Abstract
Wnt/β-catenin canonical pathway is critical for normal embryonic development; mutations and aberrant expression of specific components of this pathway can be oncogenic. Mitogen-activated protein kinase (MAPK) pathways, prominent in intracellular signaling, have been shown to have unique and provocative roles that impact the Wnt/β-catenin signaling. We discuss recent insights that implicate the three major pathways of the MAPK network, i.e., mediated by p38, c-Jun N-terminal (JNK) kinase, and Extra-cellular-Regulated Kinases (ERK) and their downstream signaling elements in Wnt/β-catenin signaling. Novel “crosstalk” among MAPK and Wnt/β-catenin canonical signalling pathways is essential. A fuller understanding of how such signaling is integrated during development is a high-value target for future research.
Introduction
Mitogen-activated protein kinases are a family of signaling molecules whose activity is regulated in response to a variety of stimuli, including growth factors, cytokines, or even environmental stress. MAPKs are the primary end points for a cascade that included upstream activation of MAP kinase kinase kinases (e.g., MAPKKK, MAP3K) that phosphorylate MAP kinase kinases (e.g., MAPKK, MAP2K) that, in turn, phosphorylate the MAPKs themselves (i.e., p38 MAPK, JNK1/2 and ERK1/2) ().Citation1,Citation2 MAPK pathways of this level of complexity and reversible phosphorylation/dephosphorylation constitute a large set of targets for input from cell signaling molecules not routinely associated with their operation, especially the canonical Wnt/β-catenin signaling.
Wnt signaling is essential for normal embryonic development, patterning and cellular proliferation; dysregulation of the function/expression of components of this pathway is linked to many forms of human cancer.Citation3–Citation6 Wnt ligands bind to G-protein-coupled receptors, termed “Frizzleds” (Fzs) and initiate intracellular signaling cascades ().Citation7,Citation8 The Wnt-sensitive pathways include not only the “canonical” Wnt/β-catenin,Citation7,Citation9 but also the non-canonical planar cell polarity and Wnt/cyclic GMP/Ca2+ pathways.Citation10–Citation14 In the absence of Wnt, cellular β-catenin is phosphorylated by the Serine/Threonine protein kinase, glycogen synthase kinase-3β (GSK3β), which is a component of a large multiprotein complex that includes the scaffold Axin and the product of the adenomatous polyposis coli gene. This complex is referred to as a “destruction complex” since it catalyzes the ubiquitination and proteasome-mediated degradation of β-catenin when Wnts are not present. Wnt3a binding to Fz1 activates G-proteins, Gαo and Gαq and the downstream phosphoprotein Dishevelled (Dsh/Dvl), provoking reduced GSK3β kinase activity, decreased phosphorylation of β-catenin by the complex, and increased stability and accumulation of intracellular β-catenin. Nuclear accumulation of β-catenin provokes activation of lymphoid-enhancer factor/T-cell factor (Lef/Tcf)-sensitive transcription of developmentally-related genes ().Citation15,Citation16 The central role of β-catenin in this canonical pathway is highlighted in studies of human cancers. In colon cancer, cells display increased levels of β-catenin that appear to result from high-frequency mutations in the adenomatous polyposis coli (APC) protein.Citation17 Mutations in β-catenin itself were also found to be responsible for several colon and other types of human cancers.Citation18,Citation19 The majority of the mutations of β-catenin reported are potential sites of GSK3β phosphorylation.Citation20,Citation21 Intracellular levels of β-catenin are tightly controlled, suggesting that multiple signaling pathways may be involved in regulating β-catenin levels and β-catenin-induced gene transcription. In this review, we highlight novel roles of the three MAPKs; p38 MAPK, c-Jun N-terminal kinase (JNK) and Extra-cellular regulated kinases (ERK) in the canonical Wnt/β-catenin signaling.
