Abstract
The automata theory is the mathematical study of abstract machines commonly studied in the theoretical computer science and highly interdisciplinary fields that combine the natural sciences and the theoretical computer science. In the present review article, as the chemical and biological basis for natural computing or informatics, some plants, plant cells or plant-derived molecules involved in signaling are listed and classified as natural sequential machines (namely, the Mealy machines or Moore machines) or finite state automata. By defining the actions (states and transition functions) of these natural automata, the similarity between the computational data processing and plant decision-making processes became obvious. Finally, their putative roles as the parts for plant-based computing or robotic systems are discussed.
Introduction
Natural computing is the recently growing field of research that investigates models and computational techniques inspired by nature and, dually, attempts to understand the world around us in terms of information processing.Citation1 It is a highly interdisciplinary field that combines the natural sciences and the theoretical computer science. The automata theory is the mathematical study of abstract machines commonly studied in the theoretical computer science.Citation2 Using these abstract machines called automata (singular, automaton), the computational problems can be properly solved. A finite-state automaton (FSA), known as the finite-state machine is a behavioral model used to design the computer programs, which is composed of a finite number of states associated to transitions. A transition is a set of actions allowed starting from one state and ending in the same or another state, which is started by a triggering event or condition. In biology and artificial intelligence researches, finite-state machine are sometimes used to describe the neurological systems.
In the present review article, as the chemical and biological basis for natural computing or informatics, some plants, plant cells or plant-derived molecules behaving as natural sequential machines or automata are listed and classified as natural sequential machines (namely the Mealy machines or Moore machines) or FSA. By defining the actions (states and transition functions) of these natural automata, the similarity between the computational data processing and plant decision-making processes became obvious. Finally, their putative roles as the parts for plant-based computing or robotic systems are discussed.
Handling and manipulating the life
As natural computing studies attempt to understand the world including the biological systems arrond us, some approaches may be similar to the those taken by synthetic biologists. For understanding the life, it is simpler and far easier to create a minimal model that functions as a life or to develop much more simplified models mimicking at least a part of life, which is a fundamental idea common to newly emerging system biology and synthetic biology.Citation3 Synthetic biologists engineer complex artificial biological systems to investigate the natural biological phenomena for a variety of applications.Citation4 Kurihara et al.Citation5 argued that the construction of protocells, from a materials-point of view, is important in understanding the origin of life.
A major goal of synthetic biology is to develop a deeper understanding of biological design principles from the bottom up, by building circuits and studying their behavior in the living cells.Citation6 Since the ability to quickly and reliably engineer many-component systems from libraries of standard interchangeable parts is one hallmark of modern technologies, nowadays, the synthetic biologists insist that we should adapt many of established frameworks for describing the existing engineered devices to the study and manipulation of biological objects.Citation7 To answer a question if simple biological systems can be built from standard, interchangeable parts and operated in living cells, synthetic biology is now expanding and affecting the educational sceneries. In the last decade, the International Genetically Engineered Machine Competition (iGEM) has been offering the chances for students and young researchers of various backgrounds to cope with each other to achieve designing and assembling of biological devices required for building the “genetic machines,” by using a library of standardized parts known as BioBricks.Citation8 Standard biological parts, such as BioBricks, may provide the foundation for a new engineering discipline that enables the design and construction of synthetic biological systems with a variety of applications in bioenergy, new materials, therapeutics, and environmental remediation.Citation9
Computing with Biological Materials
One of the aims in synthetic biology is the creation of intelligence based on the biological bricks. Biocomputers are man-made biological networks whose goal is to probe and control the biological hosts, namely the cells and organisms, in which they operate.Citation10 Their key design features, inspired by computer science and engineering, are programmability, modularity and versatility. While still the works are in progress, the state of the art “DNA-based biocomputer” now calculates a square root.Citation11 In the near future, due to robust growing of the area, biocomputers will eventually enable disease diagnosis and treatment with single-cell precision, lead to “designer” cell functions for biotechnology, and bring about a new generation of biological measurement tools.Citation10
Mealy Machines and Moore Machines: Two Different Types of Sequential Machines in vivo
As the machine meets an input, it makes a jump to another state, according to the transition functions defined. By defining the actions (states and transition functions) of these natural automata, the similarity between the computational data processing and plant decision-making processes became obvious. Here, we would like to compare two types of sequential machines, namely, Mealy machines and Moore machimes.
By definition, Mealy machine is represented formally by sextuple, viz., a three different sets of Q, Σ, and Δ, two functions δ, and λ, and the initial state q0.Citation2,Citation12 Note that (1) Q is a finite set of states; (2) Σ is a finite set of input symbols; (3) Δ is a finite set of output symbols; (4) δ is the state transition function which determines the next state q (∈Q) based on the combination of the present state p (∈ Q) and the input a (∈ Σ), that is, δ (p, a) = q; (5) λ is the output function which determines the output b (∈ Δ) based on the combination of the present state p (∈Q) and the input a (∈ Σ), that is, λ (p, a) = b; and (6) q0 (∈ Q) is the start state, that is, the state of the machine before any input has been processed.
