Abstract
It has become increasingly clear that glucocorticoid (GC) signaling not only comprises classic nuclear receptor binding—that is, glucocorticoid receptors (GRs) to their response element in the nucleus—but also involves rapid, non-genomic efforts to regulate signaling cascades and other cell functions in the cytoplasm as well as other cell organelles. In a recent study, we found that GRs form a complex with B-cell lymphoma 2 (Bcl-2), translocate to mitochondria in response to corticosterone (CORT), and modulate mitochondrial calcium and oxidation in an inverted U–shaped manner. It is also well-established that steroid and thyroid hormone receptors regulate mitochondrial function to protect cells against various challenges and modulate synaptic plasticity. Here, we explore how such work reveals a fundamental mechanism whereby GCs regulate mitochondrial functions, and provides a mechanistic basis for therapeutic methods to rescue mitochondrial dysfunction during chronic stress or related psychiatric and neurodegenerative disorders.
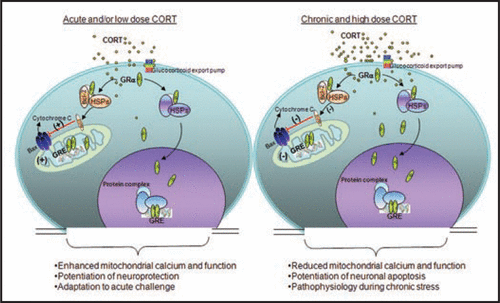
Glucocorticoids (GCs) act through their nuclear receptors in the brain to influence behavior and physiology.Citation1,Citation2 In addition to their well-known genomic effects on gene transcription, GCs also exert rapid, non-genomic effects on neurons in the brain.Citation3 Accumulating data have also shown that certain receptors traditionally considered to be nuclear receptors, particularly glucocorticoid receptors (GRs) translocate into mitochondria and modulate mitochondrial gene expression.Citation4–Citation6 However, the molecular mechanism of this regulation remains unclear. Recently, we found that in rat brain cells treated with corticosterone (CORT), GRs latched onto B-cell lymphoma 2 (Bcl-2), a protein that affects cytochrome C and calcium release from mitochondria. This GR/Bcl-2 complex moves into mitochondria and regulates mitochondrial functions in an inverted “U”-shaped manner (). Specifically, short-term exposure to CORT enhanced mitochondrial functions, while high doses or long-term treatment with CORT decreased levels of GRs and Bcl-2 in mitochondria (). Similar results occur in rats exposed to chronic CORT.Citation7
Such work suggests that, under physiological conditions, GCs enhance mitochondrial functions to provide cells with more energy for coping with and adapting to acute challenges. However, chronic stress may lead to chronically elevated levels of GCs, which in turn may reduce cell functioning via the interaction between GRs/Bcl-2 and mitochondria (). The decrease in proper cell function may contribute to the pathophysiology of several stress-related mental disorders, including major depressive disorder (MDD), and post-traumatic stress disorder (PTSD).
How Does the GR Protein Complex Translocate to the Mitochondria?
Previous studies have shown that GRs form protein complexes with heat shock protein 70/90 (HSP70/90) and Bcl-2-associated athanogene (Bag-1) in response to GC treatment.Citation8–Citation10 It is also well established that the proteins targeting mitochondria associate with chaperones that help in their mitochondrial translocation. One of the major chaperones in this category is HSP70.Citation11,Citation12 Based on the signal information in the precursor protein, it could be targeted to any of the four locations: the mitochondrial outer membrane, the mitochondrial inner membrane, the intermembrane space, or the mitochondrial matrix.Citation13 Furthermore, Bcl-2 is one of the tail-anchored molecules localized at the outer membrane of mitochondria.Citation14 In contrast, GRs travel to the matrix of the mitochondria and modulate mitochondria-coded protein expression.Citation15 It is possible that the GR/Bcl-2 complex shares the machinery for mitochondrial protein translocation by binding to HSP70/90 chaperone proteins, in much the same manner that estrogen receptors (ERs) can.Citation16
Bag-1, which binds to Bcl-2, is also a GR chaperone protein. Bag-1 attenuates nuclear translocation of GR, activates the extracellular receptor kinase (ERK) pathway, and potentiates the anti-apoptotic function of Bcl-2.Citation10,Citation17 In addition, Bag-1 transgenic mice showed less anxious-like behavior on the elevated plus maze test and more resilience in recovering from learned helplessness behavior and amphetamine-induced manic-like behaviors.Citation18 Because Bag-1 binds to both Bcl-2 and GRs, its role in GR/Bcl-2 complex translocation to mitochondria becomes a key issue.
