Abstract
Despite their name, resting state networks (RSNs) provide a clear indication that the human brain may be hard-working. Unlike the cardiac and respiratory systems, which greatly reduce their rate of function during periods of inactivity, the human brain may have additional responsibilities during rest. One particularly intriguing function performed by the resting brain is the consolidation of recent learned information, which is known to take place over a period of several hours after learning. We recently reported that resting state brain activity is modulated by recent learning. We measured the brain activity using functional MRI during periods of rest that preceded and followed learning of a sensorimotor task, and found a network of brain areas that changed their resting activity. These areas are known to be involved in the acquisition and memory of such sensorimotor tasks. Furthermore, the changes were specific to a task that required learning, and were not found after motor performance without learning. Here we discuss the implications and possible extensions of this work and its relevance to the study of memory consolidation.
The human brain has a high metabolic demand, and much of this energy is used to sustain spontaneous synaptic processing.Citation1 Such resting activity is not random; instead, there are highly organized patterns of coherent activity.Citation2 So while most functional neuroimaging experiments are based on detecting temporal correlations between brain signals and known input stimuli or output behaviors, newer analytical techniques allow identification of spatial networks defined only by their within-network correlations, unrelated to external events. This makes possible the identification of “resting state” brain systems without defining their time-course. The pattern of correlated activity defines specific segregated networks which are consistent across studies and even remain under sedation or during sleep.Citation3–Citation5
The function of such activity remains poorly understood but absence of a time-locked brain-behavior relationship does not preclude meaningful relationships between resting brain functions and behaviour. For example, spontaneous resting activity within the primary motor cortex can predict future response times to external stimuli.Citation6 Thus, resting activity may make the human brain highly responsive to external events, and so the resting brain affects our future behavior. Recently, we set out to examine the reciprocal relationship: the capacity for earlier behaviors to affect subsequent resting brain activity, and so in turn develop a deeper understanding of the resting brain.
Memory in the Resting Brain
We measured spontaneous patterns of activity within the resting brain before and after learning a new sensorimotor skill or sensorimotor performance, and showed that two circuits within the resting brain were specifically affected by sensorimotor learning.Citation7,Citation8 One included the frontal and parietal cortices, the other included the bilateral cerebellum. Both of these circuits have been implicated in sensorimotor learning,Citation9–Citation11 and it may be that these same areas continue to be engaged during memory consolidation processes that occur during the “off-line” period between performances. Unfortunately, in our recent work a measure of consolidation was never taken, and so within-group correlations between the amount of consolidation and the extent of the resting state changes were not possible. Nonetheless, we showed that the resting brain was affected by prior sensorimotor learning, and not by the mere performance of a sensorimotor skill, making an important connection between neuroplastic changes induced by learning and the resting brain. This is a necessary first step towards testing the idea that spontaneous resting activity is responsible for the off-line processing of memories during consolidation.Citation12
Identifying Task-Specific and Task-Independent Memory Consolidation Systems
Two circuits within the resting brain were affected by prior sensorimotor learning,Citation7 and some have suggested that two circuits underlie motor skill consolidation.Citation13 Our results may provide additional evidence for these dissociable consolidation systems. We found no learning related modulation of primary motor cortex (M1), which has been shown to m ake a critical contribution to consolidation of simple motor skills.Citation14,Citation15 It may be the case that the challenging visuomotor task learned in our experiment depends on consolidation within fronto-parietal circuits and does not require the contributions of M1.Citation16 Future work might usefully distinguish between those circuits that are differentially related to the consolidation of different types of skills—or even underlie the consolidation of other forms of memory—semantic or episodic—or learning (e.g., sequential or perceptual tasks).
Resting State and Brain State
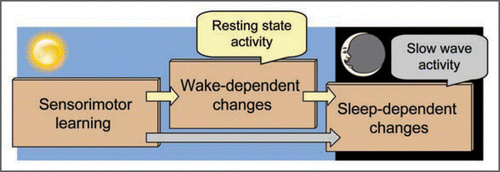
Memory consolidation can occur over wakefulness; but, some consolidation can only occur over sleep.Citation10,Citation13 Current theory suggests that anatomically distinct circuits support wake-dependent and sleep-dependent consolidation.Citation13,Citation17 One prediction from this theory is that two distinct circuits within the resting brain should be affected by prior sensorimotor skill learning.Citation7 It may be the case that resting state changes within one of these sensorimotor circuits correlates with wake-dependent performance changes;Citation7,Citation18 whereas, changes within the other circuit could correlate with sleep-dependent performance changes (). So our data might indicate that the fronto-parietal circuit is supporting memories that will undergo sleep-dependent consolidation, whereas the cerebellum is supporting memories undergoing wake-dependent consolidation. This hypothesis must be tested. Others have observed that increases in slow-wave activity (SWA) occur over the parietal cortex during sleep.Citation19 It may be that changes in spontaneous activity during wakefulness are responsible for triggering the subsequent increases in SWA during sleep. SWA appears to lead to synaptic modulations which may be a key mechanism for sleep-dependent consolidation.Citation19 This concept predicts that spontaneous resting state activity within the parietal cortex during wakefulness may be correlated with parietal SWA changes during sleep. Showing such a correlation would provide a strong link between the resting and the sleeping brain.