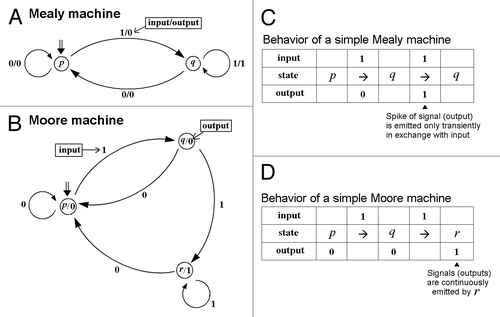
Similarly to a Mealy machine, a Moore machine is also represented formally by sextuple,Citation13 where Moore machine M = (Q, Σ, Δ, δ, λ, q0). The key difference between the Mealy and the Moore machines are temporal features of the output as illustrated in . This point is discussed in the later section.
Figure 1. State transitions in Mealy machine and Moore machine. These sequential machines (A) and (B) consist of states (represented by circles), and transitions (represented by arrows). The initial states are shown by the double arrows. As the machine meets an input, it makes a jump to another state, according to the transition function defined (based on the current state and the recent symbol of inputs). Above illustrations were adopted from ref.1 Temporal difference in the behaviors of two simple sequential machines are compared in (C) and (D). As shown in (C), Mealy machine's action is just to exchange an input event with an output event. In contrast, many signaling molecules may behave similarly to a Moore machine during signal transduction in aqueous computing or biological systems (D). Thus, any given single (chemical) event can be considered as an input for a receptor or protein involved in signaling. Once the molecule of interest is activated by single (chemical) event such as phosphorylation, binding to calcium, binding to the ligands, etc., the molecule becomes activated for certain length of time. During the activated state, the molecule (Moore machine) might keep acting by emitting multiple signals.
Volatile Memory and FSA in Plants
By considering an intact plant as an individual sequential machine, the signal processing (perception and transduction) in inside the living plant can be attributed to the automaton-like decision-making processes. The most notable case is the action of Venus flytrap (Dionaea muscipula) which possesses an active trapping mechanism to capture small insects.Citation14 Each of five to seven leaves on this plant has two parts, the upper and the lower leaves, which coordinately capture the insects. The trapping action by Venus flytrap has three unique features attracting experts not only in biology but also in chemistryCitation15 and physics,Citation16 namely, (1) it has one of the fastest movements in the plant kingdom, (2) it exhibits a “decision-making intelligence” to determine, from a semi-closed state, whether to proceed to be completely closed or fully open, and (3) the Venus flytrap has a “memory” of touch that two consecutive mechanical stimuli within about 30 sec are usually necessary to trigger the trap closing. As the sensory organs, three sensitive trigger hairs are protruding from the upper leaf epidermis of the Venus flytrap, adjacent to the red anthocyanin pigment that attracts the insects. According to recent studies, touching of the trigger hairs (by insects) rapidly activates the mechano-sensitive ion channels and generates receptor potentials, which induce an action potential required for trap closure. Recently, Volkov et al.Citation17 discovered that closing of the traps can be also induced upon electrical stimulation. The Venus flytrap can accumulate small sub-threshold charges, and when the threshold value is reached, the trap closes. Thus, repeated stimulation by smaller charges are counted and summed up to induce the closure of the leaves. Therefore, the researchers have argued that this plant has “volatile memory” for either mechanical touching or electrical stimulation.
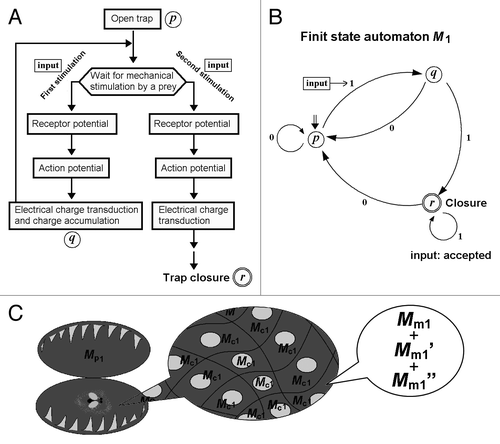
By assuming that Venus flytrap itself is an abstract sequential machine or there is an array of sensory cells or molecules equipped within the Venus flytrap, thus, functioning as a sequential machine, the functioning signal “transducer” or “recognizer” processing the mechanical stimuli in Venus flytrap must be interpreted either as a Mealy machine or Moore machine. Since a Moore-type sequential machine can be readily converted into a FSA, the system conserved in Venus flytrap is now described as a FSA M1 ().
Figure 2. Volatile memory processing determining the closure of the trap in Venus flytrap can be attributed to “automata.” (A) The simplified signaling mechanism of trap closure induced after processing the mechanical input in Venus flytrap, supported by experimental and theoretical analyses (Modified from Volkov et al.13). (B) Transition state of FSA M1 counting the number of stimuli. The states allowed in M1 are represented by circles, and the transitions are represented by the arrows. The initial state is shown by the double arrow and the final state is shown with the double circle. Input can be accepted (thus, closure induced) only after repeated stimuli. FSA M1 = (Q1, Σ1, δ1, q01, F1), where Q1 = {p, q, r}, Σ1 = {0, 1}, δ1(p, 0) = p, δ1(p, 1), = q, δ1(q, 0) = q, δ1(q, 1), = r, δ1(r, 0) = p, δ1(r, 1), = r, q01 = r, F1 = r. (C) The behavior of FSA M1 can be attributed to two types of metaphorical FSA Mp1 and Mc1 functioning as a whole plant and the cells composing the plant, respectively. At the level of molecular interactions, the function for Mc1 can be considered as synthesis of functions for various molecular FSA (Mm1, Mm1’, Mm1”…).
