Abstract
We investigated the symbiotic activities of fungal endophytes isolated from spotted knapweed, Centaurea stoebe. Previously, an analysis of community similarity had demonstrated differences in the endophyte communities of C. stoebe in its native and invaded ranges. Here, we found that specific endophytes can exert positive effects on their host, whereas others exert negative effects. Endophytes produced metabolites that inhibited germination of a competitor of C. stoebe. Endophytes also repelled a specialist insect herbivore, perhaps by producing biologically active volatiles. Yet other endophytes acted as cryptic pathogens of C. stoebe, suppressing its germination, reducing its growth, increasing the abundance of a generalist insect herbivore, and delaying or suppressing its flowering. Since, as reported here, endophytes are not functionally interchangeable, previously reported community differences could be contributing to the invasiveness of C. stoebe.
Recently we reported significant diversity in endophytic fungi in an invasive plant, Centaurea stoebe, or spotted knapweed.Citation1 Communities in the invaded and native ranges differed according to an analysis of similarity. Preliminary experiments to investigate functional activities of endophytes suggest that differences in the presence or absence of key endophytes could affect the invasiveness of this plant that is native to Eurasia and invasive in North America and elsewhere.
Positive Effects
Culture filtrates of 12 endophytes (Experiments 1–3, ) suppressed germination of Festuca idahoensis, a plant that competes with C. stoebe in its invaded range in western North America.Citation2 This result demonstrated that specific endophytes produce allelopathic effects that might aid C. stoebe in competition with other plants.
Symbionts can also have positive or mutualistic effects on their hosts by protecting them. Two endophytes, Alternaria CID62 and Epicoccum CID66 (CID = Cultivation Identification Number, or endophyte isolate number. A complete list of our CIDs is hereCitation1), appeared to protect C. stoebe from Larinus minutus, a seed-feeding weevil from the native range of C. stoebe, that was deliberately released in North America for biological control.Citation3 In dual-choice laboratory bioassays (Experiments 4–9), mated Larinus minutus females spent more time on uninoculated, control flowerheads than on those inoculated with either Alternaria CID62 or Epicoccum CID66, and preferred flowerheads inoculated with Epicoccum CID66 to those inoculated with Alternaria CID62 (). A similar pattern occurred when the isolated fungi were applied to cotton-flower mimics, except that the difference in preference for Epicoccum CID66 over Alternaria CID62 was not significant ().
The effects we have detected thus far are potentially mediated by chemical factors. We sampled each of 16 endophytes for their capacity to release volatile organic compounds (VOC) in pure culture (i.e., Experiment 10), following methods similar to those that have been used to detect biologically active VOC produced by an endophytic fungus.Citation4 Fourteen of these isolates in pure liquid culture produced at least one volatile sesquiterpene. Fusarium CID124 produced 20 distinct sesquiterpenes. Total production of sesquiterpenes ranged from zero to 236.8 ng/0.5 h/20 ml sample of culture. Volatile sesquiterpenes are implicated in many interorganismal interactions.Citation5
Negative Effects on Flowering
Although the endophytes reported thus farCitation1 are not overt pathogens they could be cryptic pathogens.Citation6 In Experiment 11, knapweed seedlings inoculated with Alternaria isolate ‘CID62’ produced fewer flowering heads than seedlings inoculated with Epicoccum CID66, Fusarium CID107, and an uninoculated, E− (i.e., endophyte-free) control (ANOVA F1,38 = 5.276, p = 0.03). In Experiment 12, seedlings inoculated with Alternaria CID123 and Fusarium CID124 flowered significantly later than E− controls (ANOVA F2,46 = 17.173, p < 0.001).
Negative Effects on Seed Germination
We also performed knapweed germination assays following inoculation with endophyte cultures (Experiments 13–15), or following treatment with liquid culture filtrates (Experiments 11–13); germination was 100% suppressed by Botrytis CID360, Alternaria CID120 and Fusarium CIDs 107 and 396, and many lesser, but still significant, effects were recorded.
Not only was germination of knapweed seeds entirely suppressed by Fusarium CID107, but a viability test with 0.1% unbuffered tetrazolium solution showed that seeds that failed to germinate were actually dead.
Negative Effects on Growth of C. stoebe
Some seedlings survived if they were first germinated and then inoculated with Fusarium CID107 (Experiments 14–16), but survivors had fewer and shorter leaves (ANOVA F1,52 = 8.987, p = 0.004 for number of leaves and ANOVA F1,52 = 7.307, p = 0.009 for length of maximal leaves) during a forty-day period of growth, and fewer mature, dissected leaves (χ2 test for independence, χ2 = 4.103, p = 0.043) than E− controls. Final, aboveground biomass was lower for Fusarium CID107-inoculated plants (ANOVA F1,50 = 11.292, p = 0.001) than E− controls.
