Abstract
Desert ants, Cataglyphis fortis, return to their nest when they are disturbed during their foraging trips. Training them to a landmark corridor enabled us to induce ants that were captured immediately after leaving the nest and transferred to an unknown area to start their foraging trips.1 However, most of the ants never traveled the entire foraging distance to the feeder, but aborted their runs after the landmark corridor was no longer visible. Therefore, apart from landmark information and path integrator, there are additional cues that determine the ants’ foraging behavior. Considering the reduced straightness of the outbound runs, I argue that surface structure might have a remarkable impact on foraging desert ants.
When disturbed, foraging desert ants, Cataglyphis fortis, tend to return immediately to their nest rather then to continue foraging (be it randomly or to a known food source).Citation2 They do so even if their sensory or motor systems have been severely damaged.Citation3,Citation4 The situation is similar when ants are about to leave their nest.Citation1 Due to this behavioral feature hitherto it was not possible to make desert ants proceed with or start their foraging runs after they had been displaced to an unfamiliar territory. As a consequence, we know much less about their navigational performance during foraging as compared to homing, which has been studied extensively.Citation5,Citation6
Due to their habitat, which consists of the bare desert ground of the North African Sahara where external markers and signposts are rarely present, C. fortis foragers continuously measure and integrate all their distances and directions, resulting in a vector that leads them home on a straight path (path integration).Citation6–Citation8 However, as soon as landmarks are available the ants use them to readjust their bearings and to find the location of their nest or a known feeding site.Citation9,Citation10 Depending on the circumstances (outbound or inbound run, distance to/from the nest) the information provided by landmarks can even out compete the information from the path integrator.Citation10,Citation11
Keeping that in mind, we showed in a recent paper that landmarks can change the foraging behavior of desert ants.Citation1 By training ants from their nest to a feeder through a corridor of landmarks (which were visible from a distance) we managed to make individuals start their foraging trips, even if they were captured shortly after leaving their nest and transferred to a remote test area. In contrast, when the landmarks were absent in the test or in both the training and the test area, only a few ants started foraging, whereas the majority searched for the nest instead.
In a previous experiment, we had already shown that the existence of landmarks can increase the number of ants that continue foraging after being disturbed half-way between their nest and a known feeder.Citation2 However, the effect of landmarks on these ants (that had already covered half of their foraging trip) was not as strong as the effect on the ants described above (that had just emerged from their nest).
In our most recent study, even when landmarks were present, we could not prevent one behavioral pattern. Most of the ants that had been transferred after leaving the nest and started foraging never reached the position of the fictive feeder. Instead, the majority returned to the point of release after only one outbound run into the trained direction (as a consequence, none of those individuals started to search for the feeder). Moreover, all ants first searched for the nest around the point of release before beginning their outbound runs.
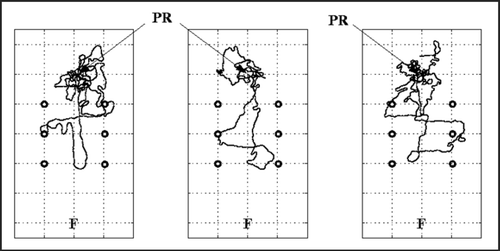
Thus, there seem to be additional cues that have an impact on the ants' foraging behavior. In other words, the motivation to forage is still not the same as compared to the training situation, even though the external circumstances were quite similar. The immediate search for the nest might be explained by the transfer procedure. After being captured and transferred the ants might have fallen back to their usual reaction when they are disturbed, which is to return to the nest (in this case, to search for the nest). However, this reasoning cannot fully explain why the ants did not cover the distance they had been trained to after they commenced foraging. They had obviously reached a motivational stage where they made at least one foraging attempt, but they did not perform a complete foraging excursion. Most of the ants aborted their foraging trips when the landmarks had vanished from their field of view (). However, even when the landmarks were still within their field of view, their runs were not very straight () as compared to runs of ants that were allowed to forage in their familiar terrain. Even ants that had been displaced at the feeder, and therefore had to perform their home runs in an unknown area, displayed straighter runs (Merkle, personal observation).