By definition,Citation2 an automaton (M) is represented formally by a quintuple, M = (Q, Σ, δ, q0, F): where (1) Q is a finite set of states; (2) Σ is a finite set of input symbols (the alphabet of the automaton); (3) δ is the state transition function (δ: Q × Σ → Q), which determines the next state q (∈ Q) based on the combination of the present state p (∈ Q) and the input a (∈ Σ), that is, δ (p, a) = q; (4) q0 (∈ Q) is the start state, that is, the state of the automaton before any input has been processed; and (5) F is a set of final states of Q (i.e., F ⊆ Q).
While the Venus flytrap model () deals with an intact plant as an individual FSA, the automata belonging to this type are not available as interchangeable biological parts to be used out of the living plants. In order to construct a library of interchangeable parts, further search for putative molecules behaving as automata is required. Furthermore, molecules encoded by specific genes, thus proteins, are preferable. In the below sections, the nature and the molecular basis for the abstract sequential machines or automata naturally equipped in living plants, as the interchangeable parts for synthetic biology are discussed.
Signaling Molecules as Moore Machines or Derived FSA
In electriccal computing models, the Mealy machines and Moore machines are considered to be equivalent, thus these machines can be interchangeable.Citation2 However, in the aqueous computing model or biological models, the molecular basis for these machines must be clearly distinguished. Especially the timing and continuity of the output are largely different as examples of the behaviors of two simple sequential machines are compared in . Here, this review propose a view that most of individual signaling molecules found in biological systems including plants can be categorized as the Moore machines or derived FSA. Let’s consider two distinct types of molecules acting as sequential machines placed in the aqueous system or an artificial cell. By definition illustrated in , upon receiving an effective input signal, a Mealy machine used as a signaling molecule may emit an output signal only once.
In contrast, a Moore machine may consecutively emit signals once the transition to the active state is manifested upon receiving an effective input signal. Thus, the input signal can be amplified by the output through Moore-type signaling molecules. The example of plant and non-plant signaling molecules acting in the manners similar to Moore machines include photoreceptors such as phytochrome and cryptochrome; many members of transporters and channel proteins such as aquaporins, cation permeable channels and anion channels; protein kinases such as mitogen-activated protein kinase (MAPK), MAPK kinase (MAPKK), MAPKK kinase (MAPKKK), protein kinase A (PKA), protein kinase C (PKC), plant Ca2+-dependent protein kinase (CDPK), Ca2+/calmodulin-dependent protein kinase (CaMK), and etc.; protein-binding and activating proteins such as calmodulins, trimeric G-proteins, samll G protein members and etc.; and DNA-binding proteins acting as transcription factors.
In case of action plants and locomotive cells of green algae, cytoskeletons and motor protein complexes controlled under signaling events can be considered as the Moore machines too. In the above models, intermediate signals such as phosphorylating events and releases of secondary messengers such as cyclic AMP and calcium ions can be considered both as the input and output signals for individual Moore machines or FSA involved in cellular signal transduction.
Plant Peroxidase as Redox-Mealy Machines
Despite the above section focused on the signal amplifying roles for the Moore-type signaling molecules, it should be also noted that plant enzymes are rich sources for the Mealy-type molecules. The given xamples with the roles of plant peroxidases in the plant signaling events involving salicylic acid (SA), aromatic monoamines (AMAs) and indole-3-acetic acid (IAA), brought in the below section clearly suggest that this type of molecules can participate the signaling pathways in plants, thus deserve being listed in additions to the Moore-type molecules and FSA from plants.
The production of reactive oxygen species (ROS), chiefly superoxide anion radicals (O2•-), hydrogen peroxide (H2O2), and hydroxyl radicals (HO•) at the cell surface, well known as the “oxidative burst” is one of the earliest events detectable during the incompatible interactions between plants and pathogens.Citation18 To date, multiple roles of ROS have been proposed in direct microbicidal actions, strengthening of cell wall through oxidative cross-linking of glycoproteins, induction of intracellular signaling pathway such as the synthesis of SA and activation of MAPK cascade, or activation of systemic acquired resistance associated with systemic propagation of the oxidative burst.
