Abstract
By developing a technique for imaging the avian neural crest epithelial–mesenchymal transition (EMT), we have discovered cellular behaviors that challenge current thinking on this important developmental event, including the probability that complete disassembly of the adherens junctions may not control whether or not a neural epithelial cell undergoes an EMT. Further, neural crest cells can adopt multiple modes of cell motility in order to emigrate from the neuroepithelium. We also gained insights into interkinetic nuclear migration (INM). For example, the movement of the nucleus from the basal to apical domain may not require microtubule motors nor an intact nuclear envelope, and the nucleus does not always need to reach the apical surface in order for cytokinesis to occur. These studies illustrate the value of live-cell imaging to elucidate cellular processes.
During an epithelial-mesenchymal transition (EMT), epithelial cells lose high-affinity adherens junctions and apical-basolateral polarity, and gain mesenchymal cell characteristics such as motility and invasiveness.Citation1,Citation2 A classic example of an EMT is the emigration of neural crest cells from the dorsal neural tube of amniotes.Citation3,Citation4 By imaging the avian trunk neural crest EMT directly at high-resolution, we have gained several valuable insights about the cellular mechanisms of this EMT.Citation3
Complete Disassembly of Adherens Junctions is Neither Always Necessary nor Sufficient for an EMT
Currently, the most popular model for the trunk neural crest EMT is that the disassembly of adherens junction-mediated adhesion controls the emigration of neural crest cells from the neural tube.Citation5,Citation6 It has also been proposed that the separation of neural tube cells from the luminal adherens junctions by the plane of cell division might be sufficient to produce neural crest cells.Citation7 By imaging the neural crest EMT directly, we provide evidence that challenges these hypotheses. For example, as neural crest cells emigrate from the neuroepithelium, a small percentage rupture their cell tail and leave behind bits of cellular debris, including components of the adherens junctions, at the lumen.Citation3 Other times the adherens junction components move along with the retracting tail, which is compatible with electron microscopic data showing intact adherens junctions at non-apical positions in other emigrating cell types.Citation8,Citation9 Live-cell analysis of the ingression of zebrafish retinal ganglion cells also reveals that adhesion complexes are not completely disassembled prior to cell tail retraction, and also slide along with the retracting cell tail.Citation10 Therefore, based on our studies and others, the complete disassembly of adhesion complexes may not be necessary in order for cells to depart from an epithelium.
We also found that when mitotic neuroepithelial cells are separated from the luminal adhesions by the plane of cell division, most daughter cells re-integrate into the neuroepitheliumCitation3 (, see movie 1 in the Suppl. material). Hence, separation of cells from the adherens junctions is not always sufficient to generate neural crest cells. Zebrafish neural keel cells also re-integrate into the neural epithelium following cell division. Interestingly, this process is disrupted if the planar cell polarity protein Vangl2/Stabismus is mutated.Citation11 Therefore, although cell adhesion complexes are important for epithelial integrity, it seems likely that other molecular mechanisms, such as those that govern cell polarity, also control whether individual cells retain epithelial identity or not.
Modes of Neural Crest Motility
During an EMT, epithelial cells acquire the ability to emigrate out of the epithelium. Time-lapse analysis of the neural crest EMT reveals that, pre-neural crest cells adopted a range of migration phenotypes, forming filopodia, blebs and the occasional “lobopodium”Citation12 (, see Suppl. Movie 2). What is the mechanism of neural crest emigration? The classic model for cellular motility in vitro involves the formation of a lamellipodium, attachment to a substratum, directional contractile forces and release of the trailing end,Citation13,Citation14 although cells migrating in three-dimensional environments often prefer “blebbing” motility.Citation15,Citation16 It is likely that these cell processes are molecularly similar since they have been shown to be interconvertable in Fundulus deep cells.Citation12 Alternatively, because most neural crest cells undergo a tail retraction during the EMT,Citation3 it may be that actin contraction at the apical surface is important.Citation17,Citation18 Further still, the forces that drive the “sorting out” of germ layersCitation19 (likely regulated by changes in cortical tension)Citation20 could also be responsible for the segregation of neural crest cells from the neuroepithelium. Interestingly, the neural crest EMT can be stimulated ectopically by expression of constitutively active RhoB (together with Sox9 to prevent apoptosis).Citation21 The role of RhoB during the neural crest EMT could potentially promote blebbing motility or to create changes in cortical tension. Finally, it is likely that a combination of mechanical properties (protrusiveness, contractility and cell-matrix adhesions) contribute to the ability of neural crest cells to emigrate from the neural tube.Citation22
