Sec7p to recruit specific effectors to the late Golgi, pik1–101ts mutants accumulate abnormal membrane structures characteristic for both classes of Golgi ArfGEF mutants. This data suggests that, in addition to its role in coincidence detection together with activated Arf, PI4P production could be important to maintain the integrity of Golgi compartments beside the TGN, and Pik1p, as the sole yeast Golgi PI4-kinase might play a direct role in controlling organelle maturation.
Figure 1. pik1–101ts cells accumulate structures observed in either sec7–4ts or gea1–6ts∆gea2. Samples from gea1–6ts∆gea2 (panel A+B), pik1–101ts (panel C+D) and sec7–4ts (panel E+F) were prepared for electron microscopy by high-pressure freezing. Both pik1–101ts (C) and sec7–4ts (E-F) preferentially accumulate Berkeley bodies. The larger membrane structures accumulating in gea1–6ts∆gea2 (A-B) are occasionally also found in pik1–101ts (D). These structures can be sorted into two categories, either multi-layered ring-like structures (asterisk) or multi-lamellar structures (arrow). Bar = 500 nm.
The unique properties of the Golgi apparatus, which are reflected both by its unusual architecture and the dynamics of its remodeling remain a fascinating field of investigation. As evidence accumulate to support the cisternal maturation model in small organisms as the budding yeast S.cerevisiae, we now have to define/determine the role of vesicular transport in this context, identify the driving forces underlying the maturation process and understand how these events are co-regulated. Both Sec7p and Pik1p are likely candidates for a regulatory network which ensures the integrity of the Golgi apparatus by coordinating the flux of material entering and leaving this organelle with evolution of the cisternae. It will be interesting to extend the present findings to other organisms as the validity of the different Golgi progression models is presumably organismCitation7 and even cell-type dependent. In animal cells having more rigidly structured Golgi stacks, maturation is likely to be a slow process and the forward transport mainly supported by vesicular carriers, while in simpler organisms like budding yeast lacking discernable stacks, cisternal maturation might take the upper hand.
Material and Methods
Yeast strains used for electron microscopy were CSY712 (MATα ura3–52 leu2–3,112 pik1–101), yGY166 (MATα ura3–52 leu2–3,112 his3∆200 gea1–6 ∆gea2::HIS3) and AFM69–1A (MATα ura3–1 leu2 his3–11,15 sec7–4). AFM69–1A is a gift from D. Gallwitz. Yeast cultures, grown to early log phase, were cryoimmobilized using an EMPACT2+RTS high-pressure freezer (Leica), freeze-substituted in acetone containing 1% OsO4 + 0.1% uranyl acetate and embedded in Epon/Araldite.Citation26 Thin sections (70nm) were stained with 2% uranyl acetate in 70% methanol and lead citrate and observed with a TECHNAI_12 (FEI) transmission electron microscope operated at 80 kV.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
This work was supported by the Max Planck Society. We thank J. Maentler for expert technical assistance.
References
- Quatela SE, Philips MR. Ras signaling on the Golgi. Curr Opin Cell Biol 2006; 18:162 - 7; http://dx.doi.org/10.1016/j.ceb.2006.02.004; PMID: 16488589
- De Matteis MA, Morrow JS. The role of ankyrin and spectrin in membrane transport and domain formation. Curr Opin Cell Biol 1998; 10:542 - 9; http://dx.doi.org/10.1016/S0955-0674(98)80071-9; PMID: 9719877
- Stow JL, Heimann K. Vesicle budding on Golgi membranes: regulation by G proteins and myosin motors. Biochim Biophys Acta 1998; 1404:161 - 71; http://dx.doi.org/10.1016/S0167-4889(98)00055-X; PMID: 9714787
- Dunphy WG, Fries E, Urbani LJ, Rothman JE. Early and late functions associated with the Golgi apparatus reside in distinct compartments. Proc Natl Acad Sci U S A 1981; 78:7453 - 7; http://dx.doi.org/10.1073/pnas.78.12.7453; PMID: 6801652
- Young WW Jr.. Organization of Golgi glycosyltransferases in membranes: complexity via complexes. J Membr Biol 2004; 198:1 - 13; http://dx.doi.org/10.1007/s00232-004-0656-5; PMID: 15209092
- Wooding S, Pelham HR. The dynamics of golgi protein traffic visualized in living yeast cells. Mol Biol Cell 1998; 9:2667 - 80; PMID: 9725919
- Pelham HR, Rothman JE. The debate about transport in the Golgi--two sides of the same coin?. Cell 2000; 102:713 - 9; http://dx.doi.org/10.1016/S0092-8674(00)00060-X; PMID: 11030615
- Rabouille C, Klumperman J. Opinion: The maturing role of COPI vesicles in intra-Golgi transport. Nat Rev Mol Cell Biol 2005; 6:812 - 7; http://dx.doi.org/10.1038/nrm1735; PMID: 16167055
- Marsh BJ, Howell KE. The mammalian Golgi--complex debates. Nat Rev Mol Cell Biol 2002; 3:789 - 95; http://dx.doi.org/10.1038/nrm933; PMID: 12360195
- Elsner M, Hashimoto H, Nilsson T. Cisternal maturation and vesicle transport: join the band wagon! (Review). [Review] Mol Membr Biol 2003; 20:221 - 9; http://dx.doi.org/10.1080/0968768031000114024; PMID: 12893530
- Kartberg F, Elsner M, Froderberg L, Asp L, Nilsson T. Commuting between Golgi cisternae–mind the GAP! Biochim Biophys Acta 2005; 1744:351-63.
