Abstract
Recently we presented how Camponotus ants in Thailand infected with the fungus Ophiocordyceps unilateralis are behaviorally manipulated into dying where the conditions are optimal for fungal development. Death incurred in a very narrow zone of space and here we compare this highly specific manipulation with a related system in Brazil. We show that the behavioral manipulation is less fine-tuned and discuss the potential explanations for this by examining differences in ant host and environmental characteristics.
Keywords: :
Parasite manipulation of host behavior is an intriguing example of parasite adaptation. The change of host behavior is considered to be an extended phenotype of the parasite, as it can be explained as an expression of parasite genes in the host phenotype to increase parasite fitness.Citation1-Citation3 One of the most dramatic examples of a parasite extended phenotype is the manipulation of ant behavior by the fungus Ophiocordyceps unilateralis s.l.Citation4,Citation5
In a recent study we showed that the manipulation of the host ants by O. unilateralis s.l. is highly specific and beneficial to parasite fitness, thereby fulfilling the criteria of an extended phenotype.Citation6 Infected Camponotus leonardi worker ants leave their nest in the canopy and seek out the underside of a leaf in the undergrowth, bite into a leaf vein and die. The parasite then quickly colonizes the ant and grows for > 2 weeks before achieving reproduction. The death location of the ants was found to be far from random: dead infected ants were located 25.20 ± 2.46 SE cm above the ground, where the humidity and temperature were optimal for fungal growth, and on the north-northwest side of the plant biting onto a vein of the leaf. Parasites in the dead ants relocated from this ‘manipulative zone’ did not grow, confirming the adaptive value of the behavioral change. O. unilateralis was until recently believed to be a globally distributed species, but morphological studies in Brazil revealed a species complex with high host specificity.Citation7 Here we wanted to explore the data on adaptive manipulation further and infer the role of host characteristics, by comparison of the data on the death position from Thailand with that of ants in the related system of Ophiocordyceps camponoti-rufipedis (= unilateralis s.l.) infecting the ant Camponotus rufipes in Brazil. In addition, as hyperparasitism by mycoparasites is extensive and likely very costly for the parasite, we wished to elucidate whether the position where the ant was manipulated to die had an effect on the risk of hyperparasitism.
Fieldwork took place in February 2011 in Mata do Paraíso, a 400 ha Atlantic rainforest nature reserve in Minas Gerais, Brazil. The location and condition of 132 dead C. rufipes ants infected with O. camponoti-rufipedis was registered along a 460 meters stretch of forest path. For each dead ant we noted the height above ground, the orientation of the ant (which compass direction the head was pointing) and the parasite life stage as one of the following four categories: (1) a freshly killed ant (n = 26), (2) a cadaver with a stroma (stem-like structure that is the precursor to the production of a mature fruiting body; n = 28), (3) a cadaver with a mature sexual fruiting body (ascoma; n = 19), (4) a cadaver at stage 2 or 3, but hyperparasitized by other fungi (n = 59). The data were analyzed in JMP 9.0.2 for Mac and PAST 1.80 (available as free download at folk.uio.no/ohammer/past/) and compared with the data on height and orientation of dead C. leonardi ants infected with O. unilateralis s.l. obtained in Thailand as reported in Andersen et al.Citation6
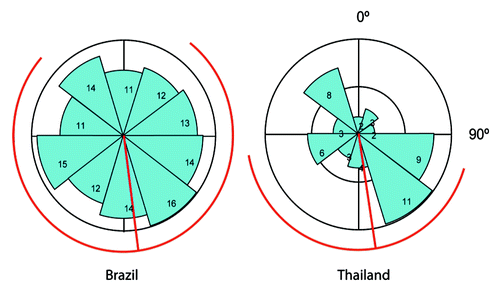
At the Brazilian site we found no difference in the height of dead infected ants between the parasite life stages or in the variation around the mean. We suggest that there is no relationship between the height at which ants are manipulated to die and the probability of the fungus reaching maturity or subsequently it self becoming a host to hyperparasites (, One-Way ANOVA Brazil: F3,128: 0.329, p = 0.804, Levene’s test of unequal variance: F3,128: 2.390 p = 0.0718). The dead infected ants were found higher up in Brazil than in Thailand and with a greater variance around the mean (, t-test assuming unequal variances t174.5: -15.506, p ≤ 0.0001, Levene’s test of unequal variance: F1,181: 89.555 p ≤ 0.0001). This may suggest a less fine tuned host manipulation in Brazil, potentially as a consequence of a wider height range of appropriate growth conditions. A temperature and humidity profile by height from the ground was not measured in Brazil, but may likely differ from Thailand, as the forest was denser. While both locations experience a wet and a dry season, the dry season in the Atlantic Rainforests of Minas Gerais, Brazil is correlated with lower temperatures and high humidity,Citation8 which may lower the risk of parasite desiccation. In addition, no pattern in the orientation of the ants was found in Brazil, in contrast to that of the Thai ants (). Note that the direction of the ant head is depicted, in contrast to location around the plant as in Andersen et al.,Citation6 so this is opposite to what we previously reported for Thailand as the majority of ants were facing the plant in Thailand. We do not know whether it is the location around the plant or the direction of the head that is relevant to the ant death location in Thailand, but neither appeared to matter in Brazil. We suggest that this is because infected ants in Brazil likely bite during the night, when the ants are most active, in contrast to at noon in Thailand.Citation9 This would eliminate the potential for using solar cues for orientation, e.g., by shade-seeking behavior. The ant C. rufipes is the dominant ant species at the field site in Brazil, where they nest on the ground in contrast to the canopy dwelling C. leonardi, which we recorded from over 20 meters up in the canopy. How this affects the parasite strategy is unknown, but it may make the host more accessible to the parasite in Brazil, as the dead infected ants are positioned in close proximity to the foraging trails of the host (R. Loreto et al. in preparation). However, the social immunity of the ant hostCitation10 is likely still the main challenge for the parasite, selecting for persistence in the environment to ensure transmission (S.B. Andersen et al. in preparation). In all, the comparison of the two host-parasite systems suggests that both parasites are highly adapted to their hosts but that environmental and host differences confer different strengths of selective pressure on the specificity of host manipulation.