In our recent paper we mentioned three cues that could have an impact on the ants' foraging behavior: surface structure,Citation12,Citation13 panoramic landmarksCitation14 and the presence of nest mates. Though one cannot be sure which of those cues is the most important, the analysis of the outbound runs' structure allows us to go one step further. Nest mates are not very likely to be important for the exact heading of foraging desert ants, though their presence might be helpful during nest search and to reduce the uncertainty of homing ants. Panoramic landmarks cues have been shown to be used as a backup-mechanism for ants that are blown from their familiar area rather than being important for the straightness of the run. Therefore, I argue that ground structures, which have been shown to be detected by desert antsCitation12 and to have an impact on their systematic search behavior were likely to be an important factor in our experiment. When the ants had both their path integrator and the landmarks showing them the way to the feeder they did not attach great importance to the surface structure. That is, they headed towards the feeder, but did not follow the route as accurately as in the training area. When the landmarks were out of their field of view, the surface structure might have become more important and might have prevented the ants from proceeding with their foraging trips.
Figures and Tables
Figure 1 Paths of three desert ants that had been trained to forage from the nest to the feeder through a landmark corridor. The ants were captured immediately after leaving the nest and transferred to an unknown test area (with the same landmark arrangement). PR: points of release, F: fictive feeder positions, circles: landmarks. Mesh width of grid: 2 m.
Acknowledgements
I thank Rüdiger Wehner for including me in his research project on path integration in desert ants, his continuous support and encouragement, and the many—and still ongoing—stimulating discussions (also regarding this study).
Addendum to:
References
- Merkle T, Wehner R. Landmark guidance and vector navigation in outbound desert ants. J Exp Biol 2008; 211:3370 - 3377
- Merkle T, Wehner R. Landmark cues can change the motivational state of desert ant foragers. J Comp Physiol A 2008; 194:395 - 403
- Müller M, Wehner R. Path integration in desert ants, Cataglyphis fortis. Proc Natl Acad Sci USA 1988; 85:5287 - 5290
- Wittlinger M, Wehner R, Wolf H. The desert ant odometer: a stride integrator that accounts for stride length and walking speed. J Exp Biol 2007; 210:198 - 207
- Wehner R. Himmelsnavigation bei Insekten. Neurophysiologie und Verhalten. Neujahrsbl Naturforsch Ges Zürich 1982; 184:1 - 132
- Wehner R. Desert ant navigation: How miniature brains solve complex tasks. Karl von Frisch Lecture. J Comp Physiol A 2003; 189:579 - 588
- Collett M, Collett TS, Wehner R. Calibration of vector navigation in desert ants. Curr Biol 1999; 9:1031 - 1034
- Wehner R, Srinivasan MV. Jefferey KJ. Path integration in insects. The Neurobiology of Spatial Behaviour 2003; Oxford Oxford University Press 9 - 30
- Collett M, Collett TS, Bisch S, Wehner R. Local and global vectors in desert ant navigation. Nature 1998; 394:269 - 272
- Wehner R, Michel B, Antonsen P. Visual navigation in insects: coupling of egocentric and geocentric information. J Exp Biol 1996; 199:129 - 140
- Andel D, Wehner R. Path integration in desert ants, Cataglyphis: how to make a homing ant run away from home. Proc Biol Sci 2004; 271:1485 - 1489
- Seidl T, Wehner R. Visual and tactile learning of ground structures in desert ants. J Exp Biol 2006; 209:3336 - 3344
- Merkle T, Wehner R. Repeated training does not improve the path integrator in desert ants. Behav Ecol Sociobiol http://dx.doi.org/10.1007/s00265-008-0673-6
- Collett TS, Fauria K, Dale K. Jefferey KJ. Contextual cues and insect navigation. The Neurobiology of Spatial Behaviour 2003; Oxford Oxford University Press 67 - 82