Insights into INM
A characteristic of the pseudostratified neuroepithelium is the occurrence of interkinetic-nuclear migration (INM).Citation23 During this event the nucleus migrates from a basal position to the apical surface where cytokinesis occurs, and then the daughter-cell nuclei are transported basally again. One of our novel observations was that the centrosome leaves the apical domain (where the cilium is nucleatedCitation24) just prior to apical nuclear migration and cytokinesis at the lumenCitation3 (). After the centrosome is released from the apical surface it may be pulled towards the nucleus by Lis1/dynein on microtubules.Citation25
Other insights into INM were acquired when we used mCherry-tubulin as a cytoskeletal marker for live-cell imaging. The biophysical forces that position the nucleus could involve myosin motors interacting with actin, or microtubule motors such as dynein or kinesin moving on microtubule tracks, both of which link to the nuclear envelope by KASH and SUN proteins.Citation26 Our imaging revealed that just prior to movement of the nucleus from a basal position to the apical domain and mitosis, the mCherry-tubulin signal in between the nucleus and the apical surface is reduced, which suggests that the microtubule track is degraded. Therefore nuclear movement in the apical direction may be microtubule independent (, see also Suppl. Movies 3–5). Instead, the shuttling of the nucleus apically prior to mitosis may involve actin/myosin, just as nuclear translocation in migratory neural precursor cells requires myosin II.Citation27 Also of note, the velocity of nuclear migration is faster in the apical direction prior to mitosis, as opposed to the basal direction following cell division,Citation3,Citation28 suggesting distinct mechanisms for moving the nucleus in opposite directions.
We also recorded a variety of behaviors suggesting that mitosis does not always occur at the apical surface. For example, there was an increase in the mCherrytubulin signal in the nuclear region prior to apical nuclear migration, indicating that cells can enter pro-metaphase (as evidenced by nuclear envelope breakdown) before the nucleus arrives at the apical surface (, see Suppl. Movie 3). Additionally, we recorded one example of mitotic spindle formation when the nucleus was still at a basal position prior to apical nuclear migration (, see Suppl. Movie 4), further indicating that the nucleus does not have to reach the lumen in order to initiate mitosis. Because the cytoskeletal-linking SUN and KASH proteins integrate into the nuclear envelope, Citation26 it is not known how the nucleus is transported apically following nuclearenvelope breakdown, although the nuclear matrix may not be completely dissolved after mitotic spindle formation.Citation29 Finally, we also discovered that cytokinesis can occur even if the nucleus does not reach the apical domain (, see Suppl. Movie 5).
In summary, our data show that the centrosome leaves the lumen prior to mitosis, that the movement of the nucleus apically prior to mitosis may be microtubule independent, and that nuclear-envelope breakdown, spindle formation and cytokinesis do not always have to occur at the apical surface.
These insights into the neural crest EMT and INM illustrate the value of livecell imaging and direct observation. In the future, the combination of molecular perturbations with live-cell imaging will be a valuable tool to further unravel the mechanisms of these events.
Abbreviations
| EMT | = | epithelial-mesenchymal transition |
| INM | = | interkinetic nuclear migration |
Figures and Tables
Figure 1 Following mitosis, dorsal neural tube cells can reinsert into the neuroepithelium. (A) A neural epithelial cell labeled with γ-tubulin-GFP (green) and mCherry-tubulin (red) and indicated within the white box is shown in the time-lapse series below. (A′) As this cell (white dot) undergoes interkinetic nuclear migration and mitosis at the apical surface, the centrosome (white arrowhead) leaves the apical surface and travels basally (0:14). In the next time frame (0:21), the nucleus moves towards the apical domain and meets the duplicated centrosomes in a non-apical location (white and blue arrowheads), and then the centrosomes and the nucleus travel together to the apical surface, where cytokinesis occurs (completed by 0:56). Note that the plane of cell division separates the basal-most daughter cell (white dot) from the apical surface. Following cell division, the centrosome (blue arrowhead) of the apical-most daughter cell (blue dot) is found at the lumen, whereas the centrosome (white arrowhead) belonging to the basal-most daughter cell (white dot) is found at a non-apical position. As the time-series progresses, the cell tail with the non-apical centrosome (white arrowhead) returns to the apical surface (complete by 4:26). Scale bars are 10 µm. D = dorsal, nt = neural tube, nc = neural crest, ap = apical.
