Abstract
Referential acts play a crucial part in our every day communication since human language is, in its essence, a referential system. Reference can be made via icons, indices and signs but also via ostensive/inferential gestural communication, in which the behavior of the actor directs the attention of the recipient to particular aspects of the environment. The earliest uses of ostensive/inferential communication can be observed in human children around the age of nine to 12 mo. However, what about comparable gestures in our closest living relatives, the nonhuman primates or other animal taxa? The present paper aims to provide a brief overview of the state of the art to encourage future research into the evolutionary origins and uses of referential gestural signaling.
Keywords: :
Human language is, in its essence, a referential system, which involves cooperation of three subjects: a sign, its referent, and its interpretant.Citation1 Signs vary in the way they relate to their referents and are classified as icons, indices or true symbols.Citation1 An icon is related to its referent by virtue of some actual physical resemblance between the two entities. For instance, a flame is an icon for fire. An index is related to its referent by virtue of some literal physical participation in the referent-object event. For example, smoke is an index for fire because smoke is an event that is created by and thus consistently co-occurs with fire. The relationship between a symbol and its referent on the other hand is truly arbitrary, in that there is no physical resemblance or participation between the two. A sign is thus related to its referent only through the conventions agreed upon by a group and/or community of users.
However, referential communication can also be achieved by means of what Sperber and WilsonCitation2 call ostensive/inferential communication, in which the behavior of the actor directs the attention of the recipient to particular aspects of the environment. Of special importance is however that the receiver has to infer the signaler’s meaning, which is, contrary to signs, not codified in the behavior.Citation3 For example, by pointing to a raven in a tree one can transmit the same information as by saying “look, there is a raven in the tree!” The pointing gesture itself does not contain such information, but the specific meaning has to be inferred by the receiver. Ostensive/inferential communication has thus been linked by some researchers with cognitive capacities such as mental state attribution (e.g., see ref. Citation4).
An additional interesting property of ostensive/inferential communication is that it can occur even without the signaler having the intention to communicate (the recipient can discover the raven in the tree by simply following the signaler’s gaze). However, it is frequently identified with “intentional communication” to underscore the fact that it is especially powerful when subject to the intentional control of signalers.Citation5 Intentional communication or illocutionary behavior are behaviors, “in which the sender is aware a priori of the effect that a signal will have on his listener, and he persists in that behaviour until the effect is obtained or failure is clearly indicated.”Citation6
Interestingly, the onset of intentional ostensive/inferential behavior can be observed in pre-linguistic human children around the age of nine to 12 mo. Human children start to use gestures such as giving (for example, food objects), offering, pointing and showing,Citation6 to coordinate attention toward a social partner and an object of mutual interest.Citation7 These gestures create a referential triangle between signaler, recipient and a third entity and are used either to make requests (imperatives, for example, ‘take this’) or as a means to obtain ‘attention’ in the form of “laughter, comment, smiles and eye contact”Citation8 (declaratives, for example, ‘look at this’).
Since the use of referential gestures has been viewed as the foundation to engage in symbolically mediated conversations,Citation6,Citation9 a considerable amount of research attention has been focusing on referential gestural abilities of other animals, especially our closest living relatives, the nonhuman primates. Surprisingly however, observations of referential gestures in nonhuman primates are relatively rare and mainly concern “language trained” great apes and/or occur in interactions between apes and their human caretakers.Citation10-Citation14 The most compelling evidence of referential gestures in natural environments so far stems from adult chimpanzee (Pan troglodytes) males at the Ngogo community, Kibale National Park, Uganda, who use so-called directed scratches, to indicate distinct spots on their bodies to be groomed.Citation15 Recipients of these gestures understand the conveyed message and respond to it in appropriate ways by grooming the indicated spot after a directed scratch ().Citation15,Citation16
Figure 1. Use of the gesture directed scratch and response to the gesture. ©MPIO/Claßen
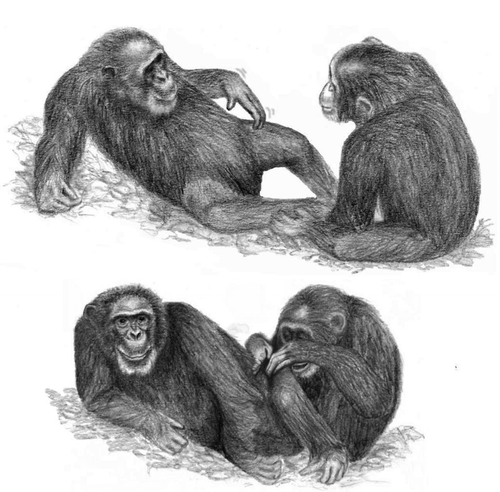