Why Are Bcl-2 Family Genes a Key Modulator for Mitochondrial Function and Neuroprotection in Response to GCs?
Bcl-2 family proteins play a key role in apoptosis because of their ability to regulate the integrity of the mitochondrial outer membrane. Proteins such as Bcl-2 and Bcl-xL prevent the release of apoptogenic proteins from mitochondria and therefore protect against outer membrane permeabilization. Pro-apoptotic Bcl-2 family members—such as Bax and Bak—induce outer membrane permeabilization and cause the release of pro-apoptotic factors from mitochondria.Citation19,Citation20 The Bcl-2 proteins act either alone or together with other proteins like Bax to regulate the permeability of the outer membrane.Citation21 Our recent findings indicate that GRs form a complex with Bcl-2 in response to CORT treatment and, furthermore, translocate with Bcl-2 into mitochondria. This may be a critical step in blocking pore formation in the mitochondrial outer membrane. Therefore, calcium and cytochrome C release from the mitochondria would be attenuated ().
Additional findings have shown that Bcl-2 overexpression leads to enhanced mitochondrial calcium levels.Citation22 Notably, increased calcium levels operate mitochondrial metabolic checkpoints. Specifically, the aspartate/glutamate metabolite carriers are activated by calcium; in turn, recombinant expression of wild-type aspartate/glutamate metabolite carriers enhances adenosine triphosphate (ATP) production in response to cell stimulation.Citation23
Is the Hormone Receptor Protein Complex a Common Mechanism for Regulating Mitochondrial Function?
GRs, ERs, androgen receptors (ARs), progesterone receptors (PRs) and thyroid hormones (TRs) have been detected in the mitochondria of various cell types by western blotting, immunofluorescence labeling, confocal microscopy and immunogold electron microscopy.Citation24 Steroid and thyroid hormones regulate energy production. Thus, both steroid and thyroid receptors modulate the expression of nuclear-encoded genes and mitochondrial-encoded genes involved in oxidative phosphorylation.Citation13,Citation24 Recent studies also indicate that, via genomic and non-genomic mechanisms, these hormone receptors play a role in neuroprotection and apoptosis.Citation13,Citation24 In this context, our recent finding that the GR/Bcl-2 complex translocates into mitochondria to regulate mitochondrial function provides a novel mechanism for understanding GR-mediated neuroprotection.
Conclusion and Future Directions
The regulation of neuronal mitochondrial function by steroids is directly linked to neuroprotection and synaptic plasticity.Citation1,Citation25–Citation27 Furthermore, the central role that mitochondria play in neurodegeneration and psychiatric disorders has become apparent over the last decade as the molecular mechanisms that influence both neuronal death and neuroplasticity have been increasingly elucidated.Citation28 Accumulating evidence regarding mitochondrial translocation and the function of traditional “nuclear receptors” has revealed interesting and novel mechanisms in mitochondria that are regulated by steroids—particularly GCs—but also estradiol.Citation2,Citation16 However, the study of steroid hormone receptor regulation of mitochondrial function is still in its infancy. Future research will be needed to address several unanswered questions. For instance, what is the protective molecule that enhances GR/Bcl-2 trafficking to mitochondria in response to GCs? Is translocation of Bcl-2 family genes to mitochondria necessary for the modulation of mitochondrial functions by GCs? Do other traditional nuclear receptors form protein complexes with the Bcl-2 family of genes? The new knowledge gained from such studies will be instrumental in understanding the protection or dysfunction that steroid hormone receptors may exert via the regulation of mitochondrial function, and their role in the etiopathophysiology of stress-related psychiatric diseases such as MDD or PTSD.