p38 MAPK
p38 is a family of MAPKs, highly conserved from yeast to mammals. p38 MAPKs also are activated in response to many extracellular stimuli including growth factors, cytokines and environmental stress.Citation22 Interestingly, Wnts have been reported to be capable of activating p38 MAPKs.Citation23–Citation25 In mesenchymal stem cells (MSCs), Wnt4 mediated activation of p38 MAPK was reported to be critical for enhancing osteogenic differentiation of MSCs.Citation24 Similarly, in C3H10T1/2 mesenchymal cells, Wnt3a induced transient activation of p38 and ERK MAPKs, which in turn regulate alkaline phosphatase activity and mineralization of nodules, suggesting a critical role for p38 in the progression of mesenchymal cells into osteoprogenitors.Citation25 Wnt3a was shown recently to stimulate p38 MAPK activation in totipotent mouse F9 teratocarcinoma cells: this activation of p38 by Wnt was shown to be dependent both on heterotrimeric G-proteins and Dishevelleds ().Citation23 More remarkably, Wnt-stimulated activation of p38 MAPK appeared to be regulating the canonical Wnt/β-catenin signaling.Citation23 The mechanism by which Wnt stimulated the p38 MAPK activation and regulated canonical Wnt/β-catenin signaling was unclear. Reports of a novel role for p38 MAPK in regulating GSK3β inactivation may provide some insight.Citation23,Citation26 By utilizing specific chemical inhibitors, gene-targeting small interfering RNAs (siRNAs), or expression of kinase-dead mutants of p38 MAPK, Wnt3a stimulation of p38 MAPK was shown to inactivate GSK3β. The loss of GSK3β function provoked an increase in cytosolic β-catenin accumulation and Wnt-sensitive gene transcription.Citation23 Thornton et al. have demonstrated that p38MAPK can phosphorylate GSK3β at Thr390 (corresponding to Ser389 in the mouse), inactivating GSK3β's kinase activity.Citation26 Thus, Wnt3a activates p38 MAPK and this p38 pathway feeds into the canonical Wnt/β-catenin pathway minimally at the level of GSK3β ().
c-Jun N-terminal Kinase (JNK) or Stress-Activated Protein Kinase (SAPK)
JNKs are activated in response to cytokines, UV irradiation or growth factor depletion, hence the name SAPKs. During Wnt signaling, activation of JNK is critical for the planar cell polarity pathway (PCP). Wnt ligands activate JNK in F9 teratocacrcinoma cells,Citation10 HEK 293 cellsCitation11 and also in WEHI-231, BAJI and Raji B-cells.Citation27 The Wnt3a-induced JNK activation operates via a pathway that is dependent on heterotrimeric G-proteins and Dishevelleds ().Citation10 Interestingly, apart from this central role in the Wnt-induced PCP pathway, JNKs also regulate the canonical Wnt/β-catenin pathway.Citation10,Citation28 Treating cells with a highly-specific JNK inhibitor, SP600125, was found to attenuate Wnt-induced Lef/Tcf-sensitive gene transcription.Citation10 Expression of kinase-dead mutants of the upstream protein kinases of the JNK MAPK cascade (RhoA, Rac1, Cdc42 and MEKKs), in contrast, had no significant effects on Wnt/β-catenin signaling.Citation10 Interestingly, JNK was found to regulate only the Wnt-induced Lef/Tcf-sensitive transcription (as measured by the Topflash luciferase activities). While, β-catenin accumulation or primitive endoderm formation in response to Wnt stimulation were largely unaffected after SP600125 treatment.Citation10,Citation28 Earlier studies suggest that phospho c-Jun (i.e., the primary product of JNK activation) interacts with the HMG-box transcription factor Tcf4,Citation29 while the AP-1 complex interacts with Lef1.Citation30 Equally provocative, Gan et al. reported that nuclear c-Jun mediates Dvl association with the TCFβ-catenin complex,Citation28 although the mechanism remains poorly described. Taken together, these observations suggest a novel insight, i.e., that the Wnt/Fz/JNK MAPK pathway and Wnt/Fz/β-catenin canonical pathway are in conversation with each other at the transcriptional level ().