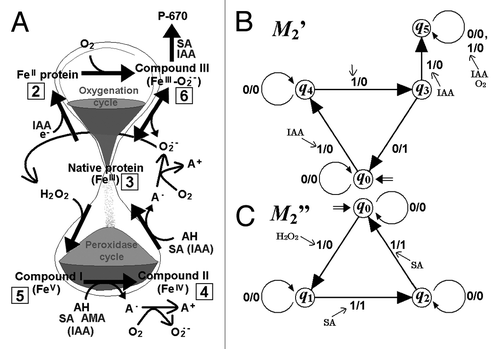
In plants, peroxidases achieve a great deal of oxidation reactions essential for the cells, using H2O2 as an acceptor of e- and a variety of substrates as e- donors, as the group of pioneering researchers of plant peroxidase in Geneva metaphorically described that plant peroxidases possess more functions than a ‘Swiss army knife’.Citation20 Indeed, highly diversified functions of plant peroxidases including regulation of H2O2 level, oxidation of various substrates, generation of ROS (coupled to oxidation of IAA, amines and SA) have been reported to date.Citation20 Oxidation of SA is one of the key functions of the ‘knife’.Citation19 In , byproducts of peroxidase-catalyzed oxidation of SA and IAA coupled to generation of O2•- are summarized. Previously, possible overall interactions between SA and plant peroxidases have been documented,Citation21,Citation22 and the estimated reactions for the generation of O2•- are as follows:
Native protein (3) + H2O2 → Compound I (5) + H2O
Compound I (5) + SA → Compound II (4) + SA•
Compound II (4) + SA → Native protein (3) + SA•
2 SA• + 2 O2 → 2 SA+ + 2 O2•-
Metabolism of IAA, the principal form of auxin in higher plants, is of great interest to plant biologists. Plant peroxidases are considered to be involved in the metabolism of IAA, by oxidizing IAA via two different mechanisms: a conventional H2O2-dependent pathway and one that requires O2 but not H2O2.Citation30-Citation32 The conventional peroxidase cycle for the oxidation of various substrates coupled to the consumption of H2O2 proceeds as follows:
Native protein (3) + H2O2 → Compound I (5) + H2O
Compound I (5) + S → Compound II (4) + P
Compound II (4) + S + H+ → Native protein (3) + H2O + P
IAA can be oxidized by plant peroxidases (chiefly by HRP in model experiments) by this mechanism but there is no strict substrate specificity in this conventional H2O2-dependent peroxidase cycle.Citation20 The plant peroxidases including HRP oxidize IAA also via the H2O2-independent pathway requiring molecular oxygen (O2), since most peroxidases of plant origins (but not animal or microbial origins) are considered to be highly specific IAA oxygenases, which possess the domains structurally similar to a specific motif in auxin-binding proteins.Citation30 The proposed reaction cycle for IAA oxidation is initiated via the formation of a ternary complex,Citation31 enzyme-IAA-dioxygen, yielding IAA cation radicals and O2•- as by-products as follows:Citation32
E + IAA ↔ [E-IAA],
[E-IAA] + O2 ↔ [E-IAA-O2],
[E-IAA-O2] → E + IAA•+ + O2•-,
Interestingly, the transitions among different oxidation states in plant peroxidases can be interpreted as the actions of Mealy machines. The models involving IAA () and SA () were shown as different redox Mealy machines. Not like the Moore machines, the members of the Mealy machines require highly repeated inputs for performing the substantially available levels of outputs by consecutively repeating the input-output cycles. Actually that is the case for plant peroxidases catalyzing a number of cycles of O2•--generating reactions to achieve the signaling upon repeated consumption of substrates such as SA and IAA.
Figure 3. Behavior of plant peroxidase as a redox-active mealy machine (M2). Based on the language (input signals) used, M2 can be separately described as two different automata (M2’ and M2”). (A) The hourglass model summarizing the superoxide generating reactions catalyzed by plant peroxidases responsive to both salicylic acid (SA, a model substrate for peroxidase cycle), aromatic monoamines (AMA) and indole-3-acetic acid (IAA, a model substrate for oxygenation cycle).20 (B) Redox-active Mealy machine M2’. M2’ = (Q2’, Σ2’, Δ2’, δ2’, λ2’, q02’), where Q2’ = {q0, q3, q4, q5,}, Σ2’ = {0, IAA, O2}, Δ2’ = {0, 1}, δ2’(q0, 0) = q0, δ2’(q0, IAA) = q4, δ2’(q0, O2) = q0, δ2’(q4, 0) = q4, δ2’(q4, IAA) = q4, δ2’(q4, O2) = q3, δ2’(q3, 0) = q0, δ2’(q3, IAA) = q5, δ2’(q3, O2) = q0, δ2’(q5, 0) = q5, δ2’(q5, IAA) = q5, δ2’(q5, O2) = q5, λ2’(q0, 0) = 0, λ2’(q0, IAA) = 0, λ2’(q0, O2) = 0, λ2’(q4, 0) = 0, λ2’(q4, IAA) = 0, λ2’(q4, O2), = 0, λ2’(q3, 0) = 1, λ2’(q3, IAA) = 0, λ2’(q3, O2) = 0, λ2’(q5, 0) = 0, λ2’(q5, IAA) = 0, λ2’(q5, O2) = 0, q02’ = q0. (c) Redox-active Mealy machine M2.” M2” = (Q2”’, Σ2,” Δ2”’, δ2,” λ2”’, q02”), where Q2” = {q0, q1, q2}, Σ2” = {0, H2O2, SA}, Δ2” = {0, 1}, δ2” (q0, 0) = q0, δ2”(q0, H2O2) = q1, δ2”(q0, SA) = q0, δ2”(q1, 0) = q1, δ2”(q1, H2O2) = q1, δ2”(q1, SA) = q2, δ2”(q2, 0) = q2, δ2”(q2, H2O2) = q2, δ2”(q2, SA) = q0, λ2”(q0, 0) = 0, λ2”(q0, H2O2) = 0, λ2”(q0, SA) = 0, λ2”(q1, 0) = 0, λ2”(q1, H2O2) = 0, λ2”(q1, SA), = 1, λ2”(q2, 0) = 0, λ2”(q2, H2O2) = 0, λ2”(q2, SA) = 1, q02” = q0. Note, Δ2’ = Δ2” = {0, 1} = {φ, O2·-}.