Figures and Tables
Figure 1 Biphasic effect of Glucocorticoids (GCs) in regulating mitochondrial function. GCs are secreted by adrenal glands in a circadian and stress-related fashion. GCs readily penetrate the cell membrane and interact with cytoplasmic glucocorticoid receptors (GRs). GRs travel to the nucleus to regulate gene expression by binding to glucocorticoid response element (GRE). Here, GRs formed a complex with the anti-apoptotic protein B-cell-lymphoma 2 (Bcl-2) in response to corticosterone (CORT) treatment, and translocated with Bcl-2 into mitochondria after acute treatment with low or high doses of CORT in primary cortical neurons; they also upregulated mitochondrial calcium levels, membrane potential and oxidation. However, after long-term (three days) treatment, high, but not low, CORT decreased GR and Bcl-2 levels in mitochondria. In addition, three independent measures of mitochondrial function—mitochondrial calcium holding capacity, mitochondrial oxidation and membrane potential—were also regulated by long-term CORT treatment in an inverted “U”-shape. Bcl-2 was able to inhibit the formation of Bax-containing pores on the mitochondrial outer membrane and reduced the release of calcium and cytochrome C from the mitochondria. This regulation of mitochondrial function by CORT correlated with neuroprotection; that is, treatment with low doses of CORT demonstrated a neuroprotective effect, whereas treatment with high doses of CORT enhanced kainic acid (KA)-induced toxicity of cortical neurons.
Acknowledgements
We acknowledge the support of the Intramural Research Program of the NIMH. Ioline Henter provided outstanding editorial assistance.
Addendum to:
Related Research Data
References
- McEwen BS. Central effects of stress hormones in health and disease: understanding the protective and damaging effects of stress and stress mediators. Eur J Pharmacol 2008; 583:174 - 185
- McEwen BS, Milner TA. Hippocampal formation: shedding light on the influence of sex and stress on the brain. Brain Res Rev 2007; 55:343 - 355
- Croxtall JD, van Hal PT, Choudhury Q, Gilroy DW, Flower RJ. Different glucocorticoids vary in their genomic and non-genomic mechanism of action in A549 cells. Br J Pharmacol 2002; 135:511 - 519
- Sionov RV, Kfir S, Zafrir E, Cohen O, Zilberman Y, Yefenof E. Glucocorticoid-induced apoptosis revisited: a novel role for glucocorticoid receptor translocation to the mitochondria. Cell Cycle 2006; 5:1017 - 1026
- Koufali MM, Moutsatsou P, Sekeris CE, Breen KC. The dynamic localization of the glucocorticoid receptor in rat C6 glioma cell mitochondria. Mol Cell Endocrinol 2003; 209:51 - 60
- Moutsatsou P, Psarra AM, Tsiapara A, Paraskevakou H, Davaris P, Sekeris CE. Localization of the glucocorticoid receptor in rat brain mitochondria. Arch Biochem Biophys 2001; 386:69 - 78
- Du J, Wang Y, Hunter R, Wei Y, Blumenthal R, Falke C, et al. Dynamic regulation of mitochondrial function by glucocorticoids. Proc Natl Acad Sci USA 2009; 106:3543 - 3548
- Filipovic D, Gavrilovic L, Dronjak S, Radojcic MB. Brain glucocorticoid receptor and heat shock protein 70 levels in rats exposed to acute, chronic or combined stress. Neuropsychobiology 2005; 51:107 - 114
- Pratt WB, Morishima Y, Murphy M, Harrell M. Chaperoning of glucocorticoid receptors. Handb Exp Pharmacol 2006; 111 - 138
- Cato AC, Mink S. BAG-1 family of cochaperones in the modulation of nuclear receptor action. J Steroid Biochem Mol Biol 2001; 78:379 - 388