Extra-Cellular Regulated Kinase (ERK1/2)
The ERK signaling cascade transmits signals from a variety of extracellular stimuli to multiple cellular processes, including cell proliferation, differentiation and development.Citation31 Wnt3a has been shown to activate ERK, stimulating proliferation of NIH3T3 cells via the Raf-1/MEK/ERK pathway ().Citation32,Citation33 Chemical inhibition of ERK activation (using MEK-specific inhibitors such as PD98059 or U0126) in either totipotent F9 teratocarcinoma cells or C3H10T1/2 mesenchymal cells has been shown to attenuate (although not knock-out) Wnt3a-sensitive Lef/Tcf gene transcription; whereas Wnt3a-induced β-catenin accumulation or primitive endoderm formation remain largely unaffected after treatment with such inhibitors.Citation10,Citation25 How Wnt3a-induced ERK activation can influence the read-out of Lef/Tcf-sensitive gene transcription, but yet not block the overall pathway to primitive endoderm formation suggests that the relationships between these pathways are neither linear nor fully appreciated in the functional sense. Wnt3a has been shown to provoke a strong nuclear accumulation of phospho-ERK in NIH3T3 fibroblasts.Citation33 Perhaps, Wnts can likewise provoke differential nuclear accumulation of phospho-c-Jun, phospho-ERK, and even phospho-p38. Such differential nuclear retention of phospho-MAPKs may in fact play some key role/s in the Wnt-induced Lef/Tcf-dependent transcription that can differentially effect the downstream signaling to the ultimate readouts ().
Conclusions and Perspectives
MAPK have long been appreciated to play key roles in cell proliferation, homeostasis and development. It is now evident that MAPK pathways also play crucial roles that can be characterized as a crosstalk with the canonical Wnt/β-catenin pathway central to development. Wnts are capable of stimulating both the Wnt/β-catenin pathway, MAPKs themselves, as well as signaling molecule intermediates, that constitute an intricate downstream network (). This organization provides for a conversation between these major signaling pathways, which are essential to not only signaling per se, but also to the broader landscape of spatial and temporal control of signaling events.Citation1,Citation2 This spatiotemporal quality of the cell signaling that undergirds development remains a vital and attracting target for novel biosensors and new technologies. Even with the current understanding on how these pathways cross-talk, probing key molecular interactions in such complex pathways will be required to eavesdrop on these subtle conversations. Molecules such as the Dishevelleds, which can clearly regulate three different Wnt-sensitive pathways as well as elements of the MAPK network, might well be the ideal nodes at which to tap into these molecular conversations. Recent studies in F9 teratocarcinoma cells and genetically modified mice clearly demonstrate Dishevelled 3, one of the three homologues of Dishevelleds, as a prime candidate for surveillance.Citation34,Citation35
Figures and Tables