Dark logic models using inhibitors converts the biological Mealy machines into Moore machine
Discussion in the above section suggested that many of plant enzymes such as peroxidases can be considered as natural Mealy machines. However, the use of speficic inhibitors targetting the enzymes may behave as the inputs for the Moore machines when dark logic was employed. For an instance, by considering the presence of active enzyme as 0 and loss of activity as 1 (thus, dark logic), addition of plant peroxidase inhibitors such as salicylhydroxamic acid or binding of carbon monooxide to heme pockets of the enzymes results in the loss of catalytic activity. Therefore, the output from the inactivated state can be considered as 1.
In vivo cellular computing and plantoids
Above sections have indicated that the similarity between the computational digital data processing and plant decision-making processes by defining the actions (states and transition functions) of the natural automata in plant systems. Today, our approaches for synthetic biology targeting the bio-inspired computing and robotics have yet started. The idea of constructing plantoids (named after analogy to androids and humanoids) was proposed by the group of Prof. Mancuso (Univ. Florence, Italy). The plantoids are by definition, the plant-inspired robots proposed for the investigation of both biological and technological issues.Citation36,Citation37 Development of technologies applicable for such novel area of robotics is highly challenging topics to be achieved. It is also challenging to develop the intelligent plant-inspired or plant-related robots equipped with plant-derived automata, by connecting computer and natural or artificial plant cellular networks, for enhanced data processing.
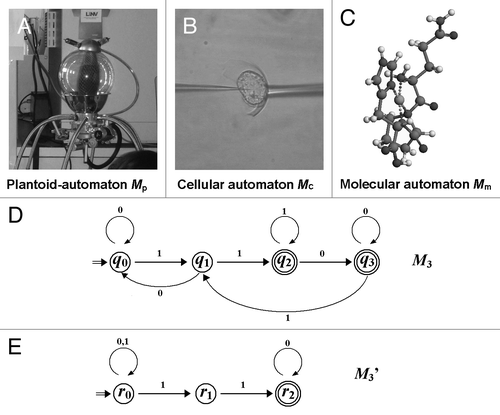
At present, we merely assume that the operational performance of plantoids can be designed based on the designs of abstract sequential machines or automata, thus, the plantoids may possibly function after installing the interchangeable parts derived from plant molecules or cells. Therefore, in , automata at different levels were proposed for future development of plantoids, namely, plantoid automata, cellular automata and molecular automata.
Figure 4. Proposed models for plantoids, cells, and molecules functioning as automata. (A) A model of plantoid produced by Prof. S. Mancuso (Univ. Florence). (B) Image of an electro-physiologically monitored cell (Prof. F. Bouteau, Univ. Paris-Diderot). (C) An artificial enzyme behaving as a plant peroxidase mimic.038 (D) State transitions in deterministic finite automata (DFA) M3. M3 = (Q3, Σ3, δ3, q03, F3), where Q3 = {q0, q1, q2, q3}, Σ3 = {0, 1}, δ3(q0, 0) = q0, δ3(q0, 1) = q1, δ3(q1, 0) = q0, δ3(q1, 1) = q2, δ3(q2, 0) = q3, δ3(q2, 1) = q2, δ3(q3, 0) = q3, δ3(q3, 1) = q1, q03 = q0, F3 = {q2, q3}. (E) State transitions in nondeterministic finite automata (NFA) M3’. M3’ = (Q3’, Σ3’, δ3’, q03’, F3’), where Q3’ = {r0, r1, r2}, Σ3’ = {0, 1}, δ3’(r0, 0) = {r0}, δ3’(r0, 1) = {r0, r1}, δ3’(r1, 0) = φ, δ3’(r1, 1) = r2, δ3’(r2, 0) = r2, δ3’(q2, 1) = φ, q03’ = r0, F3’ = r2.