- Cvoro A, Dundjerski J, Trajkovic D, Matic G. Association of the rat liver glucocorticoid receptor with Hsp90 and Hsp70 upon whole body hyperthermic stress. J Steroid Biochem Mol Biol 1998; 67:319 - 325
- Dittmar KD, Demady DR, Stancato LF, Krishna P, Pratt WB. Folding of the glucocorticoid receptor by the heat shock protein (hsp) 90-based chaperone machinery. The role of p23 is to stabilize receptor.hsp90 heterocomplexes formed by hsp90.p60.hsp70. J Biol Chem 1997; 272:21213 - 21220
- Lee J, Sharma S, Kim J, Ferrante RJ, Ryu H. Mitochondrial nuclear receptors and transcription factors: who's minding the cell?. J Neurosci Res 2008; 86:961 - 971
- Wattenberg BW, Clark D, Brock S. An artificial mitochondrial tail signal/anchor sequence confirms a requirement for moderate hydrophobicity for targeting. Biosci Rep 2007; 27:385 - 401
- Demonacos C, Djordjevic-Markovic R, Tsawdaroglou N, Sekeris CE. The mitochondrion as a primary site of action of glucocorticoids: the interaction of the glucocorticoid receptor with mitochondrial DNA sequences showing partial similarity to the nuclear glucocorticoid responsive elements. J Steroid Biochem Mol Biol 1995; 55:43 - 55
- Nilsen J, Brinton RD. Mitochondria as therapeutic targets of estrogen action in the central nervous system. Curr Drug Targets CNS Neurol Disord 2004; 3:297 - 313
- Zhou R, Gray NA, Yuan P, Li X, Chen J, Chen G, et al. The anti-apoptotic, glucocorticoid receptor cochaperone protein BAG-1 is a long-term target for the actions of mood stabilizers. J Neurosci 2005; 25:4493 - 4502
- Maeng S, Hunsberger JG, Pearson B, Yuan P, Wang Y, Wei Y, et al. BAG1 plays a critical role in regulating recovery from both manic-like and depression-like behavioral impairments. Proc Nat Acad Sci USA 2008; 105:8766 - 8771
- Gottlieb RA. Role of mitochondria in apoptosis. Crit Rev Eukaryot Gene Expr 2000; 10:231 - 239
- Daniel PT, Schulze-Osthoff K, Belka C, Guner D. Guardians of cell death: the Bcl-2 family proteins. Essays Biochem 2003; 39:73 - 88
- Armstrong JS. Mitochondria: a target for cancer therapy. Br J Pharmacol 2006; 147:239 - 248
- Murphy AN. Ca2+-mediated mitochondrial dysfunction and the protective effects of Bcl-2. Ann N Y Acad Sci 1999; 893:19 - 32
- Lasorsa FM, Pinton P, Palmieri L, Fiermonte G, Rizzuto R, Palmieri F. Recombinant expression of the Ca2+-sensitive aspartate/glutamate carrier increases mitochondrial ATP production in agonist-stimulated Chinese hamster ovary cells. J Biol Chem 2003; 278:38686 - 38692
- Psarra AM, Sekeris CE. Steroid and thyroid hormone receptors in mitochondria. IUBMB Life 2008; 60:210 - 223
- Groc L, Choquet D, Chaouloff F. The stress hormone corticosterone conditions AMPAR surface trafficking and synaptic potentiation. Nat Neurosci 2008; 11:868 - 870
- Liu F, Day M, Muniz LC, Bitran D, Arias R, Revilla-Sanchez R, et al. Activation of estrogen receptor-beta regulates hippocampal synaptic plasticity and improves memory. Nat Neurosci 2008; 11:334 - 343
- Srivastava DP, Woolfrey KM, Jones KA, Shum CY, Lash LL, Swanson GT, et al. Rapid enhancement of two-step wiring plasticity by estrogen and NMDA receptor activity. Proc Natl Acad Sci USA 2008; 105:14650 - 14655
- Bachmann RF, Schloesser RJ, Gould TD, Manji HK. Mood stabilizers target cellular plasticity and resilience cascades: implications for the development of novel therapeutics. Mol Neurobiol 2005; 32:173 - 202