Figure 1. The height above ground of dead infected ants. (A) shows that there were no significant differences between the height of dead ants in four different life stages in Brazil (fresh, stroma, mature and hyperparasitized (HP); mean height cm ± SD). The death height therefore does not affect which parasites reach maturity and which succumb to hyperparasitic fungi. (B) shows the difference between the height at which dead infected ants are found in Brazil and Thailand, where the ants die at a greater height in Brazil with a greater variance around the mean (mean height cm ± SD).
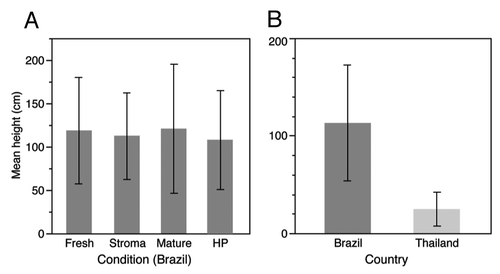
Figure 2. The direction of the heads of dead infected ants in Brazil and Thailand. In Brazil there was no pattern in the direction while the dead ants pointed toward South in Thailand. The blue slices show the number of dead ants in a given direction while the red lines indicate the mean head direction and the 95% confidence interval.
Acknowledgments
We thank Roberto Barreto and Simon Elliot at the Federal University of Viçosa, Minas Gerais, Brazil, and Harry C. Evans at the Federal University of Viçosa and CAB International, Surrey, UK for their kind hospitality. We are grateful to Jacobus J. Boomsma and Raquel Loreto for discussion. S.B.A. was funded by a PhD. Scholarship from the Science Faculty of the University of Copenhagen and D.P.H. was funded by an Outgoing International Marie Curie Fellowship.
References
- Poulin R. Manipulation of host behaviour by parasites: a weakening paradigm?. Proc Biol Sci 2000; 267:787 - 92; http://dx.doi.org/10.1098/rspb.2000.1072; PMID: 10819148
- Moore J. Parasites and the Behavior of Animals. Oxford, UK: Oxford University Press, 2002.
- Dawkins R. The Extended Phenotype. Oxford, UK: Oxford University Press, 1982.
- Evans HC, Samson RA. Cordyceps species and their anamorphs pathogenic on ants (Formicidae) in tropical forest ecosystems II. The Camponotus (Formicinae) complex. Trans Br Mycol Soc 1984; 82:127 - 50; http://dx.doi.org/10.1016/S0007-1536(84)80219-3
- Pontoppidan M-B, Himaman W, Hywel-Jones NL, Boomsma JJ, Hughes DP. Graveyards on the move: the spatio-temporal distribution of dead ophiocordyceps-infected ants. PLoS One 2009; 4:e4835; http://dx.doi.org/10.1371/journal.pone.0004835; PMID: 19279680
- Andersen SB, Gerritsma S, Yusah KM, Mayntz D, Hywel-Jones NL, Billen J, et al. The life of a dead ant: the expression of an adaptive extended phenotype. Am Nat 2009; 174:424 - 33; http://dx.doi.org/10.1086/603640; PMID: 19627240
- Evans HC, Elliot SL, Hughes DP. Hidden diversity behind the zombie-ant fungus Ophiocordyceps unilateralis: four new species described from carpenter ants in Minas Gerais, Brazil. PLoS One 2011; 6:e17024; http://dx.doi.org/10.1371/journal.pone.0017024; PMID: 21399679
- Sd CP, Venâncio MS, Barros NFd, Teixeira Dias HC. Produção de serapilheira em dois estádios sucessionais de floresta estacional semidecidual na Reserva Mata do Paraíso, em Viçosa, MG. Revista Árvore 2008; 32:545 - 56
- Hughes DP, Andersen SB, Hywel-Jones NL, Himaman W, Billen J, Boomsma JJ. Behavioral mechanisms and morphological symptoms of zombie ants dying from fungal infection. BMC Ecol 2011; 11:13; http://dx.doi.org/10.1186/1472-6785-11-13; PMID: 21554670
- Cremer S, Armitage SAO, Schmid-Hempel P. Social immunity. Curr Biol 2007; 17:R693 - 702; http://dx.doi.org/10.1016/j.cub.2007.06.008; PMID: 17714663