Possible natural automata for robotic application found in other biological systems
In the present article, we have focused on the finding and defining the natural automata acting in living plants. Our ongoing study also suggests that natural automata such as Moore-type automata or FSA can be found and defined in living cells of protozoa. To many of cell biology researchers, Paramecium species are very familiar laboratory tools for cell biological and environmental studies.Citation39 Cells of Paramecium species are now considered as model systems for studying cellular signal transduction mechanisms, obviously equipped with minimal decisin-making propeties.Citation40 Since signal perception, processing and reactions are completed within these unicellular organisms, some researchers have described the cells of Paramecium species as “swimming sensory cells”Citation41 or “swimming neurons.”Citation42
It is well known that Paramecium species including the photosynthetic species, green paramecia (Paramecium bursaria), migrate toward the anodic electrode when exposed to an electric field in a medium.Citation43 This type of cellular movement is known as the galvanotaxis. In fact, over a century ago, it has been known that Paramecium species exhibit galvanotaxis in which the ciliate cells align with an electric field or voltage gradient and swim toward the anode if the electric field is sufficiently strong.Citation44 Recently, a pharmacological study has revealed that green paramecia is responsive to the electric field and the electric stimulus applied to green paramecia is converted to a galvanotactic cellular movement with the involvement of the T-type calcium channels on the plasma membrane.Citation43 The above studies indicate the possibility for finely geared neuronal controls and engineering of unicellular micro-machineries. In fact, the galvanotactic responsiveness observed in Paramecium species (particularly P. caudatum) has attracted the attention of bioengineers in the fields of biorobotics, microrobotics or BioMEMS (biological micro-electro-mechanical systems) in order to develop electrically controllable micro-machineries.Citation45-Citation47 Furukawa and his colleagues have suggested that in vivo cellular robotics using the cells of green paramecia as micro-machines controllable under electrical and optical stimuli, has a variety of engineering applications such as transport of micro-sized particles in the capillary systems.Citation48,Citation49 In fact Moore machines and derived FSA determining the direction of cellular migration (both galvanotactic and phototactic movements) in the cells of green paramecia can be defined (data not shown).
Perspectives: Deterministic finite automata (DFA) and nondeterministic finite automata (NFA)
Lastly, the authors wish to discuss the difference between the deterministic finite automata (DFA) and nondeterministic finite automata (NFA) to be applied in designing the plantoids or study in the in vivo cellular robotics.
Many of readers with biological backgrounds may wonder if the biological systems function as machines for informatics. Can every events and outcomes observed in the biological systems be considered clearly as the series of digitalized information expressed with 1 or 0, as discussed in the above sections? In fact, the answer is yes with some cautions. It is apparently obvious that the applications of automata in the biological topics require special cautions in order to reproduce the real biological phenomena. In nature, especially in biology, accuracy of the automata may largely vary from cell to cell, from molecule to molecule; thus, an experimentally forced input “1” to the automata in biological system could be resulted in the corresponding outputs with statistically determined range of variance. This is largely due to the fact that there are both diversity and redundancy in the sets of sensory or signaling molecules and/or cells differed in thresholds to the signal intensities or quantities. One of the likely answers to describe and/or design the plant-derived automata is the modification of the automata models from DFA to NFA, although this idea is yet to be tested through future researches. compares the DFA to NFA with equivalent processing capability. The latter model () allows two different responses upon input in the initial state. This type of lowered stringency could be achieved by involvement of multiple cells or molecules in the signaling events, thus the resultant performance of the automata would be much closer to the naturally observable phenomena.
| Abbreviations: | ||
| AMAs | = | aromatic monoamines |
| DFA | = | deterministic finite automata |
| FSA | = | finite-state automaton |
| H2O2 | = | hydrogen peroxide |
| HO | = | hydroxyl radicals |
| HRP | = | horseradish peroxidase |
| IAA | = | indole-3-acetic acid |
| NFA | = | nondeterministic finite automata |
| O2 | = | superoxide anion radicals |
| ROS | = | reactive oxygen species |
| SA | = | salicylic acid |
Acknowledgments
This work was supported by a Grants-in-Aid for Scientific Research by Ministry of Education, Culture, Sports, Science and Technology (MEXT), Japan (Research Project Number:23656495).
References
- Kari L. The many facets of natural computing. Commun ACM 2008; 51:72 - 83; http://dx.doi.org/10.1145/1400181.1400200
- Tomita Y, Yokomori T. Automata and formal language. 1992; Morikita Publ. Tokyo.
- Amos M. Genesis machines. New science of biocomputing. Atlantic Books 2006; London.
- Andrianantoandro E, Basu S, Karig DK, Weiss R. Synthetic biology: new engineering rules for an emerging discipline. Mol Syst Biol 2006; 2:2006 - , 0028; http://dx.doi.org/10.1038/msb4100073; PMID: 16738572
- Kurihara K, Tamura M, Shohda K, Toyota T, Suzuki K, Sugawara T. Self-reproduction of supramolecular giant vesicles combined with the amplification of encapsulated DNA. Nat Chem 2011; 3:775 - 81; http://dx.doi.org/10.1038/nchem.1127; PMID: 21941249
- Nandagopal N, Elowitz MB. Synthetic biology: integrated gene circuits. Science 2011; 333:1244 - 8; http://dx.doi.org/10.1126/science.1207084; PMID: 21885772
- Canton B, Labno A, Endy D. Refinement and standardization of synthetic biological parts and devices. Nat Biotechnol 2008; 26:787 - 93; http://dx.doi.org/10.1038/nbt1413; PMID: 18612302
- Brown J. The iGEM competition: building with biology. Synth Biol 2007; 1:3 - 6; http://dx.doi.org/10.1049/iet-stb:20079020
- Anderson JC, Dueber JE, Leguia M, Wu GC, Goler JA, Arkin AP, et al. BglBricks: A flexible standard for biological part assembly. J Biol Eng 2010; 4:1; http://dx.doi.org/10.1186/1754-1611-4-1; PMID: 20205762
- Benenson Y. Biocomputers: from test tubes to live cells. Mol Biosyst 2009; 5:675 - 85; http://dx.doi.org/10.1039/b902484k; PMID: 19562106
- Benenson Y. Biocomputing: DNA computes a square root. Nat Nanotechnol 2011; 6:465 - 7; http://dx.doi.org/10.1038/nnano.2011.128; PMID: 21814231
- Mealy GH. Method for synthesizing sequential circuits. Bell Syst Tech J 1955; 34:1045 - 79
- Moore EF. Tgedanken-experiments on sequential machines. In: Automata studies. Princeton Univ. Press, Princeton, USA. 1956; pp. 129-153.