Figure 1 Mitogen-activated protein kinase pathways. Mitogen-activated protein kinase (MAPK) pathways are stimulated by a variety of extra-cellular stimuli, including, growth factors, cytokines or environmental stress. The MAPK cascades constitute three sequentially activated kinase complexes. MAPKs, which include p38, c-Jun N-terminal kinase (JNK) and Extra-cellular regulated kinase (ERK) are substrates for phosphorylation by MAP kinase kinases (MKKs). The MAP kinase kinases are in turn phosphorylated by MAP kinase kinase kinases (MEKKs). The activated MAPK ultimately induce an appropriate cellular response. Different stimuli activate either p38 or JNK or ERK pathways via different combinations of MEKKs and MKKs. Note: for more information on MAP kinase proteins and their upstream kinases refer to Dong et al.Citation2
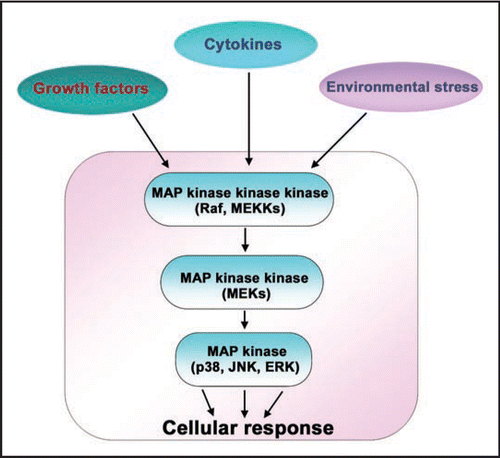
Figure 2 Wnt/-catenin signaling pathway. Wnts are secreted glycoproteins that bind to their cognate receptors, Frizzleds. Frizzleds belong to a family of heptahelical, G-protein-coupled receptors that bind specific Wnts and transduce the signal to downstream signalling pathways. Under basal conditions, cellular -catenin is phosphorylated by a Serine/Threonine kinase, Glycogen Synthase Kinase-3 (GSK3) in a complex along with Axin, adenomatous polyposis coli (APC) and other elements. The phosphorylated -catenin is constantly subjected to a proteasome-mediated degradation. Wnt-activated Frizzled (with its co-receptor low-density lipoprotein-related receptor protein-6 (LRP6)) activate the downstream signalling components including G-proteins, Go and Gq, and a phosphoprotein, Dishevelled. Dishevelleds, by an unclear mechanism, lead to reduced GSK3 kinase activity. Suppression of GSK3 activity leads to elevation of cytosolic and nuclear -catenin levels. Elevated nuclear -catenin levels activates the lymphoid-enhancer factor/T-cell factor (Lef/Tcf)-sensitive transcription, turning on the genes that are necessary for development. Note: for complete list of proteins involved in Wnt signalling, refer to the Wnt home page (http://www.stanford.edu/~rnusse/wntwindow.html).
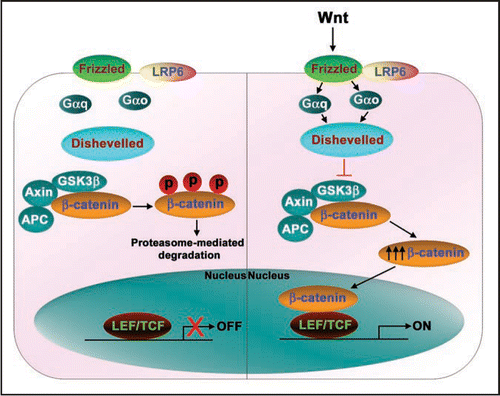
Figure 3 Conversations between Wnt/β-catenin and MAPK signaling pathways. Wnt ligands apart from activating Wnt/β-catenin pathway also activate MAPK pathways. Wnts induce a strong activation of p38 MAPK and this activation is G-protein and Dishevelled dependent. Wnt-activated p38 MAPK regulates Wnt/β-catenin signalling by inactivating GSK3β kinase activity. Similarly, Wnt-induced JNK activation occurs through G-proteins, Dishevelleds, small molecular weight GTPases (RhoA, Rac1, Cdc42), MEKKs and MKKs. Activated JNK phosphorylates its prime substrate, c-Jun, that ultimately leads induction of planar cell polarity. The phosphorylated c-Jun may also translocate into the nucleus and along with nuclear Dishevelled regulates Lef/Tcf-sensitive gene transcription. ERKs are also activated by Wnt through Raf1/MEK/pERK pathway and induce cell proliferation. In addition, the activated ERKs may also translocate into the nucleus and regulate the Lef/Tcf-sensitive gene transcription.
Acknowledgements
We thank Dr. H.-Y. Wang, Department of Physiology and Biophysics, State University of New York at Stony Brook, for the critical reading of the manuscript. We also thank members of the Malbon and Wang laboratories for their constant support and encouragement. This work has been generously supported by USPHS research awards from the National Institutes of Diabetes, Digestive, and Kidney Disease, National Institutes of Health to Craig C. Malbon.