These gesture types are of special importance, because they represent, due to their two-tiered intentional structure (combining social intention to get something done and the ‘referential’ intention to draw the attention of the recipient to some third entity), an extremely rare form of communication evolutionarily.Citation17 Although these findings emphasize that referential gesturing represents a quite rare phenomenon in the natural communication of nonhuman primates (for an anecdote on pointing in bonobos in the wild see),Citation18 they suggest that (a) referential gestural signals may only occur in species living in a highly complex social system such as the fission-fusion system of the chimpanzee; and (b) their development may be triggered by distinct behavioral contexts such as the grooming and bonding context, in which negotiation of interactions and roles play a significant role.Citation19
To investigate these two hypotheses, we therefore instigated a study on natural referential gestural skills of ravens (Corvus corax), which (i) live similarly to chimpanzees in a highly complex social system,Citation20,Citation21 and (ii) engage in complex behaviors (e.g., preening, see ) to initiate and/or strengthen social bonds.Citation22,Citation23
Figure 2. Raven preening his partner. ©MPIO/Wascher

Natural communicative interactions of individually marked members of a wild raven community were filmed in Grünau, Northern Alps, Austria during three field seasons outside the courtship and breeding season.Citation24 We observed that ravens performed two distinct referential signals, showing (see ) and offering of non-edible items to recipients, which led to frequent orientation of receivers to the object and the signaler and subsequent mainly affiliative interactions.Citation24 Both signals clearly qualified as intentionally produced communicative gesturesCitation8,Citation25 and were understood by receivers. Although these gestures seem to resemble signals of males of some bird species in the context of courtship and mating,Citation26 mainly triggered through a distinct hormonal status (for a discussion of motivational vs. referential signals, see ref. Citation27), there are two crucial differences: First, raven’s use of referential gestures is divorced from sexual reproduction since they were observed outside the courtship and breeding season; and second, it was not restricted to possible mating partners but also used between siblings ().
Figure 3. Use of the gesture showing to two other conspecifics. ©MPIO/Bugnyar

These observations thus provided the first systematic evidence, that referential gestural signaling is not restricted to the primate lineage only and may have evolved in ravens due to a process of convergent evolution rather than shared ancestry. These findings have two crucial implications: First, they suggest that humans may be far from unique in their ability to intentionally coordinate attention of multiple individuals to a common focus. Second, they challenge the view that the understanding of referential gestural signaling by receivers unambiguously demonstrates that an inference has occurred.Citation3 In addition, these results strengthen the findings of Gwinner,Citation22 who suggested that ravens have a unique multifaceted communicative repertoire characterized by a high degree of behavioral plasticity. Interestingly, GwinnerCitation22 also noted that the degree of gestural flexibility might significantly differ between ravens and other corvid species such as rooks (Corvus frugilegus) and magpies (Pica pica).
However, since the gestural domain and it’s underlying cognitive complexity and plasticity has so far been widely neglected in communicative studies of birdsCitation26-Citation29 and other non-primate vertebrates, the absence of referential signaling and gestural flexibility in other species (but see refs. Citation31–Citation30) might merely reflect a paucity of data, rather than a lack of gestural abilities on behalf of the animals. Future research is needed to understand in much more detail how referential gestural signaling systems have evolved and which key traits shaped their development, variation, distribution and their underlying cognitive complexity.
Acknowledgments
I am grateful to the late Eberhard Gwinner and Bernd Heinrich for inspiring this new research avenue. For helpful discussion and support, I thank the whole team of the KLF, Tobias Deschner, Thomas Bugnyar, Wolfgang Wickler, Lucie Salwiczek and one anonymous reviewer. I thank Dorothee Claßen for the drawing of the directed scratch and Claudia Wascher for sharing her pictures with me. This project was supported by a Sofja Kovalevskaja Award of the Alexander von Humboldt Foundation.
References
- Peirce CS. The philosophy of Peirce: Selected writings. New York: Harcourt, 1956.
- Sperber D, Wilson D. Relevance: Communication and cognition. Cambridge, Massachusetts: Harvard University Press, 1986.
- Sperber D. Understanding verbal understanding. In: Khalfa J, ed. What is intelligence. Cambridge, Massachusetts: Cambridge University Press, 1994:179-08.
- Camaioni L. The development of intentional communication: A re-analysis. In: Nadel J, Camaioni L, eds. New Perspectives in Early Communicative Development. London: Routledge, 1993:82-96.
- Gomez JC. Shared attention in ontogeny and phylogeny: SAM, TOM, Grice, and the great apes. Cah Psychol Cogn 1994; 13:590 - 8
- Bates E, Benigni L, Bretherton I, Camaioni L, Volterra V. The Emergence of Symbols: Cognition and Communication in Infancy. New York: Academic Press, 1979.
- Werner H, Kaplan B. Symbol formation. New York: Wiley, 1972.