- Volkov AG, Adesina T, Markin VS, Jovanov E. Kinetics and mechanism of Dionaea muscipula trap closing. Plant Physiol 2008; 146:694 - 702; http://dx.doi.org/10.1104/pp.107.108241; PMID: 18065564
- Ueda M, Nakamura Y, Okada M. Endogenous factors involved in the regulation of movement and” memory” in plants. Pure Appl Chem 2007; 79:519 - 27; http://dx.doi.org/10.1351/pac200779040519
- Li Y, Zhang M. Nonlinear dynamics in the trapping movement of the Venus flytrap. Proc. Am Control Conf. 2011; pp. 3514-351.
- Volkov AG, Adesina T, Jovanov E. Closing of venus flytrap by electrical stimulation of motor cells. Plant Signal Behav 2007; 2:139 - 45; http://dx.doi.org/10.4161/psb.2.3.4217; PMID: 19516982
- Yoshioka H, Bouteau F, Kawano T. Discovery of oxidative burst in the field of plant immunity: Looking back at the early pioneering works and towards the future development. Plant Signal Behav 2008; 3:153 - 5; http://dx.doi.org/10.4161/psb.3.3.5537; PMID: 19513209
- Passardi F, Cosio C, Penel C, Dunand C. Peroxidases have more functions than a Swiss army knife. Plant Cell Rep 2005; 24:255 - 65; http://dx.doi.org/10.1007/s00299-005-0972-6; PMID: 15856234
- Kawano T. Roles of the reactive oxygen species-generating peroxidase reactions in plant defense and growth induction. Plant Cell Rep 2003; 21:829 - 37; PMID: 12789499
- Kawano T, Sahashi N, Takahashi K, Uozumi N, Muto S. Salicylic acid induces extracellular superoxide generation followed by an increase in cytosolic calcium ion in tobacco suspension culture. The earliest events in salicylic acid signal transduction. Plant Cell Physiol 1998; 39:721 - 30; http://dx.doi.org/10.1093/oxfordjournals.pcp.a029426
- Kawano T, Muto S. Mechanism of peroxidase actions for salicylic acid-induced generation of active oxygen species and an increase in cytosolic calcium in tobacco cell suspension culture. J Exp Bot 2000; 51:685 - 93; http://dx.doi.org/10.1093/jexbot/51.345.685; PMID: 10938860
- Kawano T, Muto S, Adachi M, Hosoya H, Lapeyrie F. Spectroscopic evidence in support of horseradish peroxidase compound II-catalyzed oxidation of salicylic acid but not of phenylethylamine. Biosci Biotechnol Biochem 2002; 66:651 - 4; http://dx.doi.org/10.1271/bbb.66.651; PMID: 12005064
- Kawano T, Muto S, Adachi M, Hosoya H, Lapeyrie F. Spectroscopic evidence that salicylic acid converts a temporally inactivated form of horseradish peroxidase (compound III) to the irreversibly inactivated verdohemoprotein (P-670). Biosci Biotechnol Biochem 2002; 66:646 - 50; http://dx.doi.org/10.1271/bbb.66.646; PMID: 12005063
- Kawano T, Pinontoan R, Uozumi N, Miyake C, Asada K, Kolattukudy PE, et al. Aromatic monoamine-induced immediate oxidative burst leading to an increase in cytosolic Ca2+ concentration in tobacco suspension culture. Plant Cell Physiol 2000; 41:1251 - 8; http://dx.doi.org/10.1093/pcp/pcd052; PMID: 11092910
- Kawano T, Pinontoan R, Uozumi N, Morimitsu Y, Miyake C, Asada K, et al. Phenylethylamine-induced generation of reactive oxygen species and ascorbate free radicals in tobacco suspension culture: mechanism for oxidative burst mediating Ca2+ influx. Plant Cell Physiol 2000; 41:1259 - 66; http://dx.doi.org/10.1093/pcp/pcd053; PMID: 11092911
- Rutter R, Valentine M, Hendrich MP, Hager LP, Debrunner PG. Chemical nature of the porphyrin pi cation radical in horseradish peroxidase compound I. Biochemistry 1983; 22:4769 - 74; http://dx.doi.org/10.1021/bi00289a024; PMID: 6313048
- Schulz CE, Rutter R, Sage JT, Debrunner PG, Hager LP. Mössbauer and electron paramagnetic resonance studies of horseradish peroxidase and its catalytic intermediates. Biochemistry 1984; 23:4743 - 54; http://dx.doi.org/10.1021/bi00315a033; PMID: 6093863
- Kawano T, Pinontoan R, Hosoya H, Muto S. Monoamine-dependent production of reactive oxygen species catalyzed by pseudoperoxidase activity of human hemoglobin. Biosci Biotechnol Biochem 2002; 66:1224 - 32; http://dx.doi.org/10.1271/bbb.66.1224; PMID: 12162542