References
- Johnson GL, Lapadat R. Mitogen-activated protein kinase pathways mediated by ERK, JNK and p38 protein kinases. Science 2002; 298:1911 - 1912
- Dong C, Davis RJ, Flavell RA. MAP kinases in the immune response. Annu Rev Immunol 2002; 20:55 - 72
- Logan CY, Nusse R. The Wnt signalling pathway in development and disease. Annu Rev Cell Dev Biol 2004; 20:781 - 810
- Moon RT, Bowerman B, Boutros M, Perrimon N. The promise and perils of Wnt signalling through beta-catenin. Science 2002; 296:1644 - 1646
- Moon RT, Kohn AD, De Ferrari GV, Kaykas A. WNT and beta-catenin signalling: diseases and therapies. Nat Rev Genet 2004; 5:691 - 701
- Polakis P. The many ways of Wnt in cancer. Curr Opin Genet Dev 2007; 17:45 - 51
- Bhanot P, Brink M, Samos CH, Hsieh JC, Wang Y, Macke JP, Andrew D, Nathans J, Nusse R. A new member of the frizzled family from Drosophila functions as a Wingless receptor. Nature 1996; 382:225 - 230
- Liu T, DeCostanzo AJ, Liu X, Wang Hy, Hallagan S, Moon RT, Malbon CC. G protein signalling from activated rat frizzled-1 to the beta-catenin-Lef-Tcf pathway. Science 2001; 292:1718 - 1722
- Liu T, Liu X, Wang H, Moon RT, Malbon CC. Activation of rat frizzled-1 promotes Wnt signalling and differentiation of mouse F9 teratocarcinoma cells via pathways that require Galpha(q) and Galpha(o) function. J Biol Chem 1999; 274:33539 - 33544
- Bikkavilli RK, Feigin ME, Malbon CC. Galphao mediates WNT-JNK signalling through dishevelled 1 and 3, RhoA family members, and MEKK 1 and 4 in mammalian cells. J Cell Sci 2008; 121:234 - 245
- Habas R, Dawid IB, He X. Coactivation of Rac and Rho by Wnt/Frizzled signalling is required for vertebrate gastrulation. Genes Dev 2003; 17:295 - 309
- Mlodzik M. Planar cell polarization: do the same mechanisms regulate Drosophila tissue polarity and vertebrate gastrulation?. Trends Genet 2002; 18:564 - 571
- Ma L, Wang HY. Mitogen-activated protein kinase p38 regulates the Wnt/cyclic GMP/Ca2+ non-canonical pathway. J Biol Chem 2007; 282:28980 - 28990
- Ma L, Wang HY. Suppression of cyclic GMP-dependent protein kinase is essential to the Wnt/cGMP/Ca2+ pathway. J Biol Chem 2006; 281:30990 - 31001
- Behrens J, von Kries JP, Kühl M, Bruhn L, Wedlich D, Grosschedl R, Birchmeier W. Functional interaction of beta-catenin with the transcription factor LEF-1. Nature 1996; 382:638 - 642
- Molenaar M, van de Wetering M, Oosterwegel M, Peterson-Maduro J, Godsave S, Korinek V, Roose J, Destrée O, Clevers H. XTcf-3 transcription factor mediates beta-catenin-induced axis formation in Xenopus embryos. Cell 1996; 86:391 - 399
- Kinzler KW, Vogelstein B. Lessons from hereditary colorectal cancer. Cell 1996; 87:159 - 170
- Rubinfeld B, Robbins P, El-Gamil M, Albert I, Porfiri E, Polakis P. Stabilization of beta-catenin by genetic defects in melanoma cell lines. Science 1997; 275:1790 - 1792
- Chan EF, Gat U, McNiff JM, Fuchs E. A common human skin tumour is caused by activating mutations in beta-catenin. Nat Genet 1999; 21:410 - 413
- Morin PJ, Sparks AB, Korinek V, Barker N, Clevers H, Vogelstein B, Kinzler KW. Activation of beta-catenin-Tcf signalling in colon cancer by mutations in beta-catenin or APC. Science 1997; 275:1787 - 1790