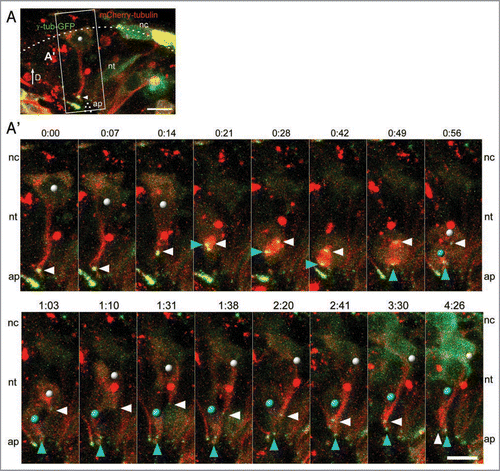
Figure 2 Neural crest emigration accompanied by filopodia and blebs. (A) The mem-EGFP-labeled neural epithelial cell indicated within the white box is shown in the time-lapse series below. (A′) Note that there are two different mem-EGFP-labeled cells close together within the field of view, a dorsal neural tube cell (red dot) and a neural crest cell (no dot). The neural epithelial cell (red dot) detaches from the lumen and migrates out of the neural tube. The formation of filopodia are indicated with red arrowheads and membrane blebs with red arrows. Scale bars are 10 µm.
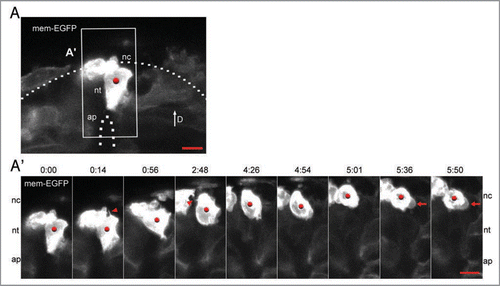
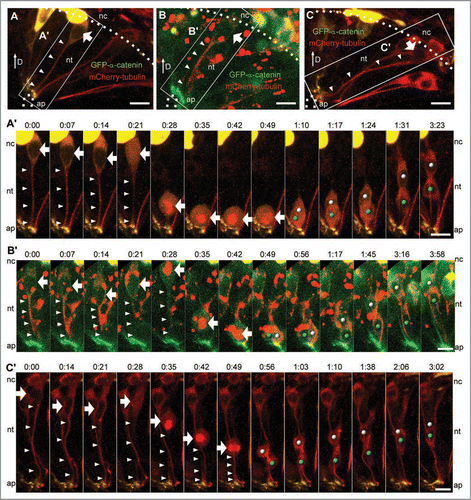
Figure 3 Examples of neural epithelial cells undergoing INM. (A–C) Neuroepithelial cells were labeled with GFP-α-catenin (green) and mCherry-tubulin (red). Cells within the white boxes are shown in the time-lapse series below. (A′) Prior to nuclear migration in an apical direction, the mCherry-tubulin signal in the cell tail is reduced (white arrowheads) and mCherry-tubulin enters the nucleus (white arrow, t = 0:21). The nucleus is then transported from a basal position to the apical surface, where the mitotic spindle forms (bright mCherry-tubulin signal) and cytokinesis occurs. The nuclei of the daughter cells (white and blue dots) are then transported basally. (B′) Before nuclear migration from the basal to apical surface, the mCherry-tubulin signal in the cell tail is reduced (white arrowheads) and mCherry-tubulin enters the nucleus (white arrow, t = 0:28). Mitotic spindle formation also occurs while the nucleus is in a basal position (bright mCherry-tubulin signal, t = 0:28). The nucleus is then transported to the apical surface where cytokinesis occurs. The daughter cell nuclei (white and blue dots) are subsequently shuttled to a basal position. (C′) Note the presence of two different mCherry-tubulin cell tails in the viewing field. Prior to nuclear migration from the basal to apical surface, the mCherry-tubulin signal in one cell tail is reduced (white arrowheads) and mCherry-tubulin enters the nucleus (white arrow, t = 0:28). Nuclear migration in the apical direction and spindle formation occurs at t = 0:35, but the nucleus does not make it all the way to the apical surface before cell division occurs (t = 0:56). After cytokinesis, the nuclei of the daughter cells (white and blue dots) are transported in a basal direction. Scale bars are 10 µm.
Additional material
Download Zip (2.6 MB)Acknowledgements
We would like to thank Dan Starr and the Erickson lab members for their helpful comments on the manuscript and discussions as this work progressed. We also acknowledge that our interpretation of INM events were shaped by ideas originating from Philip Spear, who is currently pursuing mechanisms of INM in the Erickson lab. This research was supported by grants from the NIH and the American Heart Association to CAE.