- Trucco A, Polishchuk RS, Martella O, Di Pentima A, Fusella A, Di Giandomenico D, et al. Secretory traffic triggers the formation of tubular continuities across Golgi sub-compartments. Nat Cell Biol 2004; 6:1071 - 81; http://dx.doi.org/10.1038/ncb1180; PMID: 15502824
- Losev E, Reinke CA, Jellen J, Strongin DE, Bevis BJ, Glick BS. Golgi maturation visualized in living yeast. Nature 2006; 441:1002 - 6; http://dx.doi.org/10.1038/nature04717; PMID: 16699524
- Matsuura-Tokita K, Takeuchi M, Ichihara A, Mikuriya K, Nakano A. Live imaging of yeast Golgi cisternal maturation. Nature 2006; 441:1007 - 10; http://dx.doi.org/10.1038/nature04737; PMID: 16699523
- Jackson CL, Casanova JE. Turning on ARF: the Sec7 family of guanine-nucleotide-exchange factors. Trends Cell Biol 2000; 10:60 - 7; http://dx.doi.org/10.1016/S0962-8924(99)01699-2; PMID: 10652516
- Gloor Y, Schöne M, Habermann B, Ercan E, Beck M, Weselek G, et al. Interaction between Sec7p and Pik1p: the first clue for the regulation of a coincidence detection signal. Eur J Cell Biol 2010; 89:575 - 83; http://dx.doi.org/10.1016/j.ejcb.2010.02.004; PMID: 20434792
- Audhya A, Foti M, Emr SD. Distinct roles for the yeast phosphatidylinositol 4-kinases, Stt4p and Pik1p, in secretion, cell growth, and organelle membrane dynamics. Mol Biol Cell 2000; 11:2673 - 89; PMID: 10930462
- Novick P, Field C, Schekman R. Identification of 23 complementation groups required for post-translational events in the yeast secretory pathway. Cell 1980; 21:205 - 15; http://dx.doi.org/10.1016/0092-8674(80)90128-2; PMID: 6996832
- Walch-Solimena C, Novick P. The yeast phosphatidylinositol-4-OH kinase pik1 regulates secretion at the Golgi. Nat Cell Biol 1999; 1:523 - 5; http://dx.doi.org/10.1038/70319; PMID: 10587649
- Peyroche A, Courbeyrette R, Rambourg A, Jackson CL. The ARF exchange factors Gea1p and Gea2p regulate Golgi structure and function in yeast. J Cell Sci 2001; 114:2241 - 53; PMID: 11493664
- Spang A, Herrmann JM, Hamamoto S, Schekman R. The ADP ribosylation factor-nucleotide exchange factors Gea1p and Gea2p have overlapping, but not redundant functions in retrograde transport from the Golgi to the endoplasmic reticulum. Mol Biol Cell 2001; 12:1035 - 45; PMID: 11294905
- Deitz SB, Rambourg A, Képès F, Franzusoff A. Sec7p directs the transitions required for yeast Golgi biogenesis. Traffic 2000; 1:172 - 83; http://dx.doi.org/10.1034/j.1600-0854.2000.010209.x; PMID: 11208097
- Wolf J, Nicks M, Deitz S, van Tuinen E, Franzusoff A. An N-end rule destabilization mutant reveals pre-Golgi requirements for Sec7p in yeast membrane traffic. Biochem Biophys Res Commun 1998; 243:191 - 8; http://dx.doi.org/10.1006/bbrc.1998.8084; PMID: 9473503
- Park SK, Hartnell LM, Jackson CL. Mutations in a highly conserved region of the Arf1p activator GEA2 block anterograde Golgi transport but not COPI recruitment to membranes. Mol Biol Cell 2005; 16:3786 - 99; http://dx.doi.org/10.1091/mbc.E05-04-0289; PMID: 15930122
- Jedd G, Mulholland J, Segev N. Two new Ypt GTPases are required for exit from the yeast trans-Golgi compartment. J Cell Biol 1997; 137:563 - 80; http://dx.doi.org/10.1083/jcb.137.3.563; PMID: 9151665
- Demmel L, Gravert M, Ercan E, Habermann B, Müller-Reichert T, Kukhtina V, et al. The clathrin adaptor Gga2p is a phosphatidylinositol 4-phosphate effector at the Golgi exit. Mol Biol Cell 2008; 19:1991 - 2002; http://dx.doi.org/10.1091/mbc.E06-10-0937; PMID: 18287542