- Sparks AB, Morin PJ, Vogelstein B, Kinzler KW. Mutational analysis of the APC/betacatenin/Tcf pathway in colorectal cancer. Cancer Res 1998; 58:1130 - 1134
- Martin-Blanco E. p38 MAPK signalling cascades: ancient roles and new functions. BioEssays 2000; 22:637 - 645
- Bikkavilli RK, Feigin ME, Malbon CC. p38 mitogen-activated protein kinase regulates canonical Wnt-{beta}-catenin signalling by inactivation of GSK3{beta}. J Cell Sci 2008; 121:3598 - 3607
- Chang J, Sonoyama W, Wang Z, Jin Q, Zhang C, Krebsbach PH, Giannobile W, Shi S, Wang CY. Noncanonical Wnt-4 signalling enhances bone regeneration of mesenchymal stem cells in craniofacial defects through activation of p38 MAPK. J Biol Chem 2007; 282:30938 - 30948
- Caverzasio J, Manen D. Essential role of Wnt3a-mediated activation of mitogen-activated protein kinase p38 for the stimulation of alkaline phosphatase activity and matrix mineralization in C3H10T1/2 mesenchymal cells. Endocrinology 2007; 148:5323 - 5330
- Thornton TM, Pedraza-Alva G, Deng B, Wood CD, Aronshtam A, Clements JL, Sabio G, Davis RJ, Matthews DE, Doble B, Rincon M. Phosphorylation by p38 MAPK as an alternative pathway for GSK3beta inactivation. Science 2008; 320:667 - 670
- Shi CS, Huang NN, Harrison K, Han SB, Kehrl JH. The mitogen-activated protein kinase kinase kinase kinase GCKR positively regulates canonical and noncanonical Wnt signalling in B lymphocytes. Mol Cell Biol 2006; 26:6511 - 6521
- Gan XQ, Wang JY, Xi Y, Wu ZL, Li YP, Li L. Nuclear Dvl, c-Jun, beta-catenin and TCF form a complex leading to stabilization of beta-catenin-TCF interaction. J Cell Biol 2008; 180:1087 - 1100
- Nateri AS, Spencer-Dene B, Behrens A. Interaction of phosphorylated c-Jun with TCF4 regulates intestinal cancer development. Nature 2005; 437:281 - 285
- Rivat C, Le Floch N, Sabbah M, Teyrol I, Redeuilh G, Bruyneel E, Mareel M, Matrisian LM, Crawford HC, Gespach C, Attoub S. Synergistic cooperation between the AP-1 and LEF-1 transcription factors in activation of the matrilysin promoter by the src oncogene: implications in cellular invasion. FASEB J 2003; 17:1721 - 1723
- Yoon S, Seger R. The extracellular signal-regulated kinase: multiple substrates regulate diverse cellular functions. Growth Factors 2006; 24:21 - 44
- Kim SE, Choi KY. EGF receptor is involved in WNT3a-mediated proliferation and motility of NIH3T3 cells via ERK pathway activation. Cell Signal 2007; 19:1554 - 1564
- Yun MS, Kim SE, Jeon SH, Lee JS, Choi KY. Both ERK and Wnt/beta-catenin pathways are involved in Wnt3a-induced proliferation. J Cell Sci 2005; 118:313 - 322
- Lee YN, Gao Y, Wang HY. Differential mediation of the Wnt canonical pathway by mammalian Dishevelleds-1, -2 and -3. Cell Signal 2008; 20:443 - 452
- Etheridge SL, Ray S, Li S, Hamblet NS, Lijam N, Tsang M, Greer J, Kardos N, Wang J, Sussman DJ, Chen P, Wynshaw-Boris A. Murine dishevelled 3 functions in redundant pathways with dishevelled 1 and 2 in normal cardiac outflow tract, cochlea and neural tube development. PLoS Genet 2008; 4