Changes within the resting brain may also occur as the circuits responsible for the initial encoding of a memory are reactivated for the consolidation of the memory.Citation20 Recent work has shown that memories can be reactivated during wakefulness,Citation21 and such neuronal replay may be delayed in other circuits until the onset of sleep when reactivation is thought to play a critical role in sleep-dependent consolidation.Citation22–Citation27
So learning, as our recent work shows, affects spontaneous activity within the resting brain. Understanding exactly how that resting activity is affected may provide insight into the mechanisms and pathways responsible for determining how memories consolidate over wakefulness or over sleep.
Figures and Tables
Figure 1 Consolidation across the day. Contributions of resting-state and slow-wave activity to memory consolidation may be differentiated based upon the times at which they occur—with resting-state networks supporting consolidation prior to the onset of sleep.
Acknowledgements
This work was supported by the Wellcome Trust (069439, R.C.M.) and by the US National Institutes of Health (R01 NS051446, E.M.R.).
Addendum to:
References
- Raichle ME, Mintun MA. Brain work and brain imaging. Annu Rev Neurosci 2006; 29:449 - 476
- Fox MD, Raichle ME. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat Rev Neurosci 2007; 8:700 - 711
- Vincent JL, Patel GH, Fox MD, Snyder AZ, Baker JT, Van Essen DC, et al. Intrinsic functional architecture in the anaesthetized monkey brain. Nature 2007; 447:83 - 84
- He BJ, Snyder AZ, Zempel JM, Smyth MD, Raichle ME. Electrophysiological correlates of the brain’s intrinsic large-scale functional architecture. Proc Natl Acad Sci USA 2008; 105:16039 - 16044
- Larson-Prior LJ, Zempel JM, Nolan TS, Prior FW, Snyder AZ, Raichle ME. Cortical network functional connectivity in the descent to sleep. Proc Natl Acad Sci USA 2009; 106:4489 - 4494
- Fox MD, Snyder AZ, Vincent JL, Raichle ME. Intrinsic fluctuations within cortical systems account for intertrial variability in human behavior. Neuron 2007; 56:171 - 184
- Albert NB, Robertson EM, Miall RC. The resting human brain and motor learning. Curr Biol 2009; 19:1023 - 1027
- Vincent JL. Learning and memory: while you rest, your brain keeps working. Curr Biol 2009; 19:484 - 486
- Inoue K, Kawashima R, Satoh K, Kinomura S, Sugiura M, Goto R, et al. A PET study of visuomotor learning under optical rotation. NeuroImage 2000; 11:505 - 516
- Robertson EM, Pascual-Leone A, Miall RC. Current concepts in procedural consolidation. Nat Rev Neurosci 2004; 5:576 - 582
- Galea JM, Albert NB, Ditye T, Miall RC. Disruption of the dorsolateral prefrontal cortex facilitates the consolidation of procedural skills. J Cogn Neurosci 2009; In press
- Miall RC, Robertson EM. Functional imaging: is the resting brain resting?. Curr Biol 2006; 16:998 - 1000
- Robertson EM. From creation to consolidation: a novel framework for memory processing. PLoS Biol 2009; 7:1000019
- Muellbacher W, Ziemann U, Wissel J, Dang N, Kofler M, Facchini S, et al. Early consolidation in human primary motor cortex. Nature 2002; 415:640 - 644
- Robertson EM, Press DZ, Pascual-Leone A. Off-line learning and the primary motor cortex. J Neurosci 2005; 25:6372 - 6378
- Baraduc P, Lang N, Rothwell JC, Wolpert DM. Consolidation of dynamic motor learning is not disrupted by rTMS of primary motor cortex. Curr Biol 2004; 14:252 - 256
- Cohen DA, Pascual-Leone A, Press DZ, Robertson EM. Off-line learning of motor skill memory: a double dissociation of goal and movement. Proc Natl Acad Sci USA 2005; 102:18237 - 18241
- Peigneux P, Orban P, Balteau E, Degueldre C, Luxen A, Laureys S, et al. Offline persistence of memory-related cerebral activity during active wakefulness. PLoS Biol 2006; 4:647 - 658
- Huber R, Ghilardi M, Massimini M, Tononi G. Local sleep and learning. Nature 2004; 430:78 - 81
- Marr D. Simple memory: a theory for archicortex. Phil Trans Royal Soc London Ser B, Biol Sci 1971; 262:23 - 81
- Karlsson MP, Frank LM. Awake replay of remote experiences in the hippocampus. Nat Neurosci 2009; 12:913 - 918
- Hoffman K, McNaughton B. Coordinated reactivation of distributed memory traces in primate neocortex. Science 2002; 297:2070 - 2073
- Wilson MA, Mcnaughton BL. Reactivation of Hippocampal Ensemble Memories during Sleep. Science 1994; 265:676 - 679
- Rasch B, Buchel C, Gais S, Born J. Odor cues during slow-wave sleep prompt declarative memory consolidation. Science 2007; 315:1426
- Cheng S, Frank LM. New experiences enhance coordinated neural activity in the hippocampus. Neuron 2008; 57:303 - 313
- Frank MG. The mystery of sleep function: current perspectives and future directions. Rev Neurosci 2006; 17:375 - 392
- Frank MG. Hippocampal dreams, cortical wishes: a closer look at neuronal replay and the hippocampalneocortical dialogue during sleep. Cell Sci Rev 2007; 3:161 - 171