- Gazaryan IG, Lagrimini LM, Ashby GA, Thorneley RN. Mechanism of indole-3-acetic acid oxidation by plant peroxidases: anaerobic stopped-flow spectrophotometric studies on horseradish and tobacco peroxidases. Biochem J 1996; 313:841 - 7; PMID: 8611164
- Savitsky PA, Gazaryan IG, Tishkov VI, Lagrimini LM, Ruzgas T, Gorton L. Oxidation of indole-3-acetic acid by dioxygen catalysed by plant peroxidases: specificity for the enzyme structure. Biochem J 1999; 340:579 - 83; http://dx.doi.org/10.1042/0264-6021:3400579; PMID: 10359640
- Kawano T, Kawano N, Hosoya H, Lapeyrie F. Fungal auxin antagonist hypaphorine competitively inhibits indole-3-acetic acid-dependent superoxide generation by horseradish peroxidase. Biochem Biophys Res Commun 2001; 288:546 - 51; http://dx.doi.org/10.1006/bbrc.2001.5800; PMID: 11676477
- Kawano T, Kawano N, Lapeyrie F. A fungal auxin antagonist, hypaphorine prevents the indole-3-acetic acid-dependent irreversible inactivation of horseradish peroxidase: inhibition of Compound III-mediated formation of P-670. Biochem Biophys Res Commun 2002; 294:553 - 9; http://dx.doi.org/10.1016/S0006-291X(02)00513-2; PMID: 12056802
- Kawano T, Furuichi T, Muto S. Controlled free salicylic acid levels and corresponding signaling mechanisms in plants. Plant Biotechnol 2004; 21:319 - 35; http://dx.doi.org/10.5511/plantbiotechnology.21.319
- Takayama A, Kadono T, Kawano T. Heme redox cycling in soybean peroxidase: Hypothetical model and supportive data. Sens. Mater. 2012; 24:87 - 97
- Mazzolai B, Laschi C, Dario P, Mugnai S, Mancuso S. The plant as a biomechatronic system. Plant Signal Behav 2010; 5:90 - 3; http://dx.doi.org/10.4161/psb.5.2.10457; PMID: 20023403
- Mancuso S. The roots of plant intelligence. 2010; http://www.ted.com/talks/stefano_mancuso_the_roots_of_plant_intelligence.html.
- Kagenishi T, Yokawa K, Kadono T, Uezu K, Kawano T. Copper-binding peptides from human prion protein and newly designed peroxidative biocatalysts. Z Naturforsch C 2011; 66:182 - 90; http://dx.doi.org/10.5560/ZNC.2011.66c0182; PMID: 21630593
- Miyoshi N, Kawano T, Tanaka M, Kadono T, Kosaka T, Kunimoto M, et al. Use of Paramecium species in bioassays for environmental risk management: determination of IC50 values for water pollutants. J Health Sci 2003; 46:429 - 35; http://dx.doi.org/10.1248/jhs.49.429
- Pech LL. Regulation of ciliary motility in Paramecium by cAMP and cGMP. Comp Biochem Physiol A 1995; 111:31 - 7; http://dx.doi.org/10.1016/0300-9629(95)98516-J
- Machemer H, de Peyer JE. Swimming sensory cells: electrical membrane parameters, receptor properties and motor control in ciliated protozoa. Verh. Deutsche Zool Gesellschaft, Erlangen, 1977; pp. 86-110.
- Naitoh Y. Protozoa. In: Electrical Conduction and Behaviour in “Simple” Invertebrates, Ed. Shelton G.A.B., Clarendon Press, Oxford, 1982; pp. 1-48.
- Aonuma M, Kadono T, Kawano T. Inhibition of anodic galvanotaxis of green paramecia by T-type calcium channel inhibitors. Z Naturforsch C 2007; 62:93 - 102; PMID: 17425113
- Ludloff K. Untersuchungen uber den Galvanotropismus. Arch Gesamte Physiol 1895; 59:525 - 54; http://dx.doi.org/10.1007/BF01789963
- Itoh A. Motion control of protozoa for Bio MEMS. IEEE/ASME Trans Mechatron 2000; 5:181 - 8; http://dx.doi.org/10.1109/3516.847091
- Ogawa N, Oku H, Hashimoto K, Ishikawa M. Motile cell galvanotaxis control using high-speed tracking system. Proc IEEE Int Conf Robotics Automation 2004; pp. 1646-51.
- Ogawa N, Oku H, Hashimoto K, Ishikawa M. Dynamics model of paramecium galvanotaxis for microrobotic application. Proc IEEE Int Conf Robotics Automation 2005; pp. 1258-63.
- Furukawa S, Karaki C, Kawano T. Micro-particle transporting system using galvanotactically stimulated apo-symbiotic cells of Paramecium bursaria.. Z Naturforsch C 2009; 64:421 - 33; PMID: 19678550
- Furukawa S, Kawano T. Enhanced microsphere transport in capillary by conditioned cells of green paramecia used as living micromachines controlled under electric stimuli. Sens Mater 2012;24:375-86