Addendum to:
References
- Ahlstrom JD, Erickson CA. Atala A, Lanza R, Thomson JA, Nerem RM. Molecular Organization of Cells. Principles of Regenerative Medicine
- Hay ED. The mesenchymal cell, its role in the embryo, and the remarkable signaling mechanisms that create it. Dev Dyn 2005; 233:706 - 720
- Ahlstrom JD, Erickson CA. The neural crest epithelial-mesenchymal transition in 4D: a “tail” of multiple non-obligatory cellular mechanisms. Development 2009; 136:1801 - 1812
- Le Douarin NM, Kalcheim C. The Neural Crest, Second Edition 1999; Cambridge Cambridge University Press
- Coles EG, Taneyhill LA, Bronner-Fraser M. A critical role for Cadherin6B in regulating avian neural crest emigration. Dev Biol 2008; 312:533 - 544
- Shoval I, Ludwig A, Kalcheim C. Antagonistic roles of full-length N-cadherin and its soluble BMP cleavage product in neural crest delamination. Development 2007; 134:491 - 501
- Erickson CA, Reedy MV. Neural crest development: the interplay between morphogenesis and cell differentiation. Curr Top Dev Biol 1998; 40:177 - 209
- Nichols DH. Mesenchyme formation from the trigeminal placodes of the mouse embryo. Am J Anat 1986; 176:19 - 31
- Viebahn C. Epithelio-mesenchymal transformation during formation of the mesoderm in the mammalian embryo. Acta Anat 1995; 154:79 - 97
- Zolessi F, Poggi L, Wilkinson C, Chien C-B, Harris W. Polarization and orientation of retinal ganglion cells in vivo. Neural Develop 2006; 1:1 - 21
- Ciruna B, Jenny A, Lee D, Mlodzik M, Schier AF. Planar cell polarity signalling couples cell division and morphogenesis during neurulation. Nature 2006; 439:220 - 224
- Trinkaus JP, Erickson CA. Protrusive activity, mode and rate of locomotion, and pattern of adhesion of Fundulus deep cells during gastrulation. J Exp Zool 1983; 228:41 - 70
- Lauffenburger DA, Horwitz AF. Cell migration: A physically integrated molecular process. Cell 1996; 84:359 - 369
- Sanz-Moreno V, Gadea G, Ahn J, Paterson H, Marra P, Pinner S, et al. Rac activation and inactivation control plasticity of tumor cell movement. Cell 2008; 135:510 - 523
- Charras G, Paluch E. Blebs lead the way: how to migrate without lamellipodia. Nat Rev Mol Cell Biol 2008; 9:730 - 736
- Berndt JD, Clay MR, Langenberg T, Halloran MC. Rho-kinase and myosin II affect dynamic neural crest cell behaviors during epithelial to mesenchymal transition in vivo. Dev Biol 2008; 324:236 - 244
- Lee J-Y, Goldstein B. Mechanisms of cell positioning during C. elegans gastrulation. Development 2003; 130:307 - 320
- Nakaya Y, Sukowati EW, Wu Y, Sheng G. RhoA and microtubule dynamics control cell-basement membrane interaction in EMT during gastrulation. Nat Cell Biol 2008; 10:765 - 775
- Armstrong PB. Cell sorting out: The self-assembly of tissues in vitro. Crit Rev Biochem Mol Biol 1989; 24:119 - 149
- Krieg M, Arboleda-Estudillo Y, Puech PH, Kafer J, Graner F, Muller DJ, et al. Tensile forces govern germlayer organization in zebrafish. Nat Cell Biol 2008; 10:429 - 436
- Cheung M, Chaboissier MC, Mynett A, Hirst E, Schedl A, Briscoe J. The transcriptional control of trunk neural crest induction, survival and delamination. Dev Cell 2005; 8:179 - 192
- Montell DJ. Morphogenetic cell movements: Diversity from modular mechanical properties. Science 2008; 322:1502 - 1505
- Baye LM, Link BA. Interkinetic nuclear migration and the selection of neurogenic cell divisions during vertebrate retinogenesis. J Neurosci 2007; 27:10143 - 10152
- Pugacheva EN, Jablonski SA, Hartman TR, Henske EP, Golemis EA. HEF1-dependent Aurora A activation induces disassembly of the primary cilium. Cell 2007; 129:1351 - 1363
- Tsai J-W, Chen Y, Kriegstein AR, Vallee RB. LIS1 RNA interference blocks neural stem cell division, morphogenesis and motility at multiple stages. J Cell Biol 2005; 170:935 - 945
- Starr DA. A nuclear-envelope bridge positions nuclei and moves chromosomes. J Cell Sci 2009; 122:577 - 586
- Tsai J-W, Bremner KH, Vallee RB. Dual subcellular roles for LIS1 and dynein in radial neuronal migration in live brain tissue. Nat Neurosci 2007; 10:970 - 979
- Del Bene F, Wehman AM, Link BA, Baier H. Regulation of neurogenesis by interkinetic nuclear migration through an apical-basal notch gradient. Cell 2008; 134:1055 - 1065
- De Souza CPC, Osmani SA. Mitosis, not just open or closed. Eukaryotic Cell 2007; 6:1521 - 1527