Negative Effects on Protection of C. stoebe
Fusarium CID107 also attracted a generalist herbivore, the aphid, Myzus persicae, to plants it had infected. In Experiment 17, abundance of aphid infestations differed on E+ and E− knapweed seedlings (ANOVA F3,35 = 5.023, p = 0.005). Fusarium CID107-inoculated seedlings hosted aphid populations 6.3 times higher than plants inoculated with Alternaria CID62, Epicoccum CID66, or controls, although this difference eventually disapppeared when aphid populations became very large on all treatments (ANOVA F3,36 = 0.951, p = 0.426).
Balance of Positive and Negative Effects
With both negative and positive effects on characters associated with fitness (), it seems likely that endophytes strongly influence the ecology and invasiveness of C. stoebe. The effects of endophytes were seen in all growth stages of C. stoebe, from germination to flowering. Increases in aboveground biomass due to endophytes have been observed in other plants,Citation7–Citation10 although not yet in C. stoebe (). We expect that with further experimentation, we will discover many additional, biotic interactions mediated by endophytes in C. stoebe.
Figures and Tables
Figure 1 Results of 6 dual-choice experiments to determine the settling behavior of Larinus minutus on individual flowerheads or artificial flowers of spotted knapweed with and without inoculation by endophytes. Bars show the relative proportion of observations of weevils on the two treatments being compared over a 2-hour period. Asterisks indicate whether the results depart from equal proportions on each treatment (χ2, p = 0.05).
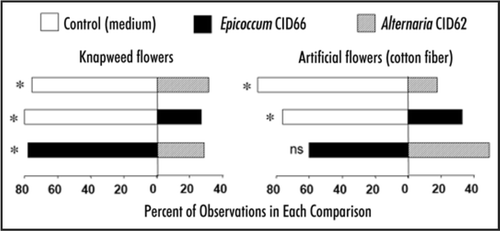
Figure 2 Growth, flowering and biotic interactions of C. stoebe, all significantly influenced by specific endophytes. Endophyte genera are followed by CID numbers that are keyed to GenBank accession numbers and to isolation frequencies in the native and invaded ranges of C. stoebe.Citation1

Table 1 A summary of experiments
Acknowledgements
R. Menjivar's participation in the project was supported by the Norman E. Borlaug International Agricultural Science and Technology Fellows Program
Addendum to:
References
- Shipunov A, Newcombe G, Raghavendra A, Anderson C. Hidden diversity of endophytic fungi in an invasive plant. Amer J Bot 2008; 95:1096 - 1108
- Callaway R, Aschehoug ET. Invasive plants versus their new and old neighbors: a mechanism for exotic invasion. Science 2000; 290:521 - 523
- Seastedt TR, Gregory N, Buckner D. Effect of biocontrol insects on diffuse knapweed (Centaurea diffusa) in a Colorado grassland. Weed Science 2003; 51:237 - 245
- Strobel GA, Dirkse E, Sears J, Markworth C. Volatile antimicrobials from Muscodor albus, a novel endophytic fungus. Microbiology 2001; 147:2943 - 2950
- Baldwin IT, Halitschke R, Paschold A, von Dahl CC, Preston CA. Volatile signaling in plant-plant interactions: “talking trees” in the genomics era. Science 2006; 311:812 - 815
- Redman RS, Dunigan DD, Rodriguez RJ. Fungal symbiosis from mutualism to parasitism: who controls the outcome, host or invader?. New Phytol 2001; 151:705 - 716
- Yuan Z-L, Dai C-C, Li X, Tian L-S, Wang X-X. Extensive host range of an endophytic fungus affects the growth and physiological functions in rice (Oryza sativa L.). Symbiosis 2007; 43:21 - 28
- Waller F, Achatz B, Baltruschat H, Fodor J, Becker K, Fischer M, et al. The endophytic fungus Piriformospora indica reprograms barley to salt-stress tolerance, disease resistance and higher yield. Proc Nat Acad Sci USA 2005; 102:13386 - 13391
- Ernst M, Mendgen KW, Wirsel SGR. Endophytic fungal mutualists: seed-borne Stagonospora spp. enhance reed biomass production in axenic microcosms. Mol Plant-Microb Interact 2003; 16:580 - 587
- Omacini M, Eggers T, Bonkowski M, Gange AC. Leaf endophytes affect mycorrhizal status and growth of co-infected and neighbouring plants. Funct Ecol 2006; 20:226 - 232
- Blair AC, Hanson BD, Brunk GR, Marrs RA, Westra P, Nissen SJ, et al. New techniques and findings in the study of a candidate allelochemical implicated in invasion success. Ecol Letts 2005; 8:1039 - 1047