- Bates E, Camaioni L, Volterra V. The acquisition of performatives prior to speech. Merrill-Palmer Quarterly 1975; 21:205 - 26
- Bruner J. The ontogenesis of speech acts. J Child Lang 1975; 2:1 - 19; http://dx.doi.org/10.1017/S0305000900000866
- Ladygina-Kohts NN. Infant Chimpanzee and Human Child. A Classic 1935 Comparative Study of Ape Emotions and Intelligence. New York.: Oxford University Press, 1935.
- Gardner RA, Gardner BT. Teaching sign language to a chimpanzee. Science 1969; 165:664 - 72; http://dx.doi.org/10.1126/science.165.3894.664; PMID: 5793972
- Miles HL. The cognitive foundations for reference in a signing orangutan. In: Parker ST, Gibson KR, eds. Language and Intelligence in Monkeys and Apes. Cambridge: Cambridge University Press, 1990:511-39.
- Savage-Rumbaugh S, Rumbaugh DM, McDonald K. Language learning in two species of apes. Neurosci Biobehav Rev 1985; 9:653 - 65; http://dx.doi.org/10.1016/0149-7634(85)90012-0; PMID: 4080283
- Leavens DA, Hopkins WD, Bard KA. Indexical and referential pointing in chimpanzees (Pan troglodytes). J Comp Psychol 1996; 110:346 - 53; http://dx.doi.org/10.1037/0735-7036.110.4.346; PMID: 8956506
- Pika S, Mitani JC. Referential gesturing in wild chimpanzees (Pan troglodytes). Curr Biol 2006; 16:191 - 2; http://dx.doi.org/10.1016/j.cub.2006.02.037
- Pika S, Mitani JC. Referential gestures in chimpanzees in the wild: Precursors to symbolic communication? In: Smith ADM, Smith K, i Cancho RF, eds. The evolution of language: Proceedings of the 7th International Conference (Evolang7). London: World Scientific Publishing, 2008:478-9.
- Tomasello M. Origins of Human Communication. Cambridge, Massachusetts: MIT Press, 2008.
- Veà JJ, Sabater-Pi J. Spontaneous pointing behaviour in the wild pygmy chimpanzee (Pan paniscus). Folia Primatol (Basel) 1998; 69:289 - 90; http://dx.doi.org/10.1159/000021640; PMID: 9751833
- Pika S. Chimpanzee’s grooming gestures and grooming sounds: What might they tell us about how language evolved? In: Dor D, Knight C, Lewis J, eds. The Social Origins of Language: Early Society, Communication and Polymodality. Oxford: Oxford University Press, in press.
- von Blotzheim UN. Handbuch der Vögel Mitteleuropas, Band 13-III. Wiesbaden: Aula Verlag, 2001.
- Heinrich B. Mind of the raven. New York: Harper-Collins, 1991.
- Gwinner E. Untersuchungen über das Ausdrucks- und Sozialverhalten des Kolkraben (Corvus corax corax L.). Z Tierpsychol 1964; 21:657 - 748; http://dx.doi.org/10.1111/j.1439-0310.1964.tb01212.x
- Heinrich B. Food sharing in the raven, Corvus corax. In: Slobodchikoff CN, ed. The Ecology of Social Behavior. San Diego: Academic Press, 1988:285-311.
- Pika S, Bugnyar T. The use of referential gestures in ravens (Corvus corax) in the wild. Nat Commun 2011; 2:560; http://dx.doi.org/10.1038/ncomms1567; PMID: 22127056
- Pika S. What is the nature of the gestural communication of great apes? In: Zlatev J, Racine T, Sinha C, Itkonen E, eds. The Shared Mind. Amsterdam, the Netherlands: John Benjamins Publishing Company, 2008:165-86.
- Armstrong EA. The ethology of bird display and bird behavior. New York: Dover Publications, INC., 1965.
- Marler P, Evans CS, Hauser MD. Animal signals: Motivational, referential or both? In: Papousek H, Jürgens S, Papousek M, eds. Nonverbal vocal communication: Comparative and developmental approaches. Cambridge: Cambridge University Press, 1992:66-86.
- Jarvis ED. Learned birdsong and the neurobiology of human language. Ann N Y Acad Sci 2004; 1016:749 - 77; http://dx.doi.org/10.1196/annals.1298.038; PMID: 15313804
- Catchpole CK, Slater PJB. Bird song: Biological themes and variation. Cambridge: Cambridge University Press, 1995.
- East ML, Hofer H, Wickler W. The erect 'penis' is a flag of submission in a female- dominated society: Greetings in Serengeti spotted hyenas. Behav Ecol Sociobiol 1993; 33:355 - 70; http://dx.doi.org/10.1007/BF00170251
- Kaplan G. Pointing gesture in a bird- merely instrumental or a cognitively complex behavior?. Current Zoology 2011